We are currently seeking applications for the role of Reviews Editor for the journal Development, our flagship journal for the developmental biology and stem cell community. This is a permanent, full-time position.
Joining an experienced and successful team, this is an exciting opportunity to make a significant contribution to one of the major journals in the field of developmental biology. Development publishes outstanding primary research articles, reviews and other front section content across the breadth of the developmental biology and stem cell fields.
Applicants should hold a PhD in developmental or stem cell biology. Post-doctoral and/or previous editorial experience is desirable, although we will provide full training on the role in-house. The successful candidate will have a broad interest in science, the scientific community and publishing. Excellent interpersonal and literary skills, enthusiasm and commitment are also essential requirements for the position.
Core responsibilities include:
Commissioning, handling peer review and developmental editing of material for the front section of the journal
Representing the journal at international conferences and within the wider scientific community
Writing press releases, article highlights and material for Development’s community website ‘the Node’
Creative involvement in the journal’s development
Additional responsibilities may be available for the right candidate. The Reviews Editor will work alongside an experienced in-house team, including the Executive Editor and one other Reviews Editor, as well as with our international team of academic editors. This position provides an excellent opportunity to gain experience on a highly successful life-science journal, and offers an attractive salary and benefits. The position will be based in The Company of Biologists’ attractive modern office on the outskirts of Cambridge, UK.
The Company of Biologists (biologists.com) exists to support biologists and inspire advances in biology. At the heart of what we do are our five specialist journals – Development, Journal of Cell Science, Journal of Experimental Biology, Disease Models & Mechanisms and Biology Open – two of them fully open access. All are edited by expert researchers in the field, and all articles are subjected to rigorous peer review. We take great pride in the experience of our editorial team and the quality of the work we publish. We believe that the profits from publishing the hard work of biologists should support scientific discovery and help develop future scientists. Our grants help support societies, meetings and individuals. Our workshops and meetings give the opportunity to network and collaborate.
Applicants should send a CV to recruitment@biologists.com, along with a covering letter that summarises their relevant experience, why they are enthusiastic about this opportunity, and their current salary level. Please direct any informal enquiries to Development’s Executive Editor, Katherine Brown: katherine.brown@biologists.com
Applications will be considered on an ongoing basis and should be sent as soon as possible. Application deadline: June 24th.
This year is the 70th anniversary of the BSDB – an obvious occasion to look back at the society’s history. As the BSDB’s communications officer I felt a need to become proactive and started, a good three years ago, to investigate the society’s past – only to find a rather blank sheet, with the laudable exception of a published article by Jonathan Slack (Slack, 2000). In his article, Jonathan provides an overview of the early decades of our society: starting as “(London) Embryologists’ Club” in 1948 (notebook #2, p.2f.; Fig.1), renamed in 1964 into “Society for Developmental Biology” (SDB; notebook #1, p.28f.) and, eventually, into “British Society for Developmental Biology (BSDB)” in 1969 (to avoid confusion with the American partner society SDB). Apart from Jonathan’s article, there was no organised information about BSDB chairs, let alone officers or committee members of the last 70 years, nor about its conferences or potential educational or political activities. I therefore started a BSDB history project trying a number of strategies to unearth some of its past, which eventually led to the launch of the BSDB Archive.
Fig. 1 Extracts from page 2 and 3 of the digitised notebook #2 documenting the birth hour of “The Embryologists’ Club” on February 19th 1948, with E.A. Fraser as chairman, Alan Fisk as secretary, Margaret Tribe as treasurer, and two further committee members.
A story of discovery
My first attempt to dig into the BSDB’s history was to contact former publications/communications officers of the last two to three decades. This yielded PDF files of newsletters covering the period since 2000. However, little newsletter information appeared to have been held for the period before that. Also former BSDB meetings officers could not help with sufficient relevant documents to reconstruct the conference history.
I also enquired at history archives. For example, the Wellcome Library holds conference documents of the years 1973, 1975, 1976, 1983, 1986-88. But these could only be viewed on-site and, if digitised, could not be made publicly available – hence another dead end.
Furthermore, I followed up on a hint that some old conferences were published as issue supplements in JEEM (precursor of the journal Development), but lengthy searches through Development’s online archive of the 70s and 80s revealed only one such issue (BSDB, 1984) – and this exact issue was also the only one we later found as a hard copy (1984-2). However, I learned that in the 80s and 90s it was a requirement for all invited speakers to provide a paper for publication in a JEEM or Development Supplement, something which all speakers seemed to have willingly agreed to. As Phil Ingham commented to me: “When you look at the calibre of speakers that provided these papers, you get a sense of the very high esteem in which the BSDB meetings were held – basically, all the top developmental biologists in the world wanted to be invited to our meetings (I am sure this is still true)“. Some examples of these special issues are linked out from our meeting documents 1987-3, 1988-1+2 and 1989-1+2.
Finally, I searched for former BSDB members and contacted them one by one. But I was usually informed that potentially helpful documents vanished when offices were cleared out upon retirement – a sign that it might in fact be too late for the BSDB history project.
Thomas Stoneman and a glance into the lower drawer of the archive.
Although none of the attempts yielded significant outcome reaching back into the last millennium, new hope arose when contacting Robert Kelsh (Secretary 2003-08) who pointed out that the same BSDB archive used by Jonathan Slack for his history article should still be with Michael Taylor (Secretary 2008-13) in Cardiff. A few days later, the archive was opened and it was agreed with the BSDB committee that Mike’s student, Thomas Stoneman, would be paid to sort through it and provide an overview. His findings revealed that we had struck gold! There were newsletters from 1979-2002 (#1 to #23-2), an almost complete list of meeting programs (and partly even abstracts) dating back to 1964, and hand-written or printed minutes of committee meetings starting with the foundation meeting in 1948, apart from membership indices of many different years, plenty of correspondence, meeting planning documents, financial statements and contemporary information about other societies, in particular ISDB and EDBO (Fig.1; for more details see the archive list and the archive-curiosities document).
Fig.2 Logo design for the digital BSDB archive (Box 1).
Following our initial euphoria, in came the sobering thoughts of what to do with these materials – and proper archiving and digitisation was (and remains) the obvious ultimate goal. Upon consultation with Carsten Timmermann, science historian here at Manchester, I contacted libraries and history archives, but none of these attempts provided a satisfactory way forward. Pragmatic solutions had to be found, and it was eventually decided that Thomas would continue his work for a while and start digitising key documents including the notebooks of the (London) Embryologists’ Club, all newsletters, and part of the conference documents. I organised the files as they became available, uploaded them as “BSDB Archive” (BSDB comms account) on the free and indexed online repository figshare.com, and designed a specific logo for future branding (Fig.2). To close remaining gaps, Mike Taylor kindly hosted me for a day in June 2018 and, together with Thomas, we searched through the archive using rapid photo-documentation to digitise further interesting finds. Through all these efforts, an important fraction of our documents has now been made publicly available (Box 1).
Clearly, I am not a historian, but even a lay person can sense the value of the BSDB archive. Here, I share some of my own thoughts and observations that arose when archiving the materials and browsing through them. And I will also explain some of my ideas of how to make practical use of the documents.
The BSDB newsletters
Newsletters are an essential pillar of scientific societies or communities (Kelty, 2012). They reveal a lot about a society’s nature and areas of engagement, and this perspective is now provided for the BSDB reaching back four decades. As shown in Box 1, BSDB newsletter #1 was published in 1979, and we hold an almost complete list of issues since then, with only 3 issues missing from this entire period! Initially, the newsletters were numbered individually up to issue #45 in 2002. From then on, the numbering occurred as single volume per year with summer and autumn editions sub-numbered as 1 and 2; consequently issue #46 was replaced by #23-2, as if newsletters had been numbered by year from start, thus making it possible to calculate the publication year and issue of previous newsletters in retrospect (A. Furley, pers. comm.; footnote in Newsletter #23-2, 2000, p.1). Since 2013, during my time as communications officer, publication has been reduced to only one newsletter per year. This latter change reflects the fact that information is now made available in more timely fashion on the BSDB website and on The Node, so that newsletters have changed into legacy items rather than carriers of urgent news and information (see editorial of Newsletter #37/38, 2016/17).
Another obvious trend is the dramatic improvement of editing and printing technology (from type writer to computer, from black-and-white print to colour; Fig. 3), as well as the advent of the internet which becomes obvious during the first two years of the millennium. The first BSDB website address (http://www.ana.ed.ac.uk/BSDB) is mentioned in issue #44 (2000, p.5), which then changed into today’s bsdb.org in issue #23-2 (2002, p.2), followed by several issues alerting people to the fact that the new website exists. To my surprise, I could not find any dedicated article introducing the BSDB website to the community. Shortly after, a website co-ordinator was introduced on the committee (see Appendix), first represented by Kate Storey (2003-4) followed by Andrew Jarman (2004-5), who then became publication secretary and website co-ordinator rolled into one (2005-10); this combined task was thereafter renamed into “communications officer”. After the turn of the millennium, web links were increasingly used in many contexts, paralleled by the disappearance of paper versions of important forms (e.g. to register for conferences, apply for travel grants or submit abstracts – which had often been grouped together in the so called “centre section”). Before the advent of the internet, it made a lot of sense to provide these forms in newsletters because it helped to reduce the burden of postage; as Phil Ingham explained to me of his time as publication officer (1991-95): he needed to get ~700 newsletters printed, stuffed into envelopes and sent out one-by-one.
Fig. 3 Two examples of newsletter cover images
When browsing through the newsletters, there are obvious phases where the emphasis lies on different forms of contents, likely due to the personal preferences of officers and chairs in charge. For example, there are periods where meeting reports are a regular feature, whereas they are completely absent at other times. Constant elements that feature in almost all newsletters throughout four decades include: (1) meeting announcements (see below for more detail); (2) lists of officers and committee members of the time (see Appendix); (3) obituaries and book reviews – although they gradually vanished during the last years likely due to the advent of The Node as a “modern newsletter” for the wider community of cell and developmental biologists covering much of the more general news (Vicente et al., 2017; see editorial of Newsletter #37/38, 2016/17); (4) reports (or at least mentions) of the winners of society awards, i.e. the Waddington medal since 1998, Beddington medal since 2004, Cheryll Tickle medal and Dennis Summerbell Lecture award since 2016.
To start capitalising on newsletters as a unique source of information, I have linked BSDB medal winners listed on our website to the newsletters which contain the respective reports/mentions of awardees. I feel that this does not only provide evidence for the otherwise anecdotal lists, but it also enriches them with contemporary views about the awardees’ achievements. Furthermore, I extracted a list of officers on the committee (see Appendix), as well as meeting information (see below for more detail); the latter includes existing meeting reports which are now part of the documents listed under “Meeting programs” (Box 1) and provide insightful contemporary views about those events. Obituaries are further valuable documents of the time which should be made accessible; for example I inserted Ed Lewis’ obituary (Newsletter#25/2, 2004, p.8) in a link collection of Drosophila research history articles, or Rosa Beddington’s obituary (Newsletter #43, 2001, p.13) on the BSDB’s Beddington medal page.
But there are many examples of interesting further discoveries that I came across. First of all, the newsletter does not restrict to BSDB-specific information, but it provides a wider insight by reporting community-relevant news of the time, such as new editors of subject-related journals, prizes in the field, conferences of other societies, etc. (i.e. the kind of information that is well covered these days by The Node; Vicente et al., 2017). Some contents touch on topics of political or societal relevance still debated these days. For example, Newletter #23-2 (2002, p.6) mentioned a plenary lecture at the BSCB/BSDB Spring meeting in Warwick in 2003 entitled “Government support for world class science” given by Lord Sainsbury of Turnville (then Government Minister for Science and Innovation); Kate Storey reported on a meeting entitled ‘“Women on Top”, Reflections on Women in Science’ (Newletter #23-2, 2002, p.10); or some pieces reflected on the newly emerging PLoS journals and the wider question of open access (Newsletters #24-1, 2003, p.3 and #27-2, 2006, p.8).
Also relevant in this context are contributions about science communication and advocacy. For example, the article entitled ‘The headless tadpole affair” (Newsletter #36, 1997, p.1) described a misleading press release about the rather forward-looking topic of mammalian organ cultures, and touches on the dangers of interacting with the media (“never get entangled with the media without some training“). Newsletter #41 (2000, p.3) contains a report about the UK Life Sciences Committee (UKLSC), a joint forum for UK bioscience societies aiming to speak “with one voice to the media, the government and the research councils, for the things we need and want”. Paul Martin acted as the BSDB liaison followed by Guy Tear. One of its legacies was the speakers database (www.biology4all.com) which still continues to lead a cryptic life. More importantly, the UKLSC merged into the Bioscience Federation in 2002 (Newsletter #23-2. 2002, p.3f.), which then combined with the Institute of Biology into the (Royal) Society of Biology in 2009 – and the BSDB became a member society in 2016 (Newsletter #36-2, 2015, p.23). In spite of all these efforts to promote biology in society, the above mentioned report in Newsletter #41 about the UKLSC reads rather sobering from today’s perspective: “There is an ongoing programme to aid quality science education in schools and the UKLSC sponsors school level videos on topics like genetic engineering so that in 10 years time we’ll all have PhD students who know significantly more than we do before they even start their bench work!” That the topic of education was taken with great enthusiasm at the time, is also reflected in the creation of education officer posts on the BSDB committee which were held by David Wilkinson (2002-2006) and Corinne Houart (2003-2007) – but these posts were not continued thereafter.
Another highlight of science communication is Ann Lackie’s article “Taking the anoraks out of fiction” (Newsletter #26-1, 2005, p.5), about the need to establish dialogue and collaboration between fiction writers and scientists, later formalised in the SciTalk initiative and website which is still live. Also the various experiments with sci-art fall into this category. Newsletter #42 (2000) shows a glass window termed “Window on Life” with Developmental Biology motifs. It was a collaboration between the MRC Centre for Developmental Neurobiology at Guy’s, Jim Cohen (Kings’ and St. Thomas’ Hospital) and the glass artist Carole Nunes. The window was installed in the north wing link corridor at St Thomas’ (any news of its present existence?). Other examples are the inclusions of sci-art exhibitions at the York Spring Meeting in 2002 (Newsletter #45, 2002, p.2 & back cover; announced in the previous newsletter) and at the Autumn Meeting 2016 in Edinburgh, where chimaera- and embryo-inspired artwork was on display (Newsletter #37/38, 2016/17, p.14). The sci-art exhibition in York prompted Phil Ingham to comment on science in society: “If the public are prepared to put their hands in their pockets to support the arts or sport, why then should they not be persuaded to support science in a similar way?” (Newsletter #23-2, 2002, p.10). Unfortunately, this topic remains as important today as it was then, and this is clearly demonstrated by the current advocacy campaign which was initiated by the BSDB together with the Company of Biologists (Maartens et al., 2018; Prokop, 2018; several articles in Newsletter #37/38, 2016/17).
An internal communications issue observed over the decades is the attempt to integrate graduate students into society life (Fig.4). For example, a graduate student meeting at the Edinburgh conference in 1981 is mentioned in Newsletter #5 (1981, p.7), and there was an announcement of a PhD student conference in Autumn 1999 – although I could not spot any traces thereafter confirming that this meeting had taken place or reflecting on its success. Furthermore, there were attempts to animate PhD students to write for the newsletter (Newsletter #23-2, 2002, p.5), which then happened for a short while starting in 2003 (Newsletter #24-1, 2003, p.6). This was further promoted also by an announcement to pay students a £50 reward for writing meeting reports (Newsletter #23-2, 2002, p.10; Fig.4). An important step was taken with the inclusion of student representatives on the committee starting in 1987, joined by a postdoc representative in 2015, which eventually led to the introduction of a separate webpage for young members (Newsletter #37/38, 2016/17, p.35). The importance of uncovering this particular part of the society’s history became clear to me during an engaging conversation I had with our newly elected postgraduate representative, Jessica Forsyth (2018-21); learning the mere fact that she has become part of a thirty year history sparked enormous enthusiasm and the immediate idea to get in contact with former graduate representatives.
Fig. 4 Attempts to animate students to actively contribute to BSDB newsletters and activities.
A further interesting read are the articles announcing and explaining the introduction of new society awards and medals, such as the Waddington medal (Newsletter #36, 1997, p.9; Fig.5 – to recognise contributions both to the field and to the Developmental Biology community in the UK), the Beddington medal to honour Rosa Beddington who had tragically died of cancer aged 45 in May 2001 (Newletter #23-2, 2002, p.2 – to recognise the achievements of PhD students), the Gurdon Summer Studentships (Newsletter #35, 2014, p.10 – to allow undergraduates to work in a lab over summer), the Cheryll Tickle medal (Newsletter #36, 2015, p.14 – to recognise achievements of female researches at their mid-stage career), and the Dennis Summerbell Lecture award (Newsletter #37/38, 2016/17, p.47 – to honour good work by postdoctoral rsearchers).
Fig. 5 First presentation of the Waddington medal design on the cover page of Newsletter #36 from 1997.
New to me was the origin of the BSDB logo, announced as a competition in 2001 (Newsletter #43, 2001, p.7). Out of 18 submissions, the design by Jeff Christiansen was the clear winner (Newsletter #44, 2001, p.25; Fig.6; logos). As Christiane Ruhrberg’s explained at her Cheryll Tickle medal lecture at the 2018 Spring meeting, she was the first person ever to have seen the logo, since Jeff stayed at her house when he designed it. – and it is little anecdotes like this that bring history to life.
Fig. 6 Jeff Christiansen’s designs that won the BSDB logo competition (Newsletter #44, 2001, p.25)
Each time I look at an issue, I stumble across other interesting features, and there will be many other topics and events worth reporting, such as the gradual development of scientific questions, themes and methodologies, the BSDB’s financial history, the development of its close and unique relationship with the Company of Biology, or little anecdotes such as the food poisoning at the York Spring Meeting in 2001 (Newsletter #45, 2002, p.2) which are also part of our society’s history. This said, the newsletters are now publicly available, and new discoveries can be made by anyone taking an interest!
The BSDB Meeting programs
The current archive contains digitised meeting programs covering most of the meetings, from the inaugural conference of the newly formed SDB in 1964 (meeting #1) through to the present. Where no meeting schedules were encountered (although they may still be hidden somewhere in the materials!), I could retrieve a lot of detailed information from the newsletters covering the period after 1996 (although relevant conference information became less detailed in newsletters after 2002, likely due to the availability of programs online that are now lost). For conferences since 2007, I could often retrieve further information from organisers who still kept relevant files – and it might be a worthwhile effort to write to former conference organisers more systematically to unearth the occasional stock of documents collecting dust on shelves or in filing cabinets.
Fig. 7 Newsletter covers from 1993, 1995 and 1996 showing illustrations for meetings of the time which might reflect the actual meeting posters.
Meetings often start featuring in newsletters about 2-3 years leading up to the conference, and interesting planning developments can be observed during this period. For example, initial announcements of the Spring Meeting 2000 were entitled “Cell death and proliferation” which then refined into “Pattern formation and control of cell number”; or the Spring Meeting of 2002 started off as “Evolution & development” which then turned into “Evolution of developmental mechanisms”. Reconstructing the meeting history from newsletters also led to the addition of insightful meeting reports to the “Meeting programs” section of the archive, and also a number of poster designs resurfaced in this way (Fig.7).
Fig. 8 Phil Ingham and Jonathan Cooke, organisers of the 1988 BSDB Spring Symposium, pose together at the 70th anniversary Spring meeting in 2018.
In general, the recovered documents will be an exciting resource for those who attended or organised those meetings, and part of that excitement came across at the 70th anniversary Spring meeting in 2018 (Fig. 8). But the conference documents also provide an important insight into the scientific topics that dominated the field at any given time. They offer an opportunity for young developmental biologists to understand the historical roots of their specific sub-fields – and perhaps the surprising revelation that many questions they address today were already asked long before they got into science. The documents also make transparent which other societies the BSDB collaborated with over the years.
Fig. 9 Nick Hopwood showing an image of the early committee meeting notes during his talk about the history of Developmental Biology at the BSDB Spring meeting 2018
The early meeting notes
The hand-written meeting notes are the archive documents which are least accessible to a lay person, but might likely be the most interesting ones for historians (Fig.9). The two note books of the “The (London) Embryologists’ Club” digitised so far, cover meeting minutes of the time from its foundation in 1948 to its transition into the SDB in 1964 (Box 1). Book #1 covers mainly committee meetings, and book #2 predominantly scientific symposia and their associated general society meetings. The precise dates for the various minutes tend to be provided at the start of each entry, and meetings are usually signed off with date on the next meeting, providing a complementary means to deduce or confirm dates. Using these materials, Jonathan Slack extracted a brief overview of this period (Slack, 2000), but more work will be required. For example, they will enable us to partly reconstruct the early conference history before 1964 (i.e. the period not covered by current meeting documents; Box 1; archive list), and I already extracted a list of early presidents/chairs, officers (Appendix). Another interesting addition is the little rule book of the newly founded SDB (SDB-1964), to which we added some documents illustrating the transition process and the thoughts leading up to it.
Conclusions, your contributions, final home for the archive
Here I have explained the story behind the digital BSDB Archive and provided my personal view of its contents, relevance and potential applications. The BSDB will likely not go further with the digitisation. But I hope that the “open source” nature of the BSDB Archive will attract wider interest and inspire others to join in and help develop its full potential – be it biologists browsing around, or (hobby) historians making systematic scientific use of it (Fig.10). Importantly, also the hard copies of our archive have now found a home: they will form part of the Historical collections of the John Innes Centre (collections.jic.ac.uk) where they will be kept together with the archive that covers 100 years of history of the Genetics Society, thus allowing comparative studies across two societies.
I hope that the BSDB archive will be of value to our community and those studying the history of science. Carsten Timmermann wrote from his perspective as science historian: “Your archive is a little treasure trove and will enable us to understand the history of Developmental Biology in this country much better. I wish other societies would follow your example. If we had a whole set of similar archives at our disposal, this would help us to study the way the life sciences overall have developed, comparing and contrasting sub-disciplines and understanding trends. For example, one could look at conference programmes in different fields within the life sciences and study how molecular methods have transformed biology.”
Fig. 10 Tweet by Cheryl Lancaster historian from Durham working on the history of stem cell science
I would like to finish this blog post by asking your help: if you read or study documents of the archive and gain any new insights, recall anecdotes, have additional background knowledge that complements available information, or realise that you hold any additional documents that might help to fill remaining gaps, please be so kind to contact comms@bsdb.org and let us know – as a contribution to the wider understanding of the history of our society and field in general.
Andreas Prokop
————————————————————-
References
British Society of Developmental Biology (BSDB) (1984). European Developmental Biology Congress (abstracts). J Embryol Exp Morphol Suppl. 1, 1-271 – [LINK]
Kelty, C. M. (2012). This is not an article: Model organism newsletters and the question of `open science’. BioSocieties 7, 140-68 – [LINK]
Maartens, A., Prokop, A., Brown, K., Pourquié, O. (2018). Advocating developmental biology. Development 145 — [LINK]
Prokop, A. (2018). What is Developmental Biology – and why is it important? Open Access Govern 17, 121-123 – [LINK]
Slack, J. M. (2000). A short history of the British Society for Developmental Biology. Int J Dev Biol 44, 79-83 — [LINK]
Vicente, C., Maartens, A., Brown, K. (2017). The Node and beyond – using social media in cell and developmental biology. Sem Cell Dev Biol — [LINK]
————————————————————-
Appendix:
BSDB chairs & officers since 1948
This list was extracted from the digitised newsletters, note books and committee/general meeting minutes (Box 1; early officers). The precise years of transition between consecutive officers were not always easy to establish and might need further refinement.
Fig.11 Description of the roles of BSDB officers (Newsletter #1, 1979, p.2)
Chairs and presidents
Ottoline Leyser (chair, 2014-2019)
Elizabeth Robertson (chair, 2009-2014)
Matthew Freeman (chair, 2004-2009)
Phil Ingham (chair, 1999-2004)
Jim Smith (chair, 1994-1999)
Michael Akam (chair, 1989-1994)
Martin Johnson (chair, 1984-1989)
Chris Graham (chair, 1979-1984) – president: David R. Newth
Anne McLaren (1975-1979)
Michael Abercrombie (1970-75 – thereafter Honorary President)
David R. Newth (chair 1963-69)
William J. Hamilton (chair 1951-63)
John Dixon Boyd (chair, 1950)
E.A. Fraser (chair; 1948-50)
James P. Hill (first president; 1948-54)
Secretaries:
Megan Davey (2018-23)
Kim Dale (2013-18)
Mike Taylor (2008-13)
Robert Kelsh (2003-08)
Ivor Mason (1998-2003)
Jonathan Slack (1993-98)
Peter Thorogood (1988-93)
Chris Ford (1984-88)
Michael Balls (ca. 1978-1983; last 4 years Secretary/Treasurer)
Postdoctoral position in the research group of Dr. Nikolay Ninov at the Center for Regenerative Therapies Dresden and the Paul Langerhans Institute Dresden (PLID) (of Helmholtz Zentrum München and the German Center for Diabetes Research (DZD e.V.).
Our goal is to understand beta-cell regeneration and function in vivo in order to develop innovative cures for diabetes. We use the zebrafish as a model organism. The zebrafish is ideal to observe the behavior of beta-cells in their endogenous environment using live imaging. To do so, we have developed new tools to visualize beta-cell function and proliferation while performing genetic and lineage-tracing analysis (see Janjuha et al., 2018; Singh et al., 2018, Alfar et al., 2017; Spanjaard et al. 2018).
Currently, we are focusing on the following projects:
At the last AGM, held at the 2018 Spring Meeting in Warwick, five new BSDB committee members were elected to take term in autumn. They will replace the five leaving members: our Graduate Representative Alexandra Ashcroft, Postdoc Representative Michelle Ware, Secretary Kim Dale, Meetings Officer Josh Brickman, and Communications Officer Andreas Prokop (see a complete list of current committee members here). Please, read below about the new committee members, their careers, research interests and plans for their time on the committee.
Jessica Forsyth – the new Graduate Representative
I’m extremely happy to be acting as the new Graduate Representative for BSDB, following Alexandra Ashcroft who has worked to represent graduate students at meetings and enhance the student experience. I hope to further this work, and make sure the BSDB meetings continue to meet the needs of students at various stages within their academic careers (see Newsletter #37/38, 2016/17, p.30ff.).
As a Physics with Medical Physics graduate, I’m relatively new to the field of Developmental Biology. I made this switch when I applied for the Quantitative and Biophysical Biology programme at The University of Manchester. Now in my first year of my PhD, I’m completing two rotation projects within the department. In my first rotation project, I worked on the pre-implantation mouse embryo with Berenika Plusa, and started to develop a mathematical tool to match single cells across imaging modalities, together with Simon Cotter from the Mathematics department. Now I am currently working with Martin Baron, and attempting to develop a mathematical model which encompasses the role of Notch in Drosophila wing vein formation, and to inform this model with live imaging studies.
Changing fields for my PhD seemed daunting when applying, but having been a part of two labs, I realise that there is a huge role for Mathematics and Physics to play in Developmental Biology. This was confirmed in my recent attendance to the BSDB Spring Meeting, where numerous talks described their collaborations with more theoretical labs. I hope to encourage the attendance of more theoretically based labs to BSDB meetings to encourage collaborations across disciplines.
If you have any questions or suggestions please feel free to contact me by email. I look forward to hearing from you and meeting you at the next BSDB meetings.
Charlotte Sophie Louise Bailey – the Postdoc Representative
Having completed my PhD in the field of vertebrate somitogenesis in the lab of Kim Dale at the University of Dundee, I am now a Marie Curie postdoctoral fellow in the lab of Elke Ober at the Novo Nordisk Center for Stem Cell Biology (DanStem) in Copenhagen. I am interested in determining the cell behavioural dynamics underpinning liver regeneration in zebrafish.
I am honoured to have been elected to serve on the BSDB committee as postdoc representative. In this role, I aim to draw on my experiences in event management and public outreach to build on the fantastic work of my predecessor Michelle Ware to support postdoctoral scientists within the BSDB community (see the PhD/postdoc website and Facebook group).
For the budding young developmental biologist, the highlight of the scientific year has to be the BSDB Spring meeting – which I encourage every postdoc to attend! With an unfailingly engaging scientific and societal programme (Newsletter #37/38, 2016/17, p.30ff.), this annual meeting consistently stands apart as the forum to network within the Developmental Biology community and beyond, as well as offering exposure to a broad range of exciting, cutting edge science and ideas. As part of my role as BSDB postdoc representative, I aim to tackle the increasing demand by postdocs for interdisciplinary training and discussion by introducing workshops at the annual Spring Meeting with a focus on introducing and developing cross-disciplinary skill sets and network connections, such as Python/Matlab programming, big data mining and biophysics. These workshops could also be used as a bridge for discussion of career choices both inside and outside of academia and the development of transferable skills.
Undoubtedly, one of the strongest attributes of the BSDB is its great sense of community and inclusion. Following Brexit, sustaining a strong feeling of unity within the scientific community will be more important than ever to preserve the UK’s reputation as a welcoming and international environment for research excellence (see also ‘Chair’s welcome note’ in Newsletter #37/38, 2016/17, p.4f.). In conjunction with the The Company of Biologists, the BSDB offers amazing support to its early-career members both financially through travel grants to attend scientific meetings in the UK and abroad, and personally at the many meetings and workshops organised annually and through multimedia such as ‘the Node’, Facebook and Twitter (Vicente et al., 2017). I implore all postdocs to take advantage of these fantastic opportunities to engage with the BSDB and other subject-specific international societies to help us preserve and nurture our supportive global scientific community.
Take part and develop your potential as a developmental biologist! Become a member of the BSDB to receive all of these great benefits. Don’t forget to follow ‘The Node’ on their website, Twitter or Facebook and check the BSDB website regularly for many interesting posts and discussions.
Got an idea for a great workshop or event? Don’t hold back – get in touch with me by email.
Tanya T. Whitfield
Tanya is Professor of Developmental Biology at the University of Sheffield, where she is a member of the Bateson Centre and Department of Biomedical Science [LINK].
Tanya studied early Xenopus development for her PhD at the University of Cambridge, under the supervision of Chris Wylie. In 1994, she was an EMBO short-term fellow in the lab of Christiane Nüsslein-Volhard in Tübingen, Germany, where she contributed to analysis of mutations affecting ear development isolated in a large-scale zebrafish mutagenesis screen for embryonic phenotypes. She continued to work on these mutants as a postdoc in the lab of Julian Lewis, first at the Imperial Cancer Research Fund Developmental Biology Unit in Oxford, and later in London.
Tanya established her lab in Sheffield in 1997 to continue work on the developing vertebrate inner ear, using the zebrafish as a model system. The ear is a fascinating system for study, due to its complex three-dimensional arrangement of interlinked ducts and chambers, and multitude of different cell types, including neurons, sensory hair cells, supporting and secretory cells. An enduring interest in the lab has been the analysis of signalling events that pattern the anteroposterior axis of the otic placode, precursor of the inner ear. More recently, a major focus has been on the dynamic epithelial rearrangements that generate the three semicircular canal ducts in the ear, and the use of light-sheet microscopy to image these events in real time in the live embryo. Additional recent highlights from the lab include the identification of glycoproteins required for otolith tethering in the ear, and use of the zebrafish as a screening tool for drug discovery.
Tanya is a committed teacher of Developmental Biology, running courses at both undergraduate and postgraduate levels at the University of Sheffield. Her lab also makes regular contributions to outreach events, introducing the public to the beauty and logic of embryonic development.
Shankar Srinivas
Shankar is Professor of Developmental Biology and a Wellcome Senior Investigator in the Department of Physiology Anatomy and Genetics at the University of Oxford [LINK].
He completed his BSc in Nizam College in Hyderabad, India. He then joined the group of Frank Costantini in Columbia University, New York, where he received a PhD for work on the molecular genetics of kidney development. Following this, he moved to the NIMR in Mill Hill, London, where he worked as a HFSPO fellow in the groups of Rosa Beddington and Jim Smith on how the anterior-posterior axis is established. Here, he developed time-lapse microscopy approaches to study early post-implantation mouse embryos, characterising the active migration of cells of the Anterior Visceral Endoderm that is essential for the correct orientation of the anterior posterior axis of the embryo.
In 2004 Shankar started his independent group at the University of Oxford as a Wellcome Trust Career Development Fellow. His group has shown that the coordinated movement of AVE cells requires Planar Cell Polarity signalling and that a stereotypic multicellular-rosette arrangement of cells in the visceral endoderm is essential for normal AVE migration. Currently, the research in Shankar’s group focuses on two main areas. The first is to understand how the coordinated cell movements that shape the mammalian embryo prior to and during gastrulation are controlled. The second, more recent area is to understand how the heart starts to beat. Shankar’s group has shown that, during cardiogenesis, the cellular machinery for calcium oscillation matures before the sarcomeric machinery for contraction. Shankar’s group takes a multidisciplinary and collaborative approach to address these questions, using techniques such as light-sheet and confocal time-lapse imaging, single cell approaches and embryo explant culture.
Shankar is also passionate about science outreach. His group participates regularly in science festivals, for which they have developed 3D printed models of developing embryos and a virtual reality based embryo and microscopy image volume explorer. For more information see Shankar’s public engagement page.
Jens Januschke
Jens is a Sir Henry Dale fellow at the School of Life Sciences at the University of Dundee running his lab in the division of Cell and Developmental Biology [LINK].
He did his undergraduate studies at the University of Cologne and moved for his PhD to the University Paris 7 where he got his degree in Genetics in the lab of Antoine Guichet, working on mRNA localization and microtubule-based transport in Drosophila oocytes trying to understand how the anterior posterior axis is specified in this system.
After his PhD he moved to the Institute for Biomedical Research (IRB) in Barcelona to start working with neural stem cells, called neuroblasts in the developing fly brain in the group of Cayetano González. During this time, he worked on asymmetric centrosome segregation and discovered that mother and daughter centrioles are differently distributed during asymmetric neuroblast division and shed light on the molecular mechanisms controlling this process. This work identified the first daughter centriole specific protein in Drosophila, called Centrobin.
In 2013, Jens started his own group in the cell and developmental biology division of the school of life sciences at the University of Dundee, for which he obtained a Sir Henry Dale Fellowship funded by Wellcome and the Royal Society. Currently, his group focusses on the cell biological mechanisms that control neuroblast asymmetric cell division, which includes studying the establishment of cell polarity, fate determinant localisation and spindle orientation. Jens has been involved in organizing the Scottish Developmental Biology group meeting twice in Dundee and is currently a co-organiser of the UK Workshop on Developmental Cell Biology of Drosophila.
To mark Mental Health Awareness Week, we’re sharing this blog post on the MRC WIMM blog from Gregorio Dias, who describes his experiences with stress and anxiety as a PhD student. If you’ve faced similar issues and want to share them with the community, just get in touch.
Working towards a PhD is an exciting, albeit challenging, narrative in a student’s life. The goals and aspirations that motivate one at the early stages of a PhD project are very likely to change over time.
In my view, this transformative process is much needed, as it builds up personal and scientific maturity. This journey is, nonetheless, accompanied by stress and low points and I discovered that taking time to do other things I enjoy and sharing my experiences with other students was as important as working hard to be successful. That is an accurate summary of my experience as a PhD student and I hope that sharing it here will help others in their own journey.
Aspiring to become an academic professor, I came straight from the Brazilian Amazon to start a DPhil (PhD) at the University of Oxford in the UK. I started an ambitious project aiming at answering a long-standing question in my field of study. In brief, I aimed at understanding how a particular protein of the innate immune system detects the presence of virus RNA in infected cells. Motivated by this exciting question, I immersed myself in the lab to achieve these goals. I had access to all the facilities, reagents and expertise I needed to tackle this question and never felt any pressure from my supervisors or colleagues.
However, the combination of being ambitious and anxious did not mean immediate results, and I soon learnt that top notch science is rather a slow process….
Read the full post over at the MRC Weatherall Institute of Molecular Medicine Blog
And in case you missed it, we’ll also direct you to Dave Reay’s piece in last week’s Nature on the his experience of debilitating depression during his PhD
A postdoctoral position is available in the Bush lab bush.ucsf.edu at the University of California, San Francisco to study the cellular basis of morphogenesis using live imaging, mouse genetic, and iPSC and ESC approaches. Our dynamic team focuses on understanding basic mechanisms of signaling control of morphogenesis particularly as related to human structural birth defects. The position is in the collaborative UCSF Department of Cell and Tissue Biology and Program in Craniofacial Biology, located at the UCSF Parnassus Heights campus in the center of San Francisco. UCSF offers an outstanding developmental and stem cell biology community, access to cutting edge technologies, and a supportive working environment. Candidates with a Ph.D. degree in a biological science and research experience in molecular biology, genetics, biochemistry, or live cell or live embryo imaging should submit a C.V. and names of at least 2 references via email to: jeffrey.bush@ucsf.edu
In September, Development is hosting the third of its highly successful series of meetings focusing on human developmental biology. Held in the Wotton House estate near Dorking in Surrey and organised by Paola Arlotta, Ali Brivanlou, Olivier Pourquiéand Jason Spence, the meeting will cover the latest developments and future prospects for this rapidly evolving field (you can find more information at the meeting homepage).
We are excited to announce a competition for a reporter to cover the meeting for the Node. The reporter will provide regular updates of the meeting via Twitter, and write a meeting report of their experience and the sights and sounds of the meeting to be published on the Node.
It’s a fantastic opportunity both to practice your science communication skills and for networking, and of course you’ll also be able to learn about the latest fascinating research.
The prize
The winner will get free registration to the meeting!
How to enter
To find the perfect reporter, we’d like to know why you’re excited about the future of human developmental biology. Send us 300 words answering one of the following questions:
Why is human developmental biology important?
What has been the biggest advance or paper in the field since the last Development meeting in September 2016?
What is the key burning question about human development you want to answer in your research?
The competition is open to PhD students and postdocs.
Please send entries to thenode@biologists.com with “Meeting reporter competition” as the subject.
Deadline = 15th May (winner announced soon thereafter)
Meeting registration details
If you have registered for the meeting already, you will get a refund on your costs.
If you have not registered but plan to, note that the meeting has limited places and we expect these places to run out. Soplease do not delay your application for the meeting while you’re writing your piece. The deadline for applying to the meeting is 22nd June – in your application, include reference to this competition in the box “Please state why this meeting is of particular relevance to your research and your reasons for wishing to attend”; you will not have to pay until the winner of the competition is announced.
The Centre for BioNano Interactions (CBNI), University College Dublin is seeking a highly motivated Post-Doctoral Research Fellow with research background in relevant areas of molecular and cellular biology to join a dynamic team dedicated to advancing the understanding and implementation of targeted medicines at the nanoscale.
The successful candidate will explore novel approaches to re-programme the epigenome in mammalian cells, including gene expression and epigenetic changes, and associated phenotypic outcomes. As such, demonstrated experience in mammalian cell culture, advanced molecular biology and biochemistry techniques such as ChIP-sequencing, CRISPR, flow cytometry and live cell microscopy is highly desirable.
In addition to conducting research, the successful candidate will help supervise and support PhD students working on the same topic, promote publication in high quality peer-reviewed academic publications, assist in the development of funding proposals and the management and reporting of projects and will generally contribute to the professional and smooth running of a highly interdisciplinary team.
About CBNI
Located in Dublin, one of the most vibrant cities in Europe, CBNI is pioneering new techniques and approaches at the research interface between nanoscale science and living systems, with applications in the biomedical arena. The team is highly interdisciplinary and dynamic and would suit young scientists that aspire to be at the leading edge of an emerging field of science and biomedical research.
Key areas of research at CBNI include:
Fundamental understanding of interactions between nanoparticles and living systems at cellular level and in vivo
Exosome detection and manipulation
Novel approaches to vaccine development
Applications of bio-nanoscale science for therapy and diagnostics
Mandatory Qualifications
PhD in molecular biology, cell biology or a related discipline
A strong background and in molecular and cell biology techniques
Experience in mammalian gene regulations, quantitative proteomics, epigenetics
Evidence of research activity (publications, conference presentations, awards) and future scholarly output (working papers, research proposals) and ability to outline a research project.
A demonstrated commitment to research and publications
An understanding of the operational requirements for a successful research project
Excellent communication skills (Oral, Written, Presentation etc), the ability to work effectively in a team and be self-motivated
Excellent organisational and administrative skills including a proven ability to work to deadlines
Salary: €36,854 – €37,383 per annum
Appointment on the above range will be dependent on qualifications and experience.
This is a full-time position for (initially) 1 year.
For further detail and to apply, please refer to the University College Dublin vacancies page available at this link with job Ref number 010298
The Centre for BioNano Interactions, University College Dublin is seeking a highly motivated Post-Doctoral Research Fellow with research background in relevant areas of molecular and cellular biology to join a dynamic team dedicated to advancing the understanding and implementation of diagnostics and therapeutics at the nanoscale.
The successful candidate will work on detection and manipulation of exosomes as complex nanostructures for use in biomedical applications. As such, demonstrated experience in production and harvesting of exosomes or viruses using bench-scale bioreactors is highly desirable. Analytical experience of extracellular media, and study of cell culture medium components using chromatographic or other techniques would be advantageous.
In addition to conducting research, the successful candidate will help supervise and support PhD students working on the same topic, promote publication in high quality peer-reviewed academic publications, assist in the development of funding proposals and the management and reporting of projects and will generally contribute to the professional and smooth running of a highly interdisciplinary team.
About CBNI
Located in Dublin, one of the most vibrant cities in Europe, CBNI is pioneering new techniques and approaches at the research interface between nanoscale science and living systems, with applications in the biomedical arena. The team is highly interdisciplinary and dynamic and would suit young scientists that aspire to be at the leading edge of an emerging field of science and biomedical research.
Key areas of research at CBNI include:
Fundamental understanding of interactions between nanoparticles and living systems at cellular level and in vivo
Exosome detection and manipulation
Novel approaches to vaccine development
Applications of bio-nanoscale science for therapy and diagnostics
Mandatory Qualifications
PhD in relevant disciplines of molecular and cellular biology
Experience in intercellular communication and cell signalling
Evidence of research activity (publications, conference presentations, awards) and future scholarly output (working papers, research proposals) and ability to outline a research project.
A demonstrated commitment to research and publications
An understanding of the operational requirements for a successful research project
Excellent communication skills (Oral, Written, Presentation etc), the ability to work effectively in a team and be self-motivated
Excellent organisational and administrative skills including a proven ability to work to deadlines
Salary: €36,854-€37,383 per annum
Appointment on the above range will be dependent on qualifications and experience.
This is a full-time temporary position for (initially) 1 year.
For further detail and to apply, please refer to the University College Dublin vacancies page available at this link with job ref number 010299
An important question in developmental biology is how regions with distinct identity are established despite the intermingling of cells that occurs during growth and morphogenesis. Our recent work revisited some old studies of how the vertebrate hindbrain is patterned, and found that sharp and homogeneous segments are formed through a combination of identity switching and border control.
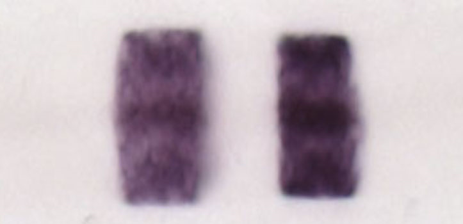
The story started in the late 1980s, in the early days of analysing developmental gene expression using in situ hybridisation. One of the genes we analysed was egr2 (a.k.a. krox20), a transcription factor which had been identified by Patrick Charnay as an early growth response gene in fibroblasts. To our amazement, we found that egr2 is expressed in stripes in the hindbrain, corresponding to two rhombomeres, r3 and r5. We then collaborated with Robb Krumlauf to show that hox genes have segmental expression in the hindbrain. egr2 and hox genes were later found to be components of a network that regulates segmental identity. A striking feature of their segmental expression is that they come to have razor sharp borders, and a clue to how these form came from the work of Scott Fraser and colleagues. They found that once morphological borders are seen in the chick hindbrain, cells do not intermingle between segments.
Sharp and homogeneous segmental expression of egr2 in the hindbrain.
In another collaboration with the Charnay lab, we carried out a screen to identify kinases that are segmentally expressed in the hindbrain. One of these, a receptor tyrosine kinase subsequently named EphA4, is expressed in r3 and r5, and we later found that it is a direct transcriptional target of egr2. We went on to show that Eph receptors and their ephrin ligands underlie cell segregation that sharpens the segment borders. This turned out to be the first example of a general role of Ephs and ephrins in border formation during development.
These findings fit the familiar idea that cell segregation sharpens and stabilises tissue organisation. However, the lineage studies in chick had found that cells marked at early stages can contribute progeny to adjacent hindbrain segments. Furthermore, experiments by Trainor and Krumlauf in mouse, and by Schilling, Prince and Ingham in zebrafish, had shown that cells transplanted between segments change identity to match their new neighbours. Intriguingly, identity switching occurs for single cells but not when groups of cells are transplanted. These findings in the early 2000s suggested that some intermingling occurs, and identity switching ensures that segments nevertheless establish a homogeneous identity. However, this idea languished as intermingling between segments had not been directly visualised, and the mechanism of switching remained a mystery. This was the problem that Megan Addison took on as her PhD project in my lab.
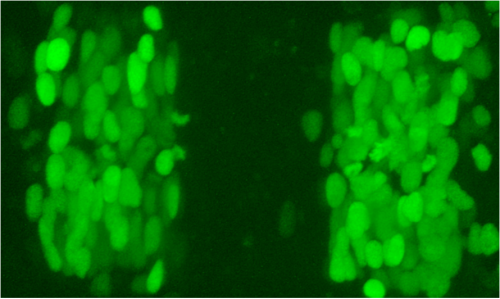
We reasoned that intermingling and identity switching of cells would mainly occur at early stages, when egr2 is first expressed but EphA4 has yet to be upregulated to sufficient levels to drive cell segregation. The key question is whether any egr2-expressing cells intermingle from r3 and r5 into adjacent segments and then downregulate egr2 expression. To address this question, we used the newly-emerging techniques for genome manipulation in zebrafish to create an enhancer trap in which a stable reporter is expressed directly from the egr2 locus. During this work, another lab reported that intermingling does not occur between hindbrain segments in zebrafish, but used reporters expressed one step downstream of egr2, which might miss the time window in which mixing occurs. Indeed, using the early reporter line that we created we found that cell intermingling and identity switching does occur.
Stable reporter for egr2 generated by CRISPR/Cas9 mediated insertion of H2B-Citrine into the egr2 locus. Some cells that have expressed egr2 are found in even-numbered segments. These cells switch identity to match their new neighbours.
We started wondering what the mechanism of switching might be, and here some other old findings came into play. The Charnay lab had reported in 2001 that mosaic ectopic expression of egr2 in the chick hindbrain causes adjacent cells to upregulate egr2. This suggested that egr2 regulates a community effect, which in classical models would involve upregulation of a signal that non-autonomously induces egr2. Such community signaling leads to homogeneous gene expression within a field of cells, and might explain why groups of transplanted cells do not switch identity. However, the puzzle of how egr2 induces egr2 in adjacent cells had also languished in the literature.
Megan analysed whether ectopically expressed egr2 acts non-autonomously in the zebrafish hindbrain. We found that it does when the egr2-expressing cells have a scattered distribution, but not when they later segregate from cells with even-numbered identity. Since our previous work had shown that the segregation is driven by EphA4, we blocked it by simultaneous knockdown of EphA4 and found that non-autonomous induction was restored. Non-autonomous induction thus depends upon how many neighbours of the same or different type you have: egr2 is only induced in cells that are surrounded by egr2-expressing cells.
What might the mechanism of the community regulation of egr2 be, and does it account for identity switching? We started thinking about retinoic acid (RA) signaling as a candidate. An RA gradient establishes segmental identity in the hindbrain, and studies of Tom Schilling in zebrafish had shown that graded expression of an RA-degrading enzyme, cyp26a1, has a key role. The lab of Cecilia Moens found that two other family members, cyp26b1 and cyp26c1, have dynamic segmental expression that also contributes to A-P patterning. We wondered if this segmental expression is under the control of segment identity genes and thus acts in a feedback loop. We found that this is indeed the case: egr2 underlies the lower expression level of cyp26b1 and cyp26c1 in r3 and r5 compared with r2, r4 and r6. Since a high level of cyp26 enzymes can non-autonomously decrease RA levels in neighbouring cells, they could decrease RA signaling in single cells that have intermingled. Indeed, loss of cyp26b1 and cyp26c1 function disrupted the identity switching of egr2-expressing cells that have intermingled into adjacent segments. We showed that in r4 the switching involves upregulation of hoxb1, which in turn represses egr2 expression.
This work has revealed parallel mechanisms of identity switching and border control that establish sharp and homogeneous segments in the hindbrain. At early stages, some cells mix into adjacent segments and switch identity to match their new neighbours. This mediated by a community effect in which there is reciprocal feedback between RA levels and segmental identity. Subsequently, Eph receptors and ephrins are upregulated and they underlie segregation that prevents intermingling and sharpens the border.
These studies of hindbrain patterning raise the question of whether similar mechanisms operate in other tissues. Since mediators of cell segregation are often regulated downstream of regional identity genes, some intermingling between adjacent regions may occur at early stages. Furthermore, plasticity in cell identity is a common feature at early stages of development. Insights will come from the creation of further reporter lines to visualise cell intermingling and cell identity.
 (No Ratings Yet)
(No Ratings Yet)










 (1 votes)
(1 votes) I’m extremely happy to be acting as the new Graduate Representative for BSDB, following
I’m extremely happy to be acting as the new Graduate Representative for BSDB, following  Having completed my PhD in the field of vertebrate somitogenesis in the lab of
Having completed my PhD in the field of vertebrate somitogenesis in the lab of  Tanya is Professor of Developmental Biology at the University of Sheffield, where she is a member of the Bateson Centre and Department of Biomedical Science [
Tanya is Professor of Developmental Biology at the University of Sheffield, where she is a member of the Bateson Centre and Department of Biomedical Science [ Shankar is Professor of Developmental Biology and a Wellcome Senior Investigator in the Department of Physiology Anatomy and Genetics at the University of Oxford [
Shankar is Professor of Developmental Biology and a Wellcome Senior Investigator in the Department of Physiology Anatomy and Genetics at the University of Oxford [ Jens is a Sir Henry Dale fellow at the School of Life Sciences at the University of Dundee running his lab in the division of Cell and Developmental Biology [
Jens is a Sir Henry Dale fellow at the School of Life Sciences at the University of Dundee running his lab in the division of Cell and Developmental Biology [