An exciting opportunity to be part of a pioneering biomedical research institute, dedicated to innovation and science. A postdoctoral research post in the laboratory of Dr. Alex P. Gould is now available. We are looking for a highly motivated researcher to identify the, as yet unknown, molecular mechanisms by which dietary nutrients and oxidative stress during development can profoundly alter adult metabolism and longevity. The successful applicant will choose which of the Drosophila and/or mouse model(s) established in the lab they wish to focus on (PMID: 21816278, PMID: 26451484, PMID: 29123106 and unpublished). Research techniques used in the lab include genetics, molecular biology, confocal microscopy, biochemistry, metabolomics and mass spectrometry imaging. Details of projects currently being undertaken in the lab can be seen at: THE GOULD LAB. The successful applicant will have good communication and organisational skills and a PhD in a relevant area (or be in the final stages of completion). Informal enquiries to alex.gould@crick.ac.uk.
The Francis Crick Institute is a biomedical discovery institute dedicated to understanding the fundamental biology underlying health and disease. Its work is helping to understand why disease develops and to translate discoveries into new ways to prevent, diagnose and treat illnesses such as cancer, heart disease, stroke, infections, and neurodegenerative diseases. An independent organisation, its founding partners are the Medical Research Council (MRC), Cancer Research UK, Wellcome, UCL (University College London), Imperial College London and King’s College London. The Crick was formed in 2015, and in 2016 it moved into a new state-of-the-art building in central London which brings together 1500 scientists and support staff working collaboratively across disciplines, making it the largest biomedical research facility in one building in Europe. The Francis Crick Institute will be world-class with a strong national role. Its distinctive vision for excellence includes commitments to collaboration; developing emerging talent and exporting it the rest of the UK; public engagement; and helping turn discoveries into treatments as quickly as possible to improve lives and strengthen the economy.
” If you are interested in this role, please apply via our website.
” The closing date for applications is Monday, 19 March 2018 at 23:30.
” All offers of employment are subject to successful security screening and continuous eligibility to work in the United Kingdom.
Key words
” Developmental biology
” Mouse genetics
” Drosophila genetics
” metabolism
” physiology
” developmental origins of health and disease
” mass spectrometry imaging
” metabolomics
” cell biology
” molecular biology
The droso4schools initiative is an educational outreach programme run by the Manchester Fly Facility, which I had the opportunity to be actively involved in during a month-long placement (see my previous blog). The aim of this programme is to re-introduce the fruit fly Drosophila melanogaster into school and college biology lessons. For this, the droso4schools team have developed a number of hands-on, readily prepared, interactive lessons with adjunct materials, which explain curriculum-relevant biology contents at the level of Key Stages 4 – 5. As has been explained in a recent blog and publication, these lessons make active use of Drosophila as a teaching tool – both to explain and illustrate contents and to perform micro experiments.
The Manchester Fly Facility team recently hosted a continuing professional development (CPD) day for a group of Key Stage 3, 4 and 5 science teachers, to showcase their existing lesson packages (and more are in production…). This was an important opportunity for the droso4schools team to explain their resources and gain feedback from a focus group of experienced teachers – namely, what are the lessons like to use? Could they be improved? Most importantly, are they curriculum-relevant?
The day began with a tour of the fly facility, a short activity using microscopes to identify common phenotypic markers used in Drosophila research, and an introduction to the droso4schools programme by academic lead Professor Andreas Prokop, and long-time collaborator Dr Catherine Alnuamaani, a teacher and keen Drosophila advocate from Trinity CoE High School, Manchester. Andreas and Catherine together explained the mutual benefits of using Drosophila as a teaching tool: the benefit for pupils and teachers lies in the fact that Drosophila provides uniquely detailed biology knowledge, delivering concepts, stories and experiments across biology – paired up with human examples to illustrate relevance. Researchers benefit in that students are introduced early on to the power of research with genetic model organisms as a key driver for scientific discovery, and will then hopefully be more open to such research strategies when entering university.
As Catherine highlighted, teaching practicals can become a little monotonous, and the experiments themselves are rarely very exciting. Using a live animal in the classroom puts the “life” back into the life sciences, and encourages pupils to engage with the subject content – Drosophila brings real-world relevance to the curriculum. Not only that, it also provides an opportunity for teachers to develop experimental skills, in a way that is both cost-effective and more interesting than starch-storing potatoes. But what benefit is there for researchers? As Andreas explained, many students who arrive at university to study life sciences, often don’t have much of an interest in invertebrates – they are attracted to applied vertebrate biology and clinically oriented research. Of course, this kind of research is important, but it is fundamental biology which forms the basis on which “translation” can occur. By demonstrating to school pupils the potential scope of fruit fly research, it enhances the chance that those who choose to study the life sciences at university, will have more of an appreciation for the impact of Drosophila research on modern science, and be keen to explore the field themselves.
The itinerary for the day included three full lessons in which the participating teachers took part from the student perspective. Each lesson consisted of a well-structured PowerPoint presentation guiding through the contents, interspersed with short activities and micro experiments. A couple of the participants had been using flies from the facility in their lessons already, mainly as a means of demonstrating genetic crosses; one had even been using elements from the “alcohol lesson” (see below), with great success, for two consecutive years. These teachers reported that students loved having live animals in the classroom, and so they had come to explore other options. The rest of the group were relatively new to the idea of Drosophila as a teaching tool. Some knew how they’d like to use the flies, but weren’t sure where to start. Others were curious, but not sure how successful flies could be as a learning resource, and how easy it would be to source materials.
Being in a classroom full of teachers is something of a surreal experience, but everyone was more than happy to get stuck in, ask questions, work in groups, and contribute to class discussions. The first lesson, concerning the fundamentals of nervous system organisation and function (“Principles of the Nervous System”), gave the participating teachers their first opportunity to work with live flies, through a series of micro behavioural experiments involving the shaking of epileptic flies into seizure (an experiment that is used to illustrated wiring principles and the mechanism of nerve impulses), and another causing transient paralysis of shibire mutant flies (which involves keeping vials of flies under your armpits for five minutes … always popular with pupils!).
Lesson two (“From gene to enzyme to evolution: using alcohol metabolism to illustrate fundamental concepts of biology”) is a fantastic revision lesson to use before the A-level exams, lining up a broad range of specifications, from gene expression to enzymes to evolution, into a consistent story. It contains a particularly hands-on practical, studying enzymatic reactions using the dissection of Drosophila larvae followed by a 5 minute staining reaction, which was very well received by the participants. This practical also provides a means to demonstrate the potential impact alcohol consumption can have on health, providing opportunities to discuss this topic most relevant to students of that age.
The third lesson (“Our vision: understanding light and light perception”) was of particular interest to me, as I had worked on its experimental component during my placement at the Manchester Fly Facility. I used the fact that flies are attracted to light (an innate behaviour called positive phototaxis) to test responses to different wave-lengths, using normal and sevenless ‘colour blind’ flies. This experiment is particularly effective, as taxis behaviours are simple automatic responses to a given stimulus, and therefore easy for pupils to identify and observe. I had some great feedback from the teachers, a number of whom commented on the short fly behaviour video, and how easy it would be for pupils to engage with – which was very rewarding! Most importantly, it was later highlighted that the observation of taxis behaviours is now a compulsory part of the AQA A-Level biology practical syllabus – so if any teachers are reading this, it is worth taking a look at this lesson!
A brief overview of another lesson (“The climbing assay: learning data analysis through live experiments with fruit flies”) was received with particular enthusiasm by everyone and sparked imaginations. This lesson makes use of a climbing assay that compares motor activities of young versus old flies to perform statistical analyses. Many of the teachers seemed quite excited to use this particular lesson with some of their younger (KS3) classes, as well as for the GCSE and A-Level students, as it is so easily translatable. Not only would it be straightforward to teach, but all agreed it would be a great way to engage pupils with what is traditionally not the most interesting part of scientific methodology.
To complement the lesson sessions, Sanjai Patel (manager of the Manchester Fly Facility) provided a short masterclass on basic fly handling and care techniques, in which teachers were given flies and had to tip them over into fresh vials. Everyone seemed to get the hang of it quite quickly, and there were hardly any escapees! Finally, teachers were given a “guided tour” through the facility’s droso4schools website (describing the resources and given additional support information), the teachers’ section of the Manchester Fly Facility website, and the figshare.com repository providing all the teaching resource files for download.
At lunch and during the concluding session of the day, there was a chance to consolidate the day’s information, reflect on the lessons, share thoughts and opinions on the ease of using Drosophila as a teaching tool, and provide feedback to the team. It was very inspiring, and also in many ways reassuring, to talk to teachers from different schools and areas, who were similarly passionate about their career choice, and their roles as science advocates. During the discussion, a number of excellent ideas were raised, and it was great to see how enthusiastic all the participants were about the project, and how keen they were to get back in the classroom and try the lessons for themselves! Positive feedback included:
how easily adaptable all the resources were – images, lessons, and other materials can be downloaded and utilised separately, or in whatever combination is convenient.
how easy it was to link the Drosophila practicals to current research, providing that real-world edge so often missing in everyday lessons.
that the lessons provide an excellent resource for pupils practicing the new synoptic essays (a synoptic essay assesses a pupil’s ability to demonstrate and apply their understanding of a topic, using a variety of examples on a given theme); often, this additional knowledge is obtained outside of the classroom, through private study.
that the depth and breadth of information available about Drosophila is easy to incorporate as “additional” information into revision lessons.
An area for improvement mentioned by some of the participants, was that there was something of a “language barrier”, i.e. the differences in basic terminology used in research, compared to that used in schools. Other barriers mentioned were the complex technicalities of the GSCE and A-level curriculum. As a potential solution to tackle these barriers, it was suggested that an experienced teacher could become involved with the droso4schools programme, to “fine-tune” the resources. As it was put by one participant: “You are almost there, but it now needs someone with A-level experience to fully adapt them for school use”.
Overall, the day was very uplifting, and all seemed to head for home brimming with new ideas – be it how to introduce flies to their lessons, or how to further improve available resources. Science lessons should never be boring – neither for pupils nor for teachers. Introducing Drosophila to the classroom provides a new and exciting way to observe science in action, in a way which is convenient, affordable, and manageable. Enthusing pupils about the possibilities presented by research with this powerful invertebrate could also help to inspire a new generation of fundamental life scientists and, who knows, maybe even a few more Nobel Prize winners!
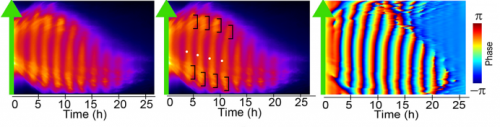
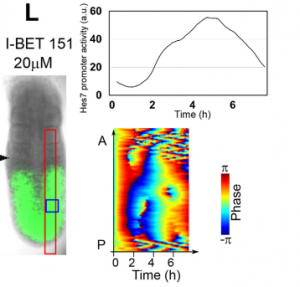
Vertebrate segmentation involves the periodic formation of somites from the presomitic mesoderm, in a manner controlled by oscillating gene expression (the oscillations of the segmentation clock must be one of the marvels of nature!). While in vivo work has provided a framework for studying the process, many aspects of segmentation dynamics are obscured in the embryo. A new Techniques & Resources paper in Development describes a stem cell-derived in vitro system for studying segmentation dynamics. We caught up with first author Marina Matsumiya and her supervisor Ryoichiro Kageyamaof Kyoto University to hear more about the story.
Ryoichiro and Marina
Ryoichiro, can you give us your scientific biography and the questions your lab is trying to answer?
RK I became Assistant Professor of Kyoto University in 1989 and started a project about transcription factors that regulate mammalian neural development. We reported the basic helix-loop-helix (bHLH)-type repressor genes Hes1, Hes3, and Hes5 in 1992 and the bHLH-type proneural gene Atoh1 (Math-1) in 1995, and since then we have been characterizing the functions of these genes in neural development. We found that the antagonistic regulation between Hes and proneural genes controls neurogenesis and gliogenesis.
In December 1997, I was promoted to full professorship in the same university and continued the bHLH project. In the same month, I met Olivier Pourquié, when he visited my group in Kyoto, and we had discussion about his finding (just published in Cell) that the expression of chick hairy1, a Hes1 homologue, oscillates during somite segmentation. This discussion made me keep pondering on how such oscillatory expression with ultradian rhythms is possible and how such dynamic expression regulates the downstream events. In the 2000s, we reported that Hes1 expression oscillates in many cell types by negative feedback, and that this oscillation is important for cell proliferation. In parallel with this finding, we identified a new member of the Hes family, Hes7, and found that its expression oscillates by negative feedback in the presomitic mesoderm (PSM), thereby regulating the somite segmentation. These findings brought us to the field of the segmentation clock. Hes7 oscillations are synchronized under the control Notch signaling, and the oscillatory expression of the Notch ligand Delta-like1 is involved in the synchronized oscillations. However, the detailed mechanism of the synchronized oscillation with traveling waves in the PSM is still unknown, and we have been trying to answer this key question.
What is the status of developmental biology research in Japan?
RK Developmental biology research is well funded in Japan if a stem cell or regenerative research field is included. However, besides this field, it seems to be more difficult to get sufficient funding. I think that translational research-oriented funding is a general trend in Japan, as observed world-wide. Collaborations are very active and actually essential to many research projects of developmental biology. Grants supporting domestic and international collaborations are also available. In addition, Riken offers a service of generation of transgenic mice as a collaboration basis, and many researchers utilize this service. I do not know the exact number of labs in developmental biology in Japan, but the Japanese Society of Developmental Biologists (JSDB) has about 1200 members. This is a good number, compared to The Society of Developmental Biology (USA), which has nearly 2000 members.
However, I am a bit concerned about the recent trend. There used to be more than 300 poster presentations in the Annual Meeting of the JSDB, but last year, there were only 193, suggesting that the number of young researchers in the field is decreasing over the years. One reason is that there were only a handful of poster presentations of the stem cell or regenerative research field in the Annual Meeting of the JSDB, while many Japanese researchers participated in the ISSCR in Boston last year. I feel that (classic) developmental biology and (fancy) stem cell or regenerative research should go hand-in-hand, but in reality these two fields seem to go separately in Japan. It would be desirable to incorporate more stem cell researchers into the JSDB meetings to advance the field properly.
And Marina, how did you come to be involved with this project?
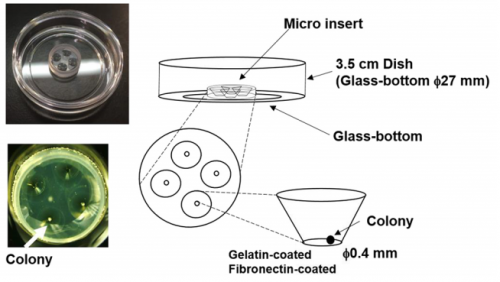
MM I was interested in the mechanism of somitogenesis, so I joined the Kageyama Lab as a master student. There were no PSM-like cell culture systems, and my colleague, Kumiko Yoshioka-Kobayashi, suggested me that such systems would be important to examine the detailed mechanism of somitogenesis. After discussion with Kumiko and RK, I decided to make a PSM-like cell culture system. At almost the same time, I found that Olivier’s group published a method to induce PSM-like cells from mouse embryonic fibroblasts (MEFs), so we first used MEFs produced from our Hes7 reporter mice. However, we were not able to induce Hes7-expressing cells, and we next tried mouse embryonic stem (ES) cells following the paper from the same group published in 2015, which focused on the induction of muscle fibers from ES cells. We also had a chance to learn an ES cell culture method from Mototsugu Eiraku at Riken CDB, who established a method to induce the eye cup from ES cells. By combining these methods, we succeeded in inducing PSM-like cells, which exhibited Hes7 oscillations.
Matsumiya, et al.’s culture method for imaging Hes7 expression in iPSM colonies, from Figure 1 in the paper.
What are the difficulties of studying the segmentation clock in embryos? And what were the limitations of previous in vitro approaches?
RK Detailed analysis of the segmentation clock has been hampered because it requires the use of live embryos. Analysis involves both genetic and pharmacological approaches: in genetic approaches, mutant animals are generated to activate or inactivate gene functions, while in pharmacological approaches, either embryos or PSM tissues prepared from embryos are used for treatment with chemicals that modulate gene activities. If in vitro cell cultures are available, both genetic and pharmacological analyses would be facilitated, and many attempts have been made to induce PSM-like tissues from ES cells. Although PSM-like tissues have been successfully induced from ES cells, there are no reports of wave-like propagation of oscillatory gene expression in such induced tissues. So, we started the in vitro PSM cell project.
Hes7 expression in an iPSM colony cultured in a fibronectin-coated plate; Movie 2 from the paper.
Can you give us the key results of the paper in a paragraph?
MM & RK We established a simple and efficient method to generate mouse ES cell-derived PSM-like tissues, in which Hes7 expression oscillates like traveling waves. In these tissues, Hes7 oscillation is synchronized between neighboring cells, and the posterior-anterior axis is self-organized as the central-peripheral axis, generating somite-like segments at the periphery. This method is applicable to not only chemical library and RNAi screening but also CRISPR-Cas9-mediated gene modifications and will facilitate the analysis of the molecular nature of the segmentation clock.
In your system, Hes7 oscillations did show some variability: do you know what underlies this variability?
MM & RK There is some variability in the amplitude, period, and patterns of Hes7 oscillations even in the same iPSM colonies, which may hamper to detect effects of chemicals and genetic modifications. This variability may depend on the size and shape of iPSM colonies, and how iPSM colonies expand on dishes. Further improvements will be required to reduce such variability.
Kymographs of Hes-7 driven luciferase, from Figure 3 in the paper.
Is your system restricted to mouse cells, or could you use it to compare segmentation between species?
MM & RK We have not examined other species yet, but we are sure that this method may be applicable to ES/iPS cells of other species. We are interested in the mechanism of how species-specific periods of Hes7 oscillations are determined.
When doing the research, did you have any particular result or eureka moment that has stuck with you?
MM My eureka moment is when I observed Hes7 oscillation in iPSM colonies for the first time. I still remember the date, and we used the data in Figure 4 of the paper. We did not get nice data for a few years, but after changing the culture conditions, all of sudden, everything went well. We were surprised to see the Hes7 oscillations propagating like waves and leading to somite-like segment formation. When I saw the movie, we celebrated with high five. I could watch it forever. It took 6 months from the first detection of Hes7 oscillation to submit the paper. Every day in this half a year was a series of joy.
Validating in vitro results in the embryo, from Figure 7 in the paper
And what about the flipside: any moments of frustration or despair?
MM When I used MEFs, I was really nervous. I got some Hes7 signals but not oscillatory expression. I tried many different culture conditions, but none of them worked. It was like a never-ending nightmare, but now I feel that it is a very important step to improve myself mentally and technically.
What next for you Marina?
MM I have one more year to go before I finish my PhD course. Until then, I will try to reveal new factors or pathways that make Hes7 oscillations synchronized by using the iPSM method. This information will be important for the somitogenesis research. After finishing my PhD course, I would like to do research on refractory diseases of bone formation related to somitogenesis. Now, I will prepare the application of a fellowship to become a postdoc.
Where will this work take the Kageyama lab?
RK By using this system, we now started screening many chemicals to find a new pathway involved in synchronization of Hes7 oscillations. We also plan to use two-color fluorescent reporters (one from Hes7 promoter and the other from another important PSM gene) to examine the oscillation at the single-cell resolution. We are trying to find chemicals that do not repress Hes7 expression but affect synchronization. These chemicals, if found, would help reveal the mechanism of synchronized oscillation besides the Notch pathway. We have recently developed an optogenetic gene induction system and plan to introduce this system to iPSM cells. This system allows precise spatiotemporal control of gene expression, so we would like to see how light-controlled gene expression affects Hes7 oscillations.
Finally, let’s move outside the lab – what do you like to do in your spare time?
MM I would like to sleep, but I also like playing modern ballet. These are very important for me to control my condition.
RK I like jogging along the Kamo river in Kyoto, which really refreshes me. I also like walking. I used to bike, but after an accident I stopped biking now.
In spite of the difference between the cell functions responsible for giving rise to a tumour and for the metastasis of this same tumour, studies at IRB Barcelona using the fly Drosophila melanogaster reveal that some genes can drive both phenome
na.
In cancer research, there has been a tendency to differentiate between the genes responsible for tumour development and those that give rise to metastasis and to study these processes separately since the cell functions required for these two mechanisms differ radically. Researchers headed by Jordi Casanova at the Institute for Research in Biomedicine (IRB Barcelona) now demonstrate that genes activated during metastasis are also able to initiate primary tumour development, and they explain the molecular mechanism involved. Made using the fly model Drosophila melanogaster, this finding has been published in PloS Genetics this week.
“The results strengthen the notion that was already being put forward in cancer research,” explains Jordi Casanova, head of the Development and Morphogenesis in Drosophila lab and CSIC professor. He goes on to say “And that is that the same genes can activate tumour growth and metastasis, and therefore some tumours are conferred metastatic capacity from their very outset”.
Two phenomena that share genes
The study is based on the observation that benign human tumours or tumours at stages well before metastasis express some genes associated with this process. “This overlap made us question whether the so-called epithelial-mesenchymal factors—which activate genes associated with cell movement and migration, which are common in embryo development and are also present in metastasis—play a role in tumour initiation,” says Casanova.
In particular, GATA genes are overexpressed in liver metastasis in colon cancer patients, as well as in several human tumours, such as those of the breast, ovary, pancreas and colon. Jordi Casanova’s team examined various genes, among them Serpent (homologue of GATA in Drosophila), and demonstrated that the overexpression of this gene is indeed sufficient to drive tumour growth. In this regard, this gene causes overgrowth of the wing, modifies tissue structure, and activates a series of genetic changes in order to facilitate initial tumour growth.
The fly wing as a model of epithelial tumour
The research was conducted on the fruit fly Drosophila, an established model for studying epithelial tumours. “Once again, studies on Drosophila have contributed to our basic understanding of the behaviour of genes involved in cancer and metastasis, knowledge that will allow cancer researchers to pursue this line further,” says the scientist.
Kyra Campbell, first author of the study, former postdoctoral fellow at IRB Barcelona and currently at the University of Sheffield, in the UK, was supported by Xavier Franch-Marro, at the CSIC’s Institute of Evolutionary Biology.
This work was supported by the Ministry of the Economy and Competitiveness, through ERDFs, and by the Government of Catalonia.
The Beddington Medal is the BSDB’s major commendation to promising young biologists, awarded for the best PhD thesis in Developmental Biology defended in the year previous to the award. Rosa Beddington was one of the greatest talents and inspirational leaders in the field of developmental biology. Rosa made an enormous contribution to the field in general and to the BSDB in particular, so it seemed entirely appropriate that the Society should establish a lasting memorial to her. The design of the medal, mice on a stylised DNA helix, is from artwork by Rosa herself. We would like to congratulate the 2018 winner of the Beddington Medal, Emilia Favuzzi, and would like to take this opportunity to give a brief overview of her career and her PhD project that was awarded the Beddington medal.
Emilia’s career and project
Emilia started her studies in 2007 at the Sapienza University of Rome and was awarded a B.Sc. in Biological Sciences with highest marks in 2010. She stayed at the same university for her Master’s project which she performed in the laboratory of Sergio Nasi at the Institute of Molecular Biology and Pathology (CNR, Rome). She completed her M.Sc. in Neurobiology in 2011, again with highest marks. In 2011 she joined the group of Beatriz Rico at the Institute of Neuroscience in Alicante (Spain) and moved with that group to the Centre for Developmental Neurobiology at King’s College London in 2014 where she terminated her project work. Her PhD in Neuroscience was awarded in 2017 by the University Miguel Hernandez of Elche (Spain) also with summa cum laude. Since 2017 she is a postdoctoral associate in Gordon Fishell’s laboratory at the Broad Institute and Harvard Medical School.
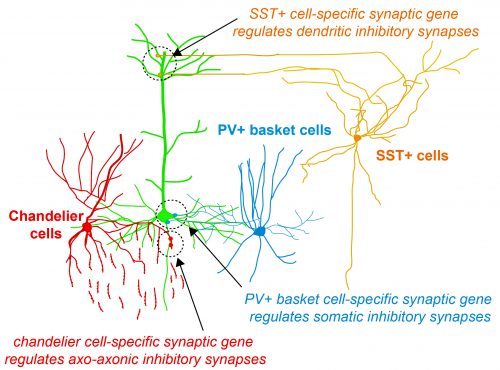
Fig.1 Activity-dependent gating of parvalbumin interneuron function by the perineuronal net protein Brevican
During her PhD, Emilia worked on two projects which were both based on candidate and genome-wide screen approaches aiming to identify genes that were involved in GABAergic synapse formation. In one project, she investigated the role of perineuronal nets during the synaptic development of GABAergic interneurons. She discovered that the perineuronal net component Brevican is involved in the gating of parvalbumin interneurons by controlling their intrinsic properties as well as extrinsic input through excitatory synapses (Fig.1). This paper was published as a featured article in Neuron (2017). Emilia also took ownership within a parallel project, where she collaborated with another lab member to set up protocols to isolate different populations of interneurons and screen for genes involved in the specific synaptic targeting of cortical interneurons to the different compartments of pyramidal cells. This work led to the discovery of validated candidate genes involved in specific interneuron synapse formation, as shown via loss and gain of function approaches (Fig.2). The respective manuscript is in preparation and Emilia will be shared first author.
Her PhD supervisor Beatriz Rico said about her: “Emilia is a gift for a supervisor: she goes ahead of you, technically and conceptually and pushes you forward. She is brilliant, extremely motivated and creative person and resistant to any difficulties she has found during the development of her project. She never gave up and pursues her aims with an impressive efficiency. She is extremely independent and hard worker. She is fully committed to science, a dream for a supervisor.”
Thesis abstract: Cell-type specific programs regulate the assembly and dynamics of cortical circuits
Understanding how neuronal connections are established and organized in functional networks during development is critical to understand brain function. In the mammalian cortex, GABAergic interneurons are characterized by a remarkable diversity of types and connectivity patterns. As such, they are uniquely suited to orchestrate functionally relevant circuit-specific roles and critically shape cortical function. Yet, how inhibitory circuit specificity is achieved during development is largely unknown. We revealed the transcriptional dynamics of different cortical interneurons during brain wiring and identified subtype-enriched synaptic molecules. Moreover, we showed that the functional connectivity of different interneurons relies on the cell-specific expression of such synaptic genes. Altogether, our results demonstrate that highly selective molecular programs emerging during development in cortical interneurons support their early wiring and underlie inhibitory circuit specificity. After their integration into canonical circuits, activity-dependent plasticity endows neurons with the flexibility required for adapting to sensory experience. Parvalbumin (PV+) interneurons have been shown to play a critical role in this process but the molecular mechanisms by which experience influences PV+ interneuron plasticity were poorly understood. We revealed how perineuronal net (PNN) proteins drive PV+ cell wiring as well as network adaptation to experience. We showed that the PNN protein Brevican simultaneously regulates the excitatory inputs and firing properties of PV+ interneurons by controlling the localization of AMPA receptors and potassium channels, respectively. We also showed that, by modulating Brevican levels, experience influences cellular and synaptic forms of plasticity in PV+ cells and this is required for normal cognitive function. These findings uncover a cell-specific molecular program through which a PNN protein dynamically gates PV+ interneuron function both during development and upon experience-dependent plasticity.
Papers by Emilia so far (* co-first authors)
Favuzzi E*, Deogracias R*, Marques-Smith A, Maeso P, Exposito-Alonso D, Balia M, Jezequel J, Kroon T, Hinojosa AJ, Rico B. Highly selective cell-type specific programs regulate structural synapse target specificity (manuscript in preparation)
Favuzzi E, Marques-Smith A, Deogracias R, Winterflood CM, Sánchez-Aguilera A, Mantoan L, Maeso P, Fernandes C, Ewers H, Rico B. Activity-dependent gating of parvalbumin interneuron function by perineuronal net proteins. Neuron (2017)
Marques-Smith A*, Favuzzi E* & Rico B. Shaping Early Networks To Rule Mature Circuits: Little MiRs Go A Long Way. Neuron (preview), (2016)
Annibali D*, Whitfield JR*, Favuzzi E, Jauset T, Serrano E, Cuartas I, Redondo-Campos S, et al. Myc inhibition is effective against glioma and reveals a role for Myc in proficient mitosis. Nature Communications (2014)
Savino M, Annibali D, Carucci N, Favuzzi E, Cole MD, Evan GI, Soucek L, Nasi S. The Action Mechanism of the Myc Inhibitor Termed Omomyc May Give Clues on How to Target Myc for Cancer Therapy. PLoS One (2011)
Cell polarization defines the spatial biological specificities in a cell. During the first cell cycle of a C. elegans zygote, its symmetry is broken by local remodeling of the cortical actomyosin network. This leads to a segregation of the dedicated polarity regulators, the PAR proteins, into two discrete cortical domains. However, it remains unclear how the mechanical changes driven by actomyosin contractions are transmitted to PAR proteins, and this regulates cellular spatial patterning. In our paper, we revealed that both actomyosin contractions and CDC-42 activity modulate the dynamics of the anterior PAR proteins (aPARs) through stimulation of their clustering at the cell cortex. In the early phase of polarity establishment, contractility of the actomyosin cytoskeleton drives cortical tension, which promotes clustering of PAR-3 (Mov.1, Fig.1). In turn, PAR-3 clusters recruit PKC-3, whereas CDC-42 activity antagonizes PKC-3 clustering during later phase of polarization (Mov.2, Fig.2). The degree of aPARs clustering is significantly associated with the stagnation of aPARs exchange at the cortex, and an effective entrainment with the advective flows of the cortical actomyosin network. This is the first report showing that the cell polarity protein PAR-3 is mechanosensitive, and senses forces to form higher order structures at the cell cortex. These findings depict how actomyosin and PAR proteins interplay and set up asymmetry (Fig.3). This feature may also play a role in other cell patterning events including neuron maturation, wound healing and cancer formation.
Movie 1: Cortical PAR-3::GFP (green) and NMY-2::mKate2 (magenta) during the first cell cycle of a C. elegans embryo.
How I started this project
I had made up my mind to become a researcher during my undergraduate days, after first trying to do a simple plasmid construction in a biological laboratory. Back then I was fascinated by all the equipment and materials in the lab. I went on to obtain my Master’s and Ph.D. degrees in Virology and Neuroscience respectively, before moving to Singapore to commence my first postdoctoral training in Frederic Bard’s lab at the Institute of Molecular and Cell Biology (IMCB), A*STAR. There, I matured as a scientist, and with access to multiple advanced microscopes across the various institutes within the campus, I was able to develop my working skills and knowledge in live-imaging techniques. These abilities, have turned out to be a great help in my career so far.
Before joining Fumio Motegi’s lab at the Temasek Lifesciences Laboratory (TLL), I had worked for over 10 years using cell culture to address different biological questions. So, I had a strong feeling that I must do something different for my next post-doc training. However, it was only after Fumio showed a fantastic movie depicting how anterior and posterior PARs are segregated in worm embryos that I decided to pursue this new area of study. My interests had been captured by how complex, yet exquisite, the underlying regulations can be. After I started working on this project and got some beautiful images of the genome-edited GFP-tagged aPARs, which were generous gifts from Kenneth Kemphus from the Cornell University, I realized that C. elegans is such a powerful tool with which we can address fundamental and important questions. I was also encouraged by the worm society, who kindly shared their materials and information, and welcomed me as a newcomer in their community. I was confident that this was what I wanted to do.
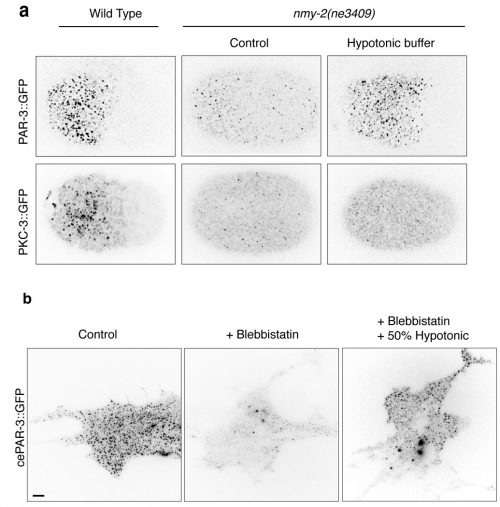
Fig. 1: Cortical tension driven by osmotic pressure change restores aPARs clustering in NMY-2 mutants. (a) Cortical images of PAR-3::GFP and PKC-3::GFP in wild-type or nmy-2(ne3409) C. elegans zygotes with or without hypotonic buffer treatment. (b) Images of live NIH3T3 fibroblast cells expressing GFP tagged-C. elegans PAR-3. Cells were treated with DMSO (control) or Blebbistatin for 30 min. GFP::PAR-3 at the superficial sections were imaged by TIRF microscopy.
How I found the key results in this paper
The first Eureka moment came when I was inspecting the cortical PAR-3::GFP and PKC-3::GFP in wild-type embryos. They showed beautiful and dynamic puncta structures. This led us to further characterize the implication of those puncta during polarity establishment. We then conducted intensive examinations on the clustering of aPARs in wild-type and polarity-mutant zygotes. Another moment of insight came to me when my colleague Ms. Tricia Low noticed that PAR-3::GFP may exhibit mechanosensitivity. PAR-3::GFP failed to form clusters in the non-muscle myosin-II (nmy-2) mutant embryos, suggesting actomyosin and PAR-3 clustering were somehow connected. The clues became clearer when we observed another two phenomena. Firstly, in wild-type embryos, PAR-3 did not reside with NMY-2, suggesting that PAR-3 clustering is not caused by the direct recruitment of PAR-3 molecules to actomyosin. Secondly, although PAR-3 was unable to form clusters in nmy-2 zygotes, PAR-3 clustering was restored when the embryos underwent shape changes, with cortical ruffling occurring during the later stage of embryogenesis. This suggested that actomyosin contractility itself was not essential for PAR-3 clustering. Instead, physical properties of the cortex appeared to play a crucial role in the control of PAR-3. Hence, we hypothesized that cortical tension driven by actomyosin could promote PAR-3 clustering. We tested and validated this assumption in worm zygotes, as well as mammalian fibroblasts NIH3T3 cells exogenously expressing C. elegans PAR-3, which is achieved with the fantastic help from Dr. Yukako Nishimura from the Mechanobiology Institute (MBI), Singapore. We found that in both cases, PAR-3 clustering was rescued under hypotonic treatment when myosin wasn’t functional. This is the first report stating that PAR-3 reorganizes to higher-order structures in response to tension!
Movie 2: GFP tagged PAR-3, PKC-3, and CDC-42 during polarity establishment in one-cell C. elegans embryos.
How we build a team to test our hypothetical model
When we discovered that PAR-3 carries mechanosensitivity and that it could sense and respond to changes in tension in worm zygotes, we wanted to validate and replicate this experiment in cultured cells and monitor PAR-3 clustering using a TIRF microscope. In the meantime, we also needed a proper method to quantify and demonstrate the clustering so that we could report its significance under different conditions. To allow the research to progress without delay, we decided to collaborate with researchers outside of our lab. The MBI and the IMCB instantly came to mind. At MBI, there are dozens of experts in the field of physical biology, as well as a comprehensive collection of advanced microscopes. We therefore felt there was no better choice for collaboration. Moreover, I had established tight connections with people in the IMCB during my post-doctoral training there. In particular, with the Computational Bioimage Analysis (CBA) Unit, which is run by truly reliable and efficient staffs who are doing great jobs in image analysis and quantification. I was fortunate to have the opportunity to establish fruitful collaborations across TLL, MBI, and IMCB. Each institute is highly-respected not only in Singapore, but worldwide, for consistently publishing in high-profile journals, and attracting talented scientists from different countries. Singapore had become one of the most splendid research hubs in the world, which is attributed to the massive investment and strong support from the government into the life science realm.
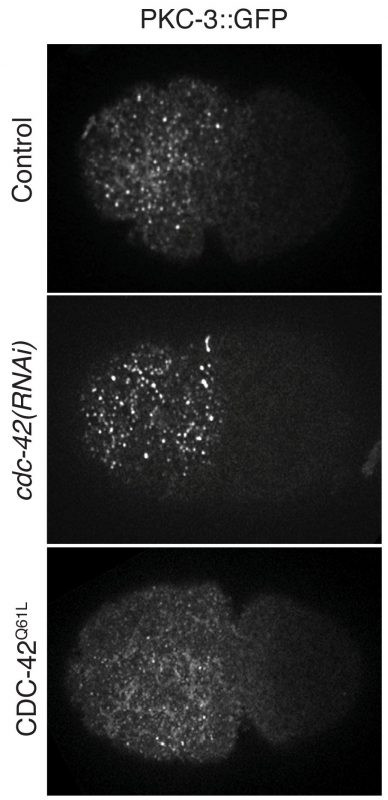
Fig. 2: PKC-3::GFP clustering under loss- or gain-of-function of CDC-42 conditions during the establishment phase of one-cell C. elegans zygotes.
How we struggled to publish our project
I think the most difficult time for us along the journey was when our manuscript was rejected by the journal in December of 2016. The reviewers appreciated the main point of our story, but they thought the quantification method we had used to measure aPARs clustering was insufficient. Time was a big concern as we considered whether to submit to another journal, which might have been the easier path to publication, or stick with the same journal and strengthen our manuscript. We were in a dilemma. But since we were confident in ourselves, and in our story, we immediately made the decision to amend the quantification method and reanalyze all of our images. Fortunately, the results returned by the modified methods were consistent with our previous conclusions. We quickly revised the manuscript and resubmitted to the same journal and it turned out to be a happy ending. We appreciate Dr. Weimiao Yu and Dr. Laurent Gole from the Computational Bioimage Analysis (CBA) Unit of IMCB for their efforts to come up with the modified code, as well as the detailed descriptions for the quantification method, for the revised manuscript. We are also very grateful to the editor and all our reviewers for their constructive comments and positive encouragements to improve our manuscript. We did enjoy the review process. I hope what we had gone through can be an inspiration to those who might currently be struggling in a difficult time like this. What we need to do is to keep calm and believe in yourself.
Publication of our work along with two other groups
We learned there were several groups working on a similar topic since 2016. Fortunately, we maintained unencumbered communication with these groups, through emails or in person during international conferences, and it is truly exciting to see that we have all successfully published our stories in high-impact journals.
The common conclusion drawn by the three groups is that the degree of PAR-3 oligomerization increases during polarity establishment, and this further promotes the formation of aPAR clusters. The clustering of aPARs supports its segregation along with the actomyosin flow. Moreover, single-molecule pull-down in single embryos, as well as particle tracking, were used by Dickinson et al. to demonstrate that clustered PAR-3 is coupled to the cortical flow and phosphorylation by the mitotic kinase PLK-1 prevents PAR-3 from undergoing oligomerization. Furthermore, through a functional assay, Rodriguez et al. showed that PKC-3 inhibition or activation depends on its interaction with pools of PAR-3 or CDC-42, respectively. We found that PAR-3 carries mechanosensitivity that responds to cortical tension driven by actomyosin contraction. It’s genuinely fantastic that all three papers support each other and yet, by employing distinct approaches, present a unique significance to the questions addressed. I believe this can be a good example to encourage those who may also be facing competition in their research.
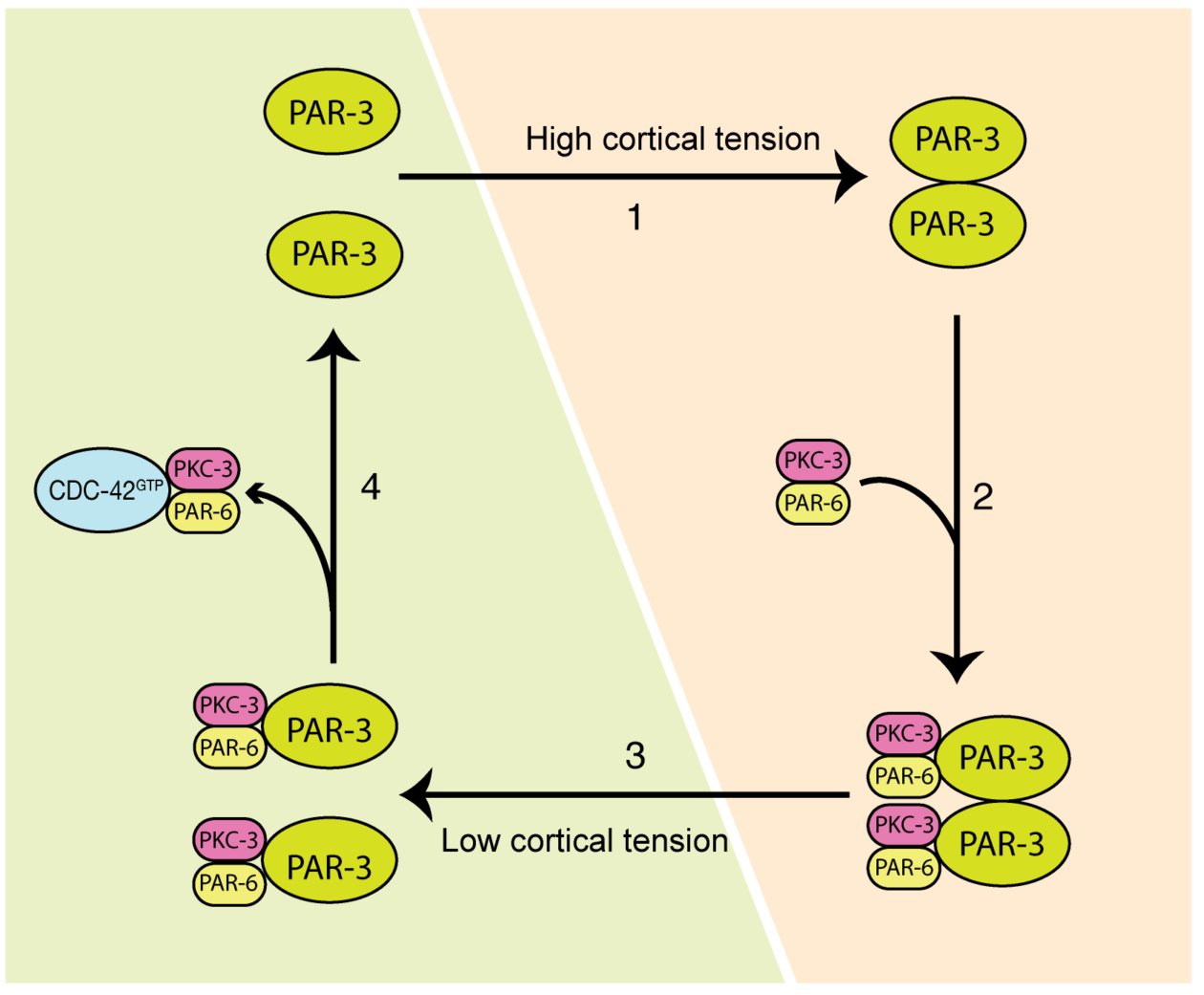
Fig. 3: A model depicting two-phases aPARs dynamics regulated by cortical tension driven by actomyosin and CDC-42 activity during polarity establishment. (1) Under a high cortical tension (orange panel), PAR-3 oligomerizes and forms clusters (the number of PAR-3 unit in the depicted model is not reflecting reality). (2) Clustered PAR-3 recruits the PKC-3/PAR-6 complex. (3) When actomyosin is less active (green panel), which leads to a lower cortical tension, PAR-3 clustering is weakened and its binding affinity to the PKC-3/PAR-6 complex is reduced. (4) Active CDC-42 recruits the PKC-3/PAR-6 complex from its cytosolic pool that is freed up from PAR-3 or by direct competition with PAR-3/PKC-3/PAR-6 complex.
Several open questions about cell polarisation
The evolutionarily conserved PAR machinery is predominantly localized at the actomyosin cortex, which lies adjacent to the cell membrane. It is likely that the principle we uncovered is applicable to other types of tissue, and other organisms. For instance, it has been shown that self-assembly of the PAR-3 N’-terminal domain is critical for axon specification of neurons. In this case, the direct binding of the oligomerized domain to the microtubule promotes microtubule bundling and stabilization. It would be interesting to test if this formation of higher-order PAR-3 can be promoted by the activation of non-muscle myosin, which in turn changes local cortical tension. Another example is during Drosophila embryogenesis. Here, during the process of dorsal closure, actomyosin repeatedly assembles and disassembles in the epithelial amnioserosa cells. In this case, PAR-3 forms patches and overlaps with myosin at the apicomedial surface to promote actomyosin contraction. It is intriguing to explore whether myosin assembly induces structural changes in PAR-3, and whether this contributes to developmental morphogenesis. Finally, some follow-up questions remain elusive and need to be further addressed. What are the molecular mechanisms by which PAR-3 clustering or CDC-42 activity apply to compete for the PKC-3/PAR-6? How do aPARs clustering and actomyosin activity regulate and feedback to each other with a temporal-spatial precision during cell-cycle progression? It would be exciting to see emerging studies resolving more of the puzzles surrounding cell patterning in response to extrinsic and intrinsic stimulation in the near future.
References
Wang, S.C., Low, T.Y.F., Nishimura, Y., Gole, L., Yu, W. & Motegi, F. Cortical Forces and CDC-42 Control Clustering of PAR proteins for C. elegans Embyonic Polarization. Nat Cell Biol19, 988–995 (2017).
Dickinson, D. J., Schwager, F., Pintard, L., Gotta, M. & Goldstein, B. A Single-Cell Biochemistry Approach Reveals PAR Complex Dynamics during Cell Polarization. Developmental Cell42, 416–434.e11 (2017).
Rodriguez, J. et al. aPKC Cycles between Functionally Distinct PAR Protein Assemblies to Drive Cell Polarity. Developmental Cell42, 1–35 (2017).
Chen, S. et al. Regulation of Microtubule Stability and Organization by Mammalian Par3 in Specifying Neuronal Polarity. Developmental Cell24, 26–40 (2013).
David, D. J. V., Tishkina, A. & Harris, T. J. C. The PAR complex regulates pulsed actomyosin contractions during amnioserosa apical constriction in Drosophila. Development137, 1645–1655 (2010).
I am writing to ask you to consider nominating individuals for membership of the BSDB committee. Due to committee retirements, three new individuals are needed to serve as ordinary committee members, beginning in October 2018. Their term of office will be five years and they will be required to (make every attempt to) attend two committee meetings per year.
All nominations must be proposed by a member of the BSDB, and seconded by another member, and must also be accompanied by a statement from the nominee that they are willing to serve if elected. The nominee has to already be a member of the BSDB.
Please send your nominations to me ASAP and by Friday 2 March 2018 at the latest.
In the event of a vote being required to decide between >2 candidates, the election will take place at this year’s BSDB AGM, which will be held during the BSDB Spring Meeting 2018 in Warwick (15th – 18th April, 2017). If there is an election, I will require someone, preferably the proposer or seconder, to give a short verbal statement of support of their nominee.
The BSDB committee also invites self-nominations for a new post graduate representative and a new post doctoral representative. Please, include a short text (max. 1 page) explaining your motivation and intentions to serve on the committee. Deadline will also be Friday 2nd March 2018 .
The Turing Centre for Living Systems aims at attracting students willing to work in an interdisciplinary life-science environment and with backgrounds in cell or developmental biology, immunology, neurosciences, theoretical physics, biophysics, computer science, bioinformatics, applied mathematics, engineering.
Candidates are asked to select the projects in order of interest (up to 3) and rank them in the application form.
The DRSC/TRiP-Functional Genomics Resources is a Drosophila community resource with three main focus areas: (1) cell-based Drosophila cell screening, (2) fly stock production, and (3) bioinformatics. Our TRiP fly stock production platform is taking nominations for production of CRISPR sgRNA fly stocks. Researchers can nominate genes for knockout (TRiP CRIPSR-KO) or activation (TRiP CRISPR-OE), or both. To search existing and in-production stocks, and to nominate genes for production of new stocks, please visit the gRNA Tracker website. In keeping with our long-time policies, all fly stocks are deposited to the Bloomington Drosophila Stock Center. We look forward to continuing to serve the needs of Drosophila developmental biologists and others through this effort!
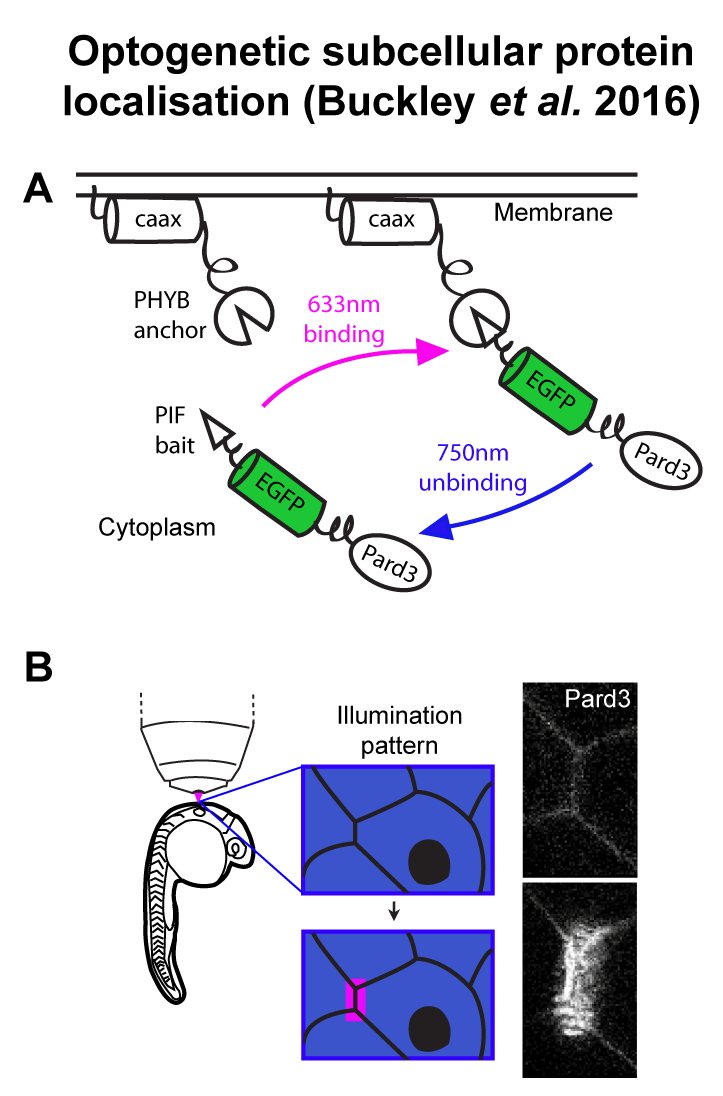
The Buckley lab at the department of Physiology, Development and Neuroscience (PDN), University of Cambridge is recruiting a postdoctoral research associate or research assistant. The lab uses cutting edge optogenetic and live confocal imaging approaches within the whole zebrafish neural tube to manipulate the polarity of single cells (Buckley et al., 2016, PMID: 26766447). In combination with CRISPR-mediated functional knock down experiments, we are directly testing the role of cell polarity in building epithelial integrity during organ development and in breaking it during developmental processes such as EMT and diseases such as carcinoma. The department of PDN is home to world-leading research in development, neuroscience, zebrafish live imaging and optogenetics. It hosts the Cambridge Advanced Imaging Centre (CAIC), which provides cutting edge microscopy systems, bespoke development of new imaging equipment and expert support.
We are seeking an enthusiastic and proactive candidate to join the team at the beginning of this exciting research. There are two main projects with which the successful candidate could be involved, depending on their interests and expertise. The first is to use optogenetics and tissue-specific CRISPR to determine how cell polarity and cell division are linked during epithelial establishment (we previously discovered a novel mechanism of cell polarisation that occurs independently to cell division: Buckley et al., PMCID: PMC3545300). We will do this within zebrafish embryos and, in partnership with our collaborators, in mammalian stem cell culture systems. The second project is to test the role of polarity dysregulation in tissue disruption. We will do this by optogenetically manipulating polarity-linked signalling pathways (such as the PI3K pathway) in the already established zebrafish neural tube epithelium. We will use 4D imaging to assess the cellular consequences of these manipulations and will model how signalling dynamics are propagated through the tissue in real time.
The successful candidate should have or be near completion of a PhD (or equivalent) in a relevant field and have a competitive history of research achievements. We are interested both in candidates with a background in developmental cell biology and those coming from a more biophysical background. Experience in molecular biology and genetics is essential and ideally the candidate should have experience in CRISPR technology. Candidates must also have a good understanding of data analysis and bioinformatics. Experience in advanced imaging and analysis would be a great advantage, as would specific knowledge of zebrafish genetics. Knowledge and interest in cell polarity and epithelial development, biochemical signalling pathways and optogenetic techniques would be desirable.
Although this is a full-time post, part-time working i.e. 80% of full-time over 4 days may be possible.
Fixed-term: The funds for this post are available for 3 years in the first instance.
 (No Ratings Yet)
(No Ratings Yet)







 (1 votes)
(1 votes)

 Emilia started her studies in 2007 at the Sapienza University of Rome and was awarded a B.Sc. in Biological Sciences with highest marks in 2010. She stayed at the same university for her Master’s project which she performed in the laboratory of
Emilia started her studies in 2007 at the Sapienza University of Rome and was awarded a B.Sc. in Biological Sciences with highest marks in 2010. She stayed at the same university for her Master’s project which she performed in the laboratory of 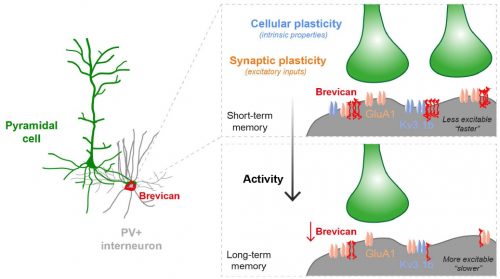
 (7 votes)
(7 votes)