A 3-year PhD fellowship in evolutionary developmental biology and genomics is available in the lab of Michalis Averof, at the Institut de Génomique Fonctionnelle de Lyon (IGFL) in France. The fellowship is funded by the Marie Curie ITN programme EvoCELL.
Some animals have the ability to regenerate parts of their body (limbs, tail, internal organs) after severe injury. However the molecular and cellular mechanisms underlying the regeneration of such complex organs are still poorly understood. The project will investigate cell differentiation during leg regeneration in the crustacean Parhyale hawaiensis, an emerging model for limb regeneration. The PhD fellow will determine the spectrum of cell types present in Parhyale limbs, their molecular profiles, and whether that diversity of cell types is fully restored after regeneration. In collaboration with other teams of the EvoCELL network, s/he will compare the transcriptional profiles of diverse cells types across phyla, in an effort to identify the evolutionary origins and conserved molecular signatures of progenitor cells.
The student will be trained in various technologies to analyze limb regeneration in Parhyale and compare it to regeneration in other species. S/he will use single-cell transcriptomics to establish the exhaustive repertoire of cell types prior to, during and after regeneration. S/he will participate in all steps of the experiment: dissociate limb cells, prepare libraries and computationally analyze the data. The fellow will also be trained in transgenesis, confocal microscopy and in situ hybridization in order to validate the identity of the detected cell populations and use markers to follow the fate of those cells during regeneration. This work will involve short periods of training/research in the laboratories of other EvoCELL partners and collaborators, and participation in yearly meetings and practical courses organized by the network, to take full advantage of the wide range of expertise available in EvoCELL.
The host lab (https://averof-lab.org) is based in the IGFL – an institute co-founded by the École Normale Supérieure de Lyon and the CNRS, whose scientific focus lies at the interface of developmental biology, functional genomics and evolution (http://igfl.ens-lyon.fr/). The lab’s working language is English. The fellow will be co-supervised by Michalis Averof and Mathilde Paris.
Applicants should have a Masters degree or equivalent (e.g. a 5-year university degree) to be eligible to embark on a PhD at the École Normale Supérieure de Lyon. To be eligible for the fellowship, candidates must be within the first four years of their research career and not hold a doctoral degree. Candidates of all nationalities may apply, but they must not have resided or carried out their main activity (work, studies, etc.) in France for more than 12 months in the 3 years immediately prior to their appointment (short stays such as holidays or compulsory national service are not taken into account).
The suggested starting date for the PhD is October 1, 2018.
Applicants should send a short letter of interest and CV to michalis.averof@ens-lyon.fr, and ask two referees to send recommendation letters to the same electronic address. The closing date for applications is February 2, 2018.
One author’s perspective on the origins of this study (Gavin Clowry)
For me the origins of this research paper stretch back nearly twenty years. I was part of a research group, led by the paediatric neurologist Professor Janet Eyre, studying the neurobiology of cerebral palsy including neurophysiological studies on infants and children. A little boy with hemiplegia generated some remarkable results. He was unusual in that he had suffered a brain lesion relatively early in development compared to most CP sufferers, around mid-gestation, resulting in destruction of the sensorimotor cortex unilaterally. Transcranial magnetic stimulation, coupled with electromyographic recording, showed that muscles contralateral to the lesion were strongly responsive to stimulation of the ipsilateral motor cortex, not unusual in hemiplegics. What was unusual, though, was the strong response of the same muscles to stimulation of the intact contralateral occipital cortex. Ten more years of experiments and imaging were required to confirm these observations [1] but it was shown that in response to the lesion, visual cortex had acquired or maintained projections to motor centres in the spinal cord (Figure 1).
Figure 1 (A) T1-weighted magnetic resonance imaging of the brain at age 4 years showing left middle cerebral artery territory infarction. (B, C) Topography of corticomuscular coherence (CMC) with the electromyogram (EMG) of the right (paretic) first dorsal interosseus (FDI) (B) at 17Hz, which is centered over O1 and (C) at 24Hz, which is significant at O1 and C4. (D) Topography of CMC with the EMG of the left FDI at 24Hz, which is centered over C4. (E, F) Transcranial magnetic stimulation (TMS) motor evoked potentials (MEPs) recorded in the EMG of the right and left FDIs when L.J. was aged 14 months (upper traces) and 48 months (lower traces) evoked when the TMS coil was placed (E) over the occipital cortex (O1) and (F) over the right motor cortex (C4). Stimulation over C4 (F) led to the expected motor responses in the contralateral (left) FDI (individual responses shown in black) and also to MEPs in the ipsilateral (right/paretic) FDI (responses shown in blue). This is a pattern previously recognized in the motor supply to the paretic hand in congenital hemiplegia. (E) Finally and uniquely, stimulation over O1 produced MEPs in the paretic (right) FDI (red lines) but no MEPs in the unaffected (left) FDI (black lines). All evoked responses were of shorter latency at 48 months than at 14 months, as expected. (G) CMC spectra recorded at O1, showing significant coherence with the EMG of the right FDI (red line) but not the left FDI (black line). (H) CMC spectra recorded at C4 showing significant coherence with the EMG of the right (blue line) and left (black line) FDI. The dashed line indicates the upper 95% confidence limits for a CMC of zero for both (G) and (H). From [1] with permission.
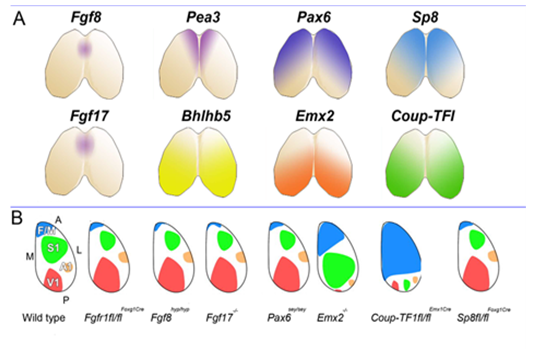
This set us speculating on the nature and plasticity of the protomap in human. Around this time strong evidence was being presented in favour of the protomap hypothesis originally proposed by Pasko Rakic [2]. The hypothesis stated that the organisation of the cerebral cortex into functional areas is determined by the co-ordinated and compartmented expression of genes in time and space at the earliest stages of its development, prior to its connection with the sensory input that could drive its maturation. Experiments in mice (Figure 2) [3, 4] were showing that altered gradients of transcription factor expression across the early cortex in mutant and transgenic mice led to an altered functional map for primary cortical areas. But does all this hold true for human development? For our next programme of research we teamed up with Prof Susan Lindsay and the Human Developmental Biology Resource to explore gene expression in the early stages of human cortex development especially to look for evidence for a human protomap.
Figure 2. A. Shows location of expression of the morphogen Fgf8 and some of its downstream effectors, all of which show high anteromedial expression (Fgf17, Pea3, Sp8) along with transcription factors expressed in an opposing gradient (Coup-TFI, Bhblb5) and Pax6 and Emx2, expressed in opposing anterolateral to posteromedial gradients. B. Summarises the effects of experiments manipulating the expression of these morphogens or transcription factors upon the size and location of primary cortical areas, usually identified and delineated in perinatal animals by expression of specific cell adhesion molecules. From [ 5 ] with permission.
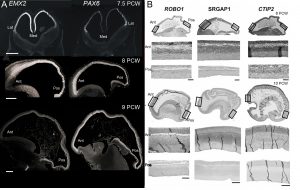
Our first success was to show that the transcription factors PAX6 and EMX2 form counter-gradients across the human cortex, just as they do in mouse, but only at the very earliest stages of cortical plate formation (Figure 3) [6]. Unlike in mouse, where expression is confined to progenitor cells, EMX2 was found to be expressed in a gradient in the post-mitotic neurons of the cortical plate as well. We also found arealised expression for other genes, by histological techniques, from quantitative measurements of mRNA expression, and in fetal cells cultured from different cortical regions [7, 8]. For instance, the layer V specific transcription factor CTIP2, along with transmembrane signalling molecules and corticospinal tract markers ROBO1 and SRGAP1, are more highly expressed in the frontal cortex between 9-12 PCW which might represent the predominance of corticofugal projection neurons in frontal sensorimotor areas.
Figure 3. (A) Expression of opposing gradients of PAX6 and EMX2 in the human forebrain between 7.5 and 9 post-conceptional weeks (PCW) by in situ hybridisation. Note that the Pax6 gradient disappears by 9 PCW. (B) Markers for corticofugal neurons, CTIP2, ROBO1 and SRGAP1 are expressed in a high anterior to low posterior gradient in the human cortex at 8-10 PCW. Ant, anterior, Pos, posterior; Med, medial; lat, lateral.
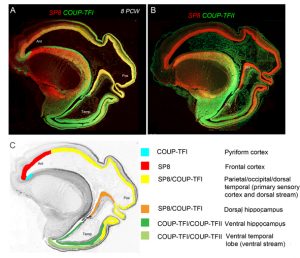
However the crucial question is whether such a developmental scheme observed in rodents is sufficient to produce all the extra areas of specialised association cortex, along with the vastly increased interconnectivity between these areas, seen in primates [9]? We believe our latest study may shed some light on this question [10]. We studied expression patterns of the arealising transcription factors COUP-TFI, COUP-TFII and SP8. What was noticeable in humans was that SP8 and COUP-TFI overlap extensively in the ventricular zone of visual, auditory and somatosensory cortex. This differs from the mouse in which COUP-TFI and SP8 show little overlap. Combinatorial expression of COUP-TFI and SP8 could maintain a common genetic identity for future primary sensory areas (visual, auditory and somatosensory) and a partially shared identity with SP8-expressing frontal motor cortex with which these sensory areas will interconnect, along with allied association cortex, via dorsal sensorimotor pathways. In mouse COUP-TFII is confined to a very small portion of the posterior cortex but in human is expressed extensively throughout the ventral temporal and ventral posterior cortex where it overlaps with COUP-TFI expression. Perhaps the expansion of cortical COUP-TFII expressing territory in human fetal brain mirrors the increased size and complexity of the association areas of the ventro-temporal cortex. An extension of this observation is that dorsal and ventral hippocampus are also differentiated by combinatorial expression of SP8/COUP-TFI and COUP-TFII/COUP-TFI respectively. Each domain has distinct functions and distinct efferent and afferent connections. It appears that the protomap for human hippocampal specialisation is laid down early in development.
Figure 4 (A) Expression of opposing gradients of SP8 and COUP-TFI in a sagittal section of human fetal telencephalon at 8 Post-conceptional weeks. (B) compartmentalised expression of SP8 and COUP-TFII in the developing cerebral cortex. (C) summarises the findings in A and B demonstrating how the progenitor zones of the cortex are subdivided into compartments by transcription factor expression that give rise to different functional areas of cortex in maturity.
It has been a 15 year journey to reach the beautiful images in Figure 4 with many interesting diversions along the way; for instance, when studying PAX6 expression we discovered a new class of progenitor cell in the subventricular zone of the cortex [11] which was subsequently shown to be a new class of radial glia [12]. We now know that humans as well as rodents have a protomap to guide cortical development, but this protomap is plastic and can be perturbed by genetic alteration, pharmacological interventions and lesions. Although more investigation is needed, it seems highly likely that the protomap has evolved in complexity as the cerebral cortex has evolved in complexity, rather than entirely new mechanisms being required to shape the cortex’s functional organisation, although, as ever, more research is needed to test this assertion. Ensuring that neurons in the cortex have the right regional and functional identity is crucial to establishing their long and short range connectivity, deficits in which are proposed to underlie neurodevelopmental orders such as autism and schizophrenia, so there is good reason to extend this journey a while longer yet.
Clowry GJ, Alzu’bi A, Harkin LF, Sarma S, Kerwin J, Lindsay S (2017) Charting the protomap of the human telencephalon. Seminars in Cell and Developmental Biology, Epub ahead of print, doi.org/10.1016/j.semcdb.2017.08.33.
Established by the British Society for Developmental Biology in 2014, The Gurdon/The Company of Biologists Summer Studentship scheme provides financial support to allow highly motivated undergraduate students an opportunity to engage in practical research during their summer vacation. Each year, ten successful applicants spend eight weeks in the research laboratories of their choices, and the feedback we receive is outstanding.
Our tenth report from the 2017 group of student awardees comes fromLiam McMulkin(student at The University of Dundee), who undertook his studentship with Dr. Marios Stavridis and Dr. David Martinat The University of Dundee.
My project’s aim was to expand British Sign Language (BSL) glossary for biology, more specifically areas relating to Developmental Biology.
British Sign Language (BSL) is a form of sign language which involves in the use of hand movements, gestures, body language and facial expressions to communicate. BSL is mainly used by deaf people. Unfortunately, BSL usually does not have signs for technical words that are not normally used in daily conversations, in biology for example, adenosine triphosphate, centriole, ectotherm, and many more which consequently lead to deaf people not having an equal access to biology compared with hearing people e.g. Interpreters don’t have signs for biological terms, which results in the use of fingerspelling. However, fingerspelling can be a lengthy process as every letter has to be spelt and is therefore not appropriate for a biology lecture, and also it is unpleasant to watch. Could you imagine a lecturer speaking out individual letter to spell out a word, more than twenty times in an hour lecture? E.g. electrophoresis and electroporation. Sometimes it can be difficult to distinguish between two words in a same lecture e.g. pluripotent, totipotent etc. especially when they are spelt out one letter at a time.
Scottish Sensory Centre has National 4 Biology BSL glossary. Unfortunately, they cannot source more funding to expand their BSL glossary for biology. Therefore, I decided to do something. I want to change this, I want deaf people to have a better access to biology with a better standard of BSL. This is a good timing to change this, as now it is an exciting time to study biology as new technologies open up novel areas of discovery e.g. genetic engineering and stem cells. Deaf people deserve to learn in their first language.
Dr. Marios Stavridis and Dr. David Martin agreed to support me to change this. They persuaded me to apply for this fantastic BSDB Gurdon Summer Studentship. After a month of waiting, I was very pleased to find out my application turned out successful. Straight away, I was given an unrestricted access to the staff in the Division of Cell and Developmental Biology at the University of Dundee, also space in the building to use as my base and meet with staff and postgraduate students to develop this glossary. In the process, I gained a first-hand experience of topics in Developmental Biology spanning the areas from Evo-Devo, live imaging of gastrulation, stem cells, neural and mesoderm development. This allowed me to get a feel for the terms beyond a dictionary definition. BSL is a very visual language and imagery is important in making good terminology. I spent a month doing this, before I ran the workshops.
The workshops were held on 8th and 10th August at University of Dundee. I sent out invitations using BSL via a video on Facebook with closed captions. The video had many shares, and I think it has shocked some people as they never knew that BSL has a limited glossary. University of Dundee also released a video about my workshops, which has reached to many people. Other news agencies such as Times Higher Education also released a news piece about my project. I am pleased that my project was well informed to the public because it raises awareness about BSL and its glossary.
Over 15 BSL users (aged from 20 and 70) attended and took part in the process of developing signs. Dr. Marios Stavridis briefly introduced what Developmental Biology is. Then, a Ph.D student provided a talk about her research work – two BSL interpreters struggled translating the talk due to lack of complex signs. Before we started developing new signs, a BSL linguistic, Gary Quinn introduced how to develop signs, and ensure they follow BSL grammar. I prepared a PowerPoint with terms we lack signs in with definitions and other helpful resources. All the participants worked together really well and developed over 70 new signs! At the end of the last workshop, the Ph.D student repeated her talk, and the participants were extremely shocked how improved the translations were from the same interpreters.
After the workshops, I spent roughly three days signing the newly developed signs. I then shared the signs online via Facebook for review. The feedbacks I received were really positive.
Now, I am at the last stage before finishing the project. The developed signs are required to be reviewed by Scottish Sensory Centre Glossary Manager, Dr. Audrey Cameron and a BSL linguistic, Gary Quinn before they are uploaded online at Scottish Sensory Centre website.
I can’t express how much I appreciated everyone’s support in this project. I millions of times thank to BSDB for selecting me to part of their studentship programme. Also, millions of times thank to The Robertson Trust for covering the costs for running the 2-day workshops at University of Dundee. I thank all the participants for all their efforts in developing signs for complex terms. Thank you to Dr. Marios Stavridis for being my supervisor. Also, for arranging an unrestricted access to the College of Life Sciences which gave me unique experience meeting world-leading scientists and observed their real work. Many thanks to Dr. David Martin for co-supervising me, and allow me to borrow his high-quality filming devices. My filming skills have improved! Finally, many thanks to Francesca Carrieri for her time observing her work and her time to come along to my workshop to deliver a talk on her research work.
In conclusion, I am very pleased I took this opportunity to improve BSL vocabulary for biology, which will improve deaf people’s access to education and science-related workplaces. Also, general science conversations using BSL. From this project, I hope more funding bodies have recognised the work of Scottish Sensory Centre and support them expanding BSL vocabulary to help deaf students in education. Finally, I really hope this project encourages Scottish Sensory Centre to add more of university-level vocabulary rather than just school-level vocabulary for help deaf higher-education students like myself.
Gaining a first-hand experience of topics in Developmental Biology at College of Life Sciences, University of Dundee. Left to Right: Anne Whittaker (communication support worker), Francesca Carrieri and myself.
Dr Marios Stavridis introducing Developmental Biology on 8th August 2017 at the first sign development workshop at University of Dundee. Challenging for interpreters. The picture was taken by Dr David Martin.
Positions are available to study the physical principles of morphogenesis in the Mechanics of Morphogenesis / Davidson Laboratory at the University of Pittsburgh in the Department of Bioengineering. Our group focuses on studying the molecular, cellular, and tissue-scale processes that regulate mechanical properties and force-production during morphogenesis. Projects can involve quantitative cell biology, biophysics, bioengineering, and embryology.
Postdoctoral candidates will have recently completed a PhD and have a background in either bioengineering, biophysics, cell and developmental biology, or cell- and tissue- mechanics. The research environment at the University of Pittsburgh includes a dynamic community of bioengineers, developmental biologists, cell- and tissue-level biomechanics, and theoretical biologists. Nearby resources include the Peterson Institute of NanoScience and Engineering and the Pittsburgh Supercomputing Center. Contemporary Pittsburgh is a diverse vibrant city undergoing a renaissance led by world class Universities and the University of Pittsburgh Medical Center. The University of Pittsburgh is an Equal Opportunity Employer. Women and minorities are especially encouraged to apply.
Interested applicants should forward their CV and statement of research interests to:
Lance Davidson (lad43@pitt.edu)
Professor of Bioengineering
University of Pittsburgh
mechmorpho.org
The Marine Biological Laboratory seeks a Research Assistant II or III to join the laboratories of Kristin Gribble and David Mark Welch in the Josephine Bay Paul Center. The successful candidate will help develop genome editing techniques, including CRISPR/Cas9, in rotifers, a novel aquatic invertebrate model system for studies of aging, neurobiology, developmental biology, ecology, and evolution. Specific goals of the project include designing guide RNAs, optimizing microinjection methodologies, phenotyping and genotyping mutant strains, and screening genes of interest.
Qualifications:
Applicants should have a B.A., B.S., or Master’s degree in biology, cell/molecular biology, biochemistry, or a related field. This position requires proficiency and previous experience in basic molecular biology techniques, microscopy, microinjection, and CRISPR/Cas9 methodology. We are seeking an independent, organized, enthusiastic, and productive individual with robust problem solving skills. Excellent interpersonal skills, attention to detail, and a strong work ethic are essential. Position level and salary will depend upon education and experience. The ideal candidate will have working familiarity with RNAi and transgenic protocols. Proficiency in bioinformatics is a plus. Previous experience in established animal model or in non-model systems is preferred.
Established by the British Society for Developmental Biology in 2014, The Gurdon/The Company of Biologists Summer Studentship scheme provides financial support to allow highly motivated undergraduate students an opportunity to engage in practical research during their summer vacation. Each year, ten successful applicants spend eight weeks in the research laboratories of their choices, and the feedback we receive is outstanding.
Our ninth report from the 2017 group of student awardees comes fromMiguel Robles Garcia(student at The University of East Anglia), who undertook his studentship with Andrea Münsterbergat The University of East Anglia.
This summer I had the opportunity to undertake an internship at Andrea Münsterberg’s Laboratory at the University of East Anglia, where I am currently studying for a Bachelor’s Degree in Biological Sciences. Under the supervision of a PhD student, Johannes Wittig, and a postdoctoral researcher, Dr Estefanía Lozano-Velasco, I was able to spend seven weeks learning the ins and outs of everyday research. During this internship, my role was to focus on the early stages of heart development in chicks.
During vertebrate development, the heart is one of the first organs to develop. It is known that during this process many malformations can occur, which are capable of affecting correct heart function leading to potential defects or death. The developmental stages between mammals and birds are similar starting with cardiac looping and resulting in chamber formation. Due to the importance of the heart, its development is tightly regulated by different transcriptional and post transcriptional signalling pathways driven by transcription factors and microRNAs (miRs) respectively. miRs are regulators of gene expression that inhibit the translation of mRNA. During this project, my focus was to aid the postdoctoral researcher with her investigation of miR-133 function, which is thought to regulate BAF60b chromatin during heart development.
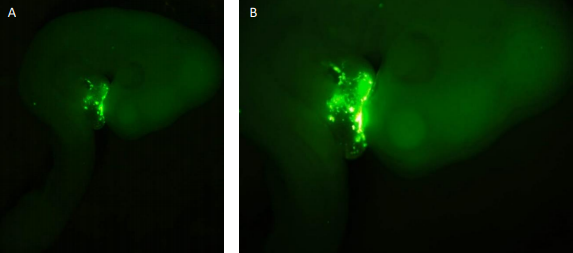
I started my time in the lab by dissecting wild type (WT) embryos from stages HH18-19 and HH23-25. The staging of the chick development follows the one described by Hamburger and Hamilton from 1951. Throughout my time in the laboratory, I became familiar with the dissection technique in which the different membranes that cover the embryo need to be removed. This is needed so that in future analyses (such as those that are mentioned) the membranes would not infer in the procedure and to make the tissue more available. My project then shifted towards the injection of embryos at stages HH 14-16 with the antagomir-133 (AM133). An antagomir is a modified oligonucleotide that binds to its specific micro RNA and inhibits its action. Therefore, this technique can be used to investigate potential effects in hearts which are deprived of a specific micro RNA. I used AM133 which is designed to inhibit miR-133. I also used scrambeled antagomiR (SCR) as a control for the procedure to see that the actual way of injecting the oligonucleotide wasn’t the one affecting the development of the heart. Both of these oligonucleotides have 5’ fluorescent label that allows the injection to be seen at first with the naked eye, but later after further incubation, with the aid of a GFP microscope (Figure 1). These antagomiRs were injected into a hollow cavity close to the heart walls while the heart is still beating without killing the embryo. After
the injection, the embryos were sealed with tape and returned to an incubator at 37°C for 24/48h depending on the intended analysis.
Figure 1. Fluorescence seen in embryos injected with the SCR probe. As depicted the fluorescence is seen only in certain parts of the heart and nowhere else. This shows that only in these places the inhibitor has bound to its target. Figure A shows the whole embryo while B is a close up of the same embryo. The fluorescence signal is stronger on the right-hand side of the heart.
While some of the embryos were used for imaging and making sure the inhibitor bound in the correct place as shown in Figure 1, other embryos were embedded for sectioning. This involved placing the embryos in a gelatine medium that would then solidify at room temperature. Two different approaches were used for this in which one meant leaving the embedded embryos at room temperature for the gelatine to solidify slowly while the other was performed using dry ice for a rapid solidification. This allowed me to section the embryos for a closer examination of the heart. SCR embryos were compared to AM133 injected embryos to see if there is any difference in morphology. However, in order to image the slides containing the embryo section I had to stain them. For this we used an Alcian blue staining procedure (Figure 2). In this case the medium surrounding the tissues of the embryos would be discarded and only the glycosaminoglycans within the actual tissue were stained blue. This was also performed for embryos that had a double knock out of miR133 and miR1.
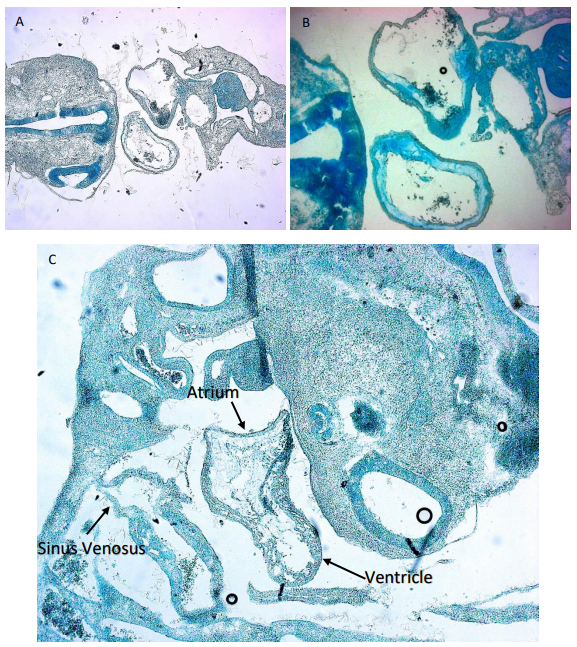
Figure 2. Image shows Alcian Blue staining of cryosections of embryos injected with antagomir-133 and SCR. A) Cross section of SCR injected embryo. The two hollow forms in the middle are the atria (down) and Truncus arteriosus (up) which can be seen connecting to the sinus venosus. B) Cross section of another SCR embryo showing a closer look at the heart. C) shows a sagital section. The top structure of the image is the head while the structure along the bottom is the back and tail of the embryo. The heart can be seen again in the middle of the embryo. Two different chambers are observable the top right one being the atria and ventricle while the bottom left one is the sinus venosus.
I want to thank the BSDB Gurdon Studentship for granting me the opportunity to have what I can just describe as an incredible experience. It has allowed me to develop my skills and scientific mind which I will be able to apply in my future studies and career. This opportunity is of great importance as, as a third-year student, I wasn’t sure of how daily lab teamwork alongside other scientists that are dedicated to their research felt like. I would also like to thank the Münsterberg lab for hosting me and Estefanía and Johannes for their guidance through these weeks.
A Brain Research UK-sponsored research assistant position for a highly motivated scientist is available in the laboratory of Dr. Claudia Barros at the Peninsula School of Medicine of Plymouth University, UK. The project is an opportunity to characterise novel targets and its human orthologue genes potentially involved in brain tumour initiation and growth, which have been identified in our laboratory. Molecular biology, cellular and biochemical techniques will be employed, including gene loss and gain of function assays, FACS, RT-qPCR, immunochemistry, western-blotting and confocal imaging. Work will make use of human brain tumour cell lines, tumour tissues and Drosophila brain as a model. A relevant 1st class or 2:1 (or equivalent) Bachelor degree is required and a postgraduate research degree may be preferred. You must have experience in some of the mentioned techniques, in particular cell culture and tissue work, and a suitable background knowledge. Good analytical, organisational and presentation skills, ability to multitask and collaborate with team members, and a genuine dedication to the research work are essential. Please include a cover letter detailing suitability/ experience/ interest and an academic CV with your application, in addition to ensure that 1-2 academic references are received.
Full or part time; Fixed term: initially 7 months if full time; Salary start: £24,983/ year.
Established by the British Society for Developmental Biology in 2014, The Gurdon/The Company of Biologists Summer Studentship scheme provides financial support to allow highly motivated undergraduate students an opportunity to engage in practical research during their summer vacation. Each year, ten successful applicants spend eight weeks in the research laboratories of their choices, and the feedback we receive is outstanding.
Our eighth report from the 2017 group of student awardees comes from Nicole Serzhantova(student at The University of Edinburgh), who undertook her studentship withJennifer Nicholsat The University of Cambridge.
Understanding the first step in the formation of organismal complexity in the mouse embryo
As biologists, we are very used to poking and prodding very complex systems, be it drosophila or homo sapiens. Rarely is it a prerogative to take a step back and really understand how such complexity arises seemingly out of nowhere. How is that we start off from a single cell and over a period as short as 48 hours (in the mouse at least) we already start seeing complexity arising?
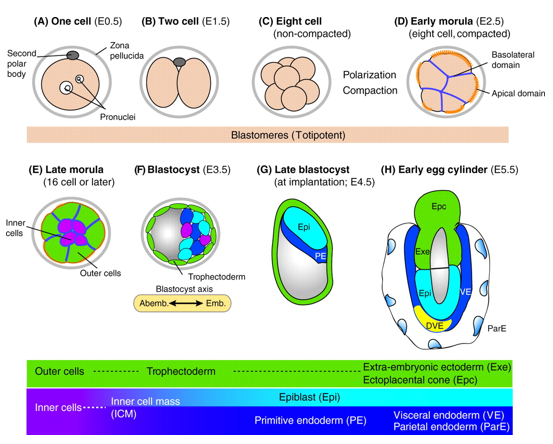
In mice, a single-cell zygote undergoes a series of cleavage cell divisions and morphogenetic changes to form a unipotent epithelial vesicle termed the trophectoderm (TE) enclosing a compact group of pluripotent stem cells known as the inner cell mass (ICM) (Fig.1F).
Figure 1. Development of the mouse embryo from one cell pre-implantation stage to a post-implantation stage embryo. (Rossant & Patrick, Development, 2009)
Restriction of cell fate takes place over an extended time period of 48 hours from the 8-cell (morula) stage (Fig.1C), when all cells are indistinguishable from one another and are capable of forming all three germ layers. This feature of cells is termed totipotency and is a transient characteristic of embryonic stem cells before they commit to become either TE or ICM.
At this 8-cell stage compaction of the embryo begins, in which the embryo surface smoothens because of an increase in intercellular adhesion. Cells epithelialise, forming tight junctions (which acts as a seal) between cells, limiting diffusion and what you are left with is fluid-filled epithelial vesicle enclosing a small number of cells (Fig. 1D-F).
It is at point that the first commitment of cells occurs, whereby the outer epithelium (Fig.1E) becomes committed to form the trophectoderm (TE), which subsequently gives rise to the trophoblast layers of the placenta and the trophoblast giant cells. Whilst the small population of enclosed cells commit to becoming the ICM, which is the pluripotent stem cell lineage of the embryo, giving rise to all of the primary germ layers of the foetus and its extraembryonic tissue.
At this stage of development, protein synthesis from maternal mRNA becomes more transient as the embryo’s own gene programs begin to switch on. Once commitment occurs, as the morulae becomes a blastocyst, differential gene expression is seen between the ICM and TE. At day 2 ½ post-fertilisation, all cells within the morula uniformly express most genes, including the pluripotency factor Oct4 and the TE marker Cdx2. Interestingly, at day 3 ½ the first real differential gene expression is observed, where the ICM begins to express Oct4 whilst loosing its ability to express Cdx2 and conversely the trophectoderm begins to express Cdx2 whilst loosing all Oct4 activity. There is a mutual repression by Oct4 and Cdx2 which further consolidates the TE and ICM segregation.
Elucidating the mechanism governing a cell’s ability to become either a unipotent TE cell or a pluripotent stem cell of the ICM can open up new avenues for generating and maintaining pluripotency in vitro. In an attempt to understand this initial lineage segregation, a stem cell line was cultured which removes the Oct4 gene upon treatment with tamoxifen. This cell line also had a florescent tdTomato reporter under the control of a constitutive promoter, meaning that these cells fluoresce red when exposed to light in the blue to green range.
The following cells were cultured in serum-LIF (a differentiation permissive medium) and different portions of the cells were treated with tamoxifen for either 24, 48 or 92 hours to allow for the excision of the Oct4 gene. Once these time points of treatment were reached, cells were microinjected into live mouse morulae and blastocysts. Approximately 5 morulae and blastocysts were microinjected with 8-10 stem cells deficient in Oct4 from each time point as well as un-induced cells form the same batch as a control. These morulae were left for two days to allow for compaction and the first lineage segregation to occur. These were then promptly fixed and immunostained to examine what effect the deletion of Oct4 had on the cell fate choices that the injected stem cells made.
Following what we already know about what happens during development, it can be hypothesized that stem cells that do not have the capacity to make Oct4 will commit to the trophectoderm lineage as they are missing a key pluripotency factor and therefore cannot become pluripotent cells competent of forming all 3 germ layers.
Unfortunately due to rising temperatures in the laboratory during summer (the air conditioning was of course broken on the days the microinjections were performed), upon imaging it was clear that in most cases only 1 stem cell, out of the 8-10 injected, integrated into the embryo. This was prevalent in the un-induced cells also, suggesting that it was not the tamoxifen treatment that caused toxicity to the cells but rather the experimental conditions. Integration rates were higher in morulae than in blastocysts as expected, since by the blastocyst stage cell commitment has already occurred making it harder for integration to ensue.
Out of the cells that did integrate, only 1 appeared to express Cdx2 markers showing that it has adopted a trophectodermal cell fate, whilst the others exhibited a mixture of pluripotency markers. The conclusion from this being that although Oct4 is necessary for cell commitment to the ICM lineage, additional molecular events must underpin this initial commitment in the mouse model. Inferring this from the data is difficult considering how the number of integrated stem cells was so low and future experiments with more tightly controlled conditions are advised.
These results however are concordant with other studies, with Wu and Schöler (2014) reporting that the “establishment of totipotency in maternal Oct4–depleted embryos was not affected, and that these embryos could complete full-term development without any obvious defect.” In addition to this, Wu and Schöler were able to form Oct4 expressing inner cell masses in embryos with complete inactivation of both maternal and zygotic Oct4 expression as well as reprogramming of fibroblasts into fully pluripotent cells by Oct4-deficient oocytes.
This, in conjunction with the results I obtained, tends to indicate that Oct4 is not essential for the initiation of pluripotency but in contrast to its role is critical for the maintenance of pluripotency.
I would like to thank Professor Jennifer Nichols, Ayaka Yanagida, Peter Baillie-Johnson, Thorsten Boroviak, Ken Jones, Tim Lohoff and everyone else at the Cambridge Stem Cell Institute for their invaluable guidance and support throughout my internship. I would also like to thank the British Society for Developmental Biology for granting me the Gurdon Studentship Award without which none of this would have been possible.
Built into the side of a tree covered hill on a remote Japanese island lies what at first sight appears to be a super-villain’s lair, or perhaps the HQ of Jurassic Park. In fact we had arrived at the Okinawa Institute of Science and Technology (OIST) for their 2 week Developmental Neurobiology Course. DNC is a 2 week intensive lecture and lab based course designed to teach, inspire and foster collaborations between the next generation of developmental neurobiologists.
OIST (Photo credit OIST website)
23 of us were accepted, a highly diverse collection of PhD students and Postdocs from all over the world to spend two weeks in paradise all there because of our love for developmental neurobiology. The course was split into two halves: Week one – lectures in the morning and a lab project in the afternoon. Week 2 – lecture bootcamp! 6-7 hours of fascinating lectures a day by some of the most distinguished researchers in the field on a vast variety of topics.
While the classes were intense there was plenty of downtime to interact, consolidate and simply think about the wider field beyond our individual specialities, with plenty of swimming off the beach attached to our accommodation! In this post I will try to summarise the course and hopefully explain why I think courses like this are so crucial for early career researchers to participate in.
Week 1:
After 27 hours of travelling I finally arrived at our Seaside House accommodation. OIST generously covered all course costs, accommodation, food and half of travel costs for all participants; a critical factor in making it accessible to early career researchers.
We started our first day with intensive short talks from all participants so we all knew a little bit about each other’s background. The range of specialities was huge, from pure molecular biologists to electrophysiologists, to using Zebrafish to study dyslexia, making for some fascinating discussions throughout the 2 week course. We got the chance to learn more about each other’s research the following day, where after a fascinating lecture on microscopy by Ulrike Engel, we gathered for a poster session to present our research to staff and participants.
A key part of DNC is the opportunity to learn new techniques that we might not otherwise get the opportunity to learn. This was taught through a choice of one of five lab projects, but before committing we got a brief rotation in each lab to learn more. The five lab project options available to us were:
Analysis of Drosophila embryonic neuromuscular development
Ca imaging and electrophysiology recording from Drosophila Larvae
In utero electroporation in mouse
Live imaging of zebrafish nervous system
Electrophysiology in zebra finch auditory system
I was lucky enough to be taught in utero electroporatation in mouse by Hiroshi Kawasaki, a powerful technique for highly spatially specific gene editing during development.
Having been assigned our lab projects the week continued with lectures in the morning and lab projects in the afternoon. Our lectures were fascinating and varied:
Cecilia Moens (Fred Hutchinson) spoke about cranial motor neuron development in the Zebrafish.
Lisa Goodrich (Harvard) explained about the peripheral nervous system, focusing on cochlear development.
Hiroshi Kawasaki (Kanazawa University) told us about his research focused on the molecular mechanisms controlling cortical development.
Michael Granato (U Penn) talked about axon guidance and regeneration in Zebrafish.
For me the length (3 hours) of the lectures was key to the immersion of the course. This time period gave the lecturers the chance to speak about their long research journey and all the side stories that would normally be left out of a shorter lecture. Although intense, every hour we were rewarded (behavioural conditioning?) with coffee and snacks, we had the opportunity to ask questions at any point, and the lecturers were able to enter into group discussions without fear of time constraints.
Embryos from the Zebrafish lab project
In the afternoons we utilised the fantastic laboratories at OIST to work on our lab projects with the help of our group leaders and the fantastic Postdoc tutors. Under their careful tuition we practised and perfected our techniques, and gathered our data to present at the end of the course. The resources available at OIST were staggering, we were able to perform CLARITY on our electroporated brains and image them with a light-sheet microscope, while the Drosophila group used super-resolution to image individual boutons in the Drosophila larvae!
This slideshow requires JavaScript.
Lab projects complete, we celebrated the end of the first week with a barbecue and fireworks at Seaside House, and of course Japanese karaoke with the course organisers!!
The in utero electroporation team with our instructor Hiroshi Kawasaki
Week 2: Lecture Bootcamp!!
Before the intensive days of lectures started, we were given a day off to recuperate and explore Okinawa. For a couple of us that meant a relaxing trip to an Onsen, for others Scuba-Diving, and for one adventurous group it involved a multi-stop tour to a pineapple farm, a Dragonball Z-themed butterfly park (…?) and the Okinawa aquarium.
The Dragonball-Z butterfly park
Re-energised after our free day we threw ourselves into the second week of lectures, learning about ideas and concepts that we had never encountered in our day to day research:
Yimin Zou(UCSD) spoke about how we must understand the processes controlling axon guidance during development, to investigate how circuits may repair themselves following injury.
Jeff Wickens (OIST) gave a lecture on the neural mechanisms that underlie learning and adaptation in the mammalian brain.
Kang Shen (Stanford) talked about the molecular mechanisms behind dendrite patterning and how they wire into neural circuits.
Tomoyuki Takahashi (OIST) we were privileged to be given a lecture by one of the founders of electrophysiology, a fact made clear by the massive increase in audience size as many OIST researchers came to hear him speak.
Mei Zhen ( University of Toronto) described her research on understanding how the C.elegans neural motor system translates into movement.
Greg Basaw (U Penn) gave as another axon guidance lecture, this time from the perspective of receptor regulation during Drosophila development.
Anne Hart (Brown) gave one of my favourite lectures as she described using a wide range of models from C.elegans to mice to study disease mechanisms in a highly interactive manner calling on us to think of research ideas.
That Friday afternoon we gave our brains a chance to consolidate as we were taken on an excursion to Shurijo Castle to experience more of the island’s rich history before enjoying a traditional Okinawa dinner with the course organisers and lecturers in the capital Naha.
The next day we received news, after several days of speculation, that there was a typhoon passing between the Okinawa prefecture and mainland Japan. OIST is built for this weather so we continued on with the course albeit with an embargo on all water activities.
Hitoshi Okamoto (Riken) gave a fascinating lecture on social conflict in Zebrafish and the role of the habenula in this. To me this was particularly interesting as I had never considered this behavioural use for the Zebrafish before.
Tadashi Isa (NIPS) gave our penultimate lecture on how networks reorganise to recover movements following spinal cord injury.
Yoko Yazaki-Sugiyama (OIST) delivered our final lecture on critical periods in Zebra Finch auditory systems during song learning.
As lectures concluded on the final day we concluded the academic side of the course by giving presentations on our lab projects from the first week. This gave us an excellent opportunity to show the skills we had developed, consolidate our knowledge and thank our teachers and tutors for their guidance and support.
We gathered for a final dinner together in Seaside house with traditional Okinawan entertainment as we ate. After dinner we had final words from the course organisers who presented us with certificates of our participation. This also gave us time to thank them extensively for all their work organising the course and providing us with the most incredible learning opportunity that will shape our careers as neurobiologists. We ended the course as a team, with fireworks and karaoke as the lightening storm from the typhoon raged around us.
DNC 2017 team photo
This course is such a fantastic opportunity for young researchers. The vast diversity of lectures will broaden your horizons and bring new directions to your project and future plans. The people I met will be my friends and colleagues for the rest of my career and I hope one day I will get to publish alongside them as we collaborate together. I cannot thank the course organisers; Yoko Yazaki-Sugiyama, Ichiro Masai, David Van Vactor, Yimin Zou and Hiroshi Kohsaka enough for this opportunity as well as all the support staff who made it happen. I encourage all young developmental neurobiologists to apply to this wonderful course, you will not regret it!
Established by the British Society for Developmental Biology in 2014, The Gurdon/The Company of Biologists Summer Studentship scheme provides financial support to allow highly motivated undergraduate students an opportunity to engage in practical research during their summer vacation. Each year, ten successful applicants spend eight weeks in the research laboratories of their choices, and the feedback we receive is outstanding.
Our seventh report from the 2017 group of student awardees comes from Agata Czap (student at University College London), who undertook her studentship withPaola Oliveri at University College London.
How to knockout a gene in sea urchin?
I am Agata Czap, an undergraduate student of MSci Human Genetics at UCL. The BDSB Gurdon Summer Studentship has given me an incredible opportunity to undertake research for 8 weeks at Oliveri lab in UCL. I applied to work with Dr. Oliveri to learn about the challenges of applying the CRISPRcas9 technology to new model systems.
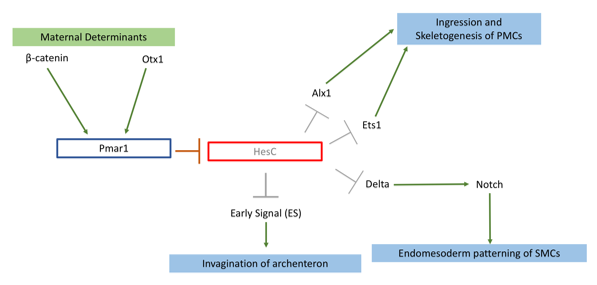
My project focused on knocking out the transcription factor Pmar1 in sea urchin S. purpuratus using the CRISPRcas9 system. Pmar1 is a part of a characterized gene regulatory network (GRN) and its role has been dissected using ectopic and dominant negative approaches (1). Specifically, pmar1 represses HesC during early development, as part of a double negative gate (DNG) of skeletogenesis. HesC is another transcription factor, which represses the downstream skeletogenesis genes (2). This DNG enables specification of micromere cell fate in the correct cells (1), from which primary mesenchyme cells (PMCs) will arise and eventually form the larval skeleton. Furthermore, the DNG enables micromeres to release two signals. Early Signal (3), which is responsible for inducing archenteron invagination, and Delta, which activates the Notch receptor in secondary mesenchyme cells for endomesoderm patterning (4) (figure 1).
Figure 1: Pmar1 is a part of double negative gate (DNG) in the skeletogenesis pathway. This gene requires β-catenin, and another transcription factor Otx, both maternally expressed. These two determinants bind to promoter of Pmar1 for activation of its expression. Pmar1 represses transcription factor HesC in micromeres. This repression enables expression of skeletogenesis genes and release of two signalling proteins to induce endomesoderm patterning in secondary mesenchyme cells (SMCs) and archenteron invagination.
So far, there have been no functional knockouts of Pmar1, as the gene has duplicated 4 times in the S. purpuratus genome. Previous experiments deduced possible function with a series of downregulation and rescue indirect strategies (1,4). Micromeres are located on the vegetal plate of the urchin embryo. One of early experiments carried by Dr. Oliveri, involved removing and transplanting micromeres into animal pole of another healthy embryo (2). This has induced a secondary, ectopic gut (2). Additionally, an embryo from which micromeres have been removed had shown no development of skeleton nor gut (3). Pmar1 expression requires maternal determinant called β-catenin, and imparing its nuclerization results in an embryo similar to micromereless embryo (3). Thus, I have hypothesized that upon successful knockout, the embryo would lack skeleton as well as other endomesoderm derivatives (e.g archenteron). Other ectodermal tissues should be unaffected.
To tackle the issue of pmar1 duplication, I have collected all 4 (Pmar1a-d) FASTA sequences from the Echinobase transcriptome. I have used Jalview to identify conserved regions between duplicates within exon 1 and 2, and generated phylogram to visualize diversification of the sequences. My results agreed with that of Cavaleri’s team (4); Pmar1b has diverged to become a monophyletic group. The sequences are still highly conserved, the first 810 bp of each transcript show 86% similarity. I used this fragment of the Pmar1b sequence, as it had the best consensus when compared to cDNA. I have inserted this sequence into 4 algorithms including CHOPCHOP to generate gRNAs. Unfortunately, the algorithms used different criteria to rate gRNAs. To filter through all 98 gRNAs, I tested each against BLAST in Echinobase for off-targets. This allowed me to obtain top 5 gRNAs, which target every Pmar1 gene.
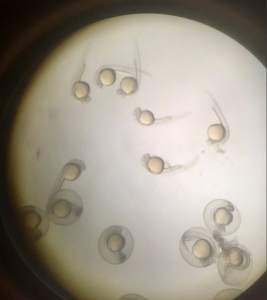
A more wet lab-based part of my project then followed. During the first two weeks, I have learned how to culture sea urchin embryos, spawn adults to obtain gametes and assess fertilisation rate. S. purpuratus has turned out to be a rewarding model system, embryos can be produced in large quantities and are transparent, allowing clear observation of tissues. Natalie, a PhD student who also worked on applying CRISPRCas9 technology to knockout neuropeptides, has taught me how to synthesize selected gRNAs using PCR and T7 in vitro translation. Once gRNAs were ready, we injected a pair of gRNAs and Cas9 mRNA into oocyte after fertilisation to induce a deletion mutation. The efficiency of knockout was tested with DNA extraction and agarose gels, which measured molecular weight of the target region. I carried out in situ hybridisation and antibody staining on genes acting downstream of Pmar1 to characterise mutants.
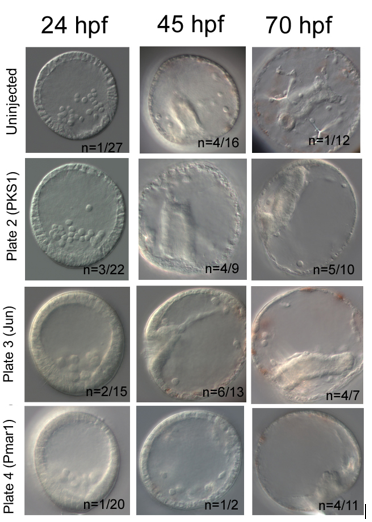
My results were promising. In each knockout, I have used PKS1 knockout in pigment cells as positive control, which proved Cas9 enzyme is non-toxic for embryos. PKS1 allows quick visual check that Cas9 works, as albino mutants are easily identified. Furthermore, it has high knockout efficiency of ~90% (5). My knockout attempt had lower PKS1 efficiency with 50% albino incidence at 45hpf. I have used additional positive control, knockout of Jun, a gene downstream of Pmar1, which caused delayed invagination in 42% at 45hpf. Pmar1 gRNA 1 and 5 have been very efficient; at both 45hpf and 70hpf, 100% of embryos were mutants. The embryos did have a blastopore and syncytial rods were present, but the gut nor skeleton were not developed (figure 2).
Figure 2: My attempt to knockout Pmar1. Jun and PKS1 knockouts act as positive controls. PKS1 knockout generated albino mutants. Jun knockout has resulted in delayed invagination characterised by undeveloped shorter gut. Pmar1 knockout caused lack of proper skeleton and gut, and round morphology of the embryos.
In conclusion, a successful knockout requires a lot of preparation, resilience and knowledge of molecular tools. The procedure gives you greater understanding of how Cas9 enzyme works and of gene function. My gRNAs will be further used by Oliveri lab to test efficiency of Pmar1 knockouts using different combinations of gRNA pairs and Cas9 concentrations.
I would like to thank Dr. Oliveri for giving me a chance to work in her lab. Also to PhD student Natalie Wood for being patient when teaching me all the techniques used. I greatly enjoyed being part of a research team and wish to pursue scientific career further. I would recommend Gurdon studentship to any student applying for a laboratory-based internship.
 (No Ratings Yet)
(No Ratings Yet)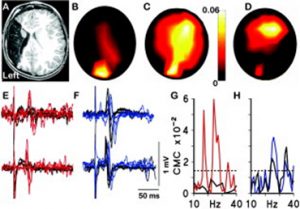
 (2 votes)
(2 votes)


 (3 votes)
(3 votes)