The Department of Neurology at the University of California, Irvine anticipates an opening for an Assistant Project Scientist in the Translational Laboratory and Biorepository (TLaB). This position requires solid background in experimental design and fluency in the use of technology germane to investigations of exosomes, including but not limited to, in vitro and in vivo methods for investigating extracellular vesicles, nanoparticle analysis, fluorescent sorting methods, immunoprecipitation, ultracentrifugation, and electron microscopy. Research efforts will involve quantitative analyses of exosomal cargo proteins and nucleic acids that are relevant in neurological disorders. The incumbent will be exposed to a wide variety of ongoing research studies within the TLaB related to neurodegenerative diseases, traumatic brain injury, and autism.
Requirements:
Candidates must hold a doctoral degree or equivalent in Neurobiological Sciences, Biochemistry, Bioengineering, Molecular or Cell Biology, or Protein and/or Nucleic Acid Chemistry and have a strong research background. Significant and creative contributions to a research or creative project in the field of Neurology and command of the English language (spoken and written) are expected. Preference will be given to candidates who hold a strong publication record and have post-doctoral experience and grant writing experience and prior funding.
Additional Information:
Positions are dependent upon extramural funding. Rank will be determined based on qualifications and experience. Initial appointments are for one year and renewal is based on continued availability of support. Salary will be commensurate with qualifications and experience.
Culture:
The UCI TLaB was established in 2015, with the research faculty and senior staff transferring from the Biomarker Laboratory and Biorepository at Georgetown University and the University of Rochester. Long-standing national and international collaborations exist on the primary research topics and provide opportunity for professional growth and job satisfaction. Current faculty and staff collaborate on a variety of human blood-based biomarker investigations, as well as in vitro and in vivo models related neurological disorders with a high potential for human translation.
The TLaB members are also involved in teaching and mentoring of medical students, graduate and undergraduate students.
Substantive inquiries about the position should be directed to:
Massimo S. Fiandaca, MD
Associate Professor
Department of Neurology
Co-Director, Translational Laboratory and Biorepository mfiandac@uci.edu
Office phone – 949-824-5579
Applicants should complete an online application profile and upload the following application materials electronically to be considered for the position:
Cover letter—Please discuss current research and future plans.
Curriculum vitae
Names and Contact Information of Three References
Diversity Statement
The University of California, Irvine is an Equal Opportunity/Affirmative Action Employer advancing inclusive excellence. All qualified applicants will receive consideration for employment without regard to race, color, religion, sex, sexual orientation, gender identity, national origin, disability, age, protected veteran status, or other protected categories covered by the UC nondiscrimination policy.
A postdoctoral fellow position is available in the laboratory of Dr. Paul Burridge in the Department of Pharmacology and the Center for Pharmacogenomics at Northwestern University Feinberg School of Medicine, Chicago, IL.
Successful candidates will participate in NIH- andAHA-funded projects to study the application of human induced pluripotent stem cells (hiPSCs) in predictive medicine. Our goal is to develop the next generation of tools for predicting drug responses and validating SNPs to allow the use of genomic information in precision medicine and drug discovery. With these data, we will probe the mechanisms of action of a range of drugs to provide individualized treatment selections and regimens to improve drug efficacy and eliminate off-target toxicity.
We are looking for highly motivated and creative candidates with an interest in studying SNPs and molecular mechanisms involved in chemotherapy-induced toxicity (primarily cardiovascular) of tyrosine kinase inhibitors and monoclonal antibodies using patient-specific hiPSC-derived cells. Projects will utilize a wide range of state-of-the-art techniques such as genome editing, high-content imaging, high-throughput drug screening, electrophysiology, whole genome sequencing,RNA-seq, and eQTL. The Burridge Laboratory is stably supported by NIH,AHA, and institutional funding. More about the lab can be found here:http://burridgelab.com/
Qualifications: PhD or MD/PhD degree (either about to graduate or graduated within the last year) and a strong record of peer-reviewed publications including first author publications are essential. Expertise in several of the following areas is required: mechanisms of chemotherapy agents and toxicity, disease modeling, pharmacogenomics,WGS,RNA-seq, eQTL,GWAS, bioinformatics,CRISPR-based genome editing, high-throughput biology and drug screening, electrophysiology, hiPSC derivation, culture, and differentiation (cardiac/vascular smooth muscle/endothelial/blood/hepatic/renal/neural), direct reprogramming, engraftment, and developmental/cardiovascular biology. Good verbal and written communication skills in English are essential. The successful candidate will join a dynamic research environment in the Department of Pharmacology, which offers both basic science and clinical translational opportunities to explore fundamental questions in pharmacogenomics.
Salary will be per theNIH(NRSA) Scale and commensurate with experience.
This position is highly suitable for those interested in pursuing a career as an independent academic scientist. Those without experience in the above fields, those more than one year after graduating their PhD, and those who wish to pursue a career in the pharma/biotechnology industry are asked not to apply.
Please send a CV, a cover letter containing a brief description of research experience and interests, and a list of 2-3 references to:paul.burridge@northwestern.edu
Northwestern University is an Equal Opportunity, Affirmative Action Employer of all protected classes, including veterans and individuals with disabilities. Women and minorities are encouraged to apply. Hiring is contingent upon eligibility to work in the United States.
I am delighted to announce that we are offering an opportunity for a 3-month internship on the Node. This is being offered as a PIPS placement to students on the BBSRC DTP program, or on similar programs where an internship forms part of the PhD training.
If you have a passion for science communication and writing, as well as a love of developmental biology, this could be the perfect internship for you! We will provide a great insight into what it’s like to work in the online scicomm environment, giving you the opportunity to come up with ideas for Node posts, talk to potential authors about writing for us, help Node users with their posts, and run the Node’s social media accounts. Working in a publishing company, you’ll also learn about how science publishing works from the inside.
You can find out more here, or please get in touch with me, Katherine Brown (Development’s Executive Editor) if you want to know more.
The Company of Biologists and its journal Development are looking for an intern, through the BBSRC DTP/PIPS or equivalent schemes, to help run the successful community website ‘the Node’ . This is a great opportunity to gain experience in the rapidly growing online science communication environment, to develop writing skills, and to learn about academic publishing.
Launched in 2010, the Node is the place for the developmental biology community to share news, discuss issues relevant to the field and read about the latest research and events. The intern will be involved in the day-to-day running of the Node, mentored by the Node’s Community Manager. The internship will be based in our office in Cambridge.
Core responsibilities of the position include:
Creating and commissioning content for the Node, including writing posts and soliciting content from the academic community, societies and other organisations
Providing user support
Running Development’s and the Node’s social media accounts (Twitter and Facebook)
The successful intern will have:
Relevant scientific expertise (ideally in developmental biology or a related field)
Strong writing and communication skills
Keen interest in science communication
Experience of and interest in blogging and/or social media (ideally including experience with WordPress)
The Company of Biologists (http://www.biologists.com) exists to support biologists and inspire advances in biology. At the heart of what we do are our five specialist journals – Development, Journal of Cell Science, Journal of Experimental Biology, Disease Models & Mechanisms and Biology Open – two of them fully open access. All are edited by expert researchers in the field, and all articles are subjected to rigorous peer review. We take great pride in the experience of our editorial team and the quality of the work we publish. We believe that the profits from publishing the hard work of biologists should support scientific discovery and help develop future scientists. Our grants help support societies, meetings and individuals. Our workshops and meetings give the opportunity to network and collaborate.
We are looking for an intern to start in Autumn 2017, though can be somewhat flexible with start dates and encourage interested candidates to submit their application as soon as possible. To apply, please send a CV and cover letter, stating why you are interested in this opportunity, to recruitment@biologists.com and Katherine Brown (Development’s Executive Editor) at katherine.brown@biologists.com. Please also direct informal enquiries to the same addresses.
Introduction
Graphs (or charts or plots) are often used for the display and summary of data. They are essential tools for the communication of results in presentations or manuscripts. One particular type of graph, the bar graph, is often used to quantitatively compare (multiple) conditions. The earliest known example of a bar graph, dates from the 18th century and its invention is attributed to William Playfair (Beniger and Robyn, 1978).
Proper use of bar graphs includes the display of counts or frequencies of observations, where the length of the bar represents the corresponding value. However, bar graphs are often used to summarise multiple data-points per condition. In this situation, the top of the bar equals the mean value calculated from the data points. The bars are often accompanied by error bars that show the standard error of the mean (SEM) or standard deviation (SD). This type of plot has been coined a ‘dynamite plunger plot’ (footnote 1) for its cartoonesque similarity to a detonator for explosives. The dynamite plunger plot has gained wide popularity and is often the graph of choice to summarise and present data in presentations or manuscripts. One of the reasons for the popularity of bar graphs is that these are easily made by (commercial) software.
Bar graphs are bad graphs
A bar graph with errors bars has one major problem: it conceals the underlying data. Bar graphs do not allow independent interpretation of the data by the reader of a manuscript or the audience of a presentation. Moreover, it is often unclear what the error bars depict (SEM, SD or 95% confidence intervals). Many related issues that add to the notion that bar graphs are bad graphs and should no longer be used have been repeatedly addressed by others, for example here, here, here and here. Clearly, the bar graph is in need of a complete make-over.
Steps toward transparent data presentation
To improve transparency in data presentation, several features of the bar graph can be modified. Below, I describe 5 steps, departing from a standard bar graph, that aim at improving transparency and interpretation. Each step changes one aspect, as shown in figure 1 (or in this animated version). A motivation of each step is presented below.
Figure 1: A makeover of the bar graph in five steps.
Remove the bar
The top horizontal line of the bar equals the average value. This single value is the only information carried by the bar, and therefore the bar (except for the horizontal line) can be removed without loosing information. This action increases the data-ink ratio, which is defined by E.R.Tufte, pioneer in the field of data visualization, as the ratio of non-erasable data-ink to the total amount of ink used in the graph. To further increase the data-ink ratio, a single dot and two lines could be used to depict the average and error margins, respectively.
Show the data
Error bars may not give a realistic impression of the variability of the data. The error bars conceal outliers, multi-component distributions and asymmetric distributions. Therefore, it is more informative to show the actual data-points as dots. When many data-points need to be plotted (>100) the dots may start to overlap. This can be remedied by using semi-transparent dots. Alternative ways to show dense distributions is by using a bean plot or a sinaplot, which is an improved version of the violin plot.
Replace the mean by the median
The mean value, indicated by the horizontal line, informs us about the central value of the data. Mean values, however, are sensitive to outliers and may not be a proper representative of the central value in case of asymmetric data distributions. An alternative, robust measures of the central value is the median (or geometric mean). The median is not sensitive to outliers and equals the mean when the data adheres to a normal distribution. In case of asymmetric distributions, the median is a better indicator of a typical value of the data. Since the median is a more robust indicator of the central value of the data, the mean should be replaced by the median. When the number of data points, n, is low (typically n<10), it is recommended to show only the data and median, omitting any error bars. So, in case of low n, the make-over of the bar graph is completed at this stage.
Add 95% Confidence intervals
In the original bar graph, the error bars depicted the standard error of the mean (SEM). There are convincing arguments that 95% confidence intervals (95%CI) are better suited to summarise variation. First, the 95%CI give a more realistic impression of the variation in the data than SEM. Second, the 95%CI can be used for statistical inference by eye, i.e. judging whether two conditions are statistically different. If the 95%CI do not overlap, this implies a statistical difference. In this example, the 95%CI around the median is calculated by using the equation from McGill et al. (1978). An alternative strategy that can be used to calculate the 95%CI of the median is by bootstrapping, allowing for asymmetric 95%CI.
Add a box
Instead of the median (or mean) with 95%CI, a notched boxplot can be shown to summarise the data and allow for inferences. By convention, the center of the boxplot indicates the median, and the limits of the box define the interquartile range (IQR = middle 50% of the data). The notches indicate the 95%CI around the median, which is an estimation of the interval that includes the population median in 95 out of 100 cases, if the experiment was performed multiple times. The whiskers (shown here as vertical lines) can be defined in multiple ways. Here, we used the Tukey definition, i.e. the whiskers extend to data points that are no further from the box than 1.5*IQR.
Conclusion
The bar plot has infested the scientific literature. The disadvantages of bar plots have been documented by many. Several good alternatives for bar plots exist, which allow for a more transparent presentation of results and enable inferences by eye. The end result of the complete make-over of the bar graph that is presented here is the box&dotplot. The box&dotplot differs in at least five essential aspects from a bar graph with error bars and is a major improvement since the box&dotplot reports data in a transparent way, enabling independent interpretation of the results by others. In an era of increased focus on open science and data availability it is time to step away from the bar graph and expose the data.
Methods
The graphs were made using R/Rstudio with the library ggplot2. The data and code is available at http://doi.org/10.5281/zenodo.375944. A user-friendly alternative to create dotplots and boxplots online is provided by boxplotR.
Shout-out
I am grateful to anyone sharing code, the twitter community and to my colleagues for comments and the exchange of thoughts.
Footnote 1: I attempted to trace the origin of the term ‘dynamite plunger plot’ (with help from Gordon Drummond and Sarah Vowler). One of its first documented uses was in the books ”How to display data” and “Statistics at Square One“, co-authored by M.J.Campbell. In an e-mail, replying to my inquiry, Michael J Campbell states “I am pretty sure I thought of the phrase, since that is what they look like, but others may also have thought of it.”
The idea sounded intriguing at first: 30 scientists in an isolated and stunning old house discussing intercellular interactions in the context of tissues and organs. It became even more intriguing when we were asked to prepare slides about ourselves, not only about our research but also about our non-scientific life!
The day before the workshop began, a puzzled friend told me “why would you want to go to such secluded place and spend 4 days surrounded by scientists?” At the beginning the question made me laugh, but then it made me think about the unique context of the meeting. Definitely I could not anticipate what I was going to encounter.
The meeting took place at Wiston House, a historic and really charming country house filled with British history. We even got a chance to hear all the tales and chronicles of the place from the historian of Wiston House. Currently, it is used as a conference centre to promote discussion and exchange of ideas, an ideal scenario for a scientific meeting!
The first surprise came when I saw the meeting room. Instead of sitting in rows, we were around a horseshoe-shaped table. This may sound a trivial detail, but I think it definitely helped the discussion and the interactions. The first session was opened by one of the organisers, Andy Ewald, who described specific molecular requirements for mammary epithelial cell invasion. Among other examples, he showed that collective invasion requires leader cells to adopt an epithelial program (Cheung et al., 2013). Parallels were drawn between the cellular features of branching morphogenesis during development and of collective epithelial migration during cancer invasion (Ewald et al., 2008). This idea was especially relevant, given that the workshop brought together cellular and developmental biologists and cancer researchers. During gastrulation, cells undergo changes in shape, position and fate at the primitive streak. Kees Weijer presented a very detailed light-sheet microscopy study of primitive streak formation in the chick embryo, which is characterized by large-scale tissue movements (Rozbicki et al., 2015). On the other hand, Peter Friedl presented beautiful two-photon microscopy studies of collective cancer cell invasion in vivo. A cross-fertilization of ideas could help us understand the complexity of embryonic and cancer cell behaviors. To achieve this goal, the development of new and improved imaging techniques will be fundamental, as was nicely shown by Scott Fraser.
The second surprise came at dinnertime: seats were assigned! We have all been at big meetings in which students and postdocs end up sitting together, and PIs, who in many cases have known each other for a really long time, interact among themselves. The idea of assigning seats turned out to be simple and very effective. We all had the opportunity to the engage in stimulating conversations with PIs, postdocs, students and editors. Scientific interactions are critical, and so are cellular interactions, both in physiology and disease. Elke Ober showed how interactions between epithelial and mesenchymal cells regulate organ positioning during development. In particular, Ephrin-mediated crosstalk between lateral plate mesoderm cells and hepatoblasts coordinates the migration of these two cell types to position the embryonic liver (Cayuso et al., 2016). Another example is the effect of stromal cells on branching of the mammary gland. Johanna Ivaska showed that expression of SHARPIN in mesenchymal cells is required to organize the extracellular matrix allowing ductal outgrowth (Peuhu et al., 2017). By using simplified model systems, we can also gain relevant information on the mechanisms of organogenesis. With this in mind, Anne Grapin-Botton talked about the cellular interactions that regulate 3D pancreatic organoid formation (Greggio et al., 2013). On the other side of the coin, interactions between Cancer-Associated Fibroblasts (CAFs) and tumor cells are known to promote cancer growth and invasion, but whether this requires a direct physical interaction has not been addressed. Erik Sahai showed a mechanical coupling between CAFs and tumor cells that drives collective invasion (Labernadie et al., 2017). Paul Timpson took a translational approach and discussed the results of targeting the stroma for cancer treatment (Pajic et al., 2015).
The third unconventional event was the out of the box sessions, dedicated to reflect on specific concepts and topics. A recurring theme was junctional tension. We discussed how to define it, how to measure it, and what are the implications for tissue morphogenesis, aspects that were also covered by different speakers. Alpha Yap and Carien Nissen analysed the role of adherens junctions as mechanotransducers using different epithelial models; and Valerie Weaver discussed how the mechanical properties of the substrate impact on human embryonic stem cell differentiation (Przybyla et al., 2016). When discussing junctional tension, there is a key element to take into consideration: the actomyosin cytoskeleton. Benedicte Sanson discussed how a planar polarized localization of actomyosin regulates collective cellular movements during Drosophila germ-band extension (Tetley et al., 2016). This was complemented by the studies of John Wallingford and Danelle Devenport on the role of planar cell polarity proteins in the control of epithelial morphogenesis. Michael Way and Laura Machesky presented their latest research on the regulation of actin dynamics and reorganization using different model systems (vaccinia infection and melanocyte migration respectively).
By the end of the workshop I remembered the words of my friend and wondered why I wanted to be part of it. The workshop was superb in so many different ways. Indeed, there were only reasons to be part of it! But probably the best reason was inspiration. Many talks and ideas were inspirational, and I would like to highlight the concept of “building instead of destroying” that was presented by Dan Fletcher and Darren Gilmour. By interfering with the function of genes through genetic studies we have learned a lot about the function of specific molecules and pathways. The time has come to move ahead, and the current challenge is to interfere with protein function in a spatiotemporal-controlled manner, and to devise ways to study how the different components of a cell self-assemble to generate cells, tissues and organs. Meetings like this provide the context to make it happen.
I am grateful to Katherine Brown for comments on the text.
Heather M. Meyer1, José Teles2, and Pau Formosa-Jordan2
1 Weill Institute for Cell and Molecular Biology and the graduate field of Genetics, Genomics, and Development, Cornell University, Ithaca, NY, 14853 USA
2 Sainsbury Laboratory, University of Cambridge, Cambridge CB2 1LR, UK
A longstanding question that has fascinated both biologists and physicists alike is how a group of genetically identical cells differentiate into specialized cell types and form intricate spatiotemporal patterns. To form cell patterns, undifferentiated cells must somehow adopt specialized cell fates as well as achieve specific spatiotemporal control over their division patterns. In the field of biology, much effort has been made to identify the genes that facilitate the patterning process. Yet, due to past limitations in imaging and quantification methods, little experimental research has been performed to observe and test the dynamic behaviors of these regulators in vivo. Theoretical approaches, such as mathematical/computational models, have thus been used to recapitulate the behavior of such genes and predict the possible underlying regulatory mechanisms. For instance, Lewis Wolpert’s French Flag model proposed that a population of cells will differentiate into different cell types by interpreting varying concentrations of a diffusible morphogen (Wolpert, 1969). This model, in complement with previous experimental data, led to the realization that Bicoid, a maternal effect gene that controls anterior patterning, acts as a diffusible morphogen to initiate the anterior-posterior patterning process of the Drosophila embryo (Briscoe and Small, 2015; Ephrussi and Johnston, 2004; Grimm et al., 2010).
Interestingly, there are various patterning systems that form without the use of a known diffusible signal. In these systems, modelers such as Alan Turing have suggested that stochastic gene expression may initiate those patterns by creating small differences that are then amplified through regulatory feedback mechanisms, ultimately leading to different fates (Turing, 1952). Although extensively explored, these behaviors have been sparsely observed in vivo within a multicellular patterning system (Kondo and Miura, 2010). Thus, it was the idea of visualizing and quantifying the emergence of cell patterns in systems where no known morphogen had been identified that brought together the collaboration between Heather- experimental biologist and confocal imaging specialist from Cornell University, José- computational biologist from Sainsbury Laboratory (SLCU), and Pau- biophysicist with a modeling background also from SLCU, to analyze the formation of giant cells on the Arabidopsis sepal. Here, we describe how our past and present interests inspired the collaborative efforts and discoveries of the dynamic nature of ATML1, and how it impacts giant cell fate decisions in the sepal, which was published recently in eLife (Meyer et al., 2017).
Heather’s narrative:
I have always been interested in the development of patterns. When I was an undergraduate I worked in an algal systematics lab, in which I used oospore membrane patterning together with phylogenetics to taxonomically classify different algal species. I was so fascinated by the different patterns observed, that I left the field of systematics and decided to pursue a PhD at Cornell University with a focus on developmental genetics.
During my PhD, I worked with Dr. Adrienne Roeder. Adrienne was a new professor at Cornell University studying the role of variability in giant cell patterning. To study this, Adrienne used a methodology called computational morphodynamics, an analytical approach that melds imaging living plants, image processing and computational modeling to study in vivo developmental processes (Chickarmane et al., 2010; Jönsson et al., 2012; Roeder et al., 2011). I was enamored by the elegancy and novelty of applying computational morphodynamics to study patterns, especially the use of time-lapse imaging for analyzing cell growth and patterning dynamics. I therefore joined Adrienne’s lab, in hopes to employ similar techniques and aid in investigating the giant cell patterning system.
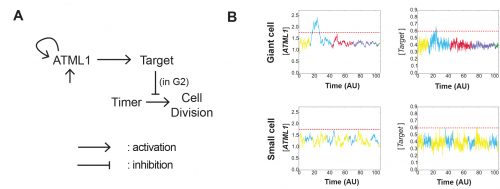
Adrienne had previously performed a forward mutagenesis screen to look for genes that affect the formation of giant cells. In this screen, Adrienne found a handful of mutants, including the epidermal specification gene ATML1 (Roeder et al., 2010; Roeder et al., 2012). ATML1 had been previously shown to be needed for epidermal specification during embryogenesis (Abe et al., 2003). Additionally, Adrienne published that ATML1 was required for the formation of giant cells and was expressed in all sepal epidermal cells (Roeder et al., 2010; Roeder et al., 2012). Thus, a major question I wanted to better understand, was if ATML1 is expressed in all epidermal cells, then why do only certain cells become giant on the sepal? To start addressing this question, we started a collaboration with Gwyneth Ingram and her graduate student at the time Rita San Bento, who had created an ATML1 overexpression line by expressing ATML1 under the epidermal specific PDF1 promoter. To our surprise, these lines produced an ectopic giant cell phenotype, consequently leading us to the initial hypothesis that a cell’s ability to become giant is sensitive to ATML1 dosage. To test this hypothesis, I created plants containing different doses of ATML1. When analyzing these plants, I found that indeed, epidermal cells were sensitive to the amount of ATML1 and that the concentration of ATML1 was correlated with the number of giant cells that formed on the sepal (Figure 1).
Figure 1. Sepals from genetically modified plants expressing different levels of ATML1. ATML1 expression levels in sepals descend in order from A-F. Reproduced from Meyer et al. (2017).
With this in mind, Adrienne and I wondered whether during development, different cells either express different amounts of ATML1 or if cells express the same amount of ATML1 but just exhibit differential perceptions. To answer this question, I created transgenic plants that possessed a fluorescently labeled ATML1 protein, in order to visualize ATML1 in individual cells. In these plants, I noticed that by eye sepal primordial cells seemed to contain different amounts of ATML1, whereas in the meristem ATML1 levels seemed uniform. With this discovery, I knew that I would have to live-image and track ATML1 through development quantitatively analyze its dynamics over time (Video 1). However, I was unsure of the best way to quantify expression levels of ATML1. Thus, Adrienne and I went to attend the 2014 EMBO conference on Interdisciplinary Plant Development at the SLCU, with the intention to find collaborators that would be knowledgeable about quantifying ATML1 and modeling its expression.
Video 1. A time-lapse video of a developing flower expressing fluorescently-tagged ATML1. These images were analyzed using our image quantification pipeline. Reproduced from Meyer et al. (2017).
At the EMBO conference, I was asked to give a lightning talk. This is a 5-minute short talk to advertise your poster. During this talk, I introduced the ATML1 giant cell story and asked anyone with questions and/or comments to come talk to me after. Both José Teles and Pau Formosa-Jordan independently approached me about collaborating on the project. José, had had previous experience studying stochasticity and had extensive image analysis experience and Pau had previously modeled tissue patterning, and had experience in salt-and-pepper patterning mediated by Notch and Delta. Thus, the three of us came together to both qualitatively and quantitatively understand how the behavior of ATML1 led to giant cell patterning. We skyped multiple times a week, often bouncing ideas off of one another via skype messenger.
José’s narrative:
For the past ten or so years, the main focus of my research has been to understand how cells make decisions. There is something intrinsically beautiful in how such complex molecular networks coordinate to effect discreet – and often times radical – outcomes regarding growth and division (Schmoller et al., 2015), dormancy and activity (Kuchina et al., 2011), life and death (Paek et al., 2016) in individual cells. During my PhD at Lund University and a short postdoc at UCL immediately after, I addressed these questions by developing deterministic and stochastic models of cell fate decision in the hematopoietic system, grounded on both population-averaged and single cell gene expression data. The work being developed in Henrik Jönsson’s group at the SLCU at the time I was finishing my postdoc was very appealing to me because they were applying computational tools to quantify live-imaging data during Arabidopsis growth. This contrasted with my previous experience where datasets were static snapshots of populations and/or developmental time points. It rapidly became clear to me that for my next research project I wanted to work in developing models of plant tissue that could incorporate spatiotemporal dynamics of gene expression at the single cell resolution. I specifically focused on studying the regulatory dynamics of stem cell identity and differentiation in the shoot apical meristem.
It was in this context that I attended and was immediately captivated by Heather’s presentation in Cambridge. The Arabidopsis sepal is an incredibly elegant system for studying cell fate decisions and pattern formation, and the beautiful live imaging data presented by Heather made that exceedingly clear. The fact that we could follow the expression dynamics of ATML1 as cells underwent the decision to endoreduplicate (i.e. an alternative cell cycle where the genome doubles without dividing) or divide opened a world of possibilities, and to this day, I am glad I approached Heather and, together with Pau, we were able to tackle this question with such a complete experimental and computational toolkit. The first challenge we faced was how to quantify ATML1 dynamics in single cells in the time lapses. I was working on image processing for my own project and realized there were a number of tools we could use: Costanza allowed for very accurate nuclear segmentation, MorphographX had a very convenient graphic interface for visual inspection and manual corrections and at the desk next to mine a postdoc in the Jönsson group, Yassin Refahi, was expanding ALT, which showed great promise for image registration and cell tracking. The issue, as is often the case with computational tools, was that these programs did not really communicate well with each other. It just so happened that in August 2015 their developers met under the same roof for two weeks, for a computational workshop organized at the SLCU every two years. Incompatibilities in file formats were resolved during that period and it became possible to analyze any particular image using the combined strength of the different tools. As I developed the image processing pipeline, Heather visited Cambridge for two weeks and together we ran through a number of configurations, selecting the one that allowed the most accurate results in quantifying ATML1 as well as cellular parameters of size and shape (Figure 2).
Figure 2. Initial tentative outline (A) and final implementation (B) of the image analysis pipeline for spatiotemporal quantification of ATML1 expression and cellular parameters. Adapted from Meyer et al. (2017).
The initial results were immediately interesting, since we could see a difference in ATML1 distributions between the meristem and the early sepal, with the latter exhibiting more heterogeneity (Figure 3A).
Figure 3. Two graphs demonstrating variation in ATML1 expression levels. (A) A histogram of fluorescently-tagged ATML1 in the sepal (red) and in the meristem (blue). ATML1 expression levels are more widely distributed in the sepal. (B) A plot of the coefficient of variation (CV) for fluorescently-tagged ATML1 in the meristems and sepals, as well as fluorescently-tagged proteins VIP1, AP2, and SEC24A, also expressed in the sepal. Note that the CV for ATML1 expressed in the sepal is higher than in all other cases. Reproduced from Meyer et al. (2017).
This suggested that ATML1 levels fluctuated more in the sepal than in the meristem, and we later confirmed that this was not the case for other proteins expressed in the sepal heterogeneity (Figure 3B). The dynamics of ATML1 in the sepal however were less clear. Our initial approach was to identify giant and small cells at the end of the time course and backtrack to the starting point, hoping to find differences in expression patterns that correlated with cell fate. When we did this, there were no immediately apparent differences between the two (Figure 4A). Upon closer visual inspection, Heather seemed to notice that cells which exhibit high peaks of ATML1 expression right before entering endoreduplication went on to become giant cells. This was consistent with our working hypothesis that a threshold-based mechanism could be involved in the decision but we needed to quantify the data in order to determine if such a threshold existed at all. Making use of my experience in a previous project, I approached this as a classification problem. If our hypothesis was true, and we compared maximum ATML1 expression peaks in cells that endoreduplicated (giant) versus cells that divided (small), we should be able to find a peak ATML1 expression value that accurately classified cells as giant or small and this value would essentially constitute the decision threshold. When we first applied this method, we realized that ATML1 peaks predicted giant cell fate with a mean accuracy of 0.7 (Figure 4B), which is a reasonable value (using a receiver operator characteristics (ROC) analysis for quantification; for reference, accuracy=1 is a perfect classifier and 0.5 is not better than flipping a coin).
Figure 4. (A) ATML1 concentration levels tracked throughout time for cells that eventually endoreduplicate and become giant (red) and cells that go on to divide and stay small (blue). ATML1 peak concentration levels observed in each small or giant cell lineage (left) and the corresponding ROC curve (right) for all cells regardless of ploidy (B), only for 2C cells (C) and only for 4C cells (D). Accuracy quantified by the area under the ROC curve (AUC) for each case; dashed lines on the ATML1 peak concentration plots mark the optimal predictive ATML1 concentration threshold. Reproduced from Meyer et al. (2017).
The most striking result however was when we observed that all the predictive value came from peaks observed in 4C cells (Figure 4D, mean accuracy 0.8), associated with the G2 stage of the cell cycle, and none at all from the 2C cells (Figure 4C, mean accuracy 0.5), associated with the G1 stage. This strongly suggested that in order to make the decision to endoreduplicate, cells must both experience fluctuations of ATML1 above a specific threshold level and be specifically in the G2 stage of the cell cycle (Figure 5). From an evolutionary perspective, it is exciting to speculate that in order for the proportion of giant to small cells to be robust across plants, there must have been selective pressure on the dynamical features of the ATML1 fluctuations, the ATML1 threshold level, the time window in which a peak in ATML1 above the threshold is productive or a combination of these.
Figure 5. Schematic depiction of the threshold-based mechanism of cell fate decision and pattern initiation. (A) Fluctuations in ATML1 concentration above a threshold level can lead to endoreduplication if they occur during the G2 (responding) but not the G1 (impervious) stage of the cell cycle. (B) A 4C cell (corresponding to the G2 stage) will endoreduplicate and become 8C if it experiences high concentrations of ATML1 above the threshold, or divide and originate two 2C daughter cells if it only experiences low concentrations below threshold. The ATML1 threshold inferred by ROC analysis for 4C cells is 80% accurate in predicting giant versus small cell identity. Reproduced from Meyer et al. (2017).
Pau’s narrative:
For a long time, I have been fascinated by the patterning process of developing tissues, especially in how initial differences in gene expression can become amplified to spur the patterning of different cell types. To start addressing this question, I did my PhD at the University of Barcelona, focusing on how Notch and Delta drive a pattern of two interspersed cell types in different developmental scenarios in animal tissues. After my PhD, I pursued a joint postdoc at the SLCU with the Locke, Jönsson and Meyerowitz groups, in order to better understand different patterning phenomena.
During my first year of my postdoc, I met Heather at the EMBO conference. I found the sepal patterning system extremely captivating and realised that jumping into such collaboration with the Roeder lab would give me the possibility to study in vivo gene expression data of the key regulator that presumably was driving a patterning process. I found this was something totally new, and I loved that I had the opportunity to model it!
Heather’s ATML1 fluorescent images were really promising, but difficult to initially interpret. Together with James Locke, Henrik Jönsson and José, we decided that José could do the image analysis, and I would focus my energies on the modeling part. Modeling this patterning phenomenon would be challenging, given that it seemed to be the result of the interplay of stochastic cell decision making, cell growth and cell division. Luckily, Henrik had this great software package called Organism-Tissue (http://dev.thep.lu.se/organism/), which would greatly facilitate the implementation and integration of these different elements.
The model had many different iterations during the collaboration. Initially, it started as a stochastic positive feedback that was simulated in spherically growing cells, where the ATML1 basal gene expression per cell volume scaled with DNA ploidy. This scaling presumably stabilized a high ATML1 state, corresponding to the giant cell fate. Yet, soon after quantifying and analyzing the live-imaging data, we realized that the scaling effect did not exist, causing the model to get amended into a threshold-based mechanism model. In this model, ATML1 regulates the expression of a downstream target, and only if the target reaches a hard threshold, will it enable the cell to escape mitosis and enter endoreduplication. This hard threshold in the target naturally encoded a ‘soft threshold’ in its upstream regulator ATML1, causing cells above a certain ATML1 threshold to likely become giant cells.
The model had further modifications to provide a robust comparison between the experimental and the computational data. For instance, having a more tissue-like geometry with anisotropic growth which drives ATML1 and target concentration dilutions, introducing a noisy timer to get the normally mitotic cells to divide – while keeping under control the instability that this timer introduced into the system -, and getting the dynamics of the modeled variables in lower time resolutions as an output. Even with those modifications, it was still difficult to establish clear comparisons between the simulated and the experimental data.
A key moment was when José came up with the idea to assess the ATML1 threshold mechanism via a ROC analysis. The ROC analysis showed that high levels of ATML1 in 4C but not in 2C cells were a proxy for giant cell fate commitment. We incorporated this observation in the model, proposing the G2 phase as the time window at which cells would be sensitive to ATML1 target levels for promoting endoreduplication. Moreover, the ROC approach enabled a more straightforward comparison between the ATML1 time-traces and the simulated ATML1 traces. Indeed, this ROC approach, together with the quantification of the variability of ATML1 in the tissue, resulted in a plausible region of the parameter space in which the ATML1 model was working, where fast and relatively small ATML1 fluctuations would lead to the giant cell pattern (Figure 6 and Video 2). Non-intuitively, the model together with the data showed us that the positive feedback on ATML1 should be weak. This was later tested experimentally, nicely demonstrating how model predictions can lead to new hypotheses that can be experimentally tested.
Figure 6. A mathematical and computational model for studying how ATML1 fluctuations drive giant cell patterning. (A) Cartoon of the model. (B) Example of simulated ATML1 and target levels. ATML1 fluctuations activate a target gene that, when having higher levels above a certain threshold, cell division is repressed. The model makes this threshold mechanism is just active during the G2 cell cycle. Horizontal lines in B represent the target threshold and the ATML1 soft threshold. Color codes in B represent ploidy levels (see Video 2). Modified from Meyer et al. (2017).
Video 2. In silico growing tissue emulating a developing sepal, where ATML1, target, cell timer levels and cell ploidies are represented. Cells that cannot divide, increase their ploidy, becoming giant cells. Reproduced from Meyer et al. (2017).
Although the model was pivotal in understanding the dynamics of ATML1, there are still a lot of open questions that the model did not address. For instance, how ordered is the emerging pattern? Is there any cell-to-cell communication that shapes the pattern? How are ATML1 fluctuations transduced to its downstream targets and ultimately perform the cell decision making process?
Conclusion:
We would be remiss if we didn’t stress that since the start and throughout the development of the project, there was constant communication between Cornell and Cambridge, and permanent iteration and intellectual contributions from everyone towards experimental planning, data analysis and model development. The fragmented nature of this post may have given the reader the impression that this work was sequentially produced in a compartmentalized manner, and that couldn’t be farther from the truth. This truly open and dynamic multidisciplinary environment was in our opinion the most important factor in leading this project to fruition, and one of our most rewarding experiences as scientists.
References
Abe, M., Katsumata, H., Komeda, Y. and Takahashi, T. (2003). Regulation of shoot epidermal cell differentiation by a pair of homeodomain proteins in Arabidopsis. Development130, 635–643.
Briscoe, J. and Small, S. (2015). Morphogen rules: design principles of gradient-mediated embryo patterning. Development142, 3996–4009.
Chickarmane, V., Roeder, A. H. K., Tarr, P. T., Cunha, A., Tobin, C. and Meyerowitz, E. M. (2010). Computational morphodynamics: a modeling framework to understand plant growth. Annual review of plant biology61, 65–87.
Ephrussi, A. and Johnston, D. S. (2004). Seeing is believing: the bicoid morphogen gradient matures. Cell116, 143–152.
Grimm, O., Coppey, M. and Wieschaus, E. (2010). Modelling the Bicoid gradient. Development137, 2253–2264.
Jönsson, H., Gruel, J., Krupinski, P. and Troein, C. (2012). On evaluating models in Computational Morphodynamics. Curr Opin in Plant Biol15, 103–110.
Kondo, S. and Miura, T. (2010). Reaction-Diffusion Model as a Framework for Understanding Biological Pattern Formation. Science329, 1616–1620.
Kuchina, A., Espinar, L., Çağatay, T., Balbin, A. O., Zhang, F., Alvarado, A., Ojalvo, J. G. and Süel, G. M. (2011). Temporal competition between differentiation programs determines cell fate choice. Mol Syst Biol7, 557.
Meyer, H. M., Teles, J., Formosa-Jordan, P., Refahi, Y., San-Bento, R., Ingram, G., Jönsson, H., Locke, J. C. W. and Roeder, A. H. K. (2017). Fluctuations of the transcription factor ATML1 generate the pattern of giant cells in the Arabidopsis sepal. eLife6, 635–41.
Paek, A. L., Liu, J. C., Loewer, A., Forrester, W. C. and Lahav, G. (2016). Cell-to-Cell Variation in p53 Dynamics Leads to Fractional Killing. Cell165, 631–642.
Roeder, A. H. K., Chickarmane, V., Cunha, A., Obara, B., Manjunath, B. S. and Meyerowitz, E. M. (2010). Variability in the control of cell division underlies sepal epidermal patterning in Arabidopsis thaliana. PLoS Biol8, e1000367.
Roeder, A. H. K., Cunha, A., Ohno, C. K. and Meyerowitz, E. M. (2012). Cell cycle regulates cell type in the Arabidopsis sepal. Development139, 4416–4427.
Roeder, A. H. K., Tarr, P. T., Tobin, C., Zhang, X., Chickarmane, V., Cunha, A. and Meyerowitz, E. M. (2011). Computational morphodynamics of plants: integrating development over space and time. Nat Rev Mol Cell Biol12, 265–273.
Schmoller, K. M., Turner, J. J., Kõivomägi, M. and Skotheim, J. M. (2015). Dilution of the cell cycle inhibitor Whi5 controls budding-yeast cell size. Nature526, 268–272.
Turing, A. M. (1952). The Chemical Basis of Morphogenesis. Philos. Trans. R. Soc. Lond., B, Biol. Sci.237, 37–72.
Wolpert, L. (1969). Positional information and the spatial pattern of cellular differentiation. J Theor Biol25, 1–47.
Elin Sørhus, John P Incardona, Tomasz Furmanek, Giles W Goetz, Nathaniel L Scholz, Sonnich Meier, Rolf B Edvardsen, Sissel Jentoft
Institute of Marine Research, Norway Northwest Fisheries Science Center, USA University of Oslo, Norway
Crude oil contains many chemicals which are toxic to both humans and aquatic animals. Fish embryos, in particular, are sitting ducks when it comes to oil pollution. The vast majority are deposited naked, unprotected by their mother, drifting around or stuck in a place on some plants or rocks. What happens if they drift through an oil spill or are deposited next to an oiled beach?
Oil spills are happening all the time. Well known large spills that got a lot of publicity are the Exxon Valdez oil spill in Prince William Sound in Alaska in 1989, which affected local animal populations for decades, and the Deepwater Horizon wellhead blowout disaster in the Gulf of Mexico in 2010, where we probably still don’t see the full consequences. However, we also find examples of recent spills that get very little worldwide attention. For example, there was a major oil spill near Chennai in India in February 2017 with a release of nearly 20 million litres of heavy bunker fuel oil. This latter spill was not mentioned in any European newspapers, suggesting there are more black oily numbers out there. Understandably, oil spills create a great concern for long-term effect on the environment, especially for fish spawning areas where eggs are released, and we search for tools to predict and measure these consequences.
The Deepwater Horizon disaster has really intensified the amount of studies investigating developmental abnormalities in fish induced by oil exposure. These studies identified the developing heart as the major target of crude oil. Exposure results in both functional and developmental abnormalities by directly interfering with ion currents in heart muscle cells (cardiomyocytes). This means that in addition to immediate effects on function during exposure, an altered heart shape resulting from an early life exposure also can generate a reduced cardiac output later in life. Reduced cardiac performance in turn will diminish swimming ability, potentially decreasing feed intake, predator avoidance and survival. For example, a reduced cardiac output in juvenile fish exposed to oil as embryos may have contributed to the collapse of the herring stock in Prince William Sound in Alaska a few years after the Exxon Valdez oil spill.
Deepwater Horizon wellhead blowout. Over 700 million liters of oil was released in the Gulf of Mexico, threatening marine life. Photo credit: US coast guard.
I work for a government institute that manages Norway’s marine resources, most importantly the fish stocks that are a major part of our economy. As an applied science agency, we need to understand how Norway’s other big economic activity, offshore oil production, potentially affects the fitness of our fish stocks. Crude oil’s effect on the heart and potential impacts on juvenile survival were therefore the primary focus when we performed our first experiment in 2013. At the time, I was a newly hired PhD student thirsting for challenges. I had just left my position as a lab technician with 10 years experience in the vast field of molecular biology. As a student in a Genomics program, I was recruited to perform RNA sequencing studies in oil-exposed marine fish with our institute’s Ecotoxicology group. The goal was to relate changes in the whole-animal embryonic and larval transcriptomes to developmental defects. But I was the rookie and totally a blank slate when it came to crude oil toxicity and developmental abnormalities. Fortunately, I was born into this world with an exceptional overload of energy which is easily transferred to hard work, long hours and optimism despite some downfalls. Also, past experience with experiments that did not turn out as expected taught me efficient trouble shooting, and when to change course and to easily create solutions to unexpected problems.
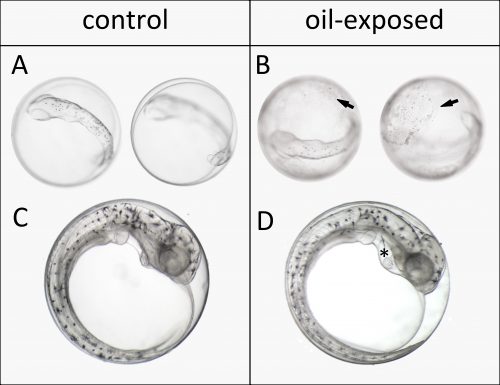
We started out working on embryos of Atlantic haddock (Melanogrammus aeglefinus) because it is an important fisheries resource in Norway and the rest of the North Atlantic. The main spawning grounds for haddock in Norway are targeted for expanded offshore oil production. In the first year our plan was to do two experiments, exposing early embryos and newly hatched larvae to dispersed crude oil. However, dealing with a non-model marine fish broodstock, getting embryos at the right time wasn’t so simple. Embryos were produced before we were really ready, but we managed to pull off an experiment exposing embryos after completion of major organogenesis. Even though this wasn’t the ideal experiment, we made a very striking and disturbing observation. Oil droplets stuck to the apparently relatively sticky chorion of haddock eggs. Presumably this increased and prolonged their exposure. Thus, despite not being exposed at their most vulnerable stages of development, these embryos still showed surprisingly high deformity rates by the time they hatched. This included large amount of edema formation in the heart sac, which is indicative of heart failure. But we weren’t able to assess these defects in sufficient detail.
Oil droplets bind to haddock eggs. Observation of oil droplets on the chorion after 72 hours of exposure to the highest oil dose (top). Edema (*) in the heart sac of 10 days old embryos exposed to the lowest oil dose (bottom).
To assess these effects more thoroughly, the year after, we invited John Incardona and his group to participate in the experiments. As a developmental biologist with over a decade of oil exposure experience, John taught us how to perfectly position and video image these delicate creatures. He showed us excitedly almost invisible development markers in our tiny haddock larvae, like the urinary tract and lateral line neuromasts. For me, imaging and sampling day in and day out, I dreamt about beating fish hearts and doing the swoop through methylcellulose to position the larvae in my sleep. Still, in the end I regret I didn’t take that extra shot. Nevertheless, I somewhat unknowingly created a sufficiently large and detailed picture and video library during development, which proved to be crucial for interpreting the RNA sequencing data.
The expert, John Incardona, at the microscope. Teaching us how to position and immobilize larvae and embryos in methylcellulose while imaging.
An applied science issue for natural resource agencies like ours is that there are too many species in the wild to develop species-specific tools on the level of model species like zebrafish. Working with non-model species like Atlantic haddock, for example, in situ hybridization probes and protocols are not readily available. Designing suitable species-specific assays and probes is time consuming and often very difficult because of lack of optimized methods and genome sequence knowledge. However, most of the fish species we care about, including haddock, have translucent embryos. The embryos therefore often can tell us directly what is happening with their development, if it’s normal or abnormal. We exploited this benefit when we used RNA sequencing to study normal development, linking changes in developmental processes and appearance of structures to temporal mRNA levels in a previous paper (Sørhus et al., 2016), showing that for example cardiac determination and differentiation genes preceded expression of genes related to cardiac function. However, would we be able to “see” abnormal structures represented in a temporal snapshot of mRNA levels? We know that altered structures typically have different distributions of cell types, theoretically this should be reflected in different levels of gene expression corresponding to cell type-specific markers. One example can be found in John´s early work (Incardona et al., 1998). He showed that the neural tube in embryos are mis-patterned after exposure to hedgehog-blocking teratogens. One pool of cell-types was replaced by another, and was measured by counting cells using antibodies for cell-type specific protein markers. If we can count cells this way, we should also be able to quantify them by mRNA sequencing. One important assumption however, is that mRNA transcript levels reflect protein levels. This is generally true in developing embryos, where genes are being turned on and off because gene function needs to be dynamic. Over-expression studies support this: generally if a cRNA is injected into an embryo, it makes a functional protein.
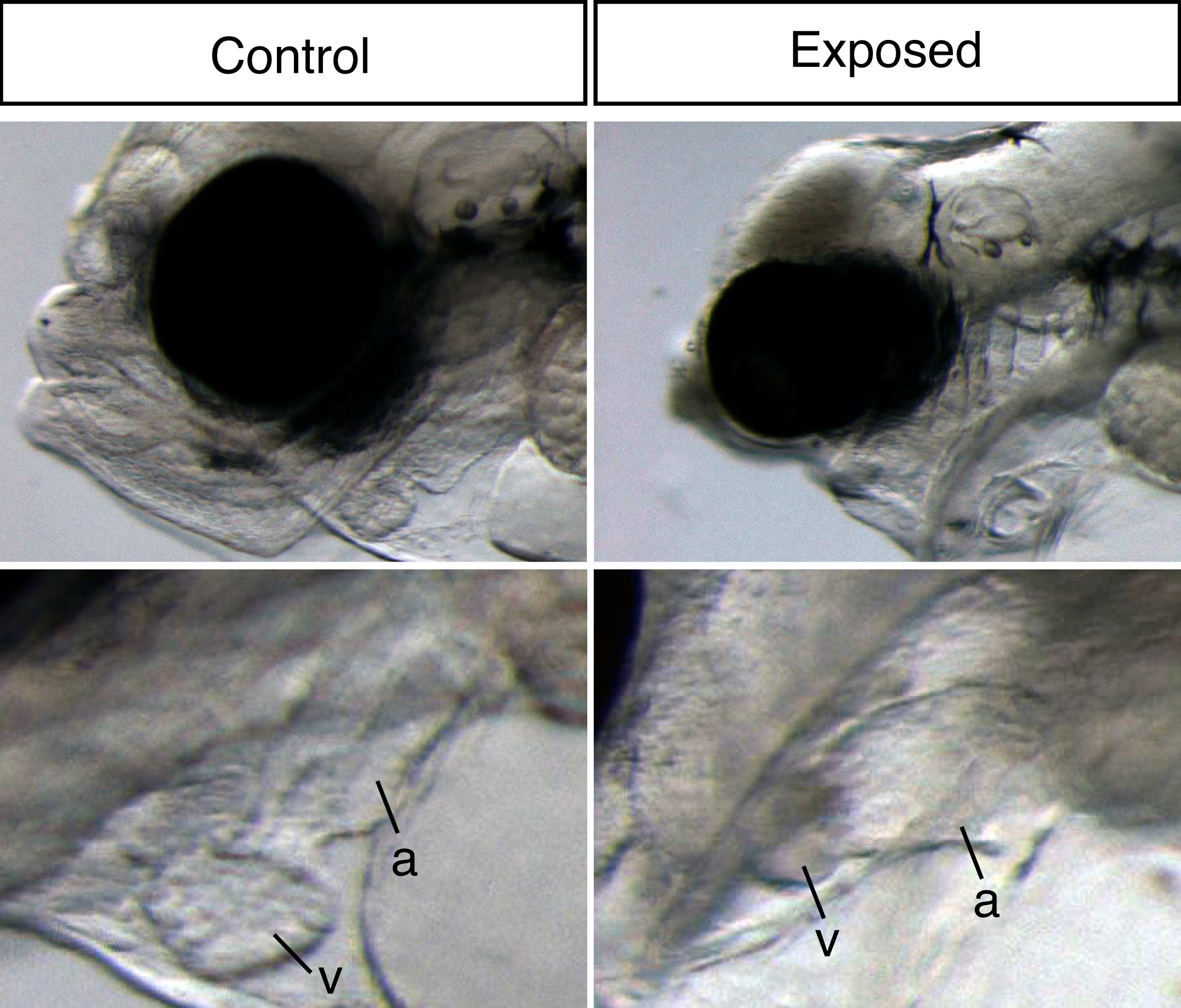
Embryonic oil exposure results in severe larval developmental abnormalities. Most striking was the extreme craniofacial abnormalities (top) and underdeveloped heart (bottom). a = atrium, v = ventricle.
Our large picture and video library monitoring abnormal development almost continuously through the exposure experiment gave us a unique possibility to link gene expression changes to visible phenotypes. Thus, our main aim was to link gene expression to oil induced phenotypes and ultimately identify initiating events potentially leading to these phenotypes. The ultimate goal is to create a toolbox of molecular indicators that can help predict the developmental consequences and thus survival after either accidental real life oil spills or chronic low-level oil pollution. With both the RNA sequencing and image data wrapped inside our computers, the newbie (myself) and the expert (John) started a year long interpretation journey.
When dealing with 20 000 genes and 11 developmental stages, the use of bioinformatics tools is, of course, inevitable. This included the use of tools developed in-house as well as several commercially available pathway analysis databases. These tools are highly based on the biomedical literature focusing on mammalian species, and thus, do not necessarily represent the same processes in a marine fish species. So while they are quite powerful at identifying altered pathways, the challenge is figuring out which of those altered pathways are relevant to phenotypes. To avoid potential pitfalls of these bioinformatics pathway tools and not use them simply as a meat grinder, we therefore manually inspected individual genes underlying affected pathways. For example, in our oil exposed haddock, “salivary gland” was a pathway that came up as affected, which is quite peculiar considering fish do not produce spit. However, when inspecting the underlying genes, we saw that they were clearly involved in ion and water balance, which fit our oil exposed phenotype perfectly. For the same reason, we also performed our own “pathways analysis” the poor man’s way. This meant (simply!) diving into literature of developmental biology and studying known mutant phenotypes resembling our oil exposed fish larvae, making our own massive lists of genes involved in, for example, craniofacial and cardiac development, tissues that visually were highly affected in oil exposed haddock. For me, this was like venturing into the deep end of the pool.
A major problem we grappled with was how to determine whether individual genes of interest were the correct tissue-specific isoform. In other words, how could we have some degree of confidence that tissue-specific genes could be seen in the pool of mRNA extracted from whole embryos. Although we experimentally lacked information about localization of expression, underway we realized how to exploit the RNA sequencing data for this information. What bioinformatics pathway analyses don’t do is account for expression patterns, they generally rely on genetic interactions identified in the literature. We therefore turned to expression pattern databases in zebrafish (like Zfin.org). Based on known mRNA localization from other species like zebrafish, we could correlate the number of transcripts determined by the RNA sequencing with tissue-specific patterns at any particular time point. For example, genes shown to be expressed only in the developing heart in zebrafish all had transcript numbers in a limited range. Thus, if a gene of interest had transcript levels well above that range, we knew it had to have a broader expression pattern. Similarly, pathway knowledge and mutant studies in the literature showed us what to expect for the down-stream consequences of an abnormal up or down-gene regulation. When manually curating gene pathways of craniofacial and cardiac development, this information was always in our minds.
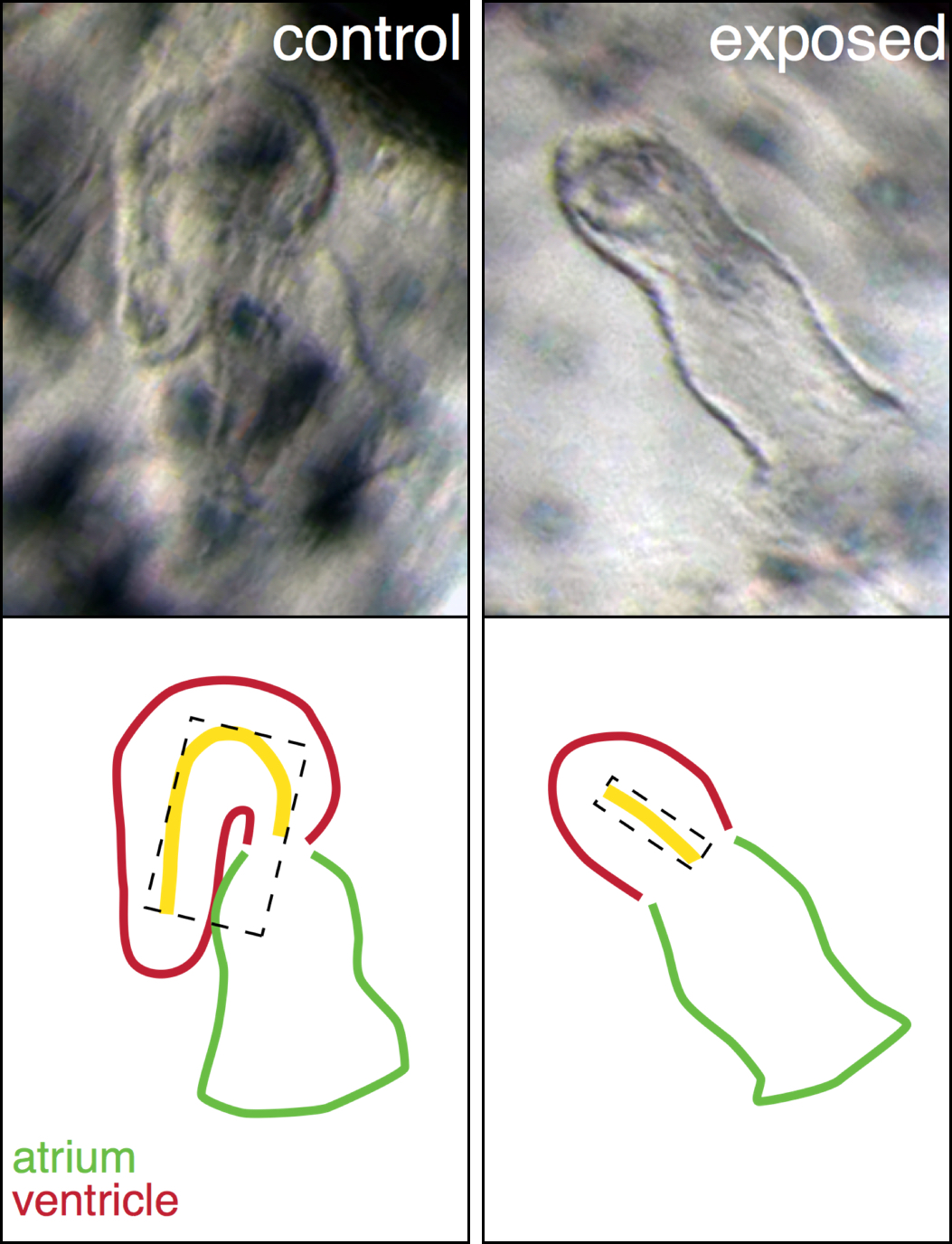
Abnormal heart shape in oil exposed haddock. The ventricle in the exposed animal is severely underdeveloped and un-looped.
A perfect example is our explanation for the abnormal heart shape in oil exposed haddock. Most of the hearts in the highest exposure group were un-looped, with the atrial and ventricular chambers in a linear rather than side-by-side arrangement, and the ventricle was extremely underdeveloped. From a large list of genes involved in cardiogenesis, one gene, bmp10, stood out and was abnormally regulated at one of the earliest time points. This gene was therefore a strong candidate for one of the first transcriptional events leading to the abnormal heart development. Bmp10 is a strong morphogen that is normally involved in the trabeculation process, the outgrowth of fingerlike structures in the ventricle. However, trabeculation is a process that is occurs much later in heart development, around hatching stage in fish. Remarkably, we detected bmp10 up-regulation when the heart was just at the “cone” stage, before the chambers are really separated and cardiomyocytes are even contracting. At this stage, another Bmp gene is normally expressed, bmp4, and it is needed to lateralize the cone and drive the asymmetric looping process to bring the chambers side-by-side. Functional studies in zebrafish demonstrated that, as a potent morphogen, the activity of bmp4 must be tightly regulated, otherwise either too much or too little leads to an un-looped heart. Therefore, inappropriately turning on the bmp10 signal too soon most likely overrides the normal bmp4 signal, leading to an abnormal, un-looped heart. Downstream targets of Bmp10 were also up-regulated such as nkx2.5, a key transcription factor in cardiac development. An inappropriate overexpression of nkx2.5 yields a reduced ventricle in zebrafish, just like the one we see in our oil exposed haddock.
The bmp10-story is an excellent example of the hypothesis-based search for changes in gene expression linked to a visible phenotype. However, an advantage of unbiased RNA-sequencing dataset is the opportunity to conduct analysis where no main hypothesis is fully settled. The differentially expressed genes and pathways pointed to phenotypes of which we were peripherally aware, but didn´t pay strict attention to. This leading us to also focus on edema accumulation in the context of abnormal osmoregulation and linking disordered lipid metabolism and up-regulation of cholesterol synthesis to reduced yolk absorption.
Real life accidental oil spills are probably more common than we fear. The fact that an oil spill with 20 million litres of oil spread on a shore in India without being mentioned in the European media is a scary example. Therefore, it is more important now than ever to understand the underlying mechanisms of crude oil toxicity. In this study, we provide a toolbox of tissue specific biomarkers that together might be included in an important monitor apparatus in risk and damage assessment in the future. Which means that in the time ahead we can collect samples after accidental oil spills, like the one in India, Gulf of Mexico or Alaska, and predict the developmental effects and thus the consequences for the individual and eventually the population in the affected area. On the other hand, our results provide an improved general understanding about low-level, complex mixture toxicity. Even mammalian embryos and fetuses are likely impacted by the vast array of chemicals we dump into the environment in ways that we don’t currently understand. Thus, the data in this eLife paper is also generally important, even for people.
Hello, my name is Tyler Carrier and I am a second year PhD student in the laboratory of Adam Reitzel at the University of North Carolina at Charlotte. Broadly speaking, this laboratory focuses on the ecology, evolution, and development of marine invertebrates and their life history stages. The basis of this work uses echinoderms and cnidarians, with most efforts pertaining to the intertidal starlet sea anemone Nematostella vectensis. I, however, have gone against this common thread and work with larval stages of echinoids (sea urchins); namely, the contribution of associated-microbiota in their development, evolution, and response to environmental variation.
Echinoids and their larvae
Sea urchins are part of the Phylum Echinodermata (Class: Echinoidea) and are close relatives to sand dollars (Echinoidea), sea stars (Asteroidea), brittle stars (Ophiuroidea), sea cucumbers (Holothuroidea), and feather stars (Crinoidea). Sea urchins are a group of bottom-dwelling (benthic) invertebrates found throughout the world’s ocean and are the primary grazers of sub-tidal kelp beds. In many cases, urchins spawn annually by releasing gametes into the water column where fertilization and larval development take place.
Left: Purple sea urchin, Strongylocentrotus purpuratus; Center: Red sea urchin, Mesocentrotus franciscanus; Right: Green sea urchin, S. droebachiensis. Larval forms for each are below.
Urchin larvae, termed the echinopluteus, develop in the water column for weeks to months, during which they are at the mercy of ocean currents and, as a result, may be transported great distances from their parental origin. While developing the echinopluteus faces a multitude of environmental variants, including salinity, temperature, and food quantity. Over the course of evolutionary time, the echinopluteus has coped with these variants by altering their physiology and developmental trajectory.
My dissertation aims to shed light on if and how associated microflora provide larvae the genomic compliment to cope with abiotic and biotic environmental variables. Microbes associated with the animal host often differ considerably in the laboratory from that in nature. Therefore, to most accurately assess my research questions, I journey from the comforts of my bench to the field, where I collect and spawn adult urchins and rear their larvae.
A typical day in the field
Days in the field do not, in any way, reflect those at the bench. Time in Charlotte is consumed by processing samples: DNA extractions, PCRs (and re-doing PCRs because of contamination – there are non-larval bacteria everywhere!), gel electrophoresis, using QIIME and R to analyze 16S rRNA datasets, and writing grants and manuscripts. When compared to the field, those days are boring! And, as with many field-oriented biologists, the few weeks spent in the field dictate productivity for the year. Below I will outline a day from recent fieldwork at the Friday Harbor Laboratories (University of Washington), a field station located on an island wedged between the Olympic Peninsula and Vancouver Island.
Days in the field are often never-ending, but the natural beauty between the land and sea are indescribable.
06:00: My alarm goes off, I stumble out of bed, and, as it was the day before, find that the sun is just rising. To start the day refreshed, I shower and head to breakfast.
07:00: Prior to caring to my larvae, I make a list of what must be done today and another for what I’d like to do if I find some spare time. The first list typically includes a tally of which cultures of larvae need to be sampled, imaged on the resident compound microscope, and need a water change and to be fed. Depending on the day or week, I tend to note that adult urchin cultures need feeding (incase I need more larvae) and to do so need to get in a row boat and collect drift kelp around the dock.
07:30: I begin by using a hemocytometer to approximate the density of my phytoplankton cultures (for feeding larvae).
08:00: One by one, I reverse filter larval cultures (three liter glass jars) down to ~100 mL. Each takes approximately ten minutes, and, depending on the day, I filter 12 or 24 cultures.
After each jar is reverse filtered, I transfer the remaining seawater to a finger bowl and, using a dissecting microscope, count ~100 larvae and transfer these to a 1.5 mL pre-labeled Eppendorf tube. This is one of the most important steps and focal points because here I am able to survey larval health, and am able to make anecdotal observations that may be interwoven with empirical results.
Following reverse filtration and sampling, larvae caged in Eppendorf tubes are spun into a pellet and seawater is replaced with a fixative (RNAlater) to preserve most DNA and RNA. These fixed samples are then moved into a -20 °C freezer.
12:30: With sampling complete, it is time to break for lunch and to catch up with resident and visiting scientists, their students, and those taking summer courses.
13:30: Having been fed, it is time to make sure the larvae are, as well. Based on phytoplankton counts this morning, I subsample phytoplankton cultures and allocate food quantities to their respective jars.
14:00: Now that husbandry is done, it is time to image larvae from each treatment. A small aliquot of larval cultures are transferred to finger bowls and are brought to the resident compound microscope.
16:00: To clear the mind from a taxing daytime session and to start de novo for the nighttime shift, I trade my laboratory attire for running gear and explore the island in a 45-minute to an hour run before dinner. Although this may take away from research time, it allows the rest of the day to be more focused and productive. Plus, the views aren’t half bad!
Jackson Beach, Friday Harbor, WA
18:00: To be able to distinguish larval-specific microbes from environmental microbes, it is essential to sample the seawater, as well. While waiting, this semi-free time allows me to read a paper or two.
19:30: To monitor the effect of my experimental treatments on larval growth, the evening begins by analyzing the images taken before dinner. For this I use ImageJ and a cup of coffee. Hey, at least the larvae are adorable!
22:00: One activity that hasn’t been featured yet is e-mail and writing, and that’s because larvae take priority. To conclude the day, I catch (back) up on e-mail and try to write a couple paragraphs for upcoming grants and manuscripts in preparation. This takes me until about mid-night, by which time I should make my way towards bed… but tend to get distracted with a game or two of billiards or ping-pong.
The use of organoids – which can be defined asartificially grown masses of cells or tissue that resemble organs – in basic and clinical research has snowballed in recent years, providing insight into fundamental developmental processes and disease etiology. Today’s paper comes from the new Special Issue of Development devoted entirely to organoids, and reports the use of human cardiac organoids to address the regenerative capacity of the immature heart. We caught up with first author Holly Voges, and her co-supervisors Enzo Porrello and James Hudson, who jointly run the Cardiac Regeneration Lab in the University of Queensland.
James, Enzo and Hannah in the lab
So James and Enzo, how did you come to co-run the Cardiac Regeneration lab in the University of Queensland?
JH I graduated with a Bachelor of Engineering (Chemical & Biological) from the University of Queensland in 2006. Rather than getting a job in the mining/oil and gas industry I decided to do a PhD in tissue engineering as I found this a more rewarding career path for my inquisitive personality. I graduated with a PhD in 2011 under the guidance of Prof Justin Cooper-White who was one of the prominent Australian researchers at the forefront of bridging the engineering and biology disciplines. From here I really wanted to apply my skills and propel my research in cardiac tissue engineering to the next level. I therefore approached Prof Wolfram-Hubertus Zimmermann one of the world leaders in cardiac tissue engineering to do a postdoc in his lab. My work in Prof Zimmermann’s lab from 2011 to 2012 was in part supported by a postdoctoral research fellowship from the German Cardiology Society, which enabled me to develop new cardiac differentiation protocols and tissue engineering strategies as part of the team Prof Zimmermann had put together. I was then awarded a National Health and Medical Research Council of Australia Early Career Fellowship for 2013-2016 and therefore decided to move back to Australia to continue my research in cardiac tissue engineering. Upon returning to Australia my goal was now to build on my previous work and use human cardiac organoids (hCOs) to find novel therapeutics for patients with cardiac disease. This is when I teamed up with Dr Enzo Porrello who also had the same goal with his work in mice on cardiac regeneration. Ever since then we have been working together to decipher how cardiomyocytes regeneration is regulated in the hope of finding novel regenerative therapeutics for cardiac disease.
“We have been working together to decipher how cardiomyocytes regeneration is regulated in the hope of finding novel regenerative therapeutics for cardiac disease”
EP I completed my PhD at The University of Melbourne and Baker Heart and Diabetes Institute. My PhD thesis focused on the developmental origins of cardiac hypertrophy, which is when I first became interested in understanding the mechanisms controlling cardiomyocyte cell cycle arrest in the neonatal period. I was subsequently offered a postdoctoral position in Eric Olson’s lab at UT Southwestern Medical Center and I moved to Dallas in 2009. During my time in the Olson lab, working together with Hesham Sadek, we uncovered a previously unappreciated regenerative capacity of the neonatal mouse heart. My research since then has focused on understanding the mechanisms that control cardiac regenerative capacity during the neonatal period in mammals. When I returned to Australia in 2012 to set up my independent laboratory at The University of Queensland, one of the questions I was very interested in was whether the human heart retained a similar regenerative potential during foetal/neonatal life. At the time, pluripotent stem cell-derived cardiomyocytes were emerging as a new technology platform to study human cardiac biology. When I first met James Hudson in 2013 we discussed numerous aspects of cardiac developmental and regenerative biology for several hours and realised that we had a unique opportunity to combine our skill sets to address some important questions in the field. We merged our labs in 2014 and our joint research program takes advantage of our respective skill sets in human cardiac tissue engineering and in vivo mouse models to develop much needed regenerative therapies for heart disease.
What is Brisbane, and Australia in general, like for stem cell and regeneration research?
EP & JH There are world class researchers in Brisbane in this area, not only in cardiac but in different fields of research. Australia wide the community is also strong as the Australian government has supported a big stem cell and regeneration initiative “Stem Cells Australia” which started in 2011 ($21 million). The funding will run out soon so hopefully another program is funded to continue to move our research forward, as there has been some exciting progress. We also have an annual meeting run by “Australasian Stem Society for Stem Cell Research” which is attended each year by Australian and also prominent international researchers each year. The International Society for Stem Cell Research (ISSCR) annual meeting is the largest in our field, and next year it will be in Melbourne Australia so hopefully there will be a strong Australian contingent at this meeting next year.
PDMS molds and poles used in the study, from Figure 1, Voges et al, 2017
And Holly, how did you come to join the lab?
HV I was lucky enough to join the lab as an undergraduate student when Enzo was setting up at UQ. I came across the lab profile online and thought his work was really interesting so I contacted him to see if I could do a 6 month project and here are we now, five years later!
Before you started your work, what was known about the regenerative capacity of human hearts of different ages?
EP, JH & HV Landmark papers from Enzo’s earlier work for the first time demonstrated that the regenerative capacity is lost during development in mammals. In contrast to the adult, newborn mice are capable of fully regenerating their hearts after injury (resection and myocardial infarction), a finding which is now widely supported by data from multiple labs. It has been known by clinicians for some time that in early life patients undergoing cardiac insults have better outcomes and heart recovery than adults. There has also been a recent case study demonstrating full recovery from myocardial infarction in a newborn. However, it was not fully understood whether this recovery/regeneration was due to cardiac regeneration or clinical management of these patients.
And what does organoid technology bring to the table for research into heart development and repair?
EP, JH & HV It provides us with an in vitro human model of cardiac regeneration we can use to complement our in vivo studies in mice. Mice and humans have many differences in their biological and physiological properties and models such as this give us an additional tool in our quest for regenerative therapeutics.
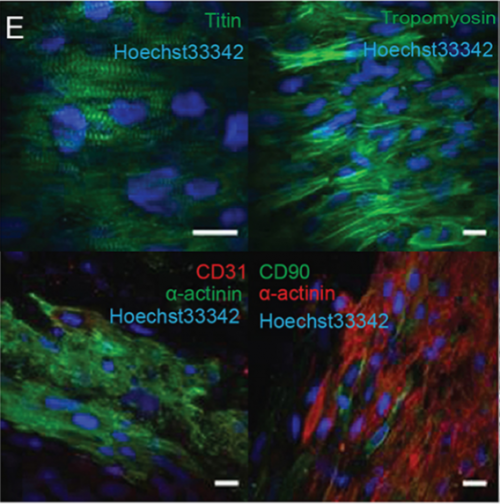
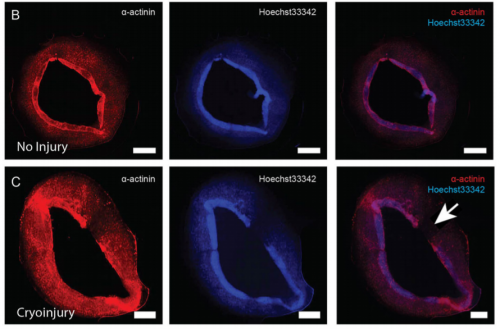
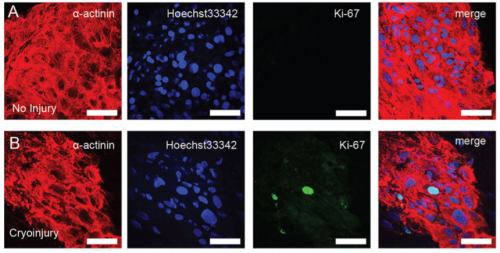
Morphological features of human cardiac organoids, from Figure 1, Voges et al. 2017.
Can you give us the key results of the paper in a paragraph?
EP, JH & HV Our hCO display properties and maturity consistent with fetal-like human heart tissue. We developed an injury model in this paper to determine how the hCO would respond to injury. In this study we chose to model disease using cryoinjury because it enables us to injure part of the tissue whilst keeping the remainder of the tissue healthy and viable. If we used ischemia-reperfusion or a chemical insult (eg. cardiotoxin) it would have affected the entire tissue. We then showed that the cryoinjured tissues have a high cell cycle activity and they can regenerate and recover their functionality following injury.
Do you have any idea why regenerative capacity is lost with heart maturation? Would more ‘mature’ organoids be poor regenerators?
EP, JH & HV In our on-going work in the hCO and in vivo in the mouse we are starting to understand more about the maturation process and how this regulates cardiomyocyte cell cycle and regeneration. Part of our work is finding the key drivers of maturation in hCO and whether mature hCO have the capacity to regeneration. We believe that more mature hCO will lose their regenerative potential.
Localized loss of α-actinin and nuclear staining in cryoinjured cardiac organoids, from Figure 2, Voges et al, 2017
Will your model have relevance for clinical avenues to cardiac repair in adults?
EP, JH & HV Our hope is to use this model to find key regulators of cardiac regeneration (together with our in vivo mouse models). Through these studies, hopefully we can find factors that can be used to facilitate cardiac regeneration in adult hearts and patients with cardiac disease.
And Holly, when doing the research, did you have any particular result or eureka moment that has stuck with you?
HV There were a number of really exciting moments along the way and it is great to be surrounded by people who share this excitement. One memory that stands out was when I finished collecting the organ bath data. This proved to be a time-consuming and painful process, so when I finally saw the exciting results I immediately ran down the hallway to show James and Enzo.
Expression of Ki-67+ cells in cryoinjured cardiac organoids, from Figure 3, Voges et al. 2017
And what about the flipside: any moments of frustration or despair?
HV I definitely had many moments of frustration throughout this project. Most of the time it felt like the moments of despair outweighed the eureka moments, but that only made the eureka moments more exciting.
What about your plans for the future following this paper?
HV I’m currently working on my next paper, which will enhance our hCO system to be a step closer to resembling native heart tissue by incorporating other endogenous cell types.
And what next for the lab?
EP & JH We are working together and also with academic and industry collaborators to try and study the cardiac regeneration process and the molecular mechanisms governing cardiomyocyte cell cycle at different stages of development. The holy grail is to find novel regenerative therapeutics for cardiac disease.
 (No Ratings Yet)
(No Ratings Yet)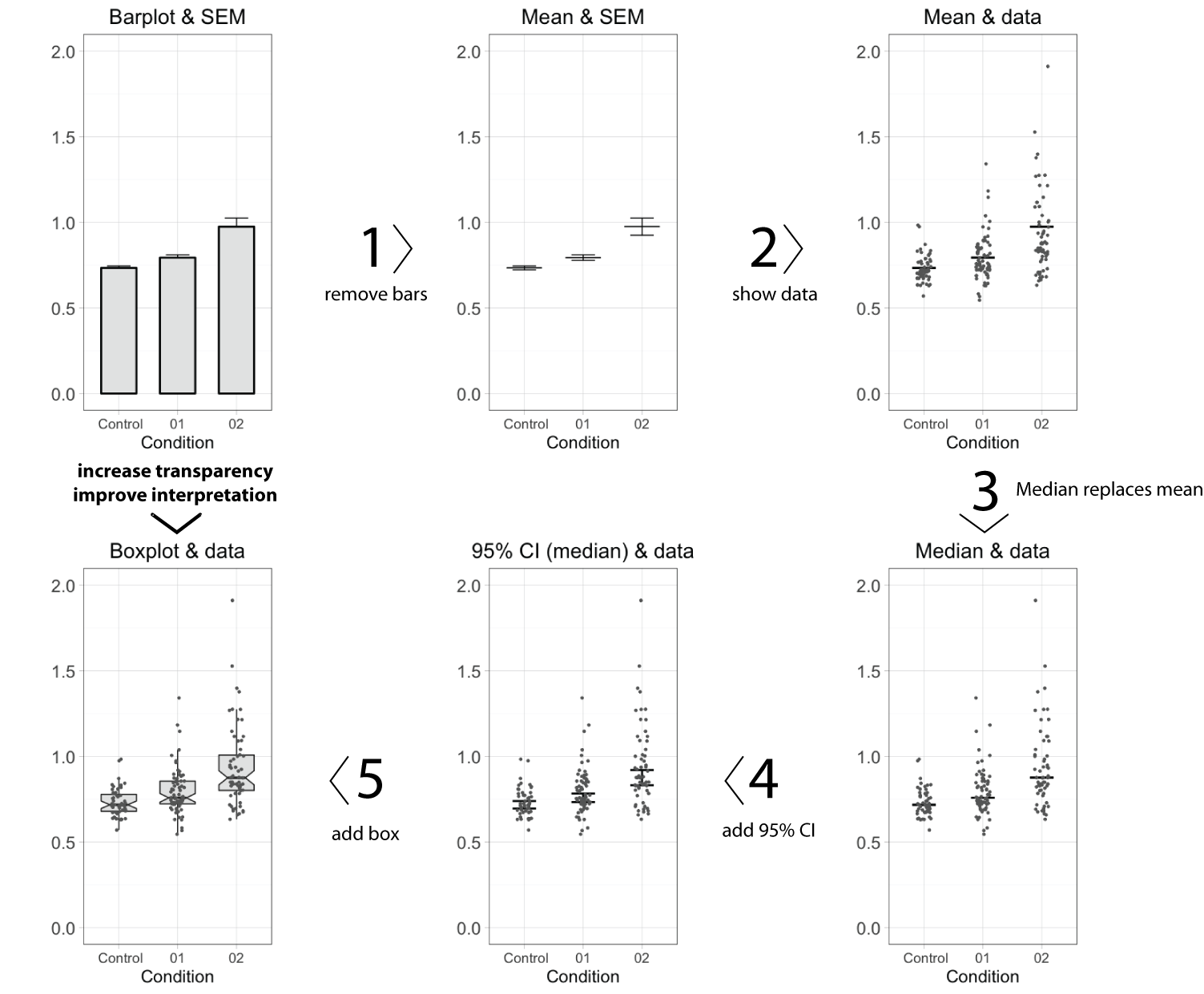
 (20 votes)
(20 votes)



 (1 votes)
(1 votes) Heather’s narrative:
Heather’s narrative: 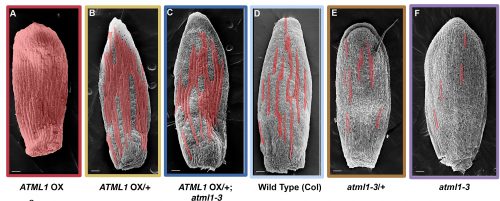
 José’s narrative:
José’s narrative: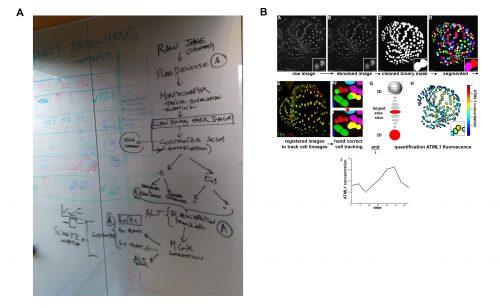
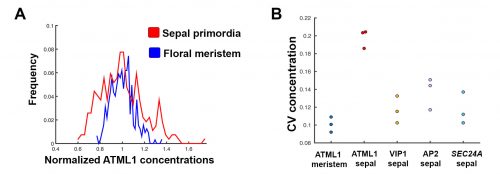
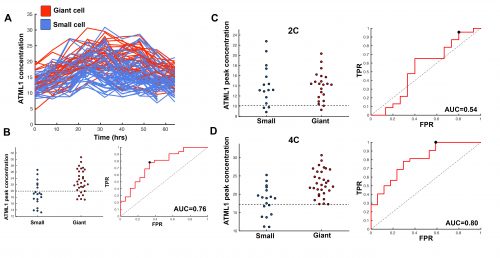
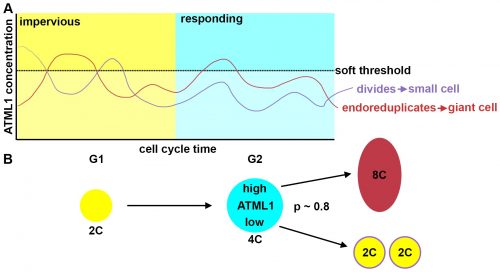
 Pau’s narrative:
Pau’s narrative: