The Company of Biologists is a not-for-profit publishing organisation dedicated to supporting and inspiring the biological community. We currently have two opportunities available in our Cambridge office.
Journal Website Content Manager
We are creating an exciting new role to enhance the community content on our journal websites.
We publish five important journals that serve the biological research community. All have effective publishing platforms and a good social media presence. We now seek to extend our community engagement, raise awareness of our charitable activities and build connections with early career scientists.
DMM is looking for an enthusiastic intern who wishes to gain experience in science publishing, including writing press releases, contributing to our social media activities, and supporting our Reviews Editor with commissioned articles. The internship is envisaged to last for 9 months at a salary of £20,000 per annum pro rata.
Our interns have a great track record of continuing on into important publishing roles.
Drosophila hematopoiesis shows striking resemblance with that of vertebrates, both at the level of signalling molecules and the phase of development. Even though there has been no report of Hematopoietic stem cells (HSCs) in Drosophila, this model has been employed extensively to understand progenitor biology and niche interactions. The Drosophila blood cells are specialised precursor cells (present within the hematopoietic organ: the lymph gland), that show several similarities with stem cells. They are known to differentiate into mature blood cells of the fruit fly. Our new study identifies the “founder cells” or the “stem cells” that give rise to these specialised blood-progenitors.
How it started off:
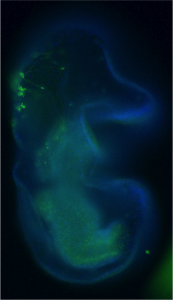
Our team was involved in detailed characterisation of signals which are required for the maintenance of the larval hematopoietic niche. To understand the dynamicity of the expression pattern of a battery of niche markers, we ventured into dissecting the larval lymph gland of the early first instar. Analysing a newly emerged larvae, about 8 hrs old, we encountered a group of large cells that always aligned themselves near the dorsal vessel or larval heart. We started calling them “Big Cells”. It turned out that these cells expressed several markers seen in other described stem cells.
Founder Cells or HSCs:
The very fact that they stood out almost every time in a early instar lymph gland caught our attention, and we felt tempted to characterise them. Our attempt to trace them throughout development revealed that although the “Big Cells” do not die, they could not be tracked beyond 22 hours post hatching. We argued that maybe they were changing their fate. Indeed, on lineage tracing of these cells and analysing them at various time point of development, we could see that they are actually converted to blood progenitor cells. Analysing the same genotype at mature third instar larval stages indicated that not only progenitors but also differentiated cells are lineage traced. This clearly indicates that the “Big Cells” are multi-potent in their nature. As a functional correlate, we genetically ablated the “Big Cells” and analysed the consequence on the mature lymph gland. Remarkably, the elimination of these 4-5 “Big Cells” drastically reduced the size of the gland.
The first instar larval lymph gland with the HSCs and its Niche marked by Dorothy-GFP expression
The final climax:
At this point we were thrilled that we had a fine story to tell about the elusive founder cells of Drosophila lymph gland. But a stem cell story without niche is not complete! We therefore entered into a spree to nail down the niche and the signal. Our suspicion was on the already described progenitor niche known to maintain the precursor cells. Since this niche is specified in embryo and is the most differentiated cell type in the first instar lymph gland, we speculated that it might also be serving as the niche for the HSCs. Our expression studies demonstrated that HSCs are enriched in phosphorylated Mad (pMAD), a pathway effector of Dpp signalling. To start with, therefore, we down regulated pMad from the HSCs. This resulted in their disappearance and a concomitant reduction in lymph gland size. Strikingly, on knock down of Dpp signalling from the progenitor niche, the HSCs failed to self-renew, clearly validating that Dpp signalling from the progenitor niche is its source of sustenance. It is to be noted that vertebrate early HSCs are also maintained by BMP (vertebrate counterpart of Dpp) signalling.
What are the implications:
With the identification of the early HSCs and the signal that maintains in Drosophila larvae, we have not only demonstrated the conservation with vertebrates, but put forward a model that can be used for answering several questions pertinent to vertebrate early blood development, both in normal as well as aberrant conditions.
Drevon, C., and T. Jaffredo. 2014. “Cell interactions and cell signaling during hematopoietic development.” Exp Cell Res 329 (2):200-6. doi: 10.1016/j.yexcr.2014.10.009.
Durand, C., C. Robin, K. Bollerot, M. H. Baron, K. Ottersbach, and E. Dzierzak. 2007. “Embryonic stromal clones reveal developmental regulators of definitive hematopoietic stem cells.” Proc Natl Acad Sci U S A 104 (52):20838-43. doi: 10.1073/pnas.0706923105.
Evans, C., Olson, J., Ngo, K., Kim, E., Lee, N., Kuoy, E., Patananan, A., Sitz, D., Tran, P., Do, M., Yackle, K., Cespedes, A., Hartenstein, V., Call, G., & Banerjee, U. (2009). G-TRACE: rapid Gal4-based cell lineage analysis in Drosophila Nature Methods, 6 (8), 603-605 DOI:10.1038/nmeth.1356
Evans, C.J., Sinenko, S.A., Mandal, L., Martinez-Agosto, J.A., Hartenstein, V., & Banerjee, U. (2007). Genetic Dissection of Hematopoiesis Using Drosophila as a Model SystemAdvances in Developmental Biology, 18, 259-299 DOI: 10.1016/S1574-3349(07)18011-X
Jung, S. H., C. J. Evans, C. Uemura, and U. Banerjee. 2005. “The Drosophila lymph gland as a developmental model of hematopoiesis.” Development 132 (11):2521-33. doi: 10.1242/dev.01837.
Mandal, L., Banerjee, U., & Hartenstein, V. (2004). Evidence for a fruit fly hemangioblast and similarities between lymph-gland hematopoiesis in fruit fly and mammal aorta-gonadal-mesonephros mesoderm Nature Genetics, 36 (9), 1019-1023 DOI: 10.1038/ng1404
Wellcome exists to improve health for everyone by helping great ideas to thrive. We’re a global charitable foundation, both politically and financially independent. We support scientists and researchers, take on big problems, fuel imaginations and spark debate.
We are seeking up to two Science Portfolio Advisers or Developers to contribute to managing the Cellular and Developmental Sciences portfolio within our Science division. You will liaise with internal and external stakeholders, including other funders, and provide our community of researchers with support and advice. You will need to keep abreast of the scientific field and attend key national and international conferences in the field.
To be successful in this role you will need to have a PhD and postdoctoral scientific experience in a relevant discipline. Some experience working outside academia in a research, funding or policy role is desirable although not necessary.
You should be confident interacting with your peers and the external scientific community. You will also be able to demonstrate that you can:
see the big picture and recognise scientific potential and opportunities
assimilate complex issues and work across science areas
lead and manage teams
communicate effectively and confidently with individuals and groups
work effectively and cooperatively within a team/matrix structure
apply your strong analytical and written skills to the development of briefing/position documents.
The salary will be between £36,000 and £52,000 plus benefits, depending on your experience.
To apply, please complete the online application process and ensure your covering letter explains how you meet the criteria for this job.
Very similar cell types are found across the animal tree of life. Much of animal diversity, therefore, arises not from the formation of new cells, but from the evolution of the developmental control of the numbers and patterns of orthologous cell types. Neural cells types are a particularly interesting example of this phenomenon. Almost all animals make serotonergic neurons, and for example, these neurons form at the anterior pole of many invertebrate larval types and in the anterior nervous systems of vertebrates. The numbers, distribution, and relative locations of serotonergic neurons are however quite varied. Thus animal gene regulatory networks (GRNs) must function to make this very conserved neural type, but be able to evolve the patterning and numbers of these cells.
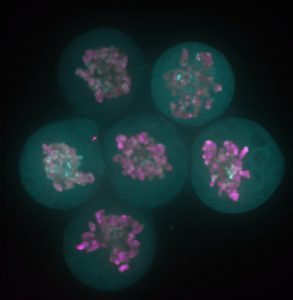
We have used the sea star, Patiria miniata, as a model system to ask how serotonergic neurons are made, and how they are positioned, so that we can understand this phenomenon. Sea stars are representatives of one class (Asteriodea) of the Phylum Echinodermata, which also include sea urchins (Cl. Echinoidea), and sea cucumbers (Cl. Holothuroidea). The sea star larvae, like the larvae of many invertebrate marine organisms has just a small number of serotonergic cells and these are readily visualized by immuno-localization using an antibody against serotonin (Fig 1).
Figure 1. Immuno-localization of serotonergic neurons in the sea star bipinnaria larva. Lateral view, mouth is to the right, anterior is up, and dorsal ganglion is top left.
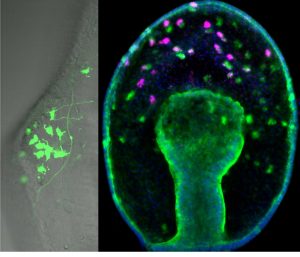
From previous work (Yankura et al., 2013) it was known that there are many soxc expressing cells scattered throughout the ectoderm in the two-day old gastrula that will form, not only the serotonergic neurons, but also other types of neurons in the later three to four-day larva. However, by the time neurons differentiate into various neural subtypes, the expression of soxc is extinguished. These cells were therefore labeled with a stable GFP construct to show that cells that originally express soxc do indeed end up forming serotonergic neurons (as well as other neural cell types) (Fig 2). Thus soxc+ cells in sea stars are neural progenitors, and these progenitors are distributed broadly across the ectoderm. The progression of these progenitors to differentiated neurons occurs through a series of asymmetric divisions (Fig 2). In particular one of the soxc+ daughters will express the LIM homeodomain transcription factor lhx2/9, and in turn lhx2/9 expressing cells will undergo both symmetric and asymmetric division, the asymmetric division producing a finally differentiated serotonergic neuron. Thus the lhx2/9 cells represent a proliferating, more restricted pool of serotonergic progenitors.
Figure 2. A stable GFP construct, driven by the soxc cis-regulatory region, is localised to neurons of the dorsal ganglion in the larval stage on left image. Right image: soxc expression cells (green) are found scattered throughout the ectoderm in the earlier gastrula. One of the soxc+ daughter cells expresses lhx2/9 (pink) in only the more anterior territory.
Next, the spatial domains in which these asymmetric divisions were occurring was examined. It has been known for many years that many animals, including the sea star larva (Yankura et al., 2010), share a remarkably conserved patterning along the AP axis. Genes with roles in patterning the anterior-most nervous system in vertebrates, for example, are also expressed in the most anterior regions of the larvae, and genes involved with patterning midbrain-hindbrain region are expressed at the posterior boundary of the ectoderm. This new work now shows that these AP domains control neural progression. That is, soxc+ cells divide to produce two soxc+ cells in the posterior zone, asymmetrically divide to produce an lhx2/9+ sister in the intermediate zone; lhx2/9+ cells will also divide symmetrically in this mid-zone, and will exit proliferation and become differentiated neuron in only the anterior-most, foxq2 expressing zone. These AP domains, therefore, establish neural proliferation zones.
Altering the size of the AP zones caused predicted changes in neural proliferation, and therefore changed the final numbers of serotonergic neurons. This shows therefore that a function of these highly conserved territories, in sea stars at least, is to regulate neural cell type progression.
This new work, therefore shows that neural progenitors form throughout the ectoderm without regard to patterning in the sea star, and that the GRN that establishes domains along the AP axis, controls neural progression. We predict that fairly simple evolutionary changes to this patterning GRN could change the size of these territories, and hence the numbers and distributions neural cell types.
REFERENCES
Yankura, K. A., Martik, M. L., Jennings, C. K. and Hinman, V. F. (2010). Uncoupling of complex regulatory patterning during evolution of larval development in echinoderms. BMC Biol8, 143.
Yankura, K. A., Koechlein, C. S., Cryan, A. F., Cheatle, A. and Hinman, V. F. (2013). Gene regulatory network for neurogenesis in a sea star embryo connects broad neural specification and localized patterning. Proc Natl Acad Sci U S A110, 8591–8596.
A postdoctoral position is available to study the cellular basis of morphogenesis in vertebrate craniofacial development. This work will integrate mouse molecular genetic and human iPSC approaches with live cell imaging, cell biology and biochemistry to study signaling mechanisms in development, and how this signaling goes wrong in congenital disease (e.g. PLos Biology 2015 13(4): e1002122, http://jcb.rupress.org/content/215/2/217.long, http://www.sciencedirect.com/science/article/pii/S0012160615301548). The position is in the laboratory of Jeff Bush (bush.ucsf.edu) in the UCSF Department of Cell and Tissue Biology and Program in Craniofacial Biology. The laboratory is located at the UCSF Parnassus Heights campus, in the center of San Francisco. UCSF offers an outstanding developmental biology community and a supportive working environment.
Candidates with a Ph.D. degree in a biological science and research experience in molecular biology, genetics, biochemistry, or live cell or live embryo imaging should submit a C.V. and names of at least 2 references via email to: jeffrey.bush@ucsf.edu
The Warwick – A*STAR Research Attachment Programme offers fully funded 4 year PhD studentships in Molecular Cell Biology and Microbiology, with students spending 2 years at Warwick and 2 years in Singapore at an A*STAR institute.
A strong 1st or an upper 2nd undergraduate degree (or equivalent international qualifications) in Physical Sciences (e.g. Engineering, Physics, Chemistry, Computer Science, Mathematics, Plant Sciences) or Life Sciences (e.g. Biology, Biochemistry, Biomedical Science).
Maths ‘A’ level or equivalent training is desirable.
Research training will be given in both Warwick and Singapore. Nonetheless, previous practical laboratory experience may be an advantage.
Application Procedure
To apply please send us:
a CV and covering letter
2 satisfactory academic references
Where appropriate, an English Language test certificate – acceptable tests can be found on the Study section of the Warwick website.
and also apply via the University’s online Postgraduate Application form HERE (please note that you are applying for course B93R PhD Molecular Biomedicine).
The story behind our recent paper (Festuccia et al., Nature Cell Biology 2016) started with a serendipitous observation made in a small room of the ISCR (Institute for Stem Cell Research), which at the time was still located in the King’s Buildings campus, south of town in my beloved Edinburgh. I was imaging a newly generated reporter ES cell line expressing a fusion between my favourite pluripotency transcription factor, Esrrb (Estrogen Related Receptor Beta), and a bright red fluorescent protein, TdTomato, when I noticed something I didn’t expect: I was able to identify dividing cells just by looking at the fluorescence signal from the Esrrb-TdTomato reporter. Esrrb was painting the mitotic chromosomes. I was surprised, and at the same time I realised I knew very little about the behaviour of transcription factors during cell division. Were all transcription factors supposed to remain bound to chromatin after condensation? And if so, with what consequences? Was this phenomenon reported in ES cells? What about other pluripotency factors?
Since then – I was finishing my PhD in the group of Ian Chambers – 4 years passed, and I learned that I was observing a phenomenon already described as “mitotic bookmarking”. Pioneering work over the last two decades in the groups of David Levens, Kenneth Zaret, Gary Stain, Marco Pontoglio and Gerd Blobel, among others, helped us understand that some proteins, including sequence specific transcription factors, remain bound to the chromatin during mitosis, marking promoters and key regulatory regions to facilitate the timely reactivation of gene expression after cell division. At the time of the observation, a friend and Post-Doc in Ian’s lab, Pablo Navarro, drove my attention to this concept. Pablo had already attempted to document bookmarking in ES cells. Unfortunately, regular fixatives such as formaldehyde delocalise transcription factors from mitotic chromatin (Pallier et al., Molecular Biology of the Cell 2003). The stimulating environment created by our usual scientific discussions made it possible for the implications of my initial observation not to remain unexplored. Thanks to the generosity of Ian, I was able to make of that observation the basis for my post-doc project, and I moved to the Pasteur Institute in Paris to join the newly established lab of Pablo. In these last 3 years, we were able to show that indeed during mitosis Esrrb remains bound to crucial enhancers regulating ES cell identity and instructs gene expression in daughter cells. We also understood that mitotic binding is very dynamic, and chiefly driven by the interaction of Esrrb with its consensus binding sites on DNA.
Nonetheless, many of the question I had at the time of the first observation of Esrrb mitotic retention remain open: the challenge that cell division, and another potentially disruptive phase of the cell cycle, DNA replication, pose to the control exerted by pluripotency factors over ES cell identity is a largely overlooked problem in our field. More generally, our understanding of the properties and structural organisation of mitotic, or newly replicated, chromatin is superficial. Can the observation made for Esrrb be extended to other pluripotency regulators? In the case mitotic binding will prove to be a general phenomenon, the current assumption of a restricted accessibility of mitotic chromatin will have to be challenged. Indeed, recent observations indicate that accessible genomic regions are largely invariant between interphase and mitosis (Hsiung et al., Genome Research, 2015). Coupled to the notion that a major portion of mitotic chromosomes is not constituted by chromatin (Booth et al., Molecular Cell 2016), this indicates that part of the DNA might reside outside domains formed of compacted nucleosomes in mitosis, possibly maintaining full or increased accessibility to transcriptional regulators, chromatin modifiers and remodelers. Or, vice-versa, we could imagine that precisely the binding of transcription factors keeps portions of the genome decompacted and localised in accessible regions of the chromatids.
If ES cells use specific mechanisms to ensure the continuity of transcription factor control over gene expression during cell division, and given that the activity of pluripotency regulators, like Esrrb, is tightly linked to the self-renewal ability of ES cells, could it be that the constraints posed by mitosis make it a favourable moment for ES cells to trigger differentiation programmes? Mitotic binding by Esrrb is restricted to only a fraction of the interphase targets. Declining levels of Esrrb and other factors might therefore trigger the further constriction of the set of accessible enhancers during cell division, irreversibly destabilising the pluripotency network in daughter cells. In fact, as part of my PhD research, I observed that Esrrb downregulation marks ES cell commitment to differentiation, and identified a set of enhancers that is inactivated during the early stages of this process (these results form the basis of a manuscript currently under revision). Many of these are indeed bookmarked enhancers.
I might just find myself in the lucky position of being able to link two parts of my previous work and contribute to our better understanding of how cell fate decisions are implemented by pluripotent cells early during development, and possibly later on in multipotent progenitors and adult stem cells.
New postdoc positions are available immediately in the laboratory of Dr. Terry Yamaguchi at the National Cancer Institute at Frederick. The group investigates how Wnt signals regulate the gene regulatory networks that control the fate of embryonic and adult stem cells. Current projects focus on understanding how Wnts regulate the self-renewal and differentiation of a recently described bipotent progenitor, the neuromesodermal progenitor (NMP), which generates the spinal cord neurons and musculoskeletal system of the trunk. Using a combination of mouse genetics, mouse and human embryonic stem cell in vitro differentiation, and genomic and proteomic approaches, Dr. Yamaguchi’s lab is investigating the molecular mechanisms underlying the activity of key transcription factors in NMP development, as well as in rare cancers such as chordoma. Please view the lab’s website for further details and a list of relevant publications at: https://ccr.cancer.gov/Cancer-and-Developmental-Biology-Laboratory/terry-p-yamaguchi
Number of Positions: 2
C.V. Required: Yes
Number of References Required: 3
Bibliography Required: Yes
Experience Required:
Interested candidates must have a Ph.D. and/or an M.D. and have less than 5 years of postdoctoral experience. Applicants that have a strong background in stem cell biology, molecular biology, embryology, genetics, functional genomics or a related discipline are encouraged to apply.
How To Apply: To apply, send a cover letter, CV including bibliography, and contact information for three references to the following address:
Dr. Terry Yamaguchi, Ph.D., Senior Investigator, Cell Signaling in Vertebrate Development Section, Center for Cancer Research, National Cancer Institute, Building 539, Room 218/205, Frederick, MD 21702-1201.
The aim of the third conference in this series is to understand the biology that underpins the success or failure of regenerative processes. We will explore the relationship between stem cell biology and regenerative biology so that both can be fully exploited to treat disease. This meeting will bring together scientists and clinicians interested in developmental and regenerative biology, stem cell research, gene therapy, tissue engineering and translational medicine. We welcome abstracts from all areas relevant to the main themes of the meeting, for both oral and poster presentations. Several oral presentations will be chosen from the abstracts submitted. A limited number of bursaries will be available for PhD students! Registration will open soon – for further details, visit: https://coursesandconferences.wellcomegenomecampus.org/events/item.aspx?e=633
 (No Ratings Yet)
(No Ratings Yet)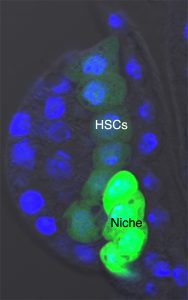
 (5 votes)
(5 votes)