In 2016 the BSCB/BSDB Spring conference has yet again been a great success, and many prizes were announced, amongst them the BSDB’s Waddington, BSCB’s Hooke, BSCB’s Women in Cell Biology Early Career Award, BSDB’s Cheryll Tickle and BSDB’s Beddington medals. See below movies of the first four of the five medal talks and find more information about all awardees of the conference in a separate blog. In addition, watch a short movie of the Uri Alon special who gave an entertaining spiel about the challenges for creative science, its opportunities, pitfalls and how to get out of the CLOUD, accompanied by his amusing but all so true songs.
All films were produced by Warwick Conferences, commissioned by the BSDB/BSCB Spring Meeting organisers.
It was on the 7th of February of 2016 when 20 leading scientists from all over the world headed to the historic Wiston House in West Sussex, England, to spend four days in focused atmosphere discussing new insights in cardiovascular research: the workshop for Transdifferentiation and Tissue Plasticity in Cardiovascular Rejuvenation. Supported by the Company of Biologists, Brian Black and Jim Martin brought together experts of the field of heart development, regeneration and tissue engineering with the aim to discuss new approaches and recent findings to improve cardiac repair. In addition to the 20 senior investigators, also 10 early-stage scientists (PhD students, postdocs, and junior PIs) were selected to participate. For me in particular, with my PhD recently completed, joining this event was a great honour and a unique opportunity. I excitedly anticipated the four intense workshop days with great interest.
by Christian Mosimann
With the heart being such a complex and specialized tissue, repair of cardiac muscle in human is difficult to achieve. In the workshop, we were introduced to and discussed a wide range of ongoing efforts, including genomic regulation of cardiomyocyte differentiation, environmental factors involved in cardiac regeneration, as well as the contribution of different cell types in this process. Although I cannot allude to all the talks here, all participants agreed about the outstanding scientific quality of the work presented in this meeting.
Liz Robertson opened the first session of the workshop on Sunday afternoon recapitulating the origin of cardiac progenitors during embryogenesis and explaining the involvement of Eomesodermin in cardiac mesoderm specification in the mouse embryo (Costello et al., 2011). To understand myocardial differentiation, it is fundamental to define transcription factors and epigenetic modifications that specify cardiac lineages. Benoit Bruneau introduced his lab’s latest study on the coordination of three cardiac transcription factors (Nkx2.5, Tbx5 and Gata4) in the regulation of cardiac gene expression and differentiation (Luna-Zurita et al., 2016). Laurent Dupays described the interaction of the two transcription factors Meis and Nkx2.5 on a specific enhancer sequence (Dupays et al., 2015). Moreover, Brian Black explained new findings from his group about the Mef2c transcriptional regulation machinery in cardiomyocytes.
We also learned about specific approaches to elucidate the functions of distinct cellular factors active in cardiomyocytes. Guo Huang is currently investigating fetal cardiac genes, which reactivate cell cycle re-entry of adult heart muscle cells for potential regenerative repair after myocardial infarction. Jim Martin reported the implication of the Hippo pathway in cytoskeletal remodelling of cardiomyocytes in the injured heart (Morikawa et al., 2015). Finally, Kathy Ivey introduced how to study human iPSC-derived cardiomyocytes to better understand protein signalling and interaction networks as well DNA-occupancy in cellular differentiation.
Another focus during this workshop was the understanding of environmental factors, which impact cardiomyocyte behaviour and fate during cardiac repair. We learned from Eldad Tzahor how the stiffness of the extracellular matrix affects the differentiation state of cardiomyocytes (Yahalom-Ronen et al., 2015). Ahmed Mahmoud described the implication of cardiac innervation and Neuregulin signalling in the regulation of cardiomyocyte proliferation in the regenerating neonatal mouse and zebrafish heart (Mahmoud et al., 2015). The fact that many different cell types are crucial for cardiac regeneration was demonstrated by Paul Riley and Nadia Rosenthal. Nadia nicely illustrated the cellular composition of the heart and discussed recent work that demonstrates the fundamental role of macrophages during cardiac repair and regeneration (Pinto et al., 2016), while Paul Riley explained the different origins and the development of lymphatic vessels in the heart and described how this developmental program is reactivated after myocardial infarction (Klotz et al., 2015). Another approach was described by Enzo Porrello, who is seeking to understand the differences between cardiac cells at different stages of life, to unveil the mechanisms that impede adult cardiac regeneration.
Multiple talks presented studies using the zebrafish, an important model of cardiac development and regeneration due to its remarkable regenerative capacity and its transparency during embryogenesis. Didier Stainier illustrated how cardiomyocytes delaminate from the compact layer to form trabeculated myocardium in the zebrafish embryo (Staudt et al., 2014). Karina Yaniv showed beautiful movies displaying the origin of lymphatic vessels during development (Nicenboim et al., 2015). Christian Mosimann presented live imaging that traces back the cardiovascular lineages to the lateral plate mesoderm (Mosimann et al., 2015) and explained what we could learn about cardiac diseases by modelling human patient mutations in zebrafish. Ken Poss, whose lab is interested in the mechanisms of heart and fin regeneration, guided us through his journey in searching for regenerative cellular programs in the zebrafish (Kang et al., 2016). Further, Nadia Mercader spoke about distinct populations of cardiomyocytes in the early and the adult zebrafish heart and injury studies to decipher the mechanisms of myocardial regeneration.
by Christian Mosimann
Another key topic discussed in this workshop was how we can investigate the underlying causes of human cardiac diseases in more depth. Alessandra Moretti showed one example of how her lab studies the cause of Arrhythmogenic right ventricular dysplasia using patient-derived iPSCs. Deepak Srivastava further reported new findings about the molecular consequences of human GATA4 mutations obtained by studying iPSCs-derived cardiomyocytes from patients. An impressive finding was reported by Eric Olson: he explained how his lab achieved the repair of mutations in the Dystrophin gene, the cause of Duchenne muscular dystrophy, by CRISPR/Cas 9 technology in mice in vivo(Long et al., 2016).
I was personally very intrigued to hear about the work of the bio-engineers participating in this workshop. Nenac Bursac illustrated how his lab gains insights into cardiomyocyte functions by using in vivo assays of cardiomyocyte patches. Moreover, from Molly Stevens we learned about the versatile use of nano needles, which can deliver substances to cells or even make measurements (Chiappini et al., 2015).
In my opinion, and I am sure all participants would agree, this workshop was a tremendous success. Fascinating data, of high scientific value were presented and openly discussed. For me, this workshop was a unique experience; I met experts in the field of cardiovascular research and learned about a vast range of experimental approaches. Moreover I had the opportunity to present and discuss our latest results on the endocardial dynamics in zebrafish heart regeneration. Finally the guided tour through the amazing 16th century historical Wiston House, the windy walk in the beautiful Sussex countryside, and the experience of watching the Super Bowl for the first time, completed the great experience of the Transdifferentiation and Tissue Plasticity in Cardiovascular Rejuvenation workshop.
Here is a short video put together by the Company of Biologists on this workshop:
I am grateful to Christian Mosimann for comments on the text.
References
Chiappini, C., De Rosa, E., Martinez, J. O., Liu, X., Steele, J., Stevens, M. M. and Tasciotti, E. (2015). Biodegradable silicon nanoneedles delivering nucleic acids intracellularly induce localized in vivo neovascularization. Nature materials14, 532-539.
Costello, I., Pimeisl, I. M., Drager, S., Bikoff, E. K., Robertson, E. J. and Arnold, S. J. (2011). The T-box transcription factor Eomesodermin acts upstream of Mesp1 to specify cardiac mesoderm during mouse gastrulation. Nature cell biology13, 1084-1091.
Dupays, L., Shang, C., Wilson, R., Kotecha, S., Wood, S., Towers, N. and Mohun, T. (2015). Sequential Binding of MEIS1 and NKX2-5 on the Popdc2 Gene: A Mechanism for Spatiotemporal Regulation of Enhancers during Cardiogenesis. Cell reports13, 183-195.
Kang, J., Hu, J., Karra, R., Dickson, A. L., Tornini, V. A., Nachtrab, G., Gemberling, M., Goldman, J. A., Black, B. L. and Poss, K. D. (2016). Modulation of tissue repair by regeneration enhancer elements. Nature.
Klotz, L., Norman, S., Vieira, J. M., Masters, M., Rohling, M., Dube, K. N., Bollini, S., Matsuzaki, F., Carr, C. A. and Riley, P. R. (2015). Cardiac lymphatics are heterogeneous in origin and respond to injury. Nature522, 62-67.
Long, C., Amoasii, L., Mireault, A. A., McAnally, J. R., Li, H., Sanchez-Ortiz, E., Bhattacharyya, S., Shelton, J. M., Bassel-Duby, R. and Olson, E. N. (2016). Postnatal genome editing partially restores dystrophin expression in a mouse model of muscular dystrophy. Science351, 400-403.
Luna-Zurita, L., Stirnimann, C. U., Glatt, S., Kaynak, B. L., Thomas, S., Baudin, F., Samee, M. A., He, D., Small, E. M., Mileikovsky, M. et al. (2016). Complex Interdependence Regulates Heterotypic Transcription Factor Distribution and Coordinates Cardiogenesis. Cell164, 999-1014.
Mahmoud, A. I., O’Meara, C. C., Gemberling, M., Zhao, L., Bryant, D. M., Zheng, R., Gannon, J. B., Cai, L., Choi, W. Y., Egnaczyk, G. F. et al. (2015). Nerves Regulate Cardiomyocyte Proliferation and Heart Regeneration. Developmental cell34, 387-399.
Morikawa, Y., Zhang, M., Heallen, T., Leach, J., Tao, G., Xiao, Y., Bai, Y., Li, W., Willerson, J. T. and Martin, J. F. (2015). Actin cytoskeletal remodeling with protrusion formation is essential for heart regeneration in Hippo-deficient mice. Science signaling8, ra41.
Mosimann, C., Panakova, D., Werdich, A. A., Musso, G., Burger, A., Lawson, K. L., Carr, L. A., Nevis, K. R., Sabeh, M. K., Zhou, Y. et al. (2015). Chamber identity programs drive early functional partitioning of the heart. Nature communications6, 8146.
Nicenboim, J., Malkinson, G., Lupo, T., Asaf, L., Sela, Y., Mayseless, O., Gibbs-Bar, L., Senderovich, N., Hashimshony, T., Shin, M. et al. (2015). Lymphatic vessels arise from specialized angioblasts within a venous niche. Nature522, 56-61.
Pinto, A. R., Ilinykh, A., Ivey, M. J., Kuwabara, J. T., D’Antoni, M. L., Debuque, R., Chandran, A., Wang, L., Arora, K., Rosenthal, N. A. et al. (2016). Revisiting Cardiac Cellular Composition. Circulation research118, 400-409.
Staudt, D. W., Liu, J., Thorn, K. S., Stuurman, N., Liebling, M. and Stainier, D. Y. (2014). High-resolution imaging of cardiomyocyte behavior reveals two distinct steps in ventricular trabeculation. Development141, 585-593.
Yahalom-Ronen, Y., Rajchman, D., Sarig, R., Geiger, B. and Tzahor, E. (2015). Reduced matrix rigidity promotes neonatal cardiomyocyte dedifferentiation, proliferation and clonal expansion. eLife4.
Every year, the British Society for Developmental Biology (BSDB) awards the Beddington Medal to the best PhD thesis in developmental biology. The 2016 award went to Elena Scarpa, who did her PhD with Roberto Mayor at University College London (UCL). We caught up with Elena at the BSCB/BSDB Spring meeting, and we asked her about her thesis work on the neural crest and what she is doing now.
Congratulations on winning the Beddington Medal. What does this prize mean to you?
I am very happy about this prize. I moved from Italy to the UK to do my PhD, which was a big change for me. I knew my science background was good, but at UCL I was surrounded by people who had come from excellent universities, like Cambridge or UCL, and my university in Italy was not recognised in the same way. Some of the other students in my PhD course were foreigners as well, but they had studied in the UK. So receiving this medal is a big achievement.
This prize also recognises the communal effort that went into this project. It was a lot of work, not just from me but also from the people I collaborated with and helped me. I think this prize also reflects well on Roberto. I am the second student from his lab that wins it, showing that his students are doing well and that he is a good supervisor.
Can you tell us a bit more about the lab where you did your PhD?
I did my PhD in Roberto Mayor’s lab at UCL. The lab works on neural crest, mostly in Xenopus but also in zebrafish. The lab started out by working on induction and specification of the neural crest, but in the last few years the majority of the lab has worked on different aspects of neural crest migration. The lab had a very nice environment. There were many students, so it was a lot of fun.
It seems that contact inhibition is on a winning streak, since last year’s winner of the Beddington Medal, John Robert Davis, was also working on contact inhibition. What is contact inhibition and in what contexts is it important?
Contact inhibition is the process by which two migrating cells interact and change their direction of motion after contact. This phenomenon was discovered by Abercrombie at UCL. Roberto likes to say that Abercrombie was based in the same room as the Mayor lab, but I am not sure this is true! Abercrombie and his colleague Heaysman observed that contact inhibition was related with the ability of malignant cells to invade other tissues. Their work was very nice but descriptive, and contact inhibition and its molecular mechanisms did not receive much attention for several years.
In 2008 Roberto’s lab published a paper about contact inhibition of locomotion, showing that this process is required for neural crest directionality, and hinting at the molecular mechanism behind it. It showed that cell-cell contacts, planar cell polarity and cadherins are required. However, what I think is interesting about contact inhibition is that it mediates many different types of cell behaviour during development. In the neural crest it mediates collective behaviour (in combination with other processes). However, as John showed in hemocytes, and also in neurons, it can mediate dispersion. In addition, it can be used by certain cancer cells as a driving force for invasion. So the presence or lack of contact inhibition can really change how cells interact with each other.
What was your thesis project about, and what were your major findings?
Contact inhibition is the process by which cells separate, but many other cells types, such as epithelial cells, stay together and make stable contacts. We wanted to understand better the nature of mesenchymal cell-cell interactions. Are mesenchymal cell-cell interactions like contact inhibition intrinsically different because cells are unable to form a junction, or is there something in the junction itself that changes the behaviour of the cells?
My main finding was that, at least in the neural crest, cells that undergo contact inhibition do not express different cadherins. The ability of cadherins to recruit the cadherin complex or the actin complex is very similar, but there is a difference in their ability to polarise the activity of small GTPases, polarising the motility of the cells. This adds to what was already previously known. In the neural crest and in cancer it is well known that E-cadherin is down regulated, and this seems to correlate with the ability of the cells to invade. However, the idea that loss of contact allows invasion does not really make sense. Cells that are unable to make contacts with other cells cannot interact properly with their environment, so in vivo this does not favour migration. Our paper showed that there are other changes in the way cells form protrusions and interact with their environment that lead to scattering and active migration, rather than just the cell-cell contact itself. Other papers had proposed this idea before, but we were able to put it in a developmental framework.
You mentioned collaborators. Which labs did you collaborate with?
Within the lab I originally worked with Eric Theveneau, and also with András Szabó who is a modeller and helped me with the quantitative analysis of traction forces.
During my project we used FRET to obtain more information about the dynamics of how contact inhibition is regulated. For this I collaborated with Maddy Parsons at King’s College London, who had already collaborated with Roberto on other contact inhibition papers. I worked with Maddy a lot, and it took a lot of effort in the first two years of my PhD to get the FRET to work. It is very nice to collaborate with Maddy because she is very available, and happy to try different things.
I also established a collaboration for the optogenetics part of the project, since were not able to get it to work at UCL. During my PhD Eric moved to Toulouse, where Xiabo Wang , who previously adapted photoactivation in vivo during his postdoc in Denise Montell’s lab and used a photo-activatable form of Rac. He was familiar with the imaging conditions, so I did my experiments in their microscope in Toulouse. This was a great opportunity to learn a new technique, and I am going to develop optogenetics further in my postdoc. So this collaboration also helped me to define my interests.
You have moved from London to Cambridge for your postdoc. What are you working on now and how did you choose your new lab?
I now work in the lab of Benedicte Sanson at the department of Physiology, Development and Neuroscience. The lab studies the Drosophila embryo at the global level, examining how extrinsic forces generate collective rearrangements in the tissue. They focus mostly on germ band extension and parasegment boundary formation. My project is concerned with oriented cell division in the early Drosophila embryo. I am looking at how mechanical cues influence the orientation of cell division.
During my PhD I looked at cell migration, and how the traction on the substrate relates with the tension mediated by cell-cell interactions. For my postdoc I wanted to develop more my knowledge of tissue mechanics. The Sanson lab is a really good place to develop this interest because the Drosophila embryo is a really powerful system. It is very simple and very easy to access. They also have a great ongoing collaboration with Guy Blanchard from Richard Adam’s lab. Their tracking software is very powerful and allows very fine image analysis. You can track mesenchymal cells like the ones I studied during my PhD, but it is never very refined because it is very difficult to segment the cells. In epithelial cells this type of analysis is possible.
Did you find it difficult to change model organism?
It is different. In the beginning working with Drosophila didn’t make sense to me. I am used to Xenopus and microinjections. When you come into the lab on a Monday you don’t know exactly what will you do, and choose your experiment based on the quality of the embryos. Because you just microinject it isn’t necessary to plan ahead, but on the other hand you can’t do clean genetics. In Drosophila you have very powerful genetic tools which are very useful. However, you need to plan your experiments. You need to cross the flies, wait, then select them… It just didn’t make sense to me why I had to wait 2 weeks to see the membrane! But you get used to it.
Do you have any advice for new students?
Choose something that you really like and that really motivates you. You are going to spend a lot of time in the lab trying to solve problems, so you need a good question that you are passionate about. Also try to choose a good environment, where you will be supported and have interesting discussions. Finally, be creative. Sometimes you get stuck, and if you tried everything and it doesn’t work then you need to think outside the box. Be a bit brave and take some risks.
Research Group: James Wakefield (www.thewakefieldlab.com)
Salary scale: £19,828 – £25,023
Duration: 1st June 2016 – 31st August 2018 (with possible 3 yr renewal)
We wish to recruit a Research Technician to support the work of Dr James Wakefield. The successful applicant will assist Dr Wakefield in the day-to-day management of the lab and work on an interdisciplinary project aimed at understanding the fundamental process of mitotic spindle formation in the model organism, Drosophila melanogaster. As such, this position represents an opportunity to work in a creative, collegiate and interdisciplinary research environment, making an essential contribution to an internationally-leading research programme.
The successful applicant will support a wide range of research activities, including supporting and training undergraduate and post-graduate research students, working closely with the PI to organise and co-ordinate aspects of lab-management, overseeing the culture and maintenance of Drosophilalaboratory stocks and undertaking their own independent research project. They will be enthusiastic, highly motivated and possess excellent verbal and written communication skills.
Applicants will possess a relevant first degree (BSc Honours) in Biological Sciences, Biochemistry, Genetics or a related subject and demonstrate sufficient knowledge of research methods and techniques to work within the established research programme. Applicants will be able to demonstrate skills in genetics, biochemical techniques, microscopy and cell biology. Previous experience working with Drosophila would be a distinct advantage.
A postdoctoral position is available to study the developmental mechanisms that pattern differentiation during organ development.
Epithelial tubes often have a functional polarity written along their P-D axis, with specialised segments carrying out distinct physiological activities. With a handful of notable exceptions, we know little about how P-D axes and segment-specific differentiation are regulated during organogenesis.
We aim to understand the molecular and cellular mechanisms that pattern and maintain functional polarity along the P-D axis in a structurally simple, but functionally sophisticated epithelial tube: the Drosophila renal tubule.
Highly motivated applicants with a PhD and strong background in cell/developmental biology are encouraged to apply.
For informal inquiries about the position please contact Barry Denholm directly: Barry.Denholm@ed.ac.uk
The origin of paired fins is a major unresolved issue in vertebrate evolutionary biology, and has been a topic of debate among palaeontologists, comparative anatomists and developmental biologists for over a century. Central to any question of “evolutionary origins” is the concept of homology: the sharing of features due to common ancestry. Homology may explain the existence of shared features between organisms (historical homology – e.g. the arms of a chimpanzee and the arms of a human are homologous because these structures have been retained from a common ancestor that possessed arms), or the existence of shared features within an organism (serial homology – e.g. the vertebral elements within a human, which exhibit a range of morphologies, but which nevertheless share a common underlying ground plan) (Roth, 1984; Wagner, 1989). In either case, homology reflects a continuity of anatomical, cellular, or genetic information, and provides a useful conceptual framework for investigating the evolutionary relationship among body plan features in distantly related taxa (Van Valen, 1982).
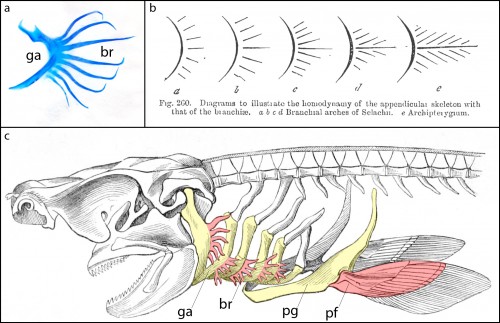
Cartilaginous fishes (sharks, skates, rays and holocephalans) are unique among living jawed vertebrates, in that they possess a series of skeletal appendages called branchial rays that project laterally from their gill arches (Fig. 1a). These branchial rays articulate with the gill arch cartilages in a way that is broadly reminiscent of the articulation between pectoral fins or limbs and the shoulder girdle, and this similarity led the comparative anatomist Carl Gegenbaur to propose that paired fins and limbs evolved by transformation of a gill arch (Gegenbaur, 1878) (Fig. 1b, c) – a hypothesis of serial homology that remains controversial to this day. Unfortunately, the fossil record currently tells us relatively little about the stepwise acquisition of paired fins during vertebrate evolution, so we decided to address this question from a developmental perspective. We were interested in determining whether the anatomical parallels that Gegenbaur noted between the gill arches of cartilaginous fishes and fins/limbs may reflect common underlying molecular patterning mechanisms in these organ systems. To this end, we conducted a series of experiments to investigate branchial ray patterning in embryos of an oviparous (egg-laying) cartilaginous fish, the little skate, Leucoraja erinacea (see video below for an overview of skate embryonic development).
Figure 1. a. Skeletal preparation of an embryonic shark gill skeleton, showing branchial rays (br) projecting from the gill arch (ga). b. Gegenbaur’s “Archipterygium” hypothesis, illustrating the hypothetical transformation of a gill arch into a fin (from Gegenbaur, 1878). c. A shark head skeleton illustrating putative serial homology of the gill arch and pectoral fin skeleton. Gill arches (ga) and the pectoral girdle (pg) are coloured yellow; branchial rays (br) and the pectoral fin (pf) are coloured red (modified from Owen, 1866).
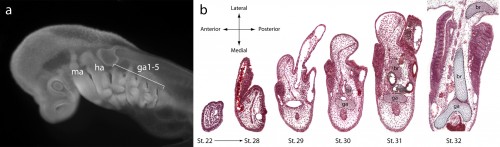
It might be helpful to first briefly introduce the gill arch, and how the gill arches of cartilaginous fishes are unique relative to those of other jawed vertebrates. All vertebrate embryos possess a bilateral series of pharyngeal arches on the sides of their developing head (Fig. 2a), and the mesenchyme within these arches gives rise to much of the craniofacial skeleton. The first pharyngeal arch is called the mandibular arch, the second is called the hyoid arch, and this is followed by a variable number of gill arches (most cartilaginous fishes have five gill arches). Primitively, the mandibular arch gave rise to the jaw skeleton, the hyoid arch gave rise to the skeletal apparatus that suspends the jaw from the braincase, and the gill arches give rise to the skeletal supports of the respiratory apparatus of the gills. These skeletal derivatives are conserved in a relatively primitive organisation in cartilaginous fishes, but are also present in other vertebrates (though often in a derived state – for example, the skeletal derivatives of the “gill arches” of mammals give rise to the laryngeal skeleton). Uniquely, though, in cartilaginous fishes, once the hyoid and gill arches have formed, they undergo a lateral expansion (Fig. 2b), and give rise to an additional set of skeletal elements – the branchial rays – that project laterally from the arches (Gillis et al., 2009). These branchial rays ultimately provide skeletal support to the fleshy flaps that protect the gills of cartilaginous fishes. As appendages, branchial rays must be patterned along the proximodistal axis (as they expand laterally) and also along the anterior-posterior axis (branchial rays exhibit a pronounced anterior-posterior polarity, and articulate proximally along the posterior margin of the hyoid and gill arch cartilages). In our paper (Gillis and Hall, 2016), we were interested in testing whether skate gill arches deploy similar axial patterning mechanisms as do fins and limbs (as would be predicted by an hypothesis of gill arch-fin/limb serial homology).
a. The head of a skate embryo (anterior to the left), showing the position of the mandibular (ma), hyoid (ha) and gill arches (ga1-5). b. Histological sections through a developmental series of skate gill arches, illustrating the lateral expansion of the arch, and the condensation and differentiation of the gill arch (ga) and branchial ray (br) cartilages.
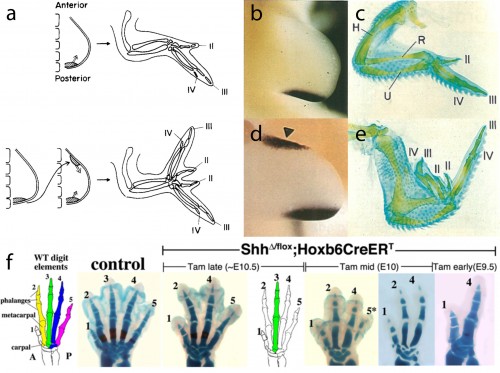
In this study, we focused our attention on the sonic hedgehog (Shh) signaling pathway, as the axial patterning function of this pathway during limb development is very well understood. Decades of experimental embryological and molecular investigation have revealed that Shh signaling is important in both anterior-posterior patterning of the limb, and in the proliferative expansion of limb skeletal progenitors. Classical chick embryo experiments by Saunders and Gasseling (1968) demonstrated that upon transplantation of donor posterior limb bud mesenchyme to the anterior region of a host limb bud, the resulting limb will form an additional set of digits that are oriented mirror-image to the normal digits (Fig. 3a). This posterior limb bud signaling centre is known as the zone of polarizing activity (ZPA). Twenty five years later, the Tabin lab demonstrated that Shh was the polarizing signal that was being secreted by the ZPA (Fig. 3b,c), and that misexpression of Shh in the anterior limb bud was sufficient to induce the ectopic, mirror-image digits noted by Saunders and Gasseling (Fib. 3d,e) (Riddle et al., 1993). In addition to this anterior-posterior patterning role, Shh signaling is also required for the expansion of limb endoskeletal progenitors, and progressively earlier loss of Shh signaling during limb development results in a progressively more profound deletions of distal limb skeletal elements (as shown, for example, by the genetic deletion of Shh from mouse limb buds, or by pharmacological manipulation of hedgehog signaling in chick and salamander) (Fig. 3f) (Towers et al., 2008; Zhu et al., 2008; Stopper and Wagner, 2007). Does Shh signaling function in a similar manner during the development of skate branchial rays?
Figure 3. a. Saunders and Gasseling demonstrated in the chick embryo that grafts of posterior limb bud mesenchyme to the anterior of a host limb bud resulted in the formation of ectopic, mirror-image digits. b. It was later determined that Shh signalling from posterior limb bud mesenchyme was the molecular effector of the zone of polarising activity, c. and was require for the development of a normal complement of digits. d. Ectopic Shh expression in the anterior limb bud mesenchyme induces e. ectopic, mirror-image digits. f. Shh signalling is also required for the proliferation of limb endoskeletal progenitor cells, and progressively earlier deletion of limb bud Shh results in a progressively greater reduction in the distal limb endoskeleton. a.–e. Images modified from Riddle et al., (1993) Sonic hedgehog mediates the polarising activity of the ZPA. Cell 75: 1401-1416 (http://www.sciencedirect.com/science/journal/00928674). Copyright Elsevier, 1993. f. Modified from Zhu et al. (2008).
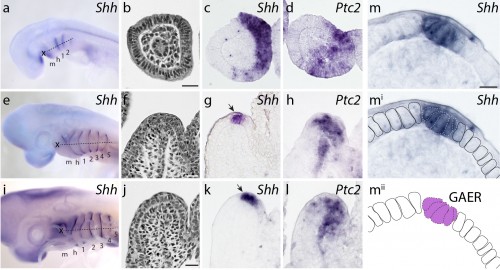
We first sought to determine whether Shh signaling components were expressed during skate gill arch development. mRNA in situ hybridization experiments revealed that, indeed, Shh is expressed in a polarized pattern during the development of the skate hyoid and gill arches – initially in the posterior epithelium of each arch, and eventually in an epithelial stripe along the leading edge of the hyoid and gill arches as they undergo lateral expansion (Fig. 4). When we looked at the expression of Ptc2 (a transcriptional readout of Shh signaling), to determine which tissues are responding to this Shh signal, we observed expression in both the distal gill arch epithelium, and in the mesenchyme beneath the Shh expression domain (Fig. 4). One key difference, of course, is that in skate gill arches, Shh is expressed in posterior-distal epithelium, while in the limb bud, Shh is expressed in posterior mesenchyme. However, in both cases, Shh signal is transduced in overlying epithelium and distal mesenchyme (so although the source of the signal is different between these appendages, the responding tissues are similar).
Figure 4. a. At stage 22, b.–c.Shh is expressed in the developing gill arches, with transcripts localizing to posterior arch epithelium. d.Ptc2 expression indicates that this signal is transduced in posterior gill arch mesenchyme, epithelium, and core mesoderm. e. By stage 27, f.–g.Shh expression has resolved into a ridge of epithelial cells (the gill arch epithelial ridge, GAER; black arrow) along the leading edge of the expanding hyoid and gill arches, and h.Ptc2 expression indicates that this signal is transduced in posterior-distal mesenchyme, epithelium and core mesoderm. i. By stage 29, j.–k. expression of Shh is maintained in the GAER, and l.Ptc2 expression indicates sustained posterior-distal transduction of this signal in posterior-distal arch mesenchyme, epithelium and core mesoderm. m. The GAER is recognizable as a pseudostratified ridge of Shh-expressing epithelial cells. m, mandibular arch; h, hyoid arch; 1-5, gill arch 1-5. Dashed lines in a.,e., and i. indicate plane of section in b.–d., f.–h. and j.–l., respectively. Modified from Gillis and Hall (2016).
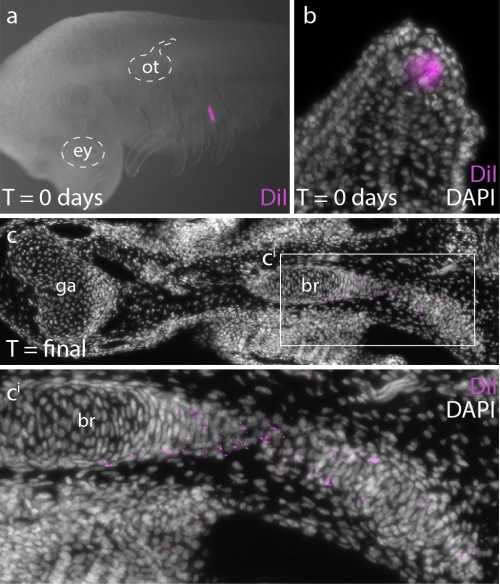
In order to determine whether Ptc2+ mesenchymal cells (i.e. mesenchymal cells responding to Shh signal) ultimately contribute to the branchial ray skeleton, we conducted a fate mapping experiment. Over the past several years, I have developed protocols for the experimental manipulation and long term in/ex ovo culture of skate embryos, and this now allows us to conduct targeted embryonic manipulations (e.g. by microinjecting and surgically manipulating specific regions of the embryo) and to focally label populations of embryonic cells, in order to trace the long-term fates of their progeny (Fig. 5a,b). For this experiments, we microinjected the lipophilic dye CM-DiI immediately subjacent to the Shh-expressing epithelium of the gill arch, so that we could assess the contribution of these cells (and their progeny) to the gill arch skeleton. CM-DiI is readily incorporated into cell membranes, and is retained in daughter cells through mitosis (although diluted somewhat with each round of cell division). Importantly, though, CM-DiI will persist through fixation and paraffin sectioning, and so upon skeletal differentiation, we can use fluorescent microscopy on thin sections to recover even very small specks of membrane-localized CM-DiI (indicating decent from mesenchymal cells that were Shh-responsive earlier in development). These experiments demonstrated that Shh-responsive gill arch mesenchyme does contribute to branchial rays (Fig. 5c), and suggest that this signaling pathway may be directly influencing the behaviour and fate of branchial ray progenitors.
Figure 5.a. CM-DiI was microinjected subjacent to the GAER at stages 27 and 29, to b. label Ptc2+ (i.e. Shh-responsive) mesenchyme (compare b. with figure Fig. 4h). c. After 10 weeks of development, CM-DiI-positive chondrocytes were recovered in branchial rays. Modified from Gillis and Hall (2016).
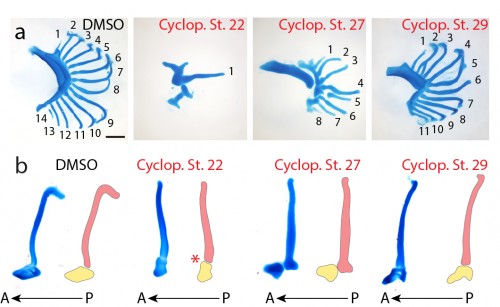
Finally, to test the function of Shh signaling during skate branchial ray development, we conducted a series of in ovo pharmacological treatments. Skate embryos develop in large, leathery egg shells, and are amenable to bath treatment by in ovo injection of small molecules. We used cyclopamine – a small molecular inhibitor of the hedgehog signaling pathway – to inhibit Shh signaling at different stages of gill arch development, and to test for stage-specific roles for Shh signaling in bronchial ray patterning. We chose three stages for treatment: stage 22 (gill arches have still formed, and Shh is expressed in posterior arch epithelium), stage 27 (gill arches are undergoing lateral expansion, with Shh signaling resolved to an epithelial stripe along the leading edge of the expanding arch) and stage 29 (just prior to the condensation of the gill arch endoskeleton). Interestingly, upon manipulation of Shh signaling at these different stages of gill arch development, we observed branchial ray defects that were broadly reminiscent of the skeletal defects observed upon manipulation of Shh signaling during limb development. For example, we observed that progressively earlier inhibition of Shh signaling resulted in a progressively greater deletion of branchial rays (i.e. cyclopamine treatments at stages 22 and 27 resulted in a significant reduction in the number of branchial rays on each arch, while treatment at stage 29 resulted in no significant difference in branchial ray number) (Fig. 6a). We also observed that cyclopamine treatment at stage 22 resulted in loss of anterior-posterior axis specification (i.e. the few branchial rays that did form articulate down the midline of the arch, rather than along the posterior margin of the arch) while cyclopamine treatment at stages 27 or 29 had no effect on the anterior-posterior axis (Fig. 6b). It therefore appears that, as in the limb bud, Shh signaling functions initially in skate gill arches to establish the anterior-posterior axis, and subsequently to maintain proliferative expansion of branchial ray endoskeletal progenitors.
Figure 6. a. Examples of branchial ray counts from control embryos (DMSO) and embryos treated with cyclopamine in ovo at stages 22, 27 and 29. Progressively earlier treatment with cyclopamine results in a progressively greater reduction in the number of branchial rays. b. The gill arches of embryos treated with cyclopamine at stage 22 also exhibit a loss of anterior-posterior axis specification (with branchial rays articulating down the midline of the gill arch, rather than down the posterior margin). This defect is not observed in control embryos (DMSO), nor in embryos treated with cyclopamine at stages 27 or 29. Modified from Gillis and Hall (2016).
So what does this shared role for Shh signaling in gill arch and limb bud patterning mean? It is possible that limbs share a patterning mechanism with gill arches because these structures are, indeed, transformational homologues (i.e. fins and limbs evolved by transformation of a gill arch in an ancestral vertebrate, as proposed by Gegenbaur). However, it may also be that gill arches and fins/limbs have independently recruited a deeply conserved “core” appendage patterning mechanism (i.e. parallel evolution, leading to serial homology), or that gill arches and fins/limbs are convergently using the Shh signaling pathway for similar purposes. Only palaeontological data can tell us about anatomical transitions, and such data are needed in order to formally test Geganbaur’s hypothesis of gill arch-paired fin transformational homology. However, it is now clear that some of the anatomical parallels that led Gegenabur to propose his gill arch hypothesis of fin origins reflect common underlying patterning mechanisms, and further investigation of the molecular basis of branchial ray patterning in cartilaginous fishes will allow us to determine whether these common mechanisms are the result of parallel evolution or convergence. I think that this study sets out an exciting path forward to address the origin and evolution of paired appendages in vertebrates, and highlights how complementary palaeontological and developmental approaches are needed in order to truly address the big, unanswered questions in vertebrate body plan evolution.
References
Gegenbaur, C (1878) Elements of Comparative Anatomy. London, UK: Macmillan.
Roth, V.L. (1984) On homology.Biol. J. Linn. Soc. 22, 13-29.
Saunders, J.W. and Gasseling, M.T. (1968). Ectodermal and mesenchymal interactions in the origin of limb symmetry. In Epithelial Mesenchymal Interactions (Ed. R. Fleischmajer and R. E. Billingham). Baltimore, William and Wilkins, pp. 78- 97.
Here are the highlights from the current issue of Development:
The origins of lung macrophages revealed
Tissue-resident macrophages are phagocytic cells that are essential for the response to injury and infection. Both within and between tissues, macrophages can show distinct characteristics, but are these attributes developmentally defined or determined by the microenvironment? In the mouse lung, there are two distinct macrophage populations: alveolar macrophages that reside within the lumen of the alveoli and interstitial macrophages that occupy the interalveolar space and elsewhere in the lung parenchyma. Some studies have suggested that alveolar macrophages originate from and are repopulated by an interstitial macrophage precursor, while others indicated that they can maintain themselves independently. Serena Tan and Mark Krasnow (p. 1318) now show that there are in fact three developmentally distinct lineages that populate the lung in three waves, with minimal interconversion between them – at least under homeostatic conditions. The first population, derived from yolk sac haematopoietic cells, populate the interstitial space in embryogenesis, but become confined to peripheral and perivascular regions postnatally. The second, of as-yet-unknown origin, initially occupy the interstitium but then become alveolar macrophages. The third population enters the lung postnatally from circulating monocytes and constitute the majority of mature adult interstitial macrophages. In the lung at least, it appears that developmental origin, rather than environmental influence, is the primary determinant of macrophage identity and diversity.
Transcription without TBP
The canonical mechanism of transcription initiation in all metazoans involves recruitment of TATA-binding proteins (TBP, TLF or TBP2 in vertebrates) to the promoter, as a rate-limiting step before binding of RNA polymerase II. TBP, TLF and TBP2 have non-redundant functions, but the degree of redundancy between them is not clear; nor is it well understood whether there are TBP-independent mechanisms of transcription initiation in vivo. On p. 1340, Gert Veenstra and co-workers now show that, during early Xenopus development, there is a small group of genes whose transcription is independent of all TBP family members (denoted TBP family-insensitive or TFI genes). These genes are enriched for factors expressed in mesoderm and at Spemann’s organiser and include several key transcription factors involved in mesendoderm specification. Strikingly, most TFI genes are bound by these TFI transcription factors. Gcn5, a component of the SAGA complex thought be involved in non-canonical transcription initiation, is recruited to TFI promoters upon TBP family knockdown, is not required for their transcription in the presence of TBP-related factors, but seems to compensate in their absence. This work provides clear evidence that alternative mechanisms of transcription initiation exist in vivo, and that they may be preferentially used for a particular set of key developmental genes.
Completing the neuroblast map
In the developing Drosophila embryonic central nervous system (CNS), the pattern of neural stem cells – neuroblasts (NBs) – is highly stereotyped, both between individuals and, in the truncal ventral nerve cord, between segments. Over the past decades, multiple studies have mapped the spatio-temporal origin and gene expression signature of the embryonic NBs in the brain, thorax and abdomen. On p.1290, Rolf Urbach and colleagues provide the final piece to this puzzle by providing a comprehensive map of the NBs in the gnathal (labial, maxillary and mandibular) segments of the embryo. In doing so, they are able to compare the NBs complement of each segment to identify homologies between NBs in different embryonic origins. Their work demonstrates the progressive loss of NBs from trunk to progressively anterior gnathal segments and analyses its cause. Despite the reduced NB number, homologies in developmental origin and expression pattern are clearly recognisable, and can also be traced into posterior brain segments. The wealth of data in this and related papers provide an essential foundation to understand the molecular and evolutionary basis of segmental diversification of the CNS.
Vascular development in technicolour
During development, the vascular endothelium becomes covered by mural cells (MCs) – vascular smooth muscle cells or pericytes – that are essential for vascular stability and homeostasis. MCs are known to be of mesodermal or neural crest origin, but little is known about how they are recruited to and cover the vessels – primarily because live imaging of this process has been challenging. Now (p.1328), Shigetomo Fukuhara, Naoki Mochizuki and colleagues overcome this hurdle by developing transgenic zebrafish lines to mark MCs fluorescently. They then use these tools to follow the origin and subsequent behaviour of MCs in both cranial and trunk regions of the embryo. The authors find that trunk MCs are mesodermal in origin, while both neural crest and mesoderm populations contribute to cranial MCs. MCs appear to be recruited to specific vessels, such as the dorsal aorta in the trunk or the basilar artery in the head, and then migrate using inter-endothelial cell junctions as a scaffold to cover other vessels – preferentially the arteries. As well as providing important insights into MC behaviour, the tools developed here should serve as a valuable resource for the community for future analyses of vascular development.
Determining dendritic diversity in Drosophila
Neuronal morphology is highly variable, particularly in terms of the complexity of dendritic arborisation, and this variability is crucial for appropriate function. But how is such diversity established and regulated? Wesley Grueber and colleagues (p. 1351) set out to address the transcriptional inputs into this process using a subset of Drosophila sensory neurons, the multidendritic (md) neurons, whose morphology is regulated by the transcription factor Cut. Cut expression is absent in neurons that have simple morphology and function as proprioceptors, but is expressed at variable levels in nociceptive or touch-sensitive neurons with more complex dendrites. Through a series of mosaic genetic analyses, the authors find that Cut represses the expression of the Pdm1/2 transcription factors in a subset of md neurons, which suppresses the ability of Pdm1/2 to restrain dendritic arborisation. Upstream of Cut, the transcriptional repressors Vestigial and Scalloped modulate Cut levels to limit dendritic elaboration – in this case repressing a complex morphology and favouring a less complex type of branching. Together, these data identify a network of repressive interactions that regulate neuronal morphology and thus help to define neuronal identity and diversity.
PLUS…
CncRNAs: RNAs with both coding and non-coding roles in development
RNAs are known to regulate diverse biological processes, either as protein-encoding molecules or as non-coding RNAs. However, a third class that comprises RNAs endowed with both protein coding and non-coding functions has recently emerged. Such bi-functional ‘coding and non-coding RNAs’ (cncRNAs) have been shown to play important roles in distinct developmental processes in plants and animals. Here, Karuna Sampath and Anne Ephrussi discuss key examples of cncRNAs and review their roles, regulation and mechanisms of action during development. See the Primer on p. 1234
Developmental origin and lineage plasticity of endogenous cardiac stem cells
Over the past two decades, several populations of cardiac stem cells have been described in the adult mammalian heart. Here, Richard Harvey, Jason Kovacic and colleagues summarize what is known about different these populations, highlighting their developmental origins and defining characteristics, and their possible contribution to heart organogenesis and regeneration. See the Review on p. 1242
Lineage-specific stem cells, signals and asymmetries during stomatal development
Stomata – the dispersed pores found in the epidermis of land plants that facilitate gas exchange – are formed from progenitor cells that execute a series of differentiation events and stereotypical cell divisions. Here, Soon-Ki Han and Keiko Torii review the intrinsic and extrinsic factors that control stomatal development, highlighting striking similarities between plants and animals with regards to their mechanisms of specialized cell differentiation. See the Review on p. 1259
Gerald (Gerry) Rubin, pioneer in Drosophila genetics, is Founding Director of the HHMI-funded Janelia Research Campus. In this interview, Gerry recounts key events and collaborations that have shaped his unique approach to scientific exploration, decision-making, management and mentorship – an approach that forms the cornerstone of the model adopted at Janelia to tackle problems in interdisciplinary biomedical research. Gerry describes his remarkable journey from newcomer to internationally renowned leader in the fly field, highlighting his contributions to the tools and resources that have helped establish Drosophila as an important model in translational research. Describing himself as a ‘tool builder’, his current focus is on developing approaches for in-depth study of the fly nervous system, in order to understand key principles in neurobiology. Gerry was interviewed by Ross Cagan, Senior Editor of Disease Models & Mechanisms.
Gerald M. Rubin was born in Boston, Massachusetts, in 1950. He undertook his undergraduate degree in Biology at MIT from 1967 to 1971, and worked at Cold Spring Harbor Laboratory during the summers of 1970 and 1971. For his PhD, he studied 5.8S ribosomal RNA under the supervision of Sydney Brenner at the MRC Laboratory of Molecular Biology, University of Cambridge. After graduating in 1974, Gerry undertook post-doctoral research in David Hogness’s lab at Stanford University. This marked Gerry’s first step into Drosophila research, which at that time was experiencing resurgence in the biomedical arena. Gerry’s first faculty appointment was at Harvard Medical School, but he moved after 4 years to the Carnegie Institute of Washington’s Department of Embryology. It was here that, together with Allan Spradling, Gerry pioneered the use of transposable elements as a tool for genetic manipulation – a breakthrough that revolutionized Drosophila genetics. In 1983, Gerry moved to the University of California, Berkeley, where his group studied pattern formation and signal transduction during development of the fly eye; he later led the sequencing of the fruit fly genome, which was completed in 2000. Currently, Gerry is a Vice President at Howard Hughes Medical Institute and the Executive Director of its biomedical research institute, the Janelia Research Campus in Ashburn, Virginia. His lab probes fly brains to gain insight into the neuronal circuits underlying learning and memory, sleep regulation, visual perception and sensory integration, and Gerry remains dedicated to the development of neurobiology and genomics tools. He has been honored with a long list of awards in recognition of his contributions to Drosophila research so far.
My first question is: how did you become interested in flies?
I was a graduate student in Cambridge, England, in the pre-cloning, pre-DNA-sequencing era, working on RNA sequencing in yeast. David Hogness came and gave a seminar in 1973 on his recent work on fly chromosomes. At that time, many labs, including his, were trying to develop methods for cloning segments of eukaryotic DNA in bacteria. Dave’s work really fascinated me and prompted me to apply to do my post-doctoral research in his lab. He agreed, and I was ready to move to Stanford in September 1974. By then, recombinant DNA technology was up and running, and the Hogness lab had generated about 200 Drosophila recombinant clones. My first project was to make a library that had enough clones in it to cover all the DNA sequences in the genome. This was the first of several times in my career when I was fortunate to be in the right place at the right time. There had been almost no prior work on cloned DNA segments, so almost any experiment led to novel insights. The experience also taught me the power of new techniques to advance science.
You and Allan Spradling pioneered the use of transposable P elements as a gene manipulation tool – this is one of the first things I learned about in Drosophila genetics. How did you come to be involved in this work?
Most of my post-doc time was spent characterizing the organization of fly DNA by making restriction maps of clones, measuring the amount of repetitive DNA (using reassociation kinetics – readers over the age of 50 will remember this technique!) and performing in situ hybridization to polytene chromosomes. We found a lot of repetitive DNA dispersed in the fly genome. Around the same time, Eric Davidson at Caltech had proposed that repetitive DNA segments might have important regulatory roles, so I continued studying them after setting up my own lab at Harvard. We soon realized that many of them were retroviral transposable elements and that they were causing spontaneous mutations in the genome. There was a growing body of literature on movable genetic elements, and Mel Green – among others – had proposed that their insertion caused some of the unstable mutations that had been described in Drosophila. We decided to try to clone the white locus – a gene that is important for eye color in Drosophila – because the many spontaneous mutations at this locus would allow us to directly test these ideas. In 1980, we succeeded in cloning the locus, by a method now known as transposon tagging, and we were soon able to show that several mutations in the white locus were in fact due to transposon insertion. That same year, I went to the Cold Spring Harbor symposium on transposable elements and heard Bill Engels give a talk on hybrid dysgenesis, which he proposed was caused by a family of transposons called P elements. In the slow lunch line following Engel’s talk, Margaret Kidwell, Paul Bingham and I designed an experiment to generate and molecularly characterize hybrid-dysgenesis-induced mutations in white. If Engel’s hypothesis were correct, the mutant whitegenes would contain insertions of the postulated P element. This turned out to be the case.
At Carnegie, I went on to characterize the structure of a large number of P elements. We discovered that there were full-length elements with a conserved structure, and a heterogeneous set of smaller elements containing internal deletions. This was immediately reminiscent of Barbara McClintock’s pioneering work in maize that defined ‘two-element systems’ of autonomously transposing and defective elements dependent on the autonomous elements for trans-acting functions. At that point it was obvious that the full-length P elements might be good vectors for making transgenic animals. Allan Spradling, my colleague at Carnegie, and I discussed trying to make this work. Allan took on the task of injecting embryos and I did most of the molecular work. It took us less than a year to have initial success and we reported our work in two articles in Science in the fall of 1982. We did all the experiments ourselves because it was a high-risk ‘nutty’ experiment!
The breakthrough dovetailed nicely with the burst of genetic screens that were happening around the same time. Did you know, when you were doing this work with Allan, the impact it would have?
We realized, from the work in yeast and bacteria, that the ability to make transgenic animals would be tremendously powerful. Methods to genetically engineer the genomes of single-cell organisms had been around for a while, but we were the first to successfully engineer the germline of a multicellular organism. A lot of people had been interested in doing this, and we were fortunate that the experiments we tried actually worked. A lot of things fell into place and it was remarkably efficient and successful. It was one of the few times you have in your career as a scientist when everything falls into place and the sort of dream experiment actually works the way you draw it up on paper. Once we had the system up and running, we passed on the reagents and so, by the time our paper was published, several other labs had already confirmed that it worked.
The P-element-based approach came along at a good time because everyone was hungry for positional cloning. Everyone had in mind the genes they wanted to identify and the experiments they wanted to do. There were a lot of fascinating developmental mutants, for example those isolated in the Nobel-Prize-winning genetic screens of Nüsslein-Volhard and Wieschaus, that hadn’t been characterized on a molecular level – we had no idea how the mutations were affecting cellular processes. The field then really took off and I think it was the synergy between molecular cloning and all the mutants and techniques that we already had that meant we could finally get at molecular mechanisms. A tremendous amount of data came out very quickly from many laboratories, particularly on developmental processes, and it was information that we hadn’t been able to glean using classical embryology in flies – or indeed any other organism. The majority of the components of all the signal transduction pathways that exist in humans were first discovered through this work. We could now ask questions that were just not possible to ask beforehand. It was as though the whole field awakened because of the sudden leap in several key technologies.
Your decision to share the reagents was critical. What made you decide to do this?
There is a tradition to share among Drosophila researchers, going back to the days of Thomas Morgan. The fly field has always seemed much more open in this respect than the mammalian field. Allan and I considered our options: we had this very useful technique and could wait until it was published to distribute it – which would give us exclusive access for 6-8 months – or to give it to people straight away. We decided that the benefits of distributing it immediately would outweigh any progress we could make in our work within a few months. I learned a lot from the experience and think it one of the better decisions I have made in my life. I consider myself to be a tool builder – the thing I’m most proud of in my scientific career is the development of tools and methods. Post-docs in my lab were eager to test hypotheses, but what has always motivated me is the building of tools that help overcome technical obstacles that block progress.
You led the fly genome project that was completed back in 2000. How did you get involved with that and what did the experience teach you?
My lab was doing interesting work on eye development at the time, and I took on the task of leading the genome project with some reluctance. Allan and I felt that the fly genome really needed to be sequenced, and we realized that we should take the lead because we were well-established and in the position to be doing something for the community. We got a grant and recruited some younger scientists, and I became the ‘figurehead’ of the project. Over time many of the younger scientists got bored or were hired by industry, and I needed to spend more and more of my time on the project. But I had staked my reputation that the project would be completed. So I was glad to be able to collaborate with Celera Genomics and Craig Venter, which was not only very enjoyable and a tremendous help in getting the project completed, but also demonstrated the power of the whole-genome shotgun method for sequencing large genomes. Once the fly sequence was available, I happily got out of the genome sequencing field and returned to the interesting questions that could now be asked about gene organization and function.
One important thing my genomics experience taught me was that I like management. I’m one of the few scientists that I know who actually enjoys management and thinks it is, in its own way, as interesting and challenging as scientific discovery and research. Managing projects, managing people, getting people to work well together and doing collaborative interdisciplinary research projects is a unique kind of challenge. This combination of enjoying scientific discovery as well as management naturally led me to my current role at Janelia.
Let’s talk a little bit about Janelia, which you have seen evolve from concept to implementation. What was the thinking behind its approach to interdisciplinary research, and what do you hope the long-term impact of the campus will be?
I feel that the way research is done in the US nowadays, where scientists depend for their livelihood on getting their grants renewed, can have a profound negative effect on research. It can make the work short-sighted by forcing scientists to pursue directions depending on what the funding agencies want, rather than by following their instincts and passions. I have always felt that this isn’t the ideal way to do all science. I moved to Janelia at a time when collaborative research and tool building were also undervalued and underappreciated. If someone wanted to build a new optical microscope, physics departments wouldn’t hire them because optics is old physics, and they wanted string theorists. In biology, people cared only about the hypothesis being tested and not about the tools. The tools are needed to advance science, yet science wasn’t – and still isn’t – funded to align well with this. There is a lack of appreciation for basic research in general, although basic science brings the fundamental breakthroughs that underlie translational research. Just look at CRISPR/Cas9, which would never have emerged if we didn’t have people studying ‘weird’ phenomena in bacteria. The pressure for recipients of grants to show short-term return on investment by tackling ‘practical’ problems – often defined top-down by funding agencies – is something I have seen steadily increase during my 40 years in science.
I felt that places like the MRC Laboratory of Molecular Biology [LMB], where I was fortunate enough to do my PhD, had a much higher rate of innovative research. I began to think about what made such places so great. As they were internally funded, people didn’t worry about convincing grant review committees to fund their research. Decisions got made much more quickly, the labs were small and people didn’t have other responsibilities; they could keep working in the lab, even at a late stage in their career. At the MRC LMB, I would see Nobel Prize winners with a pipette in hand, doing experiments 8 hours a day, 5 days a week. You would never see that in the US, and I had a rude awakening when I came back here.
I wanted to create a supportive environment for aspiring scientists who really wanted to just keep doing science without having to manage an enterprise and raise funds. And where there would be a strong synergy between the work in individual labs. I think Janelia provides that kind of environment. It’s not right for everyone – maybe it’s only right for 5% of scientists – but for those people it provides something very special that would be difficult to find elsewhere. My short-term goal is to help all the talented people we have recruited to be successful in doing important, innovative science that would be unlikely to happen elsewhere. My ultimate goal is to have a disruptive impact on scientific culture as a whole, and to change the way a significant fraction of research is funded and scientists are evaluated, especially in the US, by demonstrating the success of an alternative model. I look at Janelia as an experiment in the sociology of science and we continue to refine our working hypothesis. But I think the initial data from our first 10 years are very encouraging.
What would you say to a young scientist thinking about entering the fly field, particularly in terms of the place of Drosophila in this brave new world of translational research?
I think there’s no denying that Drosophila has been very useful as a disease model. Many components of signal transduction pathways, which are important drug targets, were discovered in yeast, worms or flies. People are working on very interesting experiments relating to growth control, cell movement and other basic mechanisms that are important in disease contexts. Drawing from my own scientific interests, I think there is a lot that flies can contribute to neurobiology. Flies show complex behavior, and I believe that the basic rules about how a biologically constructed computational device can perceive the world, navigate through it, learn and remember things, can be learned by studying flies. Flies are particularly amenable to a ‘black box approach’ in science: studying the consequences of manipulating genes and cells using clever assays can yield information that doesn’t depend on having a good hypothesis. You can gather information, in a non-biased way, on how things work.
From an intellectual point of view, I think that working on an organism like Drosophila is highly rewarding. Certainly for students, the ability to design and do really interesting experiments and see the outcome in a reasonable timeframe is tremendously powerful in terms of learning how to do science. In other fields, experiments can move so slowly that if you’re a graduate student you don’t really get to do much during the limited time you have for your PhD. The difficulty lies in convincing the funding agencies of the value of model systems such as Drosophila. There is insufficient appreciation from people making these decisions, most of who have worked in medicine or using mammalian systems, about the power of a simpler model. The work going on in vertebrate systems is important, but I think that the simpler systems deserve more investment than they get at present. Dollar for dollar the fly community has produced more insights with medical relevance than the mouse community. Similarly, in the cancer field, most of the important cell cycle mutants and checkpoints initially came from studies in yeast. The intellectual history, the source of important ideas and breakthroughs, isn’t always fully appreciated, and thus the contribution of simple systems is undervalued.
You’ve mentioned in the past that one of your biggest achievements in science is the people you have trained. Certainly, as somebody on the outside looking in, you’ve had an astonishing number of amazing post-docs that have gone on to make an impact. What is your approach to training?
It’s true that this is one of the things I’m most proud of. It certainly is as rewarding as a good publication record, and of course it’s a two-way street. The fact that I had all those great people in my lab made it easy for me to develop my own reputation based on the work they did.
I was fortunate as a graduate student; I pretty much worked on my own. I never wanted anyone telling me what to do. I was the sole author of two of the three papers I published as a graduate student; on the other one I had a co-author but it wasn’t my adviser. I had my own project and it moved in the direction I chose. I think most people are more motivated when they’re working on their idea and not an idea that someone else gives them. I was also fortunate because I entered a field that was in its early stages. The Drosophila community was growing and there weren’t that many labs who had established research programs using recombinant DNA and so I attracted to my lab a remarkable group of highly talented people. I realized that the way I would get the most out of them was to let them work on a problem they were passionate about, tell them they could take that problem with them when they left my lab, and just be on hand to provide technical advice and to act as a sounding board for ideas or help with writing manuscripts. Smart people want to be in an environment where they have freedom to explore their ideas and are challenged by other smart people. It was wonderful for me because every day there was interesting science going on, and I never felt like I needed to be the source of all the ideas or that everything was on my shoulders. I was creating an environment that facilitated other people’s ability to work, rather than directing them. As a young scientist, I was always in an environment that was positive, upbeat and optimistic. I feel tremendously fortunate and feel that I have a big obligation to try and recreate some of that for the next generation.
What advice would you give to a young start-up on how to run a successful lab?
Keep it small and be highly selective. This advice seems counterintuitive: you’re thinking, you have an empty lab, that you have some money to hire people and any reasonable pair of hands is better than nothing. But actually, if you get the wrong person, it can be disastrous. Aim to hire people smarter or more talented than yourself. Having people who are not motivated and passionate can be very detrimental – motivation and passion are probably as important as intellectual ability. Also, you should protect your time so that you can continue to do things yourself at the bench.
Finally, if not science, what would you be doing?
I got my first job working in a lab when I was 14 years old (washing glassware) and I knew right from that point that I wanted to work in science. I suppose if I hadn’t gotten a faculty job I would be teaching high-school biology. As I’ve mentioned, I’ve been extremely fortunate in every stage of my career. I’ve been in the right place at the right time on several occasions and I got tremendous support from my advisers and teachers – as well as from my wife of 38 years. My graduate and post-doc experiences were also extremely positive. I can’t imagine anything else I could have done that would have been as satisfying or as much fun – I never looked at it as work. I’m always amazed that I get paid for doing what I love doing.
Surrounded by the breathtaking landscape of the Rocky Mountains, hundreds of graduate students and researchers gathered for the 8th biennial Canadian Developmental Biology Conference at the Banff Centre, minutes outside of Banff, Alberta. This four-day conference was filled with a variety of stimulating seminars and poster sessions, reflecting some of the latest advances in the field of developmental biology; meanwhile the easy-going conference atmosphere encouraged a number of friendly and thought-provoking discussions between students, post-docs, and scientists from across Canada and abroad.
Researchers from diverse backgrounds presented in the meeting’s five main scientific sessions, incorporating a number of model organism systems and developmental processes. The first session – stem cells and regeneration – included a talk from Jeff Biernaskie (University of Calgary) on the application of adult dermal stem cells to promote wound-healing after skin grafts, using a mouse xenograft model. The session also included a talk from Rohan Khadilkar (University of British Columbia), who received the award for the best post-doctoral presentation on his research on the Drosophila hematopoietic stem cell niche and the role of septate junctions in moderating HSC differentiation. The second session focused on gene expression and development. Julie Claycomb (University of Toronto) presented on the role of an Argonaute protein, CSR-1, in licensing C. elegans germline gene expression through small RNAs and chromatin modifications. Other talks were Marie Kmita’s (University of Montréal) exploration of a Hoxa11 antisense enhancer to regulate distal limb development, and Alexandra Dallaire’s (Best student talk winner, CHU de Québec Research Centre) discussion of how microRNAs mediate mRNA stability in C. elegans.
Following were sessions on growth, differentiation and patterning – including a talk from Dominique Bergmann (Stanford University) on asymmetric cell division and fate specification in plant stomatal cells – as well as cell proliferation, migration and morphogenesis. The latter, featured, amongst others, a presentation by Vanessa Auld (University of British Columbia) on glia-ECM interactions in Drosophila and their role in protecting the peripheral nervous system. The fifth and final session focused on developmental models of disease. One of the highlights from this session was a talk by Brian Ciruna (Princeton University) on a zebrafish ciliary-defect model of scoliosis.
In addition to the five primary sessions, a major highlight was the keynote lecture and conference education session. Keynote speaker Freda Miller (University of Toronto), gave a captivating presentation on the use of the mouse cerebral cortex to study neurogenesis and the role of translational repression in regulating neuronal cell fates. This lecture was a wonderful way to start the conference and sparked many interesting discussions during the night’s opening reception. Furthermore, all attendees enjoyed an interactive presentation by Scott Barolo (University of Michigan), who demonstrated how the game Mastermind could be used to teach scientific thinking strategies.
On the final evening of the conference, attendees donned their cowboy boots and Western attire and headed over to MountainView Barbeque for a buffet-style meal and lively night of socializing. The barbeque was complete with a huge bonfire and live country music band, and to led hours of cheerful conversation and even some line-dancing.
This year’s conference provided an excellent opportunity for Canadian and international biologists from all career stages to interact and discuss leading findings in developmental biology research. On behalf of all conference attendees, I would like to say a tremendous thank you to the conference organizers, Savraj Grewal, Dave Hansen and Sarah McFarlane (University of Calgary), for putting together an excellent program of speakers and events, including the poster sessions, 60-second science presentations and conference banquet. Also to Paul Mains for organizing the judging of trainee poster presentations, and to the meeting sponsors (CIHR, SDB and many others). Congratulations to all the winners of the poster competition (below) and the Society of Developmental Biology travel and financial awards. Namely, Rohan Khadilkar for the best postdoctoral talk (SDB cash award), and Alexandra Dallaire (SDB travel award) and Anna Kobb (SDB cash award) for the best graduate student talks.
The 9th Canadian Developmental Biology conference will be held in 2018 in Mont-Tremblant, Québec. Looking forward to seeing you all there!
By: Isabella Skuplik
Student poster award winners: Aarya Chithran, Miranda Hunter, Katharine Goodwin, Rotem Lavy, Enrique Gamero-Estevez, Adam Kramer, Tanya Foley, Corey Arnold, Sonya Widen, Eric Hall, Raghda Gemae, Mriga Das, Victoria Yan, Isabella Skuplik, Bensun Fong, Dova Brenman.
Postdoctoral poster award winners: Matthew Hildebrandt, Pierre Mattar, and Sérgio Simões (SDB cash awards), Guang Yang (SDB travel award).
Travel award winners: Sarah Garner, Siavash Amon, Amanda Baumholtz, Scott De Vito, Adrienne Elbert, Sarah Gignac, Zachary Hall, Adam Kramer, Stephanie Tkachuk, Jessica Yu.
 (5 votes)
(5 votes)

 Every year, the British Society for Developmental Biology (BSDB) awards the Beddington Medal to the best PhD thesis in developmental biology. The 2016 award went to Elena Scarpa, who did her PhD with Roberto Mayor at University College London (UCL). We caught up with Elena at the BSCB/BSDB Spring meeting, and we asked her about her thesis work on the neural crest and what she is doing now.
Every year, the British Society for Developmental Biology (BSDB) awards the Beddington Medal to the best PhD thesis in developmental biology. The 2016 award went to Elena Scarpa, who did her PhD with Roberto Mayor at University College London (UCL). We caught up with Elena at the BSCB/BSDB Spring meeting, and we asked her about her thesis work on the neural crest and what she is doing now.
 (No Ratings Yet)
(No Ratings Yet)






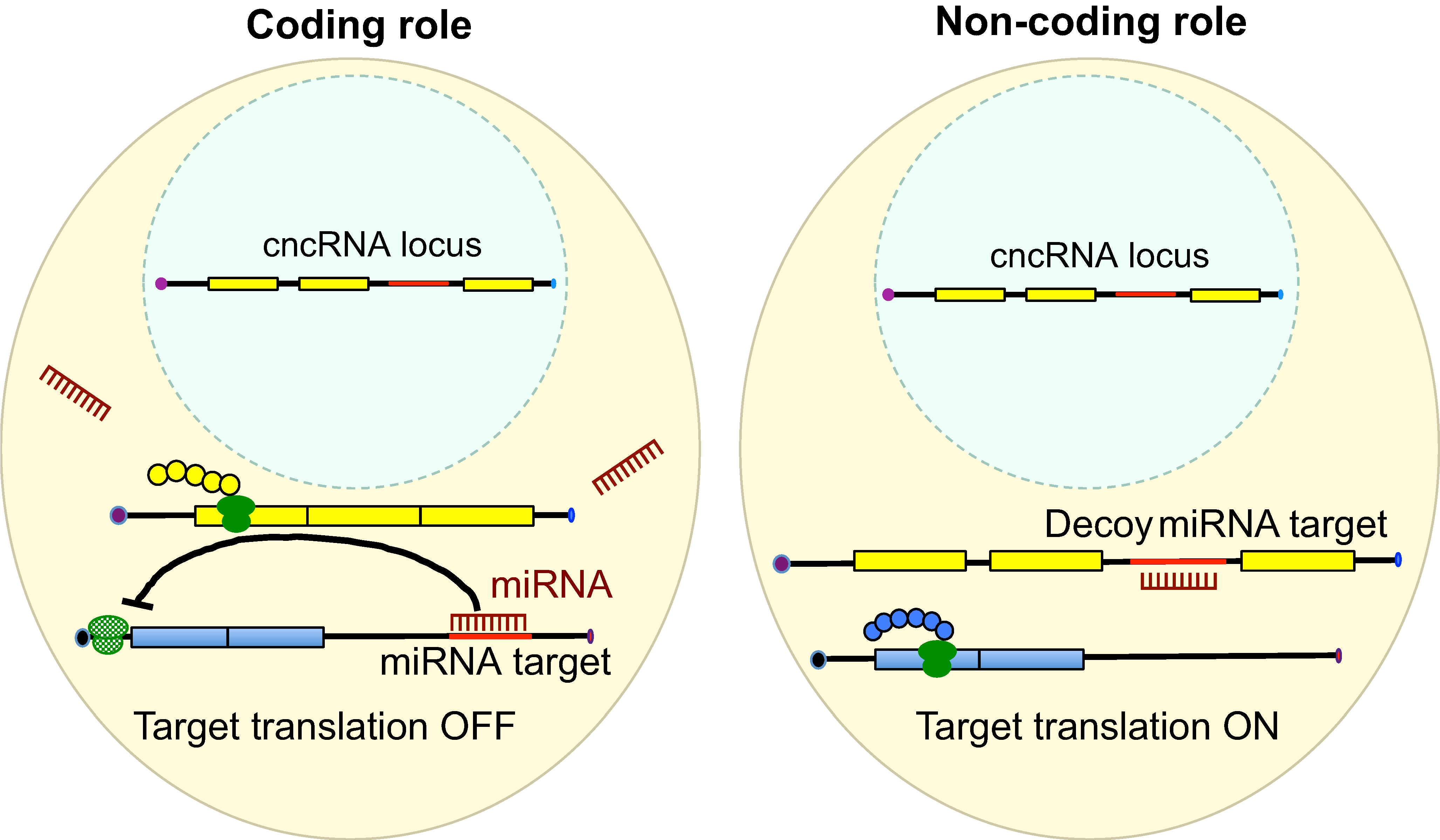 RNAs are known to regulate diverse biological processes, either as protein-encoding molecules or as non-coding RNAs. However, a third class that comprises RNAs endowed with both protein coding and non-coding functions has recently emerged. Such bi-functional ‘coding and non-coding RNAs’ (cncRNAs) have been shown to play important roles in distinct developmental processes in plants and animals. Here,
RNAs are known to regulate diverse biological processes, either as protein-encoding molecules or as non-coding RNAs. However, a third class that comprises RNAs endowed with both protein coding and non-coding functions has recently emerged. Such bi-functional ‘coding and non-coding RNAs’ (cncRNAs) have been shown to play important roles in distinct developmental processes in plants and animals. Here,  Developmental origin and lineage plasticity of endogenous cardiac stem cells
Developmental origin and lineage plasticity of endogenous cardiac stem cells Stomata – the dispersed pores found in the epidermis of land plants that facilitate gas exchange – are formed from progenitor cells that execute a series of differentiation events and stereotypical cell divisions. Here,
Stomata – the dispersed pores found in the epidermis of land plants that facilitate gas exchange – are formed from progenitor cells that execute a series of differentiation events and stereotypical cell divisions. Here,  Gerald M. Rubin was born in Boston, Massachusetts, in 1950. He undertook his undergraduate degree in Biology at MIT from 1967 to 1971, and worked at Cold Spring Harbor Laboratory during the summers of 1970 and 1971. For his PhD, he studied 5.8S ribosomal RNA under the supervision of Sydney Brenner at the MRC Laboratory of Molecular Biology, University of Cambridge. After graduating in 1974, Gerry undertook post-doctoral research in David Hogness’s lab at Stanford University. This marked Gerry’s first step into Drosophila research, which at that time was experiencing resurgence in the biomedical arena. Gerry’s first faculty appointment was at Harvard Medical School, but he moved after 4 years to the Carnegie Institute of Washington’s Department of Embryology. It was here that, together with Allan Spradling, Gerry pioneered the use of transposable elements as a tool for genetic manipulation – a breakthrough that revolutionized Drosophila genetics. In 1983, Gerry moved to the University of California, Berkeley, where his group studied pattern formation and signal transduction during development of the fly eye; he later led the sequencing of the fruit fly genome, which was completed in 2000. Currently, Gerry is a Vice President at Howard Hughes Medical Institute and the Executive Director of its biomedical research institute, the Janelia Research Campus in Ashburn, Virginia. His lab probes fly brains to gain insight into the neuronal circuits underlying learning and memory, sleep regulation, visual perception and sensory integration, and Gerry remains dedicated to the development of neurobiology and genomics tools. He has been honored with a long list of awards in recognition of his contributions to Drosophila research so far.
Gerald M. Rubin was born in Boston, Massachusetts, in 1950. He undertook his undergraduate degree in Biology at MIT from 1967 to 1971, and worked at Cold Spring Harbor Laboratory during the summers of 1970 and 1971. For his PhD, he studied 5.8S ribosomal RNA under the supervision of Sydney Brenner at the MRC Laboratory of Molecular Biology, University of Cambridge. After graduating in 1974, Gerry undertook post-doctoral research in David Hogness’s lab at Stanford University. This marked Gerry’s first step into Drosophila research, which at that time was experiencing resurgence in the biomedical arena. Gerry’s first faculty appointment was at Harvard Medical School, but he moved after 4 years to the Carnegie Institute of Washington’s Department of Embryology. It was here that, together with Allan Spradling, Gerry pioneered the use of transposable elements as a tool for genetic manipulation – a breakthrough that revolutionized Drosophila genetics. In 1983, Gerry moved to the University of California, Berkeley, where his group studied pattern formation and signal transduction during development of the fly eye; he later led the sequencing of the fruit fly genome, which was completed in 2000. Currently, Gerry is a Vice President at Howard Hughes Medical Institute and the Executive Director of its biomedical research institute, the Janelia Research Campus in Ashburn, Virginia. His lab probes fly brains to gain insight into the neuronal circuits underlying learning and memory, sleep regulation, visual perception and sensory integration, and Gerry remains dedicated to the development of neurobiology and genomics tools. He has been honored with a long list of awards in recognition of his contributions to Drosophila research so far.

