This interview first appeared in Development.
Melissa Little is a Senior Principal Research Fellow at the Murdoch Childrens Research Institute in Melbourne, Australia. Her lab has studied kidney development and regeneration for over 20 years, recently making notable advances in the generation of kidney organoids from human iPSCs. We chatted with Melissa about her career, her thoughts on the potential of the organoid and stem cell fields, and what she hopes to achieve during her guest editorship with Development.
 How did you first become interested in biology?
How did you first become interested in biology?
As a child I was fascinated by the world around me. I spent a lot of time camping in the Australian outback and found the plants, animals and insects just fascinating.
You started your research career as a cancer biologist. How did you eventually move on to studying kidney development?
At university I enrolled in science, assuming I was going to study botany or zoology. But I loved my first-year course in physiology, so my primary degree is actually in this area. Molecular biology was just beginning during my PhD, so I worked on the molecular basis of childhood cancer. I worked on Wilms’ tumour (a childhood kidney cancer), before the WT1 gene was isolated, and later worked on WT1 during my postdoc with Nicholas Hastie in Edinburgh. The paradigm at that time was the two-hit hypothesis, i.e. that to get a cancer you needed a hit in both copies of a tumour suppressor gene. This hypothesis was based on retinoblastoma. Whereas the RB1 gene is expressed everywhere, WT1, by contrast, is very confined in its expression during development, being restricted to the urogenital system. So, while mutations in this gene give rise to kidney cancer, they can also give rise to urogenital developmental anomalies. When I returned to Australia I continued to work on WT1. However, I was now based in a research institute with strong developmental biology, such as the work of Peter Koopman and Toshiya Yamada, so I changed direction to study WT1 and other genes in kidney development.
How important were the years that you spent as a postdoc in Edinburgh?
They had an enormous impact. I was a Royal Society Fellow, a relatively new scheme at that time. I had offers in the USA, but I had visited Nicholas in Edinburgh and really wanted to go to the MRC. The unfortunate rule of that fellowship was that after two years I had to return to Australia. But those years in the MRC were incredibly important to me. They shaped how I work as a scientist, how I interact with other scientists and how I approach answering questions in science. Nicholas was an extremely positive mentor and it was great to interact with people such as Wendy Bickmore, Ian Jackson and Veronica Van Heyningen. It was a very formative period of my career.
What are the challenges of establishing your lab in a relatively isolated country like Australia?
One of the surprising things about being in the UK was that amazing scientists wandered through the building on a regular basis. That didn’t happen in Australia. At that time there was no internet. The latest copy of Nature arrived by sea. By the time it appeared in the library it was already three months out of date. The isolation was immense. It was very difficult to keep pace with what was happening in science and to be at the forefront of anything because we were so far behind in our capacity to know what else was happening. I found that acutely oppressive when I came back to Australia in 1992.
The internet completely changed that. It made international collaboration feasible. We have real-time access to journals, we can search for articles (and there is far more published now than anyone could ever consume), we can electronically communicate in real time, and so on. Science has become much more feasible at an international level. Indeed, I have collaborations all around the globe. However, Australia is still a very long way to anywhere. To actually meet someone and talk face to face, which is quite important, you must travel. And I travel extensively. I don’t think people in the USA, for example, understand how taxing that is. I remember doing a talk in Italy where I was on the ground for less time than I was in the air. That is not that unusual, but it is pretty physically brutal. So, Australians travel a lot because they have to.
You initially established your lab in Brisbane but recently moved to Melbourne. What were the reasons for this change?
I was at the University of Queensland for 23 years, and that is a very long time to be in one place. I was in a really excellent institute, but my research evolved to have a regeneration and stem cell aspect to it, and I was relatively alone there in that respect. We were on a large academic campus but quite remote from any hospital. I did my PhD at the Queensland Medical Research Institute, which was located at a hospital, and my thesis supervisor was the head of oncology and haematology, so I had been in an environment quite closely associated with patients. I wanted to move back to an environment that had, first of all, more stem cell biology, and second, access to nephrologists. At the Murdoch Childrens Research Institute I am physically located in a children’s hospital. I now have very close associations with clinical geneticists and nephrologists, and we are setting up a clinic where we derive patient stem cell lines. That has been a really good part of the move.
What scientific questions is your lab working on at the moment?
We are the kidney development, disease, repair and regeneration lab, because we cover quite a wide range of kidney medicine. However, everything we do is underpinned by our understanding of kidney development. That is paramount. My most important message is that stem cell biology on any organ requires you to understand that organ intimately, so understanding development is key. We then use that information to direct stem cells towards a kidney fate in order to understand the relationship between development and disease. We also look at what the postnatal kidney can or can’t do to repair itself. We increasingly do human pluripotent stem cell work, but we still investigate really fundamental developmental biology questions – for example, how cells move, how they communicate with each other, how they self-organise during development, and what genes they express at what time.
Last year your lab published a high-profile paper in Nature, reporting the generation of kidney organoids from human iPSCs. Had this always been a goal of your lab?
It was a very specific and deliberate objective. Around 15 years ago, when stem cell biology was really starting out (around the time that Jamie Thomson derived the first human pluripotent stem cell lines and Perry Bartlett showed evidence that there were postnatal stem cells in the brain) I decided to change our research focus towards regenerative medicine. We started with every option on the table. We didn’t know what the postnatal kidney could do (we now know that it can repair quite well but can’t regenerate), whether there were postnatal stem cells or whether pluripotent stem cells could be differentiated to a kidney fate. In fact, cellular reprogramming was not even discussed at that time. There were a lot of things we wanted to try for kidney regenerative medicine and the differentiation of pluripotent stem cells was one of those. We were very systematic and it took quite a long time to get there.
Your ‘mini-kidneys’ paper was extensively covered in the mainstream media. How was your experience interacting with the media?
It would be lovely to control the media but no one can. Sometimes I cringe when I read what journalists write. They make broad generalisations that might lead a patient to think that a cure is around the corner, which is not true. This is the nature of the media. Some of the interviews I did were, I think, poorly represented, whereas some of them were great. You just roll with it. I don’t worry too much about those events that are less than perfect, because I can’t control them.
The organoid field is a new and exciting area. How much potential do you see in these techniques? Do you think it will be possible to build a full organ in a dish, as the media claims?
I think the media overestimates how far we have come. Organoids are fascinating and a really exciting area, but we are a very long way away from the clinic. ‘Organs in a dish’ is a funny expression, but I actually believe that although we have a long way to go, we will genuinely get there. I think there are some very short-term outcomes from this type of research that fall into the remit of both fundamental and translational biology. This is the first time that we can really start to pull apart human development. The developing human itself is not something we have had any access to, so we do have a circular problem: how can you know that what you are growing from a human cell is actually like a human? From what we can see so far, however, it is a pretty remarkable model. Hence, it really is a door into human development and that is very exciting. From a more practical point of view, I am very interested in what can be achieved with disease modelling and drug screening, even personalised drug screening. However, we need to be vigilant about how we develop these tools. I am already seeing publications claiming that organoids in a dish are accurately modelling disease but there are such challenges with interclonal and experimental variation that I think this still has to be definitively proven.
In the long term what we have is an approach where cells organise themselves based on embryological principles, and this is amazing from a fundamental science point of view. It will actually give us a handle on how cells self-organise. We make a lot of assumptions about how self-organisation works: cells differentially stick to each other or make growth factors that make other cells wander towards them. Now we have models where we can really pull that apart. From a translational point of view, this sort of information will then become an engineering challenge. How do we build these structures with a vasculature? How do we ensure a degree of anatomical correctness that will be helpful for patients? For example, the kidney organoids are currently of no value to a patient that requires a transplant as they have no exiting ureter to remove the urine. The kidney is, I would argue, the hardest thing you could ever try to generate in vitro. It is architecturally completely constrained and its function is totally dependent upon its anatomy. What we have at the moment is too small and indeed dysplastic, so there is a lot that we have to do better. But I think it is achievable with time.
You recently started your guest editorship with Development. What do you hope to achieve in this year with us?
I am really keen to encourage more of the development field to embrace what stem cells can give us, despite some reticence so far. There are enormous opportunities here to look at development in a different way. I want to open the door on investigating human development using directed differentiation, especially using these types of organogenic models (Little, 2016).
How do you see the relationship between the developmental biology and stem cell fields evolving?
I think the nexus between developmental biology, cell biology and stem cell biology is very exciting. There have been amazing advances in imaging in the developmental biology field, particularly by those working on what we would call ‘simple’ organisms, such as the worm or the fly. They have phenomenal expertise in temporal-spatial imaging, right down to the cellular level. Meanwhile, the cell biologists are building tools to look at mechano-biology and real-time reporting of pathway activity. To layer these advances on organoid creation from a pluripotent cell in a dish, which is where stem cell biology is going, is an enormously powerful approach.
What kind of papers would you like to see more of in Development?
I would like to see more papers looking at the fundamental processes governing how cells organise themselves, whether that’s during normal organogenesis in vivo or in a model in vitro. In the past, molecular biology superseded fundamental, anatomical developmental biology. Looking at aspects such as self-organisation was put to one side because it was considered too descriptive. I think it would be valuable to bring these fields back together and ask ‘what is the molecular basis of self-organisation during embryogenesis?’.
You were a member of the Australian Government’s Strategic Review of Health and Medical Research in 2013. Do you think it is important for scientists to play a role in policy?
Yes. I have had the opportunity to play a role in national scientific policy throughout my career. I was not only involved in this review but also in a seminal review of health and medical research in Australia in 1998, when I was still a young scientist. I have played roles within the Research Committee of the National Health Medical Research Council, have advised the federal government on science policy and around the debate on embryonic stem cells, and the state government on biotechnology policy. I simply see this as part of my professional obligations as a scientist. Too many young scientists forget that science is a less tangible product, not like making bricks or building boats. We are still primarily funded by tax payers, in Australia almost exclusively. The tax payers expect health outcomes, particularly in health and medical research. If we do not engage, not just in policy discussions but also in public communication, we only have ourselves to blame if the public loses interest in funding science, or worse, loses faith in scientists. One thing that I learnt very early is that there is a tendency for scientists to think that we just need to educate everyone and then they’ll understand why science is important. It is not about education. People can make decisions without facts and often do. They don’t need to be educated, they need to be engaged with. That is a very different process. It means you need to be in the room, to be having the conversation and discussing what you are doing and why.
What is your advice for young scientists?
Follow your heart. You have to be fundamentally passionate about finding an answer to a question. Have lateral vision and take every opportunity that comes your way. Don’t assume that what you read is right and question everything.
What would people be surprised to find out about you?
First and foremost, I’m a mum with two kids. In high school I was the top student in my year at art and English, not science. Perhaps this is why developmental biology is attractive to me. It is so beautiful. I actually won a prize in creative writing as a teenager. Someone recently asked me whether I still do any creative writing. I write grants and manuscripts. That is a creative process, even if it is describing data. I still paint for leisure, although not often enough.
 (1 votes)
(1 votes)
 Loading...
Loading...
 (No Ratings Yet)
(No Ratings Yet)
 The latest issue of Disease Models & Mechanisms highlights the translational impact of Drosophila research. In this issue, Moulton and Letsou review several Drosophila models of human inborn errors of development (read
The latest issue of Disease Models & Mechanisms highlights the translational impact of Drosophila research. In this issue, Moulton and Letsou review several Drosophila models of human inborn errors of development (read 
 Thummer, Edenhofer and colleagues analysed the outcomes of Nanog gain-of-function in various cell models employing a recently developed cell-permeant version of this protein. They show that Nanog blocks cellular senescence of fibroblasts through transcriptional regulation of cell cycle inhibitor p27KIP1. Read the paper
Thummer, Edenhofer and colleagues analysed the outcomes of Nanog gain-of-function in various cell models employing a recently developed cell-permeant version of this protein. They show that Nanog blocks cellular senescence of fibroblasts through transcriptional regulation of cell cycle inhibitor p27KIP1. Read the paper  Skeletal development
Skeletal development MOBP in oligodendrocyte differentiation
MOBP in oligodendrocyte differentiation
 How bacteria can affect development
How bacteria can affect development
 Planar cell polarity sigalling directs the polarization of cells within the plane of many epithelia.
Planar cell polarity sigalling directs the polarization of cells within the plane of many epithelia.  From fibroblasts to neural crest
From fibroblasts to neural crest 




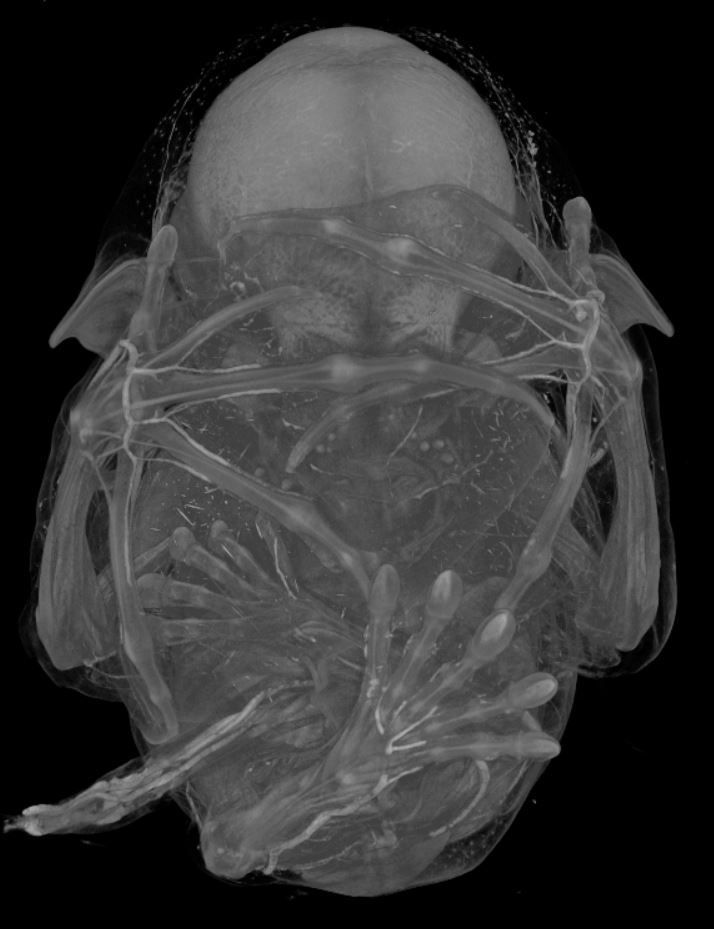

 This post is part of a series on a day in the life of developmental biology labs working on different model organisms. You can read the introduction to the series
This post is part of a series on a day in the life of developmental biology labs working on different model organisms. You can read the introduction to the series  Prof. Melissa Little – who recently published a spectacular
Prof. Melissa Little – who recently published a spectacular