The 4 September 2024 Development presents… webinar was chaired by Development Senior Editor Alex Eve and featured two talks on the topic of cardiac development and regeneration. Catch up on the talks below.
On the topic of environment, evolution and development, chaired by Development’s Guest Editor, Karen Sears (UCLA).
Wednesday 2 October – 15:00 BST
Girish Kale (University of Hohenheim) ‘Elevated temperature fatally disrupts nuclear divisions in the early Drosophila embryo’
Natasha Shylo (Stowers Institute for Medical Research) ‘Gastrulation and Left-Right patterning in veiled chameleons’
Sergio Menchero (The Francis Crick Institute) ‘Understanding temporal diversity in mammalian developmental programmes using marsupial single-cell transcriptomics’
At the speakers’ discretion, the webinar will be recorded for viewing on demand. To see the other webinars scheduled in our series, and to catch up on previous talks, please visit: thenode.biologists.com/devpres
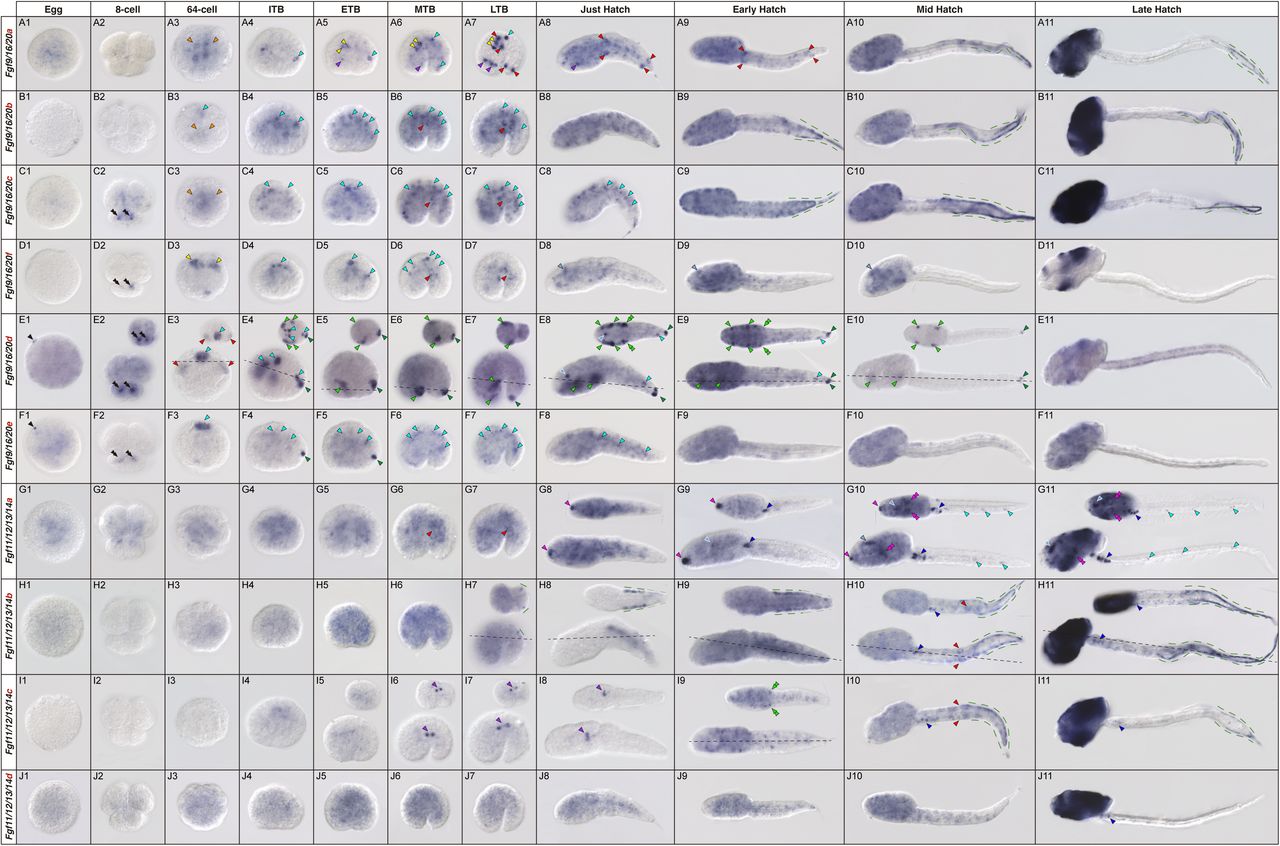
Timothy J. Duerr, Melissa Miller, Sage Kumar, Dareen Bakr, Jackson R. Griffiths, Aditya K. Gautham, Danielle Douglas, S. Randal Voss, James R. Monaghan
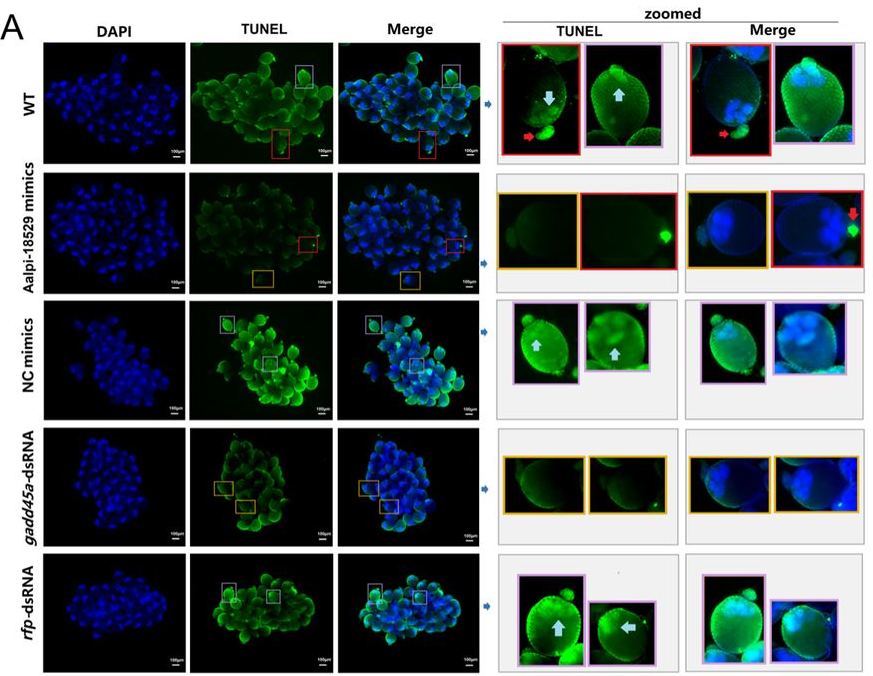
Bart Theeuwes, Luke TG Harland, Alexandra Bisia, Ita Costello, Mai-Linh Ton, Tim Lohoff, Stephen J Clark, Ricard Argelaguet, Nicola K Wilson, Wolf Reik, Elizabeth Bikoff, Elizabeth J Robertson, Berthold Gottgens
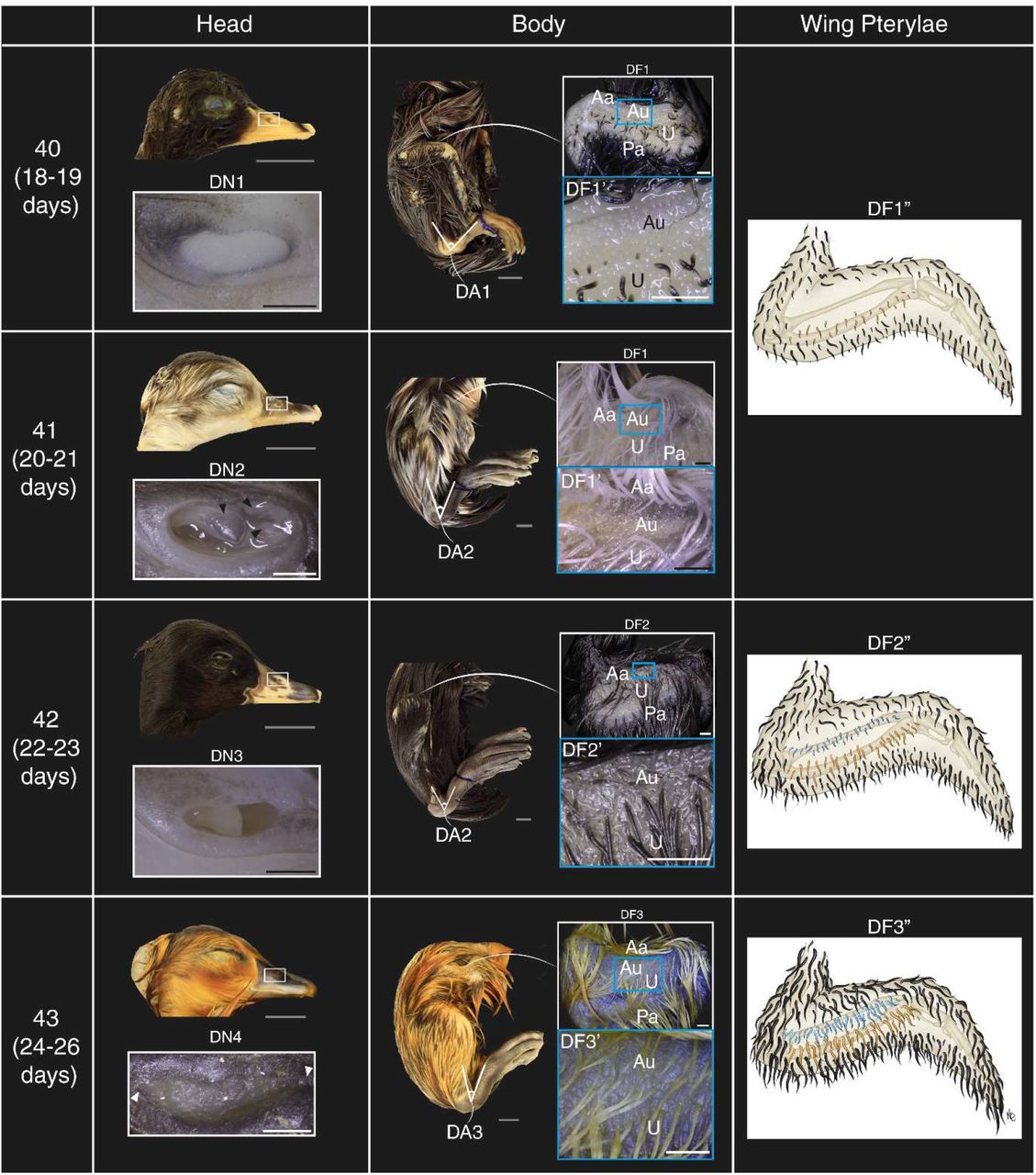
View ORCID ProfileSonja D. C. Weterings, Hiromune Eto, Jan-Daniël de Leede, Amir Giladi, Mirjam E. Hoekstra, Wouter F. Beijk, Esther J. M. Liefting, Karen B. van den Anker, Jacco van Rheenen, Alexander van Oudenaarden, Katharina F. Sonnen
Anna Fleming, Elena V. Knatko, Xiang Li, Ansgar Zoch, Zoe Heckhausen, Stephanie Stransky, Alejandro J. Brenes, Simone Sidoli, Petra Hajkova, Dónal O’Carroll, Kasper D. Rasmussen
Shreeta Chakraborty, Nina Wenzlitschke, Matthew J. Anderson, Ariel Eraso, Manon Baudic, Joyce J. Thompson, Alicia A. Evans, Lilly M. Shatford‑Adams, Raj Chari, Parirokh Awasthi, Ryan K. Dale, Mark Lewandoski, Timothy J. Petros, Pedro P. Rocha
Alessa R. Ringel, Andreas Magg, Natalia Benetti, Robert Schöpflin, Mira Kühnlein, Asita Carola Stiege, Ute Fischer, Lars Wittler, Stephan Lorenz, Stefan Mundlos, Lila Allou
Bohou Wu, Jae Hyun Lee, Kara M. Foshay, Li Zhang, Croydon J. Fernandes, Boyang Gao, Xiaoyang Dou, Chris Z. Zhang, Guoping Fan, Becky X. Xiao, Bruce T. Lahn
Sara Y. Guay, Prajal H. Patel, Jonathon M. Thomalla, Kerry L. McDermott, Jillian M. O’Toole, Sarah E. Arnold, Sarah J. Obrycki, Mariana F. Wolfner, Geoffrey D. Findlay
Yan Zhao, Andrea Fernández-Montoro, Greet Peeters, Tatjana Jatsenko, Tine De Coster, Daniel Angel-Velez, Thomas Lefevre, Thierry Voet, Olga Tšuiko, Ants Kurg, Katrien Smits, Ann Van Soom, Joris Robert Vermeesch
Atena Yasari, Monika Heinzl, Theresa Mair, Tina Karimian, Shehab Moukbel Ali Aldawla, Ingrid Hartl, Andrea J. Betancourt, Peter Lanzerstorfer, Irene Tiemann-Boege
Gal Finer, Mohammad D. Khan, Yalu Zhou, Gaurav Gadhvi, George S. Yacu, Joo-Seop Park, R. Ariel Gomez, Maria Luisa Sequeira-Lopez, Susan E. Quaggin, Deborah R. Winter
Meilin Fernandez Garcia, Kayla Retallick-Townsley, April Pruitt, Elizabeth Davidson, Yi Dai, Sarah E. Fitzpatrick, Annabel Sen, Sophie Cohen, Olivia Livoti, Suha Khan, Grace Dossou, Jen Cheung, P.J. Michael Deans, Zuoheng Wang, Laura Huckins, Ellen Hoffman, Kristen Brennand
Gal Finer, Mohammad D. Khan, Yalu Zhou, Gaurav Gadhvi, George S. Yacu, Joo-Seop Park, R. Ariel Gomez, Maria Luisa Sequeira-Lopez, Susan E. Quaggin, Deborah R. Winter
Yuki Ishii, Jessica C. Orr, Marie-Belle El Mdawar, Denise R. Bairros de Pilger, David R. Pearce, Kyren A. Lazarus, Rebecca E. Graham, Marko Z. Nikolic, Robin Ketteler, Neil O. Carragher, Sam M. Janes, Robert E. Hynds
Ian C Tobias, Sakthi D Moorthy, Virlana M Shchuka, Lida Langroudi, Mariia Cherednychenko, Zoe E Gillespie, Andrew G Duncan, Ruxiao Tian, Natalia A Gajewska, Raphaël B Di Roberto, Jennifer A Mitchell
Adam T Lynch, Naomi Phillips, Megan Douglas, Marta Dorgnach, I-Hsuan Lin, Antony D Adamson, Zoulfia Darieva, Jessica Whittle, Neil A Hanley, Nicoletta Bobola, Matthew J Birket
Luis G. Palma, Daniel Álvarez-Villanueva, María Maqueda, Mercedes Barrero, Arnau Iglesias, Joan Bertran, Damiana Álvarez-Errico, Carlos A. García-Prieto, Cecilia Ballaré, Virginia Rodriguez-Cortez, Clara Bueno, August Vidal, Alberto Villanueva, Pablo Menéndez, Gregoire Stik, Luciano Di Croce, Bernhard Payer, Manel Esteller, Lluís Espinosa, Anna Bigas
Vincent Truong, Jackson Brougher, Tim Strassmaier, Irene Lu, Dale George, Theodore J. Price, Alison Obergrussberger, Aaron Randolph, Rodolfo J. Haedo, Niels Fertig, Patrick Walsh
Nigel Kee, Mélanie Leboeuf, Silvia Gómez, Charles Petipré, Irene Mei, Salim Benlefki, Daniel W Hagey, José Dias, François Lallemend, Samir EL Andaloussi, Johan Ericson, Eva Hedlund
Abigail M. Guillemette, Guillian Hernández Casanova, John P. Hamilton, Eva Pokorná, Petre I. Dobrev, Václav Motyka, Aaron M. Rashotte, Courtney P. Leisner
Trevor Weiss, Maris Kamalu, Honglue Shi, Zheng Li, Jasmine Amerasekera, Zhenhui Zhong, Benjamin A Adler, Michelle Song, Kamakshi Vohra, Gabriel Wirnowski, Sidharth Chitkara, Charlie Ambrose, Noah Steinmetz, Ananya Sridharan, Diego Sahagun, Jill Banfield, Jennifer Doudna, Steven E. Jacobsen
Yijia Yan, Jaqueline Mellüh, Martin A. Mecchia, Hyung-Woo Jeon, Katharina Melkonian, Clemens Holzberger, Anne Harzen, Sara Christina Stolze, Rainer Franzen, Yuki Hirakawa, Ana I. Caño Delgado, Hirofumi Nakagami
Carlos Henrique Cardon, Victoria Lesy, Catherine Fust, Thales Henrique Cherubino Ribeiro, Owen Hebb, Raphael Ricon de Oliveira, Mark Minow, Antonio Chalfun Junior, Joseph Colasanti
Michael C Wilson, Alexander H Howell, Anika Sood, Youngwoo Lee, Pengcheng Yang, Heena Rani, Elena Yu, Eileen L. Mallery, Sivakumar Swaminathan, Corrinne E. Grover, Jonathan F. Wendel, Olga A. Zabotina, Jun Xie, Chelsea S. Davis, Daniel Szymanski
Martijn J. Jansen, Stuart Y. Jansma, Klaske M. Kuipers, Wim H. Vriezen, Frank F. Millenaar, Teresa Montoro, Carolien G.F. de Kovel, Fred A. van Eeuwijk, Eric J.W. Visser, Ivo Rieu
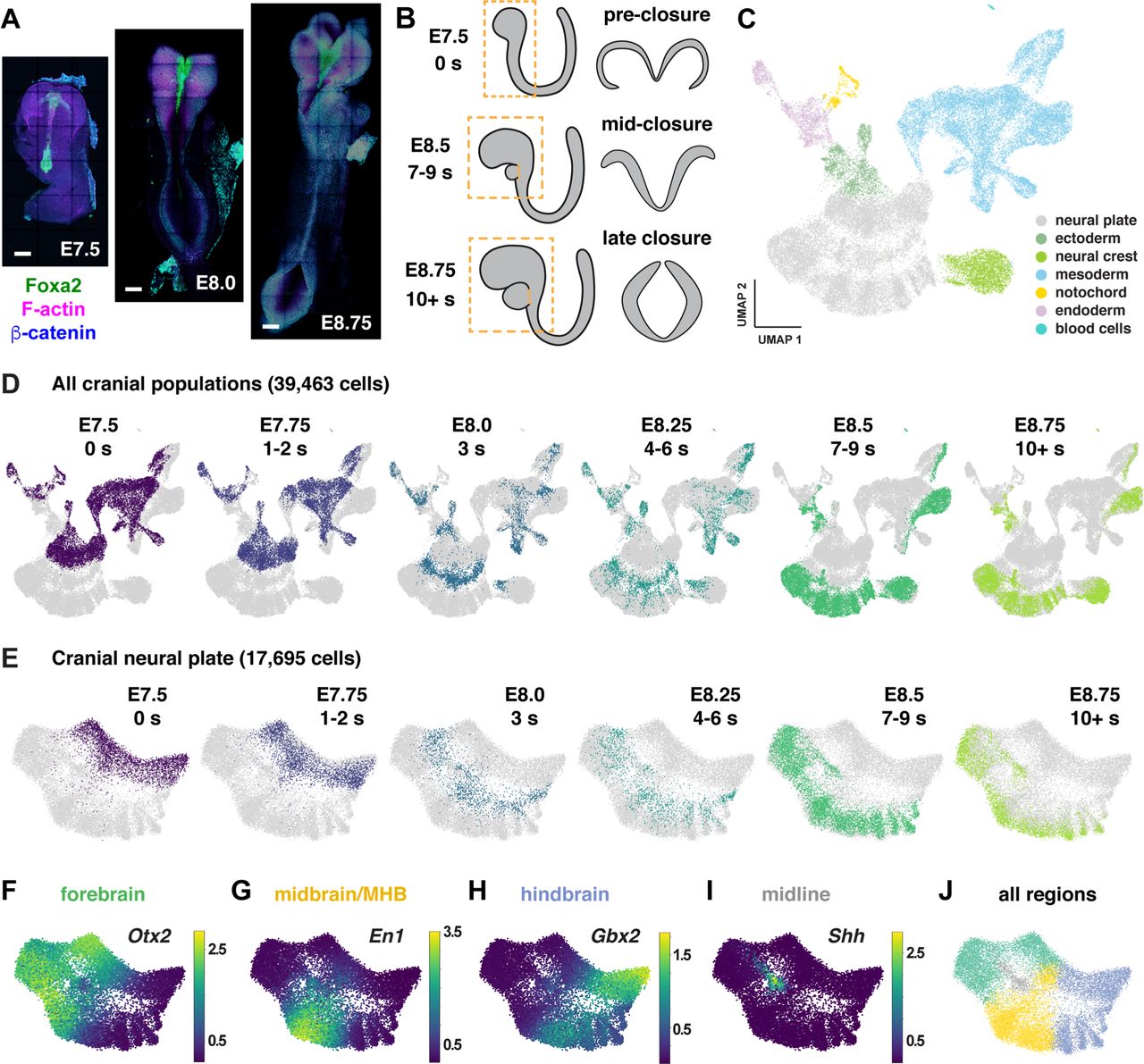
Shota Nakanoh, Despina Stamataki, Lorena Garcia-Perez, Chiara Azzi, Hayley L Carr, Alexandra Pokhilko, Lu Yu, Steven Howell, Mark Skehel, David Oxley, Simon Andrews, James Briscoe, Teresa Rayon
Gaspar Sánchez-Serna, Jordi Badia-Ramentol, Paula Bujosa, Alfonso Ferrández-Roldán, Nuria P. Torres-Águila, Marc Fabregà-Torrus, Johannes N. Wibisana, Michael J. Mansfield, Charles Plessy, Nicholas M. Luscombe, Ricard Albalat, Cristian Cañestro
Huanhuan Liu, Anupama Binoy, Siqi Ren, Thomas C. Martino, Anna E. Miller, Craig R. G. Willis, Shivakumar R. Veerabhadraiah, Abhijit Sukul, Joanna Bons, Jacob P. Rose, Birgit Schilling, Michael J. Jurynec, Shouan Zhu
Lisa K. Iwamoto-Stohl, Aleksandra A. Petelski, Maciej Meglicki, Audrey Fu, Saad Khan, Harrison Specht, Gray Huffman, Jason Derks, Victoria Jorgensen, Bailey A.T. Weatherbee, Antonia Weberling, Carlos W. Gantner, Rachel S. Mandelbaum, Richard J. Paulson, Lisa Lam, Ali Ahmady, Estefania Sanchez Vasquez, Nikolai Slavov, Magdalena Zernicka-Goetz
Caroline Hookway, Antoine Borensztejn, Leigh K. Harris, Sara Carlson, Gokhan Dalgin, Suraj Mishra, Nivedita Nivedita, Ellen M. Adams, Tiffany Barszczewski, Julie C. Dixon, Jacqueline H. Edmonds, Erik A. Ehlers, Alexandra J. Ferrante, Margaret A. Fuqua, Philip Garrison, Janani Gopalan, Benjamin W. Gregor, Maxwell J. Hedayati, Kyle N. Klein, Chantelle L. Leveille, Sean L. Meharry, Haley S. Morris, Gouthamrajan Nadarajan, Sandra A. Oluoch, Serge E. Parent, Amber Phan, Brock Roberts, Emmanuel E. Sanchez, M. Filip Sluzewski, Lev S. Snyder, Derek J. Thirstrup, John Paul Thottam, Julia R. Torvi, Gaea Turman, Matheus P. Viana, Lyndsay Wilhelm, Chamari S. Wijesooriya, Jie Yao, Julie A. Theriot, Susanne M. Rafelski, Ruwanthi N. Gunawardane
Alessandro Dei, Carlemi Calitz, Joep Korsten, Nina Johannesson, Eline Freeze, Allen Eaves, John Stingl, Ryan K Condor, Wing Chang, Dasja Pajkrt, Katja C. Wolthers, Adithya Sridhar, Salvatore Simmini
Yinan Wan, Jakob El Kholtei, Ignatius Jenie, Mariona Colomer-Rosell, Jialin Liu, Joaquin Navajas Acedo, Lucia Y. Du, Mireia Codina-Tobias, Mengfan Wang, Ahilya Sawh, Edward Lin, Tzy-Harn Chuang, Susan E. Mango, Guoqiang Yu, Bogdan Bintu, Alexander F. Schier
Jialin Liu, Sebastian M. Castillo-Hair, Lucia Y. Du, Yiqun Wang, Adam N. Carte, Mariona Colomer-Rosell, Christopher Yin, Georg Seelig, Alexander F. Schier
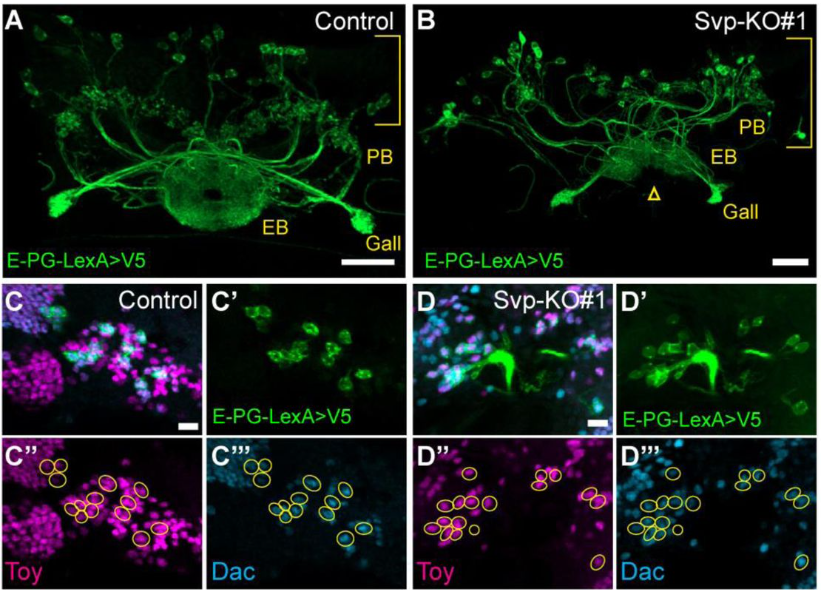
Convergent evolution — the independent emergence of analogous structures among species whose last common ancestors lacked the trait— has long fascinated me. This phenomenon represents an exciting opportunity to study the genomic constraints that shape organisms during development to produce specific forms and functions. In the fall of 2018, as I was searching for a PhD program, I heard a talk by Dr. Ricardo Mallarino. He was interested in studying one such example of convergence, the evolution of the patagium, a skin membrane that, like a wingsuit, allows animals to glide through the air as a means of locomotion. Of particular interest was the patagium of a small marsupial possum: sugar gliders. The patagium has evolved many times across the tree of life including rodents, primates, marsupials, lizards, and frogs. Marsupials however, offered a uniquely tractable group to study how similar novel morphological traits evolved independently, as three closely related possums had acquired this adaptation. Here I will cover the findings of our recent publication.
Adult sugar glider extending its patagium (red arrow).
Biology strikes back: identifying candidate regulatory elements
Previous RNAseq-based exploration of the developing patagium indicated that a major difference between the early patagium and neighboring skin was the differential regulation of an early gene regulatory network1. Given this, I set out to identify cis-regulatory elements (CREs) which had similar patterns of evolution in gliding species, which may harbor the causative changes driving differential gene regulation, evidenced by previous work2,3. We produced 15 new marsupial genomes which included gliding (sugar glider, greater glider, and feathertail glider) and non-gliding species. To identify cis-regulatory elements in the developing patagium tissue, we used ChIP- and ATAC-seq, which when used in combination provided a candidate list of active and poised CREs. Then using both our biological data and our genomes, we measured the rate of nucleotide change across these identified cis-regulatory elements, giving us an indication of which CREs are experiencing selection as these species have evolved their patagium. Through this analysis, we identified thousands of candidate glider accelerated regions (GARs)— elements which showed a substantial increase in nucleotide substitution. By focusing only on the GARs that showed shared patterns across the three gliding species we analyzed, we hoped to find a smaller pool of CREs that could be involved in the evolution of the patagium. We were surprised to find that not a single CRE matched that description. Then in our darkest times, a moment of brilliance: what if the cause was not a single CRE but instead different CREs across species all regulating the same gene. We had previously conducted Micro-C on the developing patagium and thought to use this data and identify topologically associating domains (TADs) to inform an analysis of GAR distribution and abundance. Using these TADs, we assigned CREs to the genes that were in the same TADs and asked if any gene had an overabundance of GARs.
Return of a key gene
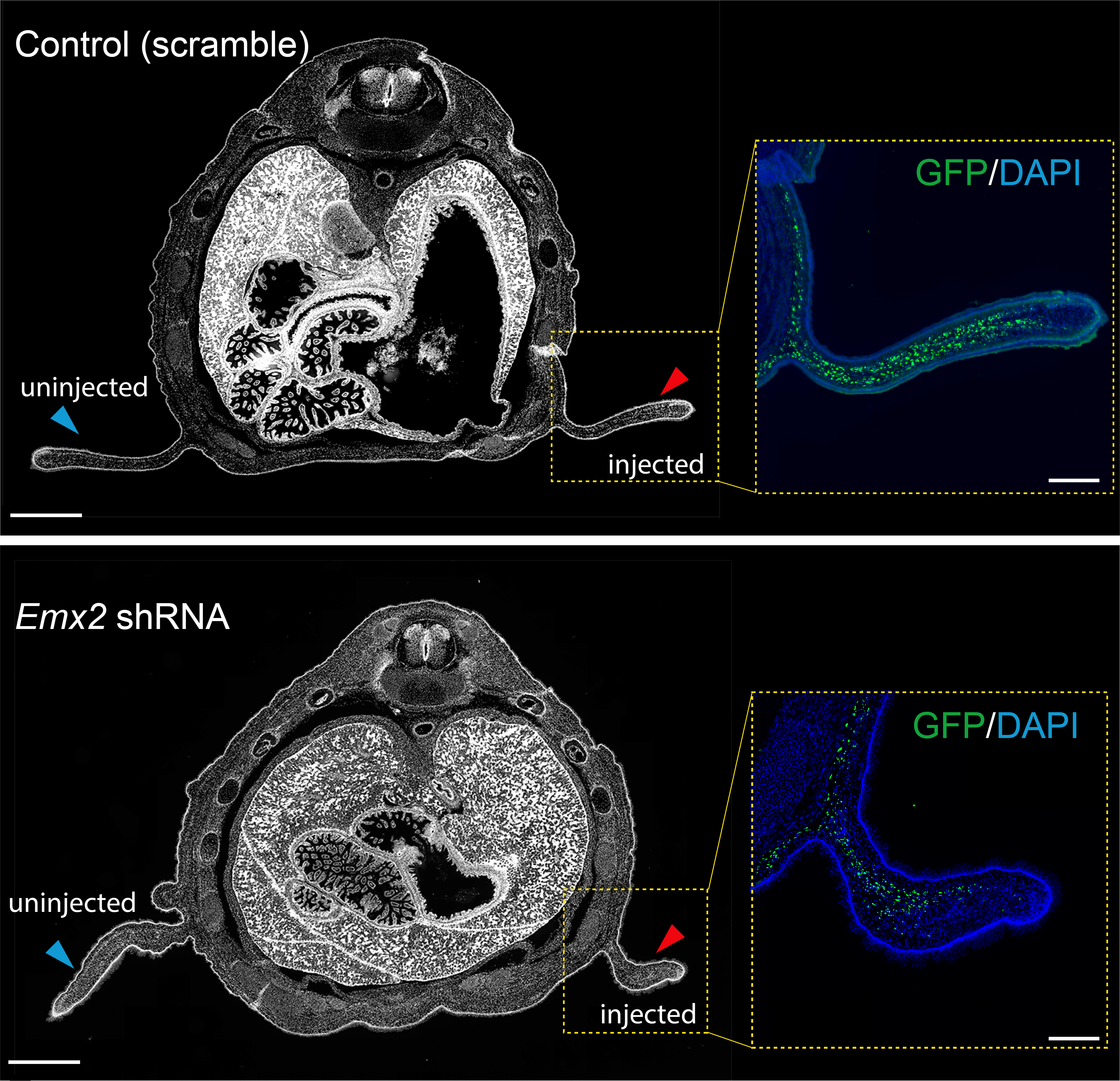
Previous work from our group uncovered that the sugar glider patagium develops through the deployment of a conserved network of genes, and that Wnt5a is heavily involved in the early development of the gliding membrane1. One of the other genes identified was the transcription factor Emx2. It just so happened, that our analysis for GAR enrichment identified Emx2 as our strongest candidate, having GARs from each of the three gliding species in its vicinity. We produced an shRNA lentivirus for Emx2 and began testing the effects of downregulating Emx2 in the developing patagium. Taking advantage of marsupial biology, that is they give birth to their young, or joeys, quite early in their development, we could then probe the early patagium. We conducted injections into the patagium primordium of sugar glider joeys and found that indeed downregulation of Emx2 caused a decrease in the area of the developing patagium. This was one of my favorite experiments and served to remind me how fascinating working in science can be. For some time, I was one of the only people in the world who knew that lack of Emx2 led to incorrect patagium growth. We then explored how Emx2 may be regulated. We had several candidates to test but initially focused on just two, one was positioned in what we presumed to be the promoter of Emx2 while the second seemed to be a distal enhancer located 1mb away from the Emx2 promoter but with a strong contact loop in the Micro-C dataset. The former was accelerated in the sugar glider and the latter accelerated in the feathertail glider. We wanted to test if the acceleration observed had an effect on the element’s ability to regulate expression and so we decided to use luciferase assays in an immortalized sugar glider cell line. This experiment works by placing your CRE of interest upstream of the luciferase gene (which originates from fireflies) to measure the amount of fluorescence produced by the cells to see if your CRE has regulatory function. In our case, our results indicated that the distal enhancer had accumulated changes that made it a stronger enhancer in the feathertail glider compared to its non-gliding sister and the sugar glider. We later found that the other gliding species, the greater glider, also had an enhancer that showed the same pattern.
Emx2 shRNA injection results in a decrease in patagium size
Duo of fate: Our two favorite genes are important for patagium development
Now we knew that Emx2 had a phenotypic effect on the patagium and had some clues as to how it could end up being highly expressed in gliding species, but I became interested in what was happening at the molecular level in the patagium when we disrupted Emx2 expression. We did another round of shRNA injections, and this time collected the tissue for RNA sequencing. We found that many of the genes that were normally upregulated in the patagium when compared to surrounding skin were now downregulated. Among the ~400 genes downregulated, Wnt5a was one of the more strongly affected genes. This prompted us to investigate if Emx2 regulates all these genes directly and specifically how it may be regulating Wnt5a. We did an Emx2-ChIP-seq experiment and found that many genes did indeed have Emx2 binding sites, and we were able to identify multiple binding sites in the Wnt5a promoter. An example of great peer reviewing led to another of my favorite experiments as we set out to test the Emx2 binding sites found in the Wnt5a promoter. We again used luciferase this time testing two versions of the same promoter, one was unaltered while in the second we mutated the Emx2 binding sites. We found that the loss of these sites led to a complete loss of regulatory ability. Then to see if Emx2 was responsible for activating this promoter we co-transfected an Emx2-producing plasmid with our luciferase reporter plasmids. This experiment showed that the wild-type promoter increased its production of luciferase nearly 4-fold when Emx2 was present.
Emx2 awakens a conserved pathway
Our final goal for the paper was to establish if the spatial expression and function of Emx2 was novel to sugar gliders or if it was conserved in non-gliding mammals. We decided to test this hypothesis in mice, a much more amenable system for testing overexpression of genes. We found that Emx2, as reported previously4, was expressed in mice in a similar spatial pattern as that of the patagium, however this expression was only present for ~2 days whereas in the developing sugar gliders it was present for at least 14 days. To test whether overexpression of emx2 was sufficient to produce early patagium phenotypes, like we had previously observed with Wnt5, we extended the duration of Emx2 expression in mice while maintaining its endogenous spatial pattern. This however resulted in mice whose forebrain grew uncontrollably and resulted in non-viability; previous work had implicated Emx2 in brain development5. Therefore, we restricted the overexpression of Emx2 to only the skin. We found that indeed this overexpression was capable of recapitulating phenotypes observed in the early patagium such as increased cell proliferation, density, and the thickening of the epidermis1. These experiments showed that Emx2 has a conserved role in driving proliferation, potentially via regulation of the Wnt pathway, further indicating that evolution has re-used existing cellular programs to evolve a new adaptation.
The last remarks
In my opinion, the key message of this paper is that the evolution of convergent traits can occur independently via similar pathways/mechanisms, but the path to get there can be different. Our work showed that the redeployment of a shared developmental pathway can be an effective mechanism by which adaptations evolve.
I am very happy to see this work published; it took many years to get to this point. There were many bumps along the way, and it was the culmination of the hard work of many people involved. I want to thank the reviewers for their candid and helpful words, truly they made the paper better than when we first submitted. As someone who finds great joy in working on emerging model systems and has received countless advice in the past to just work on “model” organisms, I am incredibly pleased with how this paper is received. I continue to work on non-model systems with fascinating biology now as a postdoc and advise anyone who is interested in working on new models to go for it! It is rewarding to work on questions that can only be asked in a new system and uncover and share new findings.
References
1 Feigin, C. Y. et al. Convergent deployment of ancestral functions during the evolution of mammalian flight membranes. Science Advances9, eade7511 (2023). https://doi.org/doi:10.1126/sciadv.ade7511
2 Booker, B. M. et al. Bat Accelerated Regions Identify a Bat Forelimb Specific Enhancer in the HoxD Locus. PLOS Genetics12, e1005738 (2016). https://doi.org/10.1371/journal.pgen.1005738
3 Capra, J. A., Erwin, G. D., McKinsey, G., Rubenstein, J. L. R. & Pollard, K. S. Many human accelerated regions are developmental enhancers. Philosophical Transactions of the Royal Society B: Biological Sciences368, 20130025 (2013). https://doi.org/doi:10.1098/rstb.2013.0025
4 Pellegrini, M., Pantano, S., Fumi, M. P., Lucchini, F. & Forabosco, A. Agenesis of the Scapula in Emx2 Homozygous Mutants. Developmental Biology232, 149-156 (2001). https://doi.org/https://doi.org/10.1006/dbio.2001.0159
5 Yoshida, M. et al. Emx1 and Emx2 functions in development of dorsal telencephalon. Development124, 101-111 (1997). https://doi.org/10.1242/dev.124.1.101
We need your help to optimise the SAFE Labs Handbook before it is disseminated throughout the academic community. Please complete our this survey to help optimize this tool for the academic community.
This handbook is an outcome of the 2024 SAFE Labs workshop, where new bioscience group leaders from across Europe discussed Starting Aware, Fair, and Equitable Labs. The primary goal of this workshop (funded through the UCL Global Engagement award) was to better-understand the common, and divergent, problems faced by new researchers trying to build successful, equitable, fair, and environmentally sustainable labs with a positive research culture. There were no scientific talks as part of the program (there are plenty of meetings for that!).
Through this workshop, we realised that a major obstacle to improving lab culture was the lack of any resource with actionable “commitments” for group leaders that could be implemented in the absence of institutional support. Existing resources raised important issues, but lacked specific actions for group leaders to take.
The thirteen attendees unanimously agreed to collaborate on creating a SAFE Labs Handbook, covering commitments that span different topics: Policies, Teams, and Careers. Of course, there are many effective strategies to run a team, reflecting differences across groups, institutions, and countries. Thus, the Handbook is not prescriptive; rather it aims to increase transparency and minimise expectation mismatch between the group leader and lab members to create positive and equitable lab culture. Commitments may require a policy to be documented, but the content of that policy is decided by the group leader.
No commitment relies on institutional support to implement, and all commitments are actionable, and can be verifiably implemented. This is because:
Implementable commitments are the most helpful to group leaders and lab members.
Verifiable commitments allow for accountability and specific feedback.
For example, “I commit to supporting a healthy work-life balance for my lab members” will not be in this handbook because the commitment cannot be verified.
The handbook does not represent an optimal or exhaustive template: it is the product of thirteen group leaders who are all passionate about improving lab culture, and are all fallible. Although this handbook will not solve every problem, we hope it takes a significant step toward addressing key challenges faced in daily lab life. The handbook will improve through community feedback and further dedicated workshops.
However, to ensure that this ensure that the SAFE Labs Handbook reflects the needs of our entire community, we need a diversity of feedback on the initial document: from all career levels and countries. The results will directly influence the content of the handbook and ensure that the commitments are something that are universally valued.
Please take the time to complete this survey, which can take less than 15 minutes (or 30 minutes if reading the commitments in more detail).
We hope the SAFE Labs Handbook becomes a valuable tool to community. Our intention is that labs implementing all commitments would feature the SAFE Labs logo on their website. This demonstrates to potential and existing lab members that the group leader is committed to creating a SAFE Lab.
We have secured funding to hold a 2025 workshop to further improve, expand, and refine this initiative. If you are interested in participating, please add your email address at the end of the survey.
Thank you for helping us to make the SAFE Labs Handbook as useful as possible!
The SAFE Labs organisers,
Pip Coen, Letizia Mariotti, Stéphane Bugeon, and Federico Rossi
[This report is co-authored by Alex Neaverson, Rachel McKeown and Anna Guixeras Fontana.]
The clock struck 1PM in the UK, the Zoom room opened, and in flooded over 50 scientists. All were drawn together by a passion for effective science communication and were here to sharpen their skills. It was time for the inaugural SciCommConnect workshop, a collaboration between the three community sites of The Company of Biologists (FocalPlane, The Node and preLights).
Though hosted from the UK, this was truly science without borders. Participants tuned in from as far apart as Seattle, USA and Melbourne, Australia. The fact that some were joining as early as 5AM and as late as 10PM local time showcased just how dedicated and engaged the participants were to science communication, which only became more apparent as the event progressed.
With all participants settled and eager to dive in, our hosts welcomed us to the first SciCommConnect event. Launching straight in, we began with a few tips and tricks on the spoken and the written word from two true science communication pros.
What do your research and raw vegetables have in common?
As scientists, we understand our work is of vital importance to us and those in our community. Your immediate motivation is probably your thesis, that conference talk, that paper, or that grant proposal. But how often do you take a step back to see the bigger picture – what are we, as biologists, working towards as a sector? This is what Jamie Gallagher, award-winning freelance communicator, calls the “collective endeavour”; learning how to frame your research in a way that the general public can understand and appreciate is a skill we should all aim to harness.
But why should we care about what the public think of our research? Jamie posed this question to the audience and it produced many insightful answers, including:
People have a right to knowledge
To help prevent the spread of misinformation – openly sharing accurate information helps the public put their trust in scientists
To share the information relevant to human health and medicine
Presenting our science to wider audiences can help to de-bunk these stereotypical views about what a scientist should look like.
The public indirectly fund a lot of our research through taxes
How do we go about turning our data into an interesting, engaging, and comprehensible presentation? Consider your research to be the equivalent of raw vegetables: healthy and good for all of us, but not everyone enjoys consuming them. One way of making people eat them is to cut them into bite-sized pieces – the same can be said for our research. Even better, is to cook the vegetables and add some herbs and spices, creating a delicious meal. In the context of a scientific presentation, the herbs and spices are the extra bits that add ‘flavour’: these include things like emotion, storytelling, pauses, analogies, questions, and varying the tone and pace of your speech. You must be careful though – just as adding too much spice can ruin a meal, adding too many of these flavour elements can ruin your presentation.
Imagine your grandma in the audience
The thought of presenting to a large audience conjures up feelings of anxiety and dread in many of us. Jamie talked us through his top tips for delivering a great scientific presentation, some of which I found quite surprising, as they challenged my preconceived ideas about what an effective presentation should include:
1. Introduce yourself the same way every time – knowing exactly what you’re going to say at the beginning will help to calm your nerves, so take the time to rehearse this part.
2. Nerves are normal, and only become a problem if others can tell that you are nervous. Some tips for dealing with nerves include speaking slower than you think; taking a sip of water to give your thoughts time to catch up; telling yourself you’re excited, not nervous; and my favourite – imagining your grandma cheering you on in the audience!
3. Eye contact is not the pièce de résistance for an engaging presentation. The model for good communication is inherently ableist, and doesn’t account for those who are neurodivergent or disabled. These tropes should be challenged, as they do not take away your ability to communicate effectively.
4. Instead of making written notes for each slide of your presentation, consider recording yourself talking, then transcribing it into text to create a more natural sounding script.
5. A general audience will not care about your data – only about what it means. Take the time to re-frame your findings in a way they will understand, and share only as much as is absolutely necessary.
Over to us – Three-minute research talks
After absorbing all of Jamie’s expert advice on presenting, the ball was then placed in our court. Twelve back-to-back research presentations, just three minutes each. In that time, we had to showcase not only what our research is about, but do so in an accessible and engaging way. Let the three-minute research competition begin!
Each participant was armed only with a single PowerPoint slide, but this by no means limited their ability to communicate. Their scientific stories were brought to life verbally in short presentations where each put into action some of the points we’d all been discussing earlier in the programme. Similes and metaphors were used to great effect, with the brain being compared to the contents of your fridge, tissue movements to toothpaste and the cellular environment to a well-known movie franchise (The Matrix). Emotive and personifying language were particularly powerful techniques to drive home the importance and significance of the projects – think of ‘healing a broken heart’ and cells having ‘social’ and ‘lonely’ personalities. Short, impactful sentences, often to summarise or conclude, left lasting memories in the audience. ‘I’m imagining a world without osteoarthritis’ – now we’re imagining it too.
The topics covered were incredibly diverse, but it was fascinating to see common threads emerge as the competition progressed – some were united by their love of zebrafish, others by their fascination with the cytoskeleton. The presentation structures also shared some key elements – opening with an introduction to the wider field, establishing how their own research contributes to it, and rounding up with the overall significance of the work. Jamie, who moderated the competition, was so engaged by the presentations that he could not help but ask questions to each participant after their time was up. His eagerness to dive more into the story was a sure sign that the audience had effectively been captured. The enthusiasm that each participant brought to the stage was infectious, and by the time the competition drew to a close, it felt as if no time at all had passed.
From twelve competitors, two emerged victorious, one chosen by Jamie and the other by audience vote. The worthy winners both delivered fantastic presentations, accompanied by visually striking slides (one being hand-drawn by a clearly talented digital artist, the other boasting stunning microscopy images). Despite this, all talks were highly praised not just by the event hosts, but by all of the fellow participants. The Zoom chat box was flooded with compliments and positivity, with all of us cheering each other on. As a result, by the time the competition drew to a close, we all felt like winners.
The 5 Rules of Fight ClubDevBiolWriteClub
After learning all about how to give an engaging scientific talk, the second half of the event focused on another form of science communication — writing. John Wallingford, professor at UT Austin and a regular contributor on The Node, shared some insights on how we can improve as science writers.
The first (and second) rule of DevBiolWriteClub is to Do The Work. As with any hobby or skill, you cannot expect to improve your writing without practice. This should be intentional – set aside time each day to write, either setting yourself a word limit or a timer. Surprisingly, this was a new concept to me, as someone who has only ever written with a specific goal in mind – a report, an article, a paper – and never purely for the sake of writing.
The third rule is to Revise and Edit: Again, and Again and Again. You will likely have many versions by the time you are finished writing – keep track of these carefully, and never permanently delete anything! You never know when you might want to come back to something you wrote earlier.
The fourth rule is to Read with Intent. So much of how we write is influenced by what and how we read. For this reason, plan to read every single day, and read widely – not just papers, but novels, poetry, comics, or whatever interests you. While you read, reflect on what kind of writing is most and least effective for its purpose.
The fifth rule is: You Cannot Do It Alone. Writing may seem like a lonely endeavour, but it doesn’t have to be. In fact, getting involved with writing groups or having a writing buddy can be very effective in providing accountability and feedback. Normalise sharing your rough first drafts with friends and colleagues – most people do not share their work until they think it is perfect, but by this point you will be attached to it and less receptive to feedback.
Over to us – Writing sprints
After we were all inducted into the DevBiol Write Club, it was our turn to put the writing tips from John into practice.
The collaborative writing sprint at SciCommConnect brought together diverse groups of participants, each of them focusing on eight critical topics in science communication and dissemination. The participants were split into eight breakout rooms on Zoom and over a dedicated hour, each group worked intensively to draft articles on their respective subjects. These subjects include 1/ Communication between Developmental Biologists and the Public, 2/ The Importance of Model Organism Databases, 3/ SciCommConnect Workshop Report, 4/ Sci-Comm “Behind-the-Scenes”, 5/ Science Writing & AI, 6/ Open Science & Preprints, 7/ AI in Microscopy and 8/ Frugal Microscopy.
The authors of this SciCommConnect report, that you, the reader, are reading now, formed one of the writing sprint groups. Within the hour, we drafted a report providing an overview of the SciCommConnect workshop, summarising key presentations and interactive sessions.
To find out how other writing sprints went, we reached out to the other participants after the event. The Node groups focused on producing posts about ‘Communication between Developmental Biologists and the Public’ and ‘The Importance of Model Organism Databases’. Joyce Yu, who led the first group, reflected, “The topic was very broad, and we all had many ideas, but we managed to narrow down the scope of the article and come up with the different sections of the piece.” Similarly, Beatrice and Mansi, participants from the second group, shared, “We have now drafted a piece calling for the continued support of Model Organism Databases. We’re still editing it.” We look forward to seeing their finished articles on the Node soon.
Meanwhile, one of the preLights writing groups worked together to draft a preLights post about a preprint on SciComm ‘Behind the scenes’. The moderator of this group, Martin Estermann, said, “My first attempt at moderating a preLights writing sprints exceeded my expectations; it involved engaging discussions on the status and value of science communication and produced an almost complete preLights post within an hour. This was definitely a collaborative effort.” You can now read their finished preLights article.
In a similar way, the other preLights writing groups created preLights articles on ‘Science Writing & AI’ and ‘Open Science & Preprints’ both of which are now available to read on preLights. Jennifer Ann Black, the moderator of one of these groups, reported, “Together, we enjoyed a constructive session chatting over the use of AI in science. For or against it, AI is here to stay, and we need to find ways of accommodating it.” Reinier Prosee, who led the group on Open science and preprints, added, “We exchanged some relevant personal experiences and discussed a preprint showing that Open Science practices can lead to a higher visibility of research papers. We all agreed though that the potential benefits of adopting Open Science practices go beyond citation metrics.”
Finally, the two FocalPlane writing groups explored the impact of artificial intelligence in microscopy and the concept of ‘frugal microscopy’. Helen Zenner, who led the AI in microscopy group, noted, “It is a huge topic, but we have a few ideas of how we can present our discussion on FocalPlane, so look out for our future posts!”
The collaborative writing sprint at SciCommConnect showcased the power of teamwork in science communication. By bringing together experts and enthusiasts from various fields, the sprint not only produced insightful reports but also strengthened the community’s commitment to effective and inclusive science communication. As the reports are finalised and shared on The Node, preLights, and FocalPlane platforms, they will contribute valuable perspectives and knowledge to the broader scientific community.
Final thoughts
SciCommConnect brought together a group of ~50 like-minded individuals from across the world to share their research and learn how to improve their skills in doing this. Something that was clear from the outset was just how supportive an environment can be created, even over Zoom – each 3 minute research talk was followed by dozens of messages giving praise to different aspects of each talk. It is difficult to anticipate the level of engagement that you’ll get in online workshops, but in this case it truly felt like the presenters were talking to a group of friends. Science is international, yet opportunities to network with researchers from across the world are relatively infrequent and can sometimes depend on opportunities to travel to conferences. Online events enable us to be brought together with far fewer financial and environmental caveats. While many online seminars are passive in nature, SciCommConnect shows how active engagement and audience participation can, with a little forethought and planning from a dedicated organising team, be enabled. A strong sense of community was cultivated, despite the audience stretching over almost 10,000 miles.
In the paper “Mechanics of human embryo compaction”, Jean-Léon Maître and colleagues mapped the mechanical forces controlling compaction during human embryogenesis. Find out more about the behind the paper story from corresponding author Jean-Léon Maître and first author Julie Firmin, who is a medical doctor specializing in Assisted Reproductive Technology.
How did the project start?
Jean-Léon: In 2017, shortly after I had started my team at Institut Curie, Edith Heard, who was the director of my department, introduced me to Catherine Patrat with whom she had worked a few years before. Catherine heads the largest IVF clinic in France at Hospital Cochin, which is only a few minutes’ walk from Curie. Fortunately, Catherine got interested in our research and we decided to start with the simplest possible project about human embryo mechanics, which is compaction. It was then only a matter of getting the permit, money and the right person for the project. Aaaaand, it was the end of 2019 🙃.
Julie, what led you to join Jean-Léon’s lab?
Julie: I am a medical doctor, specialized in Assisted Reproductive Technology (ART). ART is still a relatively recent medical discipline—the first IVF births date back to 1978—and success rates after embryo transfer are only around 30%. Meanwhile, a recent WHO report estimates that one in six people is or will be affected by infertility at some point in their lives. My attraction to research stems from the conviction that better understanding the embryo is crucial for improving ART.
I discovered the work conducted by Jean-Léon on preimplantation embryos during my second year of Master degree, which I was pursuing alongside my medical studies. At that time another student, co-supervised by Jean-Léon and Professor Catherine Patrat, head of the ART department at Cochin Hospital in Paris, was studying compaction of human embryos using time-lapse images of embryos obtained after IVF. Using quantitative measurements inspired by knowledge of mouse embryo mechanics, the student aimed at identifying more robust parameters than the qualitative ones routinely used in ART laboratories to select which embryo to transfer. This piqued my interest in the basic research being conducted in Jean-Léon’s laboratory and motivated me to learn the biomechanical approaches they use to study embryonic development.
Following an internship as a medical resident in Professor Patrat’s ART laboratory, during which I was able to develop my skills as an embryologist in a human IVF unit, I had the opportunity to join Jean-Léon’s team full-time at the end of 2019 as a PhD student.
What was known about the mechanics driving human embryo morphogenesis before your work? Since the advent of in vitro culture, the morphology of the human embryo has been observed and described in numerous studies. In fact, along with cell number, the morphology of the human embryo is a primary determinant in clinicians’ assessment of the implantation potential of human embryos. Despite the importance of human embryo morphology for its development, we know very little about the mechanisms responsible for shaping the human embryo and can only extrapolate what we know from model organisms.
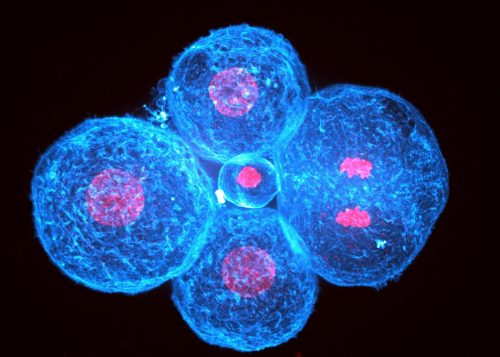
One of our favorite picture of a cleaved human embryo at 4 cell-stage showing one mitotic cell that just separated its chromosomes (the right cell)
Human embryonic development starts with cleavage divisions without much change in the morphology of the blastomeres. Human morphogenesis starts with compaction and is the first step leading to the formation of the blastocyst, which will implant the embryo into the maternal uterus. During compaction, blastomeres come closer together, forming a tighter structure. The compaction process was long thought to be driven by increased cell adhesion via modifications of the calcium-dependent cell-cell adhesion machinery, particularly CDH1-dependent adhesion, since removing extracellular calcium prevents CDH1 binding and causes embryos to decompact. However, the possibility that cells could also use cell contractility to pull themselves together, as suggested by 20 years of study of animal morphogenesis, had not been explored in human embryos.
Julie Firmin dissecting human embryos out of their zona pellucida with a dual pipette aspiration setup under Jean-Léon Maître’s supervision.Focus on dual pipette aspiration setup: here you can observe the two arms installedon the microscope with one of them bringing the pipette required for precise embryo manipulation.
Can you summarise the key findings in a paragraph?
To measure the forces responsible for human embryo compaction, we used micropipette aspiration. This technique is mostly non-invasive, compatible with long-term development and we had used it extensively with mouse embryos, i.e. we knew it would work right away with precious donated human embryos. Micropipette aspiration reveals increased forces at the cell surface and stable forces at cell-cell contacts throughout compaction. Inhibition of cell contractility or cell-cell adhesion shows that only contractility controls the large forces present at the cell surface. Therefore, as observed in compacting mouse embryos and during morphogenetic processes of other animals, cell contractility is responsible for pulling cells together and plays an important role in shaping the embryo. Interestingly, some human embryos show compaction defects, which we can relate to abnormal contractile forces. Together, these experiments further confirm the key role of contractility in animal morphogenesis and identifies defective contractility as a potential cause for failed human embryo development.
How do the mechanics of mouse and human embryo morphogenesis differ?
At the scale of the embryo, it is fascinating to see how embryonic structures can be both conserved between mice and humans while still have their own unique shapes. For example, although both species form a blastocyst, the human blastocyst has an inner cell mass that protrudes more into the blastocoel than that of the mouse. This strongly suggests that the mechanical properties of these two species differ.
Considering compaction, the shape changes in mouse and human embryos are quantitatively the same: both increase the external contact angles formed by cell-cell contacts from ~80 to ~150°. Naïvely, one could think (JL: as I definitely did), that identical shape changes in related species would result from the same mechanical changes. What surface tension measurements reveal is that mouse embryos compact by reducing their tensions at cell-cell contacts and doubling their tensions at their free interfaces whereas human embryos rely exclusively on a four-fold increase of tension at their free interfaces. Mechanically speaking, the mouse embryo adopts a more efficient way of compacting than human embryos do. We do not know how mechanical efficiency might be related to metabolism or any kind of evolutionary fitness but we think that mechanical measurements can provide new and original insights into our understanding of the evolution of morphogenesis. For example, here we see that the same morphogenetic movement in related species does not necessarily rely on the same mechanical strategy.
What are the implications of your findings on the understanding of human embryos compaction failure during IVF?
After IVF, only 40% of embryos undergo complete compaction. The remaining embryos may partially compact, containing cells that remain loosely attached to the compacted part of the embryo. These cells are suspected to be aneuploid and their exclusion was proposed to serve as some kind of protection or repair mechanism that would prevent unfit cells from being part of the embryonic tissue. However, we did not know how such mechanism would work. We now know that excluded cells are systematically less strong than their compacting neighbors, which outcompete the weak cells for position in the inner cell mass that will make all embryonic tissues (see https://doi.org/10.1038/nature18958 for more on this).
Also, our measurements reveal the very high tension at the surface of human embryos, which we think is also connected with cell fragmentation, a deleterious process that is common in human embryos. Indeed, in another study from the lab we found that increased tension in mouse embryos causes them to fragment (https://doi.org/10.15252/embj.2023114415).
Julie, were there any particular result or eureka moment that has stuck with you?
Julie: When Jean-Léon measured surface tension of mouse embryos, the question of measuring excluded cells did not arise because compaction in mouse embryos is always perfect and complete. Therefore, we had not initially planned to look into excluded cells from partially compacting human embryos. From the start, I measured all cells of all embryos, regardless of their quality. Jean-Léon admitted that he would not have measured all of these cells since he cared mostly about normal development but I could not resort myself to give up on any of these embryos! After accumulating and analyzing the data from all embryos, good or bad looking ones, we could see the distinct mechanical signatures linked to abnormal compaction in human embryos. That was nice!
And the flipside: any moments of frustration or despair?
Julie: There were some moments of frustrations with inhibitors or antibodies that would not work with human embryos in the same way as they did with mouse embryos. Since donated human embryos are very precious, we could not troubleshoot too much and it was always stressful to plan experiments and decide whether it was worth spending embryos to troubleshoot or if it was better to give up.
Thesis defense celebration with all PhDs who graduated in the lab so far. From left to right: Özge Özgüç, Julie Firmin and Markus Schliffka proudly wearing their PhD hats that were crafted by the lab and Jean-Léon Maître without anything on his head…
What’s next for you, Julie?
Julie: I am now what’s called a Hospital-University Assistant. This involves IVF work at Cochin Hospital in Paris, and teaching at the University of Paris Cité. Also, I continue to do research by collaborating with Jean-Léon and his team.
I am delighted to have these three roles, as they allow me to continue my research in developmental biology, to take care of patients suffering from infertility, and to contribute to the education of future professionals who will, in turn, take care of these patients.
And JL, where will this story take the lab?
Jean-Léon: I am very excited to look into more aspects of the mechanics of human embryos. For all our biophysical approaches in the lab, we always keep in mind that we may be interested to try them on human embryos. Therefore, a new measurement needs to be as little invasive as possible and to provide a metric with a physical unit so that we can compare human embryos to those of other species. We have a couple of new approaches in the pipeline that hopefully will be used on human embryos in the coming years.
Reference list
Firmin J, Ecker N, Rivet Danon D, et al. Mechanics of human embryo compaction. Nature. 2024;629(8012):646-651. doi:10.1038/s41586-024-07351-x
Maître JL, Turlier H, Illukkumbura R, et al. Asymmetric division of contractile domains couples cell positioning and fate specification. Nature. 2016;536(7616):344-348. doi:10.1038/nature18958
Pelzer D, de Plater L, Bradbury P, et al. Cell fragmentation in mouse preimplantation embryos induced by ectopic activation of the polar body extrusion pathway. EMBO J. 2023;42(17):e114415. doi:10.15252/embj.2023114415
At the speakers’ discretion, the webinar will be recorded for viewing on demand. To see the other webinars scheduled in our series, and to catch up on previous talks, please visit: thenode.biologists.com/devpres
On my first day at the ZMBP last October I saw our lab space and my first thought was ‘I have a lab!’. A little over half a year later, this May, our lab consisted of me, Steffi, Yadhu (PhD), VJ (PhD), Friederike (BSc), and Sarah (BSc). One afternoon in May we were all working in the lab: cloning, dissecting embryos, optimizing staining protocols. And then that thought repeated itself ‘I have a lab!’. And it wasn’t untrue in October it wasn’t true, but it was true differently in May. It felt different in May, bigger somehow but mostly just more meaningful. We are now doing experiments, developing projects, training new lab members, teaching students, writing a review, finishing BSc theses having lab lunches, and so much more. The lab is labbing and I’m loving it.
But how does a lab grow? Recruiting is an essential, but challenging part of building a lab, especially when starting out. I think lab culture is tremendously important; science itself provides you with enough challenges and you need those around you to support you in handling those setbacks and opportunities. I want to grow a group where we all love doing great science and asking critical, helpful questions, but also where we like being around each other and communicate well. So, what am I thinking about when recruiting new lab members? As I am now recruiting a Postdoc and we have new BSc students starting this fall I thought recruiting was the perfect theme to cover in this blog. With some advice for those recruiting and for those wanting to be recruited. Take this with a grain of salt, I am no expert and I am very new to this. But I do care and I am continuously working to improve.
For those starting to recruit
My main advice is to get advice. I talked with others in the department about their strategies, I next asked them for opinions on specific candidates, and I read books on lab management to think more about strategy and come up with questions. My goal is to be fair, consistent, and objective as much as possible but to not ignore my intuition.
With BSc students joining the lab I’ve been super lucky. I think those are difficult to judge as they are often very much still figuring out their interests and motivations. So far, my approach has been: if they seem motivated and communicate well it’ll probably be ok. And then when the lab is full, we are full. It’s worked out this far, but we’ll see, maybe in the future I need to figure out better criteria for judging who’ll be a good fit. Because while the BSc students only stay for 3-4 months, they do require time to teach and mentor, and like every other lab member they can have a big influence on lab culture.
For hiring the PhDs, for the position I posted I received 189 applications … So, I had to streamline things. To be blunt, most of these are spam. People want a PhD position, any PhD position, and just send their material without any personalization. And any PhD position might be fine for them, but I want someone who wants this specific PhD position. Therefore, my first goal was to see who was actually interested. I scanned through all CVs and motivation letters for indications that they like plants and developmental biology, that they read the paper mentioned in the job ad, etc. I kept track of my ‘scoring’ and ended up selecting an initial top 34. The others I advised to take a look at ‘Ten simple rules for writing a cover letter to accompany a job application for an academic position’. For the remaining 34 I read their applications in more detail to determine who would be a good fit: what was their motivation for picking plant development? What were their goals? What were their research experiences during their MSc? From these I selected 5 candidates to interview over Zoom.
Before the interview I asked candidates to prepare a presentation on their MSc thesis as well as a few slides on what they thought logical next steps from my Dev Cell paper were. I also sent them some of the questions I would ask them, so they knew what I was looking to learn from the interview. I requested to record their interviews. I asked them questions about their MSc projects and about how they thought about the plant developmental questions our lab is interested in. And I asked about their goals, what motivates them and what they value in a lab and mentor. After the interviews I was still deciding between 3 candidates, and I asked Steffi and others in the department to weigh in after watching the interviews. And then it became difficult because I ended up with 2 candidates I was really excited about, and I didn’t know how to choose because they were quite different and difficult to compare. I talked with Marja (Timmermans) and she suggested hiring both since I had an extra PhD position because of the Emmy Noether. I did and I have no regrets, I feel super lucky to have both Yadhu and VJ in the lab.
Now I am searching for a Postdoc and I’m going through the whole process again. For the project we need someone who’s interested in taking on a technical challenge and can really dig into optimizing single nucleus approaches for our system. On the other side, I am looking forward to having a more experienced voice in the lab, someone who is passionate about the big questions in science and will contribute to lively discussions. I want it all for the lab. There were 63 applicants this time, with 16 I ended up looking at in depth. This time I decided to invite 7 for a short conversation, focused mostly on their previous experiences, goals, and motivation, and some more information on the project from my side. Finally, now I am inviting some of them for a longer interview where they will present their science, talk with people in the lab, and talk with me about the project and plans for the future. In preparation for that I have been calling with some of their references, to learn more and to figure out what I should pay attention to in the next interview. Then hopefully it’ll be easier than last time to decide…
Hiring is scary. A lab is its people and assembling a team can be tricky. I’ve seen the immense effect one misfitting/negative person can have on a lab but also the great effect of having a positive and communicative group where people genuinely get along. It’s impossible to be sure how things turn out, all we can do is hire who we think will fit the project and team and then provide a supportive environment. Fingers crossed!
Some suggestions for those looking to be recruited
Avoid being spam by doing your homework. Quantity doesn’t substitute for quality. It’s better to do several applications well, compared to doing many sloppily. When preparing an application read the entire job ad, check the lab’s website and check a recent paper from them. If you are excited about their research: make that clear in your motivation letter. If you have relevant experiences: mention them. If the topic is very different: mention why you are interested in switching. And please try to get my name/title/gender/location right. I am not sir, or prof. Harter, and I don’t work at ‘Germany University’. And please don’t say you’re impressed by the excellent research from my lab, we haven’t published anything yet and that will just stress me out.
Please don’t say ‘I think I’d be a great fit’. At least not without explaining why. I’ve read so many applications where people say this but to me it is often not clear. You studied pharmacology, what courses/experiences make you a good fit for this position?
Imagine what you would want to know if you were in my position. I think this is always a great exercise for writing motivation letters and for preparing for interviews in any application. What do you think I want to know and how can you get that across. Also think about the big picture: avoid focusing only on the details in your presentation, show you’re able to identify the bigger questions, and share what you find important in a lab environment.
Ask questions. I’m sure that from your previous experiences you’ve identified some things that are important to you in a project, a mentor, and a lab. One goal is to get offered the position, but another should be to figure out whether you actually want the position. Asking questions helps you figure out whether the project and lab align with what you find interesting and important. And as a bonus it shows that you are interested and thoughtful in how you approach taking the next step in your scientific career.
Finally, both for hiring and applying: be open to feedback and be willing and excited to grow. I am sure that my approach will change as both I and the lab continue to develop. I might not always take all the advice I get but I think it’s important to consider and value it. In the end the goal is to keep learning and improving.
In June, the Node, preLights and FocalPlane organised SciCommConnect, an online event all about science writing and presentation. As part of the day, we hosted a three-minute research talk competition. We heard 12 fantastic talks on a wide range of topics. The talks were all so engaging that it’d be a shame not to share them more widely, so here is one of the talks from Rachel Mckeown, and a short interview about her research and her science communication experiences.
Rachel’s three-minute talk
What is your background?
I’m from a small village in Aberfan called South Wales, a beautiful and peaceful place. I’ve always had a deep love of science, biology in particular, which led me to study Natural Sciences at St Catharine’s College, Cambridge. In my final year I joined the Franze lab for my undergraduate research project, which is where the field of axon guidance was first introduced to me. I was fascinated by the idea that physical properties of the brain can guide neurons through the developing brain. I’d never really thought about the mechanics of the brain before, but with hindsight, it makes perfect sense that it’s an important factor. There’s so much we don’t know about how mechanics regulates development, disease and injury in the nervous system. This is why I wanted to stay in the Franze lab and extend my project into a PhD, which is where I am today.
Can you tell us a bit more about the research mentioned in your 3-minute talk?
Sure! It’s been known for decades that chemical cues are important regulators of axon guidance, but physical cues like tissue stiffness are new to the game. I’m particularly interested in how chemical cues and physical cues ‘talk’ to each other during nervous system development, and why this might be important to ensure that our brains are wired accurately and reproducibly.
In my experiments, I culture retinal ganglion cell axons (that send visual signals from the eye to the brain) from Xenopus frog embryos on gel substrates that ‘feel’ like soft and stiff regions of the Xenopus brain. We can measure brain stiffness in living embryos using atomic force microscopy and also see how it changes over time. This is how my lab first showed that a stiffness gradient forms in the brain as the retinal ganglion cell axons are navigating, and that they turn towards the softer tissue. I can expose neurons on these gels to different chemical guidance cues that they might encounter in the brain and see if their physical environment affects how they respond.
I’m also interested in exactly how these neurons are able to sense the stiffness of their surroundings. There are plenty of candidates for mechanosensing, and I’m working on introducing CRISPR-Cas9 technology to the frog model to test which are important for this process. This is a challenge because the Xenopus species I work with actually has two genomes, but there are tips and tricks to get around this.
How did you approach preparing for the 3-minute talk?
I knew summarising my research in just three minutes would be challenging, so I first sat down and wrote notes on what I thought was the most important information. I first had to set the scene and introduce the field of research. Then I had to show where I come in and what I’m doing to contribute to the picture. To round off, I wanted to get across the potential significance of this work and give the listener a message to take home. My draft was initially way too long, but I edited and edited again, cutting down words I realised weren’t needed and finding ways to shorten sentences. I timed myself while talking to myself a lot, until I was happy that I could speak calmly and clearly and still keep within the time.
For the presentation slide, I wanted to include only information that was directly relevant to the talk. I kept the structure simple and made sure that there was nothing that went unexplained or that I did not reference in my talk. I tried lots of different arrangements, adding things in and taking things out, until I was happy with the level of detail.
Finally, I had a run-through with a fellow PhD student in my lab, who gave me some great feedback. I changed a few things based on his comments, and I was ready to go!
What are your key takeaways from SciCommConnect?
For me, the main highlight was just how important it is to be able to explain the science that you do to anyone, whether that be at a specialist research conference, to other scientists outside the field, or to the general public. You need to adapt to each of these audiences, which is such a critical skill that I think is often overlooked. Sometimes, I think we are trained to think in such detail that, when communicating, the core messages can be lost in the technical details. SciCommConnect was about exploring ways to get our message across effectively, which a 3-minute presentation is great for. It challenges you to take a step back from an area of research you know so intimately and think about how to express that to someone who might have never heard about it before.
Another fantastic part of SciCommConnect was the sense of community across the entire world. There were participants from all over the globe, all brought together by a love of science communication. Everyone was so enthusiastic and engaged, with a real positive atmosphere throughout. I’ve found Zoom workshops in the past to be very draining, but not SciCommConnect, where we all took a very active role.
Have you been involved in other SciComm activities?
Absolutely. For a year I was President of Bluesci, the science communications society of the University of Cambridge. This was an incredible opportunity to gain insight into all areas of SciComm; working alongside a dedicated and motivated team, I was involved in commissioning, designing and producing magazine articles, writing several features myself.
On top of Bluesci, I’ve also written several posts for preLights, one branch of The Company of Biologists involved in organising SciCommConnect (along with The Node!). Here I get to write for a slightly more technical audience as most readers will be fellow researchers, and I get to ask questions directly to the authors of the preprints that I write about.
Finally, I’ve written a few blog posts for Women in Neuroscience UK which are more casual in style – less about the science itself and more about my experiences as a PhD student and researcher. Again, this is a really different style of writing, much more relaxed but hopefully just as informative and interesting to read!
What are you currently working on? And what are your future plans?
I’m currently taking some time away from the lab in order to focus my attention on my PhD thesis, where I’m putting some of the skills from SciCommConnect to use. Writing up four years of work in the lab is going to take some time, but reviewing everything I’ve done over the last few years has given me a sense of pride. Even when things were not going right, resourcefulness and resilience helped me push on to the next experiment. It certainly hasn’t been a smooth journey – that’s how science goes – but now is the time to turn it into a complete story.
As for my future plans, I’m glad I have a little longer to figure that out! I’d love to find a career that brings together all of the things that I’ve found most enjoyable about my PhD years – communication, collaboration and creativity, as well as fascinating science.
 (2 votes)
(2 votes)
 (No Ratings Yet)
(No Ratings Yet)