Where are you originally from and what do you work on now?
Eva and a surfboard she unleashed her paints on. “During the first lockdown in 2020, I was running out of paper and my housemate let me paint his favourite board.”
I’m from Malaysia and am currently working with Kristian Franze at the University of Cambridge. Here I study the interplay of mechanical and chemical signalling in connecting the nervous system during development. More specifically, I study how axons that originate in the eye respond to chemical and mechanical cues in their environment that help guide their growth towards the visual part of the brain.
Has science always been an important part of your life?
When growing up, I didn’t really know that being a scientist was an option! In fact, I did an Engineering degree before Biology lured me in with its wondrous questions on how life forms and functions. I have always been curious and wanted to know how and why things are the way they are; most children are scientists at heart, guess I never really grew out of that?
And what about art ?
As a kid I loved mixing colours to make ‘new’ ones and doodling (to date, most of my textbooks, papers, and notebooks have doodle-filled margins :p). I don’t have any formal training or education in art. However, rather randomly in the final year of high school I decided to take an art GCSE, it ended up being a fun independent study experience. Since grad school I’ve had a more consistent relationship with art and try to learn a new technique every year.
What or who are your artistic inspirations?
Yayoi Kusama – one could simply disappear in her art! Bill Harris (our previous head of department) – his art hangs in our corridors and sometimes when the going gets tough, I wander out to stare at them and remember how beautiful neurons and the visual system are! And Abhishek Singh – his fabulous depictions of Vedic texts are incredible!
How do you make your art?
It depends on the technique I am playing with. I love mixing my media and enjoy a variety of art forms.
Painting is my spontaneous medium, I allow my hands and eyes to work without thinking or planning. This is the go-to medium in weeks of super structured/planned lab work, it brings me some flow.
Lino-printing is another favourite technique. Printing however relies on some planning and drafting/sketching before I jump into carving the lino. I seem to print more when my work week is less structured (2020 resulted in quite a lot of prints :p).
Does your art influence your science at all, or are they separate worlds?
At present, they are separate worlds that complement each other. While I have illustrated pieces for others science, I feel too close to my current research project to make an art piece of it. Having said that, I rely on my “art eyes” to communicate science, particularly in breaking down complex ideas through simple visual representations. Art practise also trains one to be more observant and detailed, traits that are very handy in the lab!
I rely on my “art eyes” to communicate science
What are you thinking of working on next?
Science-wise, I’m excited to be back in the lab catching up on experiments that couldn’t be done in 2020. I’m also figuring out what I want to work on next. Art-wise, I plan to shift from a 2D to 3D system by trying out clay this year!
Octopus dreams: This illustration was a commission for BlueSci Cambridge (https://www.bluesci.co.uk/), for a piece based on the PBS Nature video of an octopus rapidly changing colour while sleeping. Was it moving through different environments in its sleep? Was it feeling happy? Hungry? Threatened? Living on land?
The Maze: Part of a series of plein air paintings made in the Inner Hebrides. Painting outdoors in the bracing Scottish weather was an interesting challenge.
St John’s College, Cambridge: This was a painting I made of one of my favourite parts of college. St John’s is gorgeous, and I particularly love the flaming autumnal display of Boston Ivy on the back of New Court.
Eva’s paintings
Xenopus laevis: My model organism!
Dionaea muscipula: This piece was inspired by a paper linking signal memory to calcium dynamics in a transgenic Venus flytrap! (paper link: https://www.nature.com/articles/s41477-020-00773-1)
Phylliidae: masters of camouflage. This was part of a month-long ink art series.
Eva’s inks
Little Barn: In 2020 I defended my PhD at home, on Zoom, with a view of the village church from my window. I commemorated this event by making a print of the houses and the church spire peeking out behind.
Solidarity: no one should be afraid of who they love or who they are.
An otter print: carving for a friend who loves otters.
Eva’s lino prints
In memory of John Horton Conway (1937-2020). The portrait is made up of frequently occurring patterns found in The Game of Life
We’re looking for new people to feature in this series throughout the year – whatever kind of art you do, from sculpture to embroidery to music to drawing, if you want to share it with the community just email thenode@biologists.com (nominations are also welcome!).
Stowers Research Conferences are biennial meetings that bring together researchers, faculty, and graduate students to inspire creative thought and collaboration in the basic sciences.
In the latest episode of Genetics Unzipped we’re bringing you some scientific ‘tails’ -literally – as we explore the genetics of dog breeds and behaviour. Is there a gene for being a Very Good Dog or having a boopable snoot? And what happened over tens of thousands of years to turn a fearsome wolf into a pug in a party hat?
We speak with Elinor Karlsson at the University of Massachusetts Medical School and the Broad Institute who’s looking into all aspects of dog genetics, from discovering what gives different breeds their distinctive traits to finding out how genetic variations contribute to psychological characteristics.
Jessica Hekman works with Elinor Karlsson and her team studying the brains and genes of dogs to understand canine behaviour, including trying to figure out why certain dog breeds behave in certain ways.
And Jeff Schoenebeck from the Roslin Institute at the University of Edinburgh has made it his mission to understand how genetic variations contribute to the wide range of shapes and sizes of dog skulls, using CAT scans, as well as lab tests.
If you enjoy the show, please do rate and review on Apple podcasts and help to spread the word on social media. And you can always send feedback and suggestions for future episodes and guests to podcast@geneticsunzipped.com Follow us on Twitter – @geneticsunzip
Ciliopathies are genetic disorders caused by defects in the structure and function of cilia, microtubule-based organelles present on the surface of almost every cell in the human body which play crucial roles in cell signalling. Ciliopathies present a wide range of often severe clinical symptoms, frequently affecting the head and face and leading to conditions such as cleft palate and micrognathia (an underdeveloped lower jaw that can impair feeding and breathing). While we understand many of the genetic causes of human ciliopathies, they are only half the story: the question remains as to why, at a cellular level, defective cilia cause developmental craniofacial abnormalities. Researchers have now discovered that ciliopathic micrognathia in an animal model results from abnormal skeletal differentiation and remodelling. The work from Christian Bonatto Paese, Evan Brooks and others from Samantha Brugmann’s lab at the Cincinnati Children’s Hospital Medical Center in the USA is published in the journal Development.
Surface-rendered whole-mount chick mandibular prominence in wild type chick embryos, stained in blue for DAPI and in red for Sox9, an osteochondroprogenitor marker.
The researchers used the avian ta2 mutant as a model for Oral-Facial-Digital syndrome subtype 14 (OFD14), a rare human ciliopathy characterised by micrognathia. They observed defective formation of the jaw bone (mandible) at early stages of development in ta2 mutants. These defects correlated with unchecked progression through the cell cycle and over-proliferation in skeletal progenitor cells. Importantly, these progenitor cells failed to differentiate into mature osteoblasts (the cells that secrete bone), and this failure of differentiation subsequently led to a reduction in bone deposition and hence micrognathia. The researchers also identified excessive bone resorption, a process which normally contributes to final size and shape of the mandible, as an additional causative factor in the ta2micrognathia. This work informs our understanding of the etiology of human ciliopathic micrognathia.
“We have identified distinct cellular processes that are impaired during the onset of ciliopathic micrognathia,” says Samantha Brugmann. “We know from previous work that these processes are responsive to treatment with pharmacological agents, and we are currently testing a number of these agents to determine if ‘rescuing’ ciliopathic micrognathia is possible. The therapeutic implications are exceptionally real.”
“We are also currently trying to uncover how various organ systems respond to loss of cilia and which molecular and signalling pathways are affected, to continue to advance our understanding of how to tackle treatment for patients,” says Christian Bonatto Paese.
The paper exemplifies how developmental biology can shine a light on devastating genetic disorders. Brugmann concludes: “The most important impact of this study, to me, is how useful basic science and the avian embryo can be towards uncovering mechanisms for human disease.”
______________________________________
Christian Louis Bonatto Paese, Evan C. Brooks, Megan Aarnio-Peterson, Samantha A. Brugmann. (2020) Ciliopathic micrognathia is caused by aberrant skeletal differentiation and remodeling. Development 2021 148: dev194175 doi: 10.1242/dev.194175 Published 15 February 2021
The 2020 call for the Catalan “Beatriu de Pinós Postdoctoral Fellowships” is now open. This is a fantastic opportunity to join the Araújo group, who are focused on identifying the molecular mechanisms underlying single-cell branching during development and disease. We study this during morphogenesis of the Drosophila melanogaster tracheal and nervous systems (Ricolo et al. Current Biology 2016 and Ricolo and Araújo, eLife, 2020), and are also interested in knowing how single-cell branching affects the whole organismal behaviour. We are combining cell analysis and confocal tissue live imaging, with genetic approaches and CRISPR/Cas9 technologies.
We are based at the Institute of Biomedicine of the University of Barcelona (IBUB) and are looking for a motivated and enthusiastic candidate who will play a central role in the lab. You must have a Ph.D. in areas relevant to cell/developmental biology and/or in computational biology (and have defended your Ph.D. thesis before the 31st of December 2018) and have 2 years postdoctoral experience outside of Spain. Applicants are expected to have excellent interpersonal and communication skills, be highly independent and committed to research in a fast-moving and exciting field.
If you’re interested, please write to sofiajaraujo [at] ub.edu
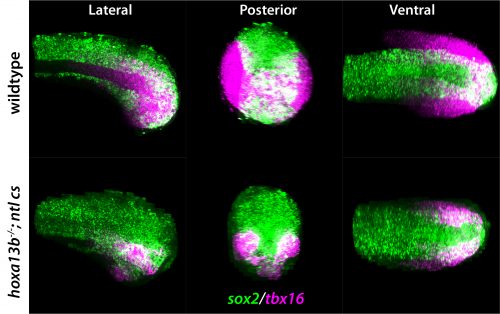
The anterior to posterior extension of the vertebrate body axis relies on a population of bipotent neuromesodermal progenitors in the tailbud. A new paper in Development uncovers a crucial and unexpected new role for Hox13 genes in sustaining these progenitors to promote axis extension in zebrafish. To hear more about the story, we caught up with the paper’s two authors: postdoctoral researcher Zhi Ye and his supervisor David Kimelman, Professor of Biochemistry and Adjunct Professor of Biology at the University of Washington, Seattle.
Zhi (L) and David (R)
David, can you give us your scientific biography and the main questions your lab is trying to answer?
DK: As a graduate student at Harvard, I worked on adenovirus, but I became fascinated by developmental biology and had the great fortune to work as a postdoc with Marc Kirschner on early Xenopus development at UC San Francisco, where we did some of the founding studies on mesoderm-inducing factors. When I started my own lab in Seattle, I continued to study Xenopus, and eventually became intrigued with the possibilities in zebrafish, which is the current focus of my lab.
The main problem we are interested in has fascinated developmental biologists for almost a century, which is how the posterior body forms from the mass of cells called the tailbud. This problem has been reinvigorated recently with a finding that we and others made: that the mesoderm and neural tissue of the posterior body arise from a very interesting neuromesodermal progenitor population. There is now a terrific collection of scientists working in this area and I am very happy to be part of this international effort.
And Zhi, how did you come to join David’s lab, and what drives your research today?
ZY: Three years ago, after I obtained my PhD from a genetics research group at Auburn University that studies farmed fish, I decided to obtain training in a developmental model system to better equip myself for an academic career. I am specifically interested in the field of developmental biology as it aims to uncover the mysterious, yet gorgeous, mechanisms that allow a single cell to develop into such a diverse variety of different organisms. In addition, I wanted to learn cutting-edge technologies that will be useful in advancing my scientific career.
When David responded to my application with a long email giving me some details about his lab, I could feel his enthusiasm and passion about science through these words. We had an interview where I was introduced to the Hox project, which really impressed me, and fortunately he agreed to train me. I feel so lucky that I made the right decision to join David’s lab and had the chance to work with the other friendly and outstanding developmental biologists at the UW. The three years of training in David’s lab has been such a great experience, not only because I have learned a lot about how to do research, but also, which I think is even more important, how to be a responsible scientist and mentor.
How has your research been affected by the COVID-19 pandemic?
ZY: Like many other people, I was not able to do bench work during the 6 weeks of full lockdown at the UW, and things were also slowed down as some reagents we needed from Germany for a key experiment were delayed for months. It turned out to be a good time for me to do some intensive reading and to learn computational skills for the bioinformatics analysis of the sequencing data that is the basis for my next paper.
DK: I spend a lot of time driving back and forth between my house, where I now have my office set up, and the lab, where my experiments are. Research feels very disjointed because of this.
What led you to initially work with zebrafish Hox13 genes?
DK: I have spent a lot of time trying to think about how the tailbud cells continually and progressively release cells into the mesoderm as the body axis extends. I was particularly intrigued by the overexpression studies in amniotes that proposed that the Hox13 genes act to terminate this process. My original plan was to see if we could just extend the amniote studies by doing overexpression in zebrafish, and then isolating the tailbud cells and performing RNA-seq to analyse the genes controlled by the Hox13 proteins, as the previous studies had only studied a few candidate downstream genes.
Can you give us the key results of the paper in a paragraph?
ZY & DK: Whereas the previous studies had argued that the Hox13 genes act to terminate body axis extension in vertebrates, our results show that they act together with the transcription factor Brachyury (now called Tbxta in many systems) to promote the formation of mesoderm from the neuromesodermal progenitors, thereby allowing the posterior body to form. This is a very different way of looking at the role of the Hox13 genes, which we believe will be applicable to all vertebrate embryos.
Three views of wild-type and hoxa13b mutant fish.
I understand the path from making the Hox13 mutants to finding a phenotype relied on some fortunate coincidences?
DK: As stated above, the original idea was to overexpress the Hox13 genes and then analyse gene expression in the tailbud with RNA-seq, as no one had comprehensively studied the downstream genes. We spent a lot of time doing this, but honestly it was a mess because, although lots of genes went up and lots of genes went down, we couldn’t make any sense of what we were seeing, and I felt terrible that I had dragged Zhi into this hellhole of a project. Fortunately, CRISPR had become relatively easy in zebrafish, and I was making a lot of mutants in genes of interest, and so I made a mutant in one of the two most abundant Hox13 genes: hoxa13b. However, to my disappointment it had no effect. I then targeted the other abundant Hox13 gene, hoxd13a, to make a hoxa13b;hoxd13a double mutant, and then I did see a small percentage of embryos with some phenotype, but it was really a major bummer after all that work and many months of raising and screening fish that the effect was so marginal.
Very fortunately I asked Zhi to give me his thoughts on the double mutant embryo phenotype, and he asked me to put the embryos at 21°C after fertilization rather than the normal 29°C, as this would allow him to watch the body form during regular work hours (at 29°C much of the body extension occurs during the middle of the night with zebrafish). I did as he requested and I was really surprised when he told me that he was seeing dramatic effects on axis formation in many of the embryos! At first I thought he had to be mistaken, but he was indeed correct. What we now know is that we were just very lucky that I had made the Hox13 mutations in the background of a homozygous cold-sensitive mutation in tbxta, which is a naturally occurring mutation that is present among the lab’s ‘wild-type’ fish. This tbxta mutant normally has no effect, such that fish with just the tbxta cold-sensitive mutation are completely normal when grown at 21°C, but when the two Hox13 genes are mutated in this background there is a very strong synergistic effect. Had I not by chance made Hox13 CRISPR mutants in a homozygous tbxta cold-sensitive background, and had Zhi not asked me to raise the embryos at 21°C, we would not have uncovered this whole story. Serendipity has often been a major factor in my scientific career, and this work completely exemplifies this point.
Serendipity has often been a major factor in my scientific career, and this work completely exemplifies this point
When doing the research, did you have any particular result or eureka moment that has stuck with you?
ZY: When I saw the larger neural tube and smaller presomitic mesoderm in the hoxa13;hoxd13 double CRISPR mutants, I was really surprised and excited! I had been working on studying RNA-seq data from the Hox13 overexpression lines for more than a year but I couldn’t make any sense of how the transcriptional changes caused by overexpressing Hox13 produced such a profound truncation of the embryos. The fate change phenotype shown in the double mutants provided us with a whole new way of thinking about the role of the Hox13 genes in early embryos.
And what about the flipside: any moments of frustration or despair?
ZY: This was a very tough project for me as we went through so many ups and downs. For example, when I told David my exciting findings with the CRISPR mutants, David warned me that as I had kept the embryos at a cooler temperature to slow them down for analysis, it might be an artefact due to the fact that some of our ‘wild-type’ fish have the cold-sensitive tbxta mutation, which would have been a huge disappointment. But it was a very lucky break as it is only when Tbxta function is reduced that the role of eliminating hox13 genes is revealed. The interaction of the Hox13 proteins and Tbxta helped us develop a new understanding of the role of the Hox13 proteins. The beauty of science, and also life, is that all these upsetting moments can also lead to great happiness. I am so grateful to David for his continuous support and encouragement during the hard times in this project.
What next for you after this paper?
ZY: We are working on finishing up a second exciting story about the Hox13 genes in which we identified the direct targets of Hox13 in vivo, and these results strongly back up the findings in our first paper. We plan to submit this work soon. After leaving David’s lab, I am going back to China and I will spend time studying interesting targets of Hox13 that we were not able to cover in our second paper. I am currently seeking a faculty position that will allow me to continue this work; combining my postdoctoral research with my graduate studies, I plan eventually to use a developmental biology perspective to improve aquaculture production.
Where will this story take the Kimelman lab?
DK: Zhi has adapted a very cool new method called CUT&RUN that has allowed him to identify genes that the Hox13 factors bind in vivo. One of the major problems with the studies of all Hox proteins, particularly in vertebrates, has been determining real in vivo targets; Zhi’s work has identified many interesting target genes. While Zhi will study some in his own future lab, there are plenty of targets for both of us to analyse.
Finally, let’s move outside the lab – what do you like to do in your spare time in Seattle?
ZY: Hiking is absolutely my first choice. Seattle is such a wonderful place to live with a variety of natural landscapes and hiking trails available within a 2-hour driving distance. Cooking is another way of relaxing; it’s like doing experiments but you can get your results (dishes) and publication (photos on social media) much quicker!
DK: For the past 10 years I have been working on Sundays with a terrific organization called Rebuilding Together Seattle, for which I go to houses throughout the Seattle metropolitan area and do free home repairs for people on a low income. It is a bit like science in that I am constantly trying to solve problems, but also very gratifying in that I can directly improve people’s lives.
In our fifth SciArt Profile we meet Sofia Araújo, a Professor in the University of Barcelona.
Where are you originally from, where do you work now, and what do you work on now?
I am originally from Portugal. I took my Ph.D. in the UK and now, after a postdoc and a career development award at the IBMB-CSIC and IRB Barcelona, I am a Professor at the Department of Genetics Microbiology and Statistics and the Institute of Biomedicine at the University of Barcelona (http://www.ub.edu/ibub/research-group/sofia-araujo/), where I run a research lab on the genetics of cell behaviour during development. In my research group, we work on the cell biology of development, more specifically on how single-cells branch.
Where you always going to be a scientist?
Science has always been a very important part of my life. I knew from early on that I wanted to be a researcher and the real problem has always been on what to focus on, since I get excited about many things and like to follow the lead from whatever interesting results we have in the lab.
And what about art – have you always enjoyed it?
Art, like science, has always been an important part of my life, since I was a child. I remember spending long hours drawing and painting and finding it a great way to express myself. I got extra help from my mum, a secondary school arts teacher, which made things easier whenever I wanted to try a new technique! Unfortunately, it is art that mainly suffers from when I don’t have enough time, so it always has had its ups and downs. However, I do need it as a balance in my life, so I always end up finding the time to do it! I am mainly a drawer and a painter, but I have also done some pottery and woodwork and, of course, the good old embroidery.
Who are your artistic influences?
I like art and I enjoy the works of many artists, but I cannot say I have an artistic influence. Because I like drawing and painting nature, animals and scientific images, I do like the influence of artists like Henry Rousseau, Vincent Van Gogh, Marianne North and of course, Ramon y Cajal!
How do you make your art?
Nowadays, I only use charcoal, watercolour and oil. And lately, I have been mainly working with oil on canvas. Approaches vary depending on which painting I work on. If I am painting a dinosaur for my kids, I search for good images on the internet. Sometimes, I paint landscapes from my own photos. Or I draw and paint cells and tissues from our own confocal images.
Does your art influence your science at all, or are they separate worlds?
They are both part of my life, so they do influence each other, of course. But I cannot pinpoint if painting has ever made me compose my microscopy photographs in any special way. I think it is more like my science has influenced my art, since I started painting confocal microscopy images with oil on canvases!
What are you thinking of working on next?
I will certainly continue painting confocal images and animals, intercalated with other types of images, like landscapes, whatever inspires me as I go along. I only wish I had more time to experiment with new techniques and ways of painting!
Neurons – Oil on canvas
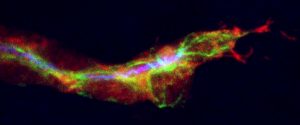
Tracheal Terminal Cell – Oil on canvas
Leziria (Portuguese landscape) – Oil on canvas
Torre de Belem – charcoal on paper
T-Rex – Oil on canvas
Gallery of Sofia’s pieces (click for full size image & caption)
We’re looking for new people to feature in this series throughout the year – whatever kind of art you do, from sculpture to embroidery to music to drawing, if you want to share it with the community just email thenode@biologists.com (nominations are also welcome!).
In the context of an ERC funded project, the laboratory of Dr. Bomont (NeuroMyoGene institute, Lyon, France) is recruiting several fellows.
We are seeking highly motivated candidates for neuronal cytoskeleton, to work on a project aiming at fueling innovation in fundamental neurobiology and therapeutic development.
This project is multidisciplinary and will be conducted in an enthusiastic lab fascinated about the biology of the cytoskeleton, together with collaborators expert in mathematics and biophysics. If you are passionate about pushing the frontiers of developmental neurobiology using live microscopy & OMICS in zebrafish, this project is for you!
The group of Prof. Dr. T. Vogel at the Institute for Anatomy and Cell Biology, Department Molecular Embryology, University of Freiburg is looking for a highly motivated Postdoc. One research focus of Prof. Vogel´s group lies on epigenetic mechanisms. In particular, we study histone modifications in stem cell specification during neurodevelopment and in CNS disease (Ferrari et al, Nat Comm, 2020, Gray de Cristoforis et al, Mol Brain, 2020, Franz et al, NAR, 2019, Bovio et al, Mol Neuro, 2018, Grassi et al, Cerebral Cortex, 2017, Roidl et al, Stem Cells, 2016). We are looking for a postdoctoral researcher to integrate into the current research projects of the lab. These focus lies on the role of transcriptional regulation through histone modifications with regard to interneuron specification and/or on hippocampus development. As a surplus the group gives possibility to teach macro- and microscopic anatomy in German language. Starting date: from now on.
You are convincing through
a strong background in biochemistry, biology, molecular medicine, medicine or equivalent, as well as an outstanding PhD or MD,
documented experience in techniques of cell biology, protein biochemistry and molecular biology, applied to understand the development or function of the central nervous system,
experience with bioinformatical analyses of large data sets and next-generation-sequencing, in live cell microscopy, or electroporation techniques,
very good communication skills in English,
enthusiasm for science, strong motivation, the ability to learn new techniques and to work synergistically in a team,
motivation to teach medical students in Anatomy.
We are offering
to work on high topical research projects with different techniques in a collaborative setting,
an international work atmosphere,
a scheme towards academic qualification and education in teaching and research.
Employment will be temporary.
Please send applications including the usual documentation (CV, degrees, a minimum of 2 references, short letter of motivation for application to the lab, list of publication) via email until March 15th to:
We offer one fully funded postdoctoral position up to five years in the Laboratory of Genome Integrity located at the National Institutes of Health (NIH, Bethesda, MD). NIH is the largest biomedical research agency in the world, fosters world-renowned researchers and provides access to state-of-the art innovative technologies and scientific resources.
Our laboratory uses human and mouse embryonic stem cells (ESCs) as well as mouse embryos to understand the molecular mechanisms underlying cell fate decisions. The applicant should have or about to have a PhD in Developmental Biology, Genetics or similar, and must have demonstrated expertise on molecular biology/mammalian cell culture (preferably in embryonic stem cells). Knowledge in mouse embryology, single-cell RNAseq, chromatin architecture and/or next generation sequencing technologies will be considered as an advantage.
The applicant will be involved in a very exciting project investigating the relation between cell plasticity/totipotency and chromatin architecture (see our last publication about this topic, https://doi.org/10.1101/2020.12.20.423692). We seek a highly motivated, creative individual, eager to learn and develop new technologies and complex cell systems based on live cell/embryo imaging, single-cell technologies and CRISPR-based editing interested in understanding how a single cell can develop into a complex multicellular organism in vitro and in vivo.
Please send a brief cover letter, CV and three reference letters via e-mail to:
Vega-Sendino, et al (2021) The ETS Transcription Factor ERF controls the exit from the naïve pluripotent state. BioRxiv, doi: https://doi.org/10.1101/2021.02.01.429223.











 (3 votes)
(3 votes) (No Ratings Yet)
(No Ratings Yet)

 (1 votes)
(1 votes)