Bilaterality, the property of having two symmetrical sides, is widely conserved among animals. It is estimated that 99% of all animal species are bilaterians, with the remaining 1% composed by sponges and radial animals, which lack or have radial symmetry respectively (1). Although bilaterality is widespread among animals, little is known about how it is developmental patterned or shaped.
Observations from naturally occurring gynandromorphs such as birds, lobsters and butterflies (to name a few) suggest that cells from left and right sides remain on their own (ipsilateral) side, with little mixing seen through out life (2). In these animals the right and left sides, which have different phenotypic colors, meet at the body midline without crossing to the contralateral side. In fact, a similar phenomenon has been reported in humans: in a clinical case-report, a patient was found to have an ovary on the left side and a testicle on the right side, and further karyotyping of skin fibroblasts revealed XX sex chromosomes on left side and XY on the right (3). This information hints at the presence of mechanisms ensuring the patterning of ipsilaterality during development.
In the case of amniotes, this is particularly intriguing given that in early body patterning (gastrulation), cells undergo epithelial to mesenchymal transition (EMT) and these cells are known to be highly invasive (4). Therefore in the recent publication Maya-Ramos and Mikawa (5), we addressed the question of how is ipsilaterality patterned during amniote gastrulation. This question was best addressed using the chick embryo, given its long-standing history as a model system in gastrulation, its handling, easy accessibility, high temporal and spatial transfection control parameters and live imaging robustness.
Our results demonstrated that ipsilaterality is patterned during gastrulation; that is, right epiblast cells undergoing EMT gave rise to right mesoendodermal cells while left epiblast cells resulted in left mesoendodermal cells. Epiblast cells undergoing EMT seldom crossed the embryonic midline. These findings are consistent with the observations of bilateral gynandromorphs and human clinical case reports, and argue that left and right sides in bilaterians are established early in development.
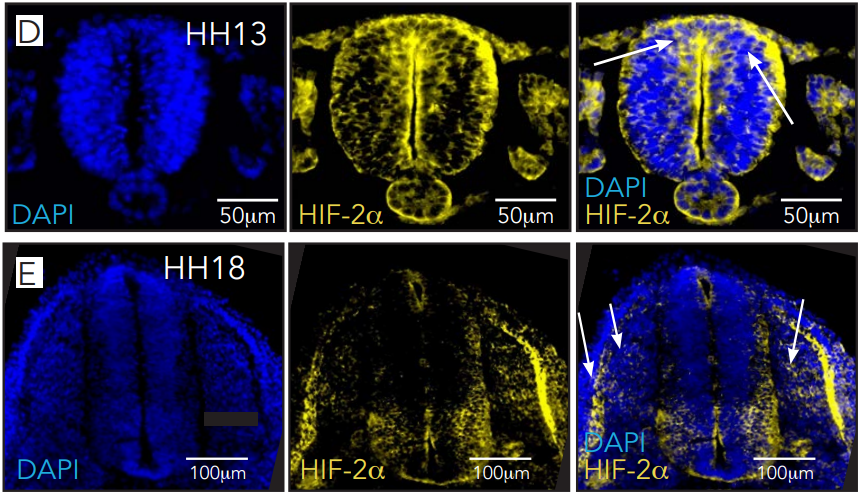
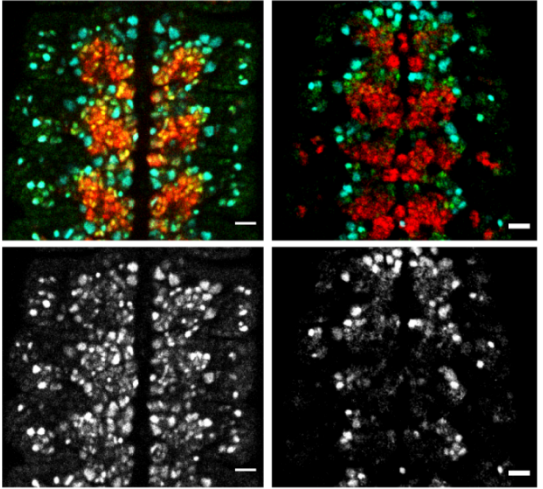
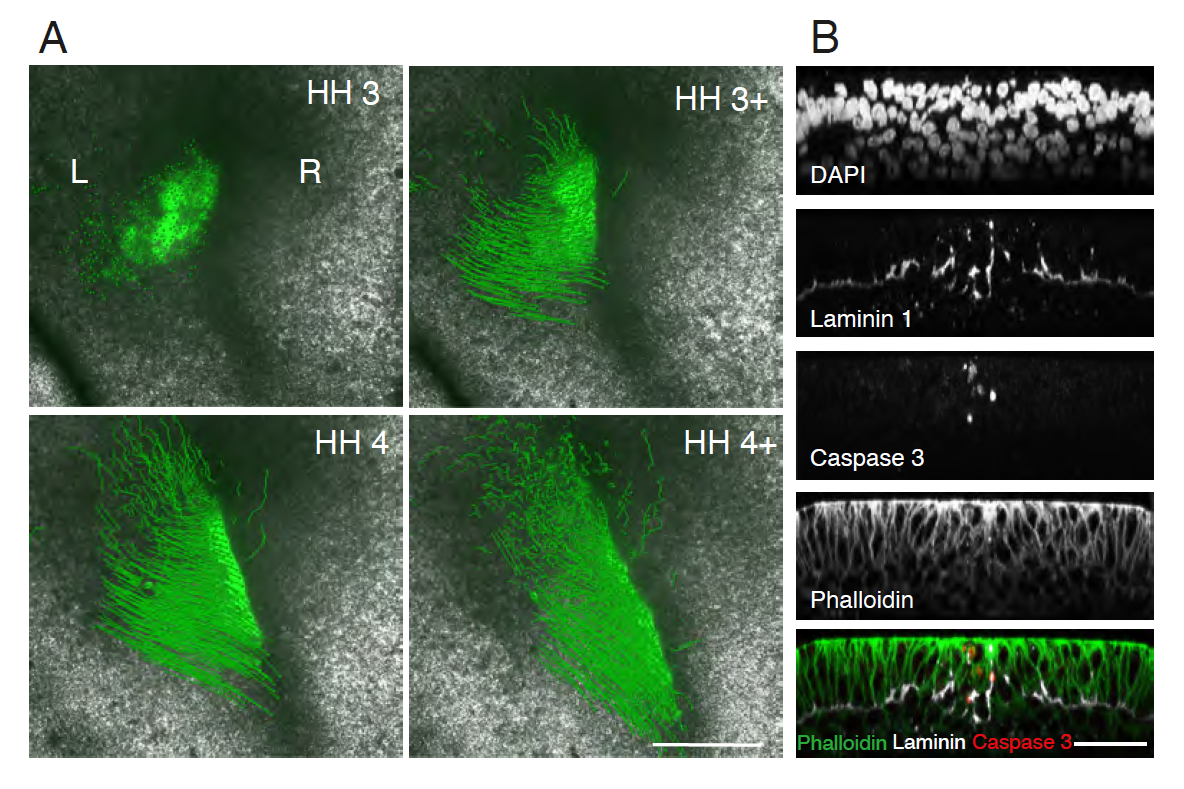
Gastrulation is ipsilateral and the PS midline is defined by ECM and PCD. (A) Left sided electroporation with Flag:2A:H2B-GFP. The majority of cells remain on the ipsilateral side without crossing the PS midline. Scale 500 μm. (B) PS cross-section immunofluorescent staining for laminin and cleaved caspase-3, both enriched at the PS midline. Scale 50 μm.
We found that the mechanism preventing cellular mixing was at the primitive streak (PS) midline. The PS midline was cellularly and molecularly distinct from PS lateral cells, as it was enriched with both extracellular matrix (ECM) proteins and programmed cell death (PCD). The origin of PS midline cells undergoing PCD was traced to a unique posterior embryonic region, embedded within the early PS. ECM and PCD loss of function resulted in crossing of the embryonic midline to the contralateral side. However, ipsilateral gastrulation was only restored with exogenous PCD.
These results highlight two key points. One is that PCD serves as a signal that prevents cell migration – this gives PCD a positive functional role in development. It is still unclear, however, what is the mechanism by which PCD prevents contralateral migration, for instance whether steps leading to PCD or the persisting cellular debris is responsible for this phenotype. Lingering cellular debris leading to intracellular content release has being associated with pathological processes, including Alzheimer’s disease, Parkinson’s disease and Systemic Lupus Erythematosus (6-8). Therefore, is not inconceivable that these same signals may take on a physiological role in development.
Second, these results suggest that ipsilaterality is programmed within bilaterality and that upstream signals are in place to specify PS midline cells before they undergo PCD. Therefore a persisting question is, how is the midline defined?
The Wellcome – MRC Cambridge Stem Cell Institute is an international centre of excellence for stem cell research and regenerative medicine. Scientists in the Institute collaborate to advance our knowledge of various stem cell types and to perform pioneering work in translational research areas, providing the foundation for new medical treatments (https://www.stemcells.cam.ac.uk/).
The Living Systems Institute (LSI) pioneers transformative science to engineer control of complex biological systems. LSI merges research in biology and medicine with ground breaking physical sciences technologies and powerful mathematical modelling capabilities. https://www.exeter.ac.uk/livingsystems
A post is open for a Research Assistant/Associate bioinformatician in the laboratory of Professor Austin Smith to work on an ERC Advanced Grant project on Plasticity of the Pluripotency Network (PLASTINET). You will contribute to analyses of the fundamental biology of pluripotency and pluripotent stem cells in different mammals. The laboratory is currently based in the Wellcome-MRC Cambridge Stem Cell Institute https://www.stemcells.cam.ac.uk/People/pi/smith and will relocate to the Living Systems Institute, University of Exeter, in July 2020 https://www.exeter.ac.uk/livingsystems/.
Research assistant candidates should have an MSc or equivalent in Bioinformatics, Computational Biology, Systems Biology or related subject. Research Associate candidates should have a PhD or equivalent research doctorate in computational biology or bioinformatics.
You will develop and implement algorithms, analysis methods and visualisation tools for dissecting multi-omics datasets. In particular, you will use scRNAseq, ChIPseq and ATAC-seq data in order to elucidate the regulatory networks governing pluripotency and undertake comparisons across species. You will also be encouraged to develop new analyses within the group’s field of interest.
Candidates will have proven capacity to understand and execute high-throughput data analysis, and should be familiar with a UNIX/Linux environment and programming/scripting language (Python, R, Matlab). General knowledge of molecular cell biology and experience in sequencing analysis will be beneficial. Specific training and support will be provided as needed.
Good communication skills and the ability to work effectively in a team are essential. This post will transfer to the University of Exeter in July 2020.
Research Assistant salary range £26,715-£30,942; Research Associate salary range £32,816-£40,322, depending on experience and qualification.
Fixed-term: The funds for this post are available for 3 years in the first instance.
To apply for this post please follow this link: http://www.jobs.cam.ac.uk/job/24910/. Click the ‘Apply’ button on Job Opportunitues to register an account with the Cambridge University recruitment system (if you have not already) and apply online.
The closing date is 20 February 2020, with interviews to be confirmed.
Please ensure that you upload a covering letter and CV in the Upload section of the online application. If you upload any additional documents which have not been requested, we will not be able to consider these as part of your application.
Please include details of your referees, one of whom must be your most recent line manager, with email address and phone number.
Please quote reference PS22234 on your application and in any correspondence about this vacancy.
The University actively supports equality, diversity and inclusion and encourages applications from all sections of society.
The University has a responsibility to ensure that all employees are eligible to live and work in the UK.
We are looking for highly motivated individuals who share our passion for science and would like to work in a friendly and collaborative environment.
Fully funded PhD student and Postdoc positions are available in the laboratory of Dr. Peng Huang in the Department of Biochemistry and Molecular Biologyat the University of Calgary, Canada.We use zebrafish as a model system to understand how tissue patterning is achieved and how tissue integrity is maintained. We study the spinal cord patterning to understand how different cell signaling pathways (Hedgehog and Notch signaling) interact during cell fate specification. We also study how non-muscle cells (e.g., tendon fibroblasts and muscle progenitor cells) contribute to muscle development, degeneration and regeneration. For more information about the lab and our recent publications, please visit: https://people.ucalgary.ca/~huangp/index.html
PhD student candidates should have a BS or MSc in Molecular Biology, Genetics, Developmental Biology or a related discipline, a strong academic background, good English skills and an enthusiasm for research. Previous lab experience with genetic model organisms is preferred but not required. Excellent written and verbal communication skills are critical.
Postdoc candidates should have a PhD in Developmental Genetics or a related discipline, excellent molecular biology skills, and a strong interest in developmental biology. Excellent written and verbal communication skills are critical. The candidate must have a track record of academic success as evidenced by peer-reviewed publications, awards and scholarships.
To apply, please send a cover letter summarizing previous research experiences and future goals and the CV with names of 2-3 references to Peng Huang, peng.huang@gmail.com with the subject line “PhD Student Position” or “Postdoc Position”.
Calgary, Canada’s fastest growing major city, is vibrant and multicultural with a population of more than 1.2 million. Situated near the Rocky Mountains, Banff National Park and Lake Louise, Calgary offers great quality of life and outstanding recreational activities.
A postdoctoral position is available in the group of Elke Ober at DanStem. The team studies fundamental mechanisms of tissue morphogenesis in the liver. The aim is to understand how different cell types communicate with each other and how the resulting cell behaviours establish an organ with a functional tissue architecture and of the correct size. To achieve this, the group utilises zebrafish genetic tools and state-of the-art molecular and cell biology techniques, such as mRNA sequencing, genome editing, confocal and light-sheet microscopy. This research project will employ a multi-disciplinary approach to understand how liver size is controlled combining zebrafish models of liver growth with human in vitro models. Starting date for this position is 1 May 2020, or upon agreement with the chosen candidate.
Institute:
The Novo Nordisk Foundation Center for Stem Cell Biology – DanStem is located at the University of Copenhagen. DanStem addresses basic research questions in stem cell and developmental biology and has activities focused on the translation of promising basic research results into new strategies and targets for the development of new therapies for cancer and chronic diseases such as diabetes and liver failure. Find more information about the Center at http://danstem.ku.dk/.
Job description:
We are seeking a highly motivated and ambitious postdoctoral candidate with a strong background in cell, developmental and/or stem cell biology to join our team. The candidate will investigate the molecular and cellular mechanisms controlling start and termination of organ growth in unique zebrafish liver mutants and human in vitro models of hepatic stem cell differentiation using e.g. advanced live cell imaging and transcriptomic approaches. The position is for 2 years with a possible extension.
Qualifications:
Candidates must hold a PhD degree in cell/developmental/stem cell biology or similar, with a track-record of successful scientific work.
Candidates should have a strong background in zebrafish genetics, cell biology and/or live cell imaging and advanced quantitative image analysis.
Previous experience in bioinformatics, next-generation sequencing and/or human stem cell differentiation are considered an advantage.
Good English communication skills, both oral and written, are prerequisite for the successful candidate.
Terms of employment:
The employment is planned to start 1 May 2020 or upon agreement with the chosen candidate. The full-time position is for 2 years with possiblity of extension. The terms of employment are set according to the Agreement between the Ministry of Finance and The Danish Confederation of Professional Associations or other relevant professional organization. The position will be at the level of postdoctoral fellow and the basic salary according to seniority. Currently, the salary starts at 34.360 DKK/approx. 4,.590 Euro (October 2019-level). A supplement could be negotiated, dependent on the candidate´s experiences and qualifications. In addition a monthly contribution of 17.1% of the salary is paid into a pension fund.
Non-Danish and Danish applicants may be eligible for tax reductions, if they hold a PhD degree and have not lived in Denmark the last 10 years.
The position is covered by the “Memorandum on the Job Structure of Academic Staff at Universities” of 11 December 2019.
Questions:
For further information contact Associate Professor Elke Ober, elke.ober@sund.ku.dk.
Application Instruction:
The application must be submitted in English, by clicking on “Apply online” below. Only online applications will be accepted.
The application must include:
Cover letter detailing the basis on which the applicant scientific qualifications meet the requirements for this position.
Curriculum vitae.
List of references (full address, incl. email and phone number)
Diplomas – all relevant certificates.
List of publications
Deadline for applications: 5 March 2020, 23.59pm.
The further process:
After the expiry of the deadline for applications, the authorized recruitment manager selects applicants for assessment on the advice of the Appointments Committee. All applicants are then immediately notified whether their application has been passed for assessment by an expert assessment committee. Selected applicants are notified of the composition of the committee and each applicant has the opportunity to comment on his/her assessment. You may read about the recruitment process at http://employment.ku.dk.
The applicant will be assessed according to the Ministerial Order no. 242 of 13 March 2012 on the Appointment of Academic Staff at Universities.
University of Copenhagen wish to reflect the diversity of society and welcome applications from all qualified candidates regardless of age, disability, gender, nationality, race, religion or sexual orientation. Appointment will be based on merit alone.
We are looking for a curious, enthusiastic and creative team member for new exciting projects in our lab (postdoc or staff scientist). The Fuhrmann Laboratory investigates the role of signaling pathways (Wnt, TGFbeta, Hippo, Hedgehog) during ocular development and regeneration in mouse. We investigate the cellular and molecular mechanisms regulating early eye patterning, RPE development and closure of the optic fissure during formation of the optic cup. We determine how signaling pathways control regeneration of the retinal pigment epithelium (RPE) in the adult eye. The lab is located in the Vanderbilt Eye Institute and is affiliated with the Cell and Developmental Biology Dept. at Vanderbilt University in Nashville, TN (https://www.vumc.org/fuhrmannlab/).
We are seeking candidates with a strong motivation in applying advanced imaging techniques and functional physiology techniques, and a solid background in cell biology, biochemistry and tissue culture paradigms to study aspects of ocular development and/or regeneration in mouse. Successful candidates must have a recent PhD in Life Sciences or equivalent with at least one first-author publication from their graduate work. We look forward to a talented and highly motivated team member. Candidates with less than 2 years of postgraduate work are specifically encouraged to apply. To apply, email a brief cover letter describing research accomplishments and future research goals, current CV with list of publications, and contact information for 3 professional references to:
Welcome to our monthly trawl for developmental biology (and related) preprints.
This month’s trawl includes a veritable farm, with developmental studies of potatoes, beetroot, tomato, maize, wheat, pigs, cows and goats, plus many of the usual suspects. They were hosted on bioRxivandarXiv. Let us know if we missed anything. Use these links to get to the section you want:
Differences in mitochondrial activity trigger cell competition during early mouse development
Ana Lima, Gabriele Lubatti, Jörg Burgstaller, Di Hu, Alistair Green, Aida Di Gregorio, Tamzin Zawadzki, Barbara Pernaute, Elmir Mahammadov, Marian Dore, Juan Miguel Sanchez, Sarah Bowling, Margarida Sancho, Mohammad Karimi, David Carling, Nick Jones, Shankar Srinivas, Antonio Scialdone, Tristan A. Rodriguez
Local retinoic acid directs emergence of the extraocular muscle functional unit
Glenda Comai, Marketa Tesarova, Valerie Dupé, Muriel Rhinn, Pedro Vallecillo Garcia, Fabio da Silva, Betty Feret, Katherine Exelby, Pascal Dollé, Leif Carlsson, Brian Pryce, Francois Spitz, Sigmar Stricker, Tomas Zikmund, Jozef Kaiser, James Briscoe, Andreas Schedl, Norbert B. Ghyselinck, Ronen Schweitzer, Shahragim Tajbakhsh
Cell-type specific impact of glucocorticoid receptor activation on the developing brain
Cristiana Cruceanu, Leander Dony, Anthi C. Krontira, David S. Fischer, Simone Roeh, Rossella Di Giaimo, Christina Kyrousi, Janine Arloth, Darina Czamara, Silvia Martinelli, Stefanie Wehner, Michael S. Breen, Maik Koedel, Susann Sauer, Monika Rex-Haffner, Silvia Cappello, Fabian J. Theis, Elisabeth B. Binder
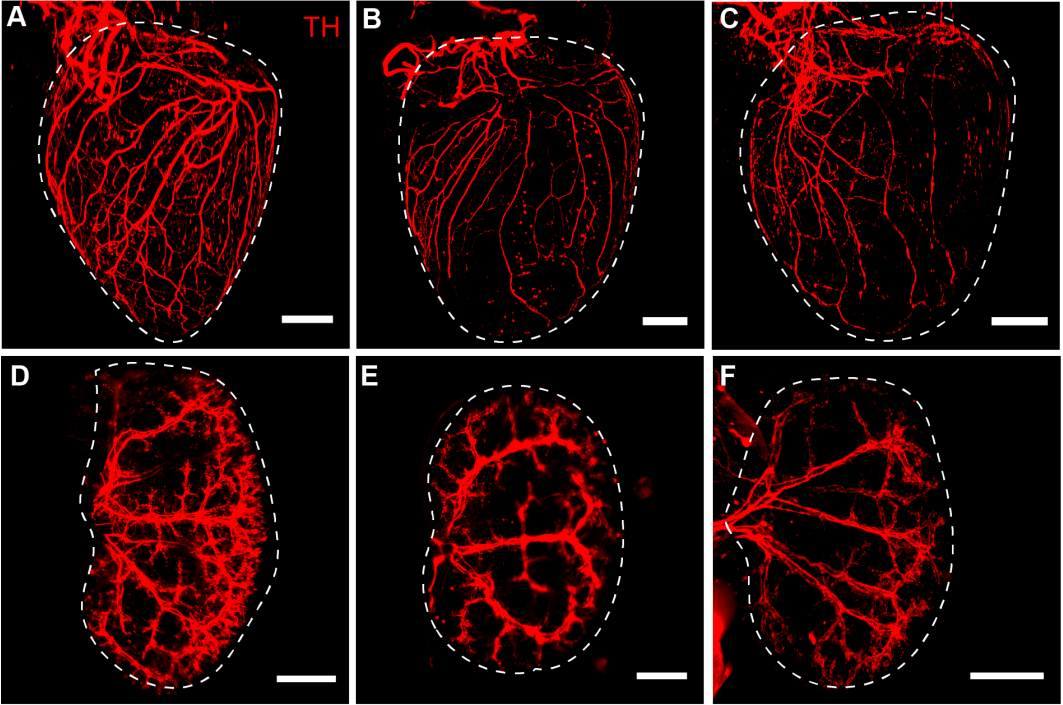
Hearts and kidneys from Cott-Solomon and Kuruvilla
Regulation of human development by ubiquitin chain editing of chromatin remodelers
David B. Beck, Mohammed A. Basar, Anthony J. Asmar, Joyce Thompson, Hirotsugu Oda, Daniela T. Uehara, Ken Saida, Precilla D’Souza, Joann Bodurtha, Weiyi Mu, Kristin W. Barañano, Noriko Miyake, Raymond Wang, Marlies Kempers, Yutaka Nishimura, Satoshi Okada, Tomoki Kosho, Ryan Dale, Apratim Mitra, Ellen Macnamara, Undiagnosed Diseases Network, Naomichi Matsumoto, Johi Inazawa, Magdalena Walkiewicz, Cynthia J. Tifft, Ivona Aksentijevich, Daniel L. Kastner, Pedro P. Rocha, Achim Werner
Hox-dependent coordination of cardiac progenitor cell patterning and differentiation
Sonia Stefanovic, Brigitte Laforest, Jean-Pierre Desvignes, Fabienne Lescroart, Laurent Argiro, Corinne Maurel-Zaffran, David Salgado, Christopher de Bono, Kristijan Pazur, Magali Théveniau-Ruissy, Christophe Béroud, Michel Pucéat, Anthony Gavalas, Robert G. Kelly, Stéphane Zaffran
Single cell epigenomic atlas of the developing human brain and organoids
Ryan S. Ziffra, Chang N. Kim, Amy Wilfert, Tychele N. Turner, Maximilian Haeussler, Alex M. Casella, Pawel F. Przytycki, Anat Kreimer, Katherine S. Pollard, Seth A. Ament, Evan E. Eichler, Nadav Ahituv, Tomasz J. Nowakowski
Molecular and genetic regulation of pig pancreatic islet cell development
Seokho Kim, Robert L. Whitener, Heshan Peiris, Xueying Gu, Charles A. Chang, Jonathan Y. Lam, Joan Camunas-Soler, Insung Park, Romina J. Bevacqua, Krissie Tellez, Stephen R. Quake, Jonathan R. T. Lakey, Rita Bottino, Pablo J. Ross, Seung K. Kim
Sox2 modulation increases naïve pluripotency plasticity
Kathryn Tremble, Giuliano G. Stirparo, Lawrence E. Bates, Katsiaryna Maskalenka, Hannah T. Stuart, Kenneth Jones, Amanda Andersson-Rolf, Aliaksandra Radzisheuskaya, Bon-Kyoung Koo, Paul Bertone, José C. R. Silva
Neural stem cells traffic functional mitochondria via extracellular vesicles to correct mitochondrial dysfunction in target cells
Luca Peruzzotti-Jametti, Joshua D Bernstock, Giulia Manferrari, Rebecca Rogall, Erika Fernandez-Vizarra, James C Williamson, Alice Braga, Aletta Van den Bosch, Tommaso Leonardi, Agnes Kittel, Cristiane Beninca, Nunzio Vicario, Sisareuth Tan, Carlos Bastos, Iacopo Bicci, Nunzio Iraci, Jayden A Smith, Paul J Lehner, Edit I Buzas, Nuno Faria, Massimo Zeviani, Christian Frezza, Alain Brisson, Nicholas J Matheson, Carlo Viscomi, Stefano Pluchino
ATR expands embryonic stem cell fate potential in response to replication stress
Sina Atashpaz, Sara Samadi Shams, Javier Martin Gonzalez, Endre Sebestyén, Negar Arghavanifard, Andrea Gnocchi, Eliene Albers, Simone Minardi, Giovanni Faga, Paolo Soffientini, Elisa Allievi, Valeria Cancila, Angela Bachi, Oscar Fernandez-Capetillo, Claudio Tripodo, Francesco Ferrari, Andrés Joaquin López-Contreras, Vincenzo Costanzo
Maternal genome dominance in early plant embryogenesis
Jaime Alaniz-Fabián, Daoquan Xiang, Gerardo Del Toro-De León, Axel Orozco-Nieto, Peng Gao, Andrew Sharpe, Leon V. Kochian, Gopalan Selvaraj, Nathan Springer, Cei Abreu-Goodger, Raju Datla, C. Stewart Gillmor
COP1 destabilizes DELLA proteins in Arabidopsis
Noel Blanco-Touriñán, Martina Legris, Eugenio G. Minguet, Cecilia Costigliolo-Rojas, María A. Nohales, Elisa Iniesto, Marta García-León, Manuel Pacín, Nicole Heucken, Tim Blomeier, Antonella Locascio, Martin Černý, David Esteve-Bruna, Mónica Díez-Díaz, Břetislav Brzobohatý, Henning Frerigmann, Matías D. Zurbriggen, Steve A. Kay, Vicente Rubio, Miguel A. Blázquez, Jorge J. Casal, David Alabadí
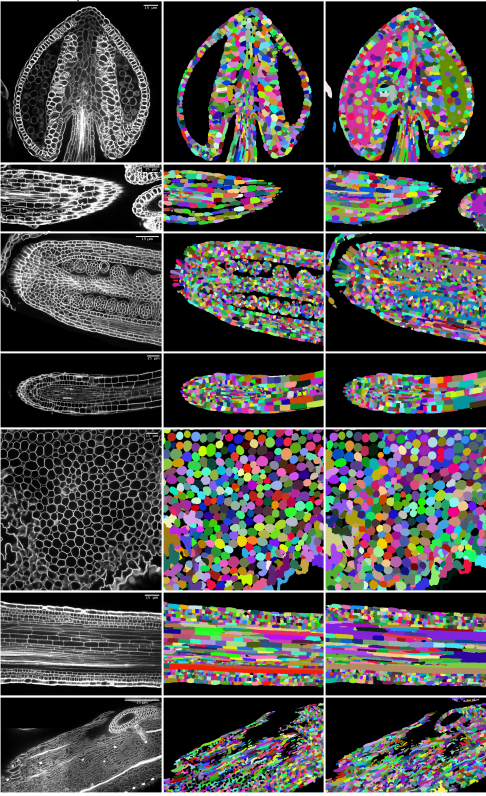
Accurate and Versatile 3D Segmentation of Plant Tissues at Cellular Resolution
Adrian Wolny, Lorenzo Cerrone, Athul Vijayan, Rachele Tofanelli, Amaya Vilches Barro, Marion Louveaux, Christian Wenzl, Susanne Steigleder, Constantin Pape, Alberto Bailoni, Salva Duran-Nebreda, George Bassel, Jan U. Lohmann, Fred A. Hamprecht, Kay Schneitz, Alexis Maizel, Anna Kreshuk
On the origin and evolution of RNA editing in metazoans
Qiye Li, Pei Zhang, Ji Li, Hao Yu, Xiaoyu Zhan, Yuanzhen Zhu, Qunfei Guo, Huishuang Tan, Nina Lundholm, Lydia Garcia, Michael D. Martin, Meritxell Antó Subirats, Yi-Hsien Su, Iñaki Ruiz-Trillo, Mark Q. Martindale, Jr-Kai Yu, M. Thomas P. Gilbert, Guojie Zhang
A chemical tool for improved culture of human pluripotent stem cells
Laurence Silpa, Maximilian Schuessler, Gu Liu, Marcus Olivecrona, Lucia Groizard-Payeras, Elizabeth Couper, Carole J. R. Bataille, Mark Stevenson, Len W. Seymour, Stephen G. Davies, William S. James, Sally A. Cowley, Angela J. Russell
CRISPR-Cas13d induces efficient mRNA knock-down in animal embryos
Gopal Kushawah, Joaquin Abugattas-Nuñez del Prado, Juan R. Martinez-Morales, Michelle DeVore, Javier R. Guelfo, Emry O. Brannan, Wei Wang, Timothy J. Corbin, Andrea M. Moran, Alejandro Sánchez Alvarado, Edward Málaga-Trillo, Carter M. Takacs, Ariel A. Bazzini, Miguel A. Moreno-Mateos
The Developing Human Connectome Project: typical and disrupted perinatal functional connectivity
Michael Eyre, Sean P Fitzgibbon, Judit Ciarrusta, Lucilio Cordero-Grande, Anthony N Price, Tanya Poppe, Andreas Schuh, Emer Hughes, Camilla O’Keeffe, Jakki Brandon, Daniel Cromb, Katy Vecchiato, Jesper Andersson, Eugene P Duff, Serena J Counsell, Stephen M Smith, Daniel Rueckert, Joseph V Hajnal, Tomoki Arichi, Jonathan O’Muircheartaigh, Dafnis Batalle, A David Edwards
Curious about novel lymphatic cell types in the meninges?
Excited to explore links between non-neuronal cells, brain activity, and sleep using zebrafish as a model system?
Then consider the Research Associate Position NOW available in the Rihel lab at University College London (Deadline Feb 22, 2020).
Details: Full Time, Grade 7.
Salary (inclusive of London allowance) £35,965 – £43,470 per annum
Duties and Responsibilities: The post-holder will research the genetic and cellular mechanisms of zebrafish sleep in the laboratory of Dr. Jason Rihel. They will join a team of researchers working on various aspects of sleep behaviours in zebrafish, with a focus on the interaction between the brain and a novel lymphatic cell type in the meninges. This position is funded for a period of 24 months in the first instance and will start immediately.
Key Requirements: Applicants must have a PhD in Neurobiology, Genetics, or a related field, together with significant experience in genetics, molecular biology, in vivo imaging, and behavioral neuroscience of a model organism such as zebrafish.
IRIBHM is a research institute of the Medical School of the Free University of Brussels (Université Libre de Bruxelles, ULB). The institute offers an internationally prominent research environment in molecular biology and life sciences, that engage different topics that span receptor pharmacology and new therapeutic targets discovery, early embryonic development, neurobiology, stem cells and cancer. The institute has trained over the years a number of talented young scientists both at the graduate and postdoctoral levels.
In order to expand internationally and keep rising its level of excellence, the IRIBHM (https://iribhm.org/) launches an international graduate programme in order to prepare the future leaders in biomedical sciences. At least 2 PhD scholarships are available. The successful candidates will have the opportunity to work in a warm and stimulating research environment aligned with the highest international standards.
As the administrative center of the European Union, Brussels is a perfect location for an International PhD programme. In addition, the city shows an active cultural life and is hosting most nationalities from all over the world.
We look forward to welcoming you in Brussels!
Requirements:
Diplomas and degrees equivalent to a European Union Master’s degree, which includes project work summarized in a written “small thesis”
Two referees willing to provide letter of recommendations
Excellent knowledge of English
Excellent interpersonal and organisational skills
Closing date for CV and letters submission: June 30.
Accepted candidates may start research projects as early as November 2020
The Lindstrom Laboratory in the Broad CIRM Center and Department of Stem Cell Biology and Regenerative Medicine, Keck School of Medicine of USC, has an opening for a postdoctoral scholar to study the transcriptional regulation underpinning the formation of functional cell-types in the kidney from stem and progenitor populations in the developing embryo. Applicants are expected to have a Ph.D. in a discipline of biological sciences or expect to complete it by the date of hire. Preference will be given to candidates with research experiences in developmental biology and genetics, stem cells and tissue engineering. However, all interested candidates are encouraged to apply.
The Lindstrom lab focuses on understanding how cells coordinate their behavior during development in order to form complex tissues that perform critical physiological functions. This enables us to make important insights into how birth defects occur, form strategies to replicate developmental process in stem-cell based systems, and to allows us to develop new regenerative therapies. Our main focus is on the kidney, where we apply state-of-the-art technologies such as single-cell omics, confocal imaging, and organoid models to delineate mechanisms controlling its formation and function. Additional information can be found at https://lindstromlab.usc.edu.
We are seeking a highly motivated and passionate individual wishing to take a leading role in developing their research projects while also contributing and gaining experience in writing grants and fellowship applications. The successful candidate will be expected to also oversee the work of other lab personnel and coordinate their research with fellow lab members. The candidate should anticipate engaging in highly collaborative and cross disciplinary research.
The Broad CIRM Center and Department of Stem Cell Biology and Regenerative Medicine, Keck School of Medicine of USC (http://keck.usc.edu/stemcell/) offers an exciting and nurturing research environment with great opportunities to conduct cutting edge science, use of latest equipment, and postdoctoral training and support. A weekly seminar series invites leading scientists in the stem / regenerative / synthetic / and developmental sciences (https://stemcell.keck.usc.edu/events/) and at regular intervals postdoctoral scientists present their data to the department.
When applying, please submit the required documentation online. A cover letter should highlight why you are interested in joining the lab, projects that you may wish to develop, and how your existing experience will synergize with our goals (2 pages max). In addition, please provide the names of a minimum of two references.
The Seifert lab in the Department of Biology at the University of Kentucky (UK) is seeking an exceptionally creative postdoctoral researcher to join our group. The successful candidate will lead an effort to isolate stable embryonic stem cell lines and/or derive iPSC lines for genomic modification and subsequent generation of transgenic spiny mice. This project will provide freedom to work on additional projects related to early embryonic development in spiny mice or projects investigating the cellular and molecular basis for complex tissue regeneration in mammals. To pursue these lines of research we maintain an active breeding colony of spiny mice (Acomys cahirinus) at UK.
Ideal candidates will have a strong background in developmental biology, experience working with ESCs and iPSCs and mouse transgenesis, experience working with model or non-model organisms, expertise in microscopy and comfortability with bioinformatics. While this is a funded position, postdocs in the Seifert lab are strongly encouraged to develop their own projects and external funding portfolios as a pathway toward independence. Salary follows NIH guidelines for postdoctoral researchers. Informal inquiries by email are strongly encouraged. For additional information visit: http://www.ashleyseifert.com/opportunities.html
Review of applications will begin on a rolling basis and will continue until the position has been filled. Ideal start date is Spring 2020. Candidates will have completed their Ph.D. prior to starting the position but need not have defended their dissertation prior to applying. Applicants should submit their application materials through the UK Jobs site (upload under Specific Request 1) at http://ukjobs.uky.edu/postings/251962. In addition, applicants should send a single pdf document to Ashley Seifert (awseifert@uky.edu) that includes their CV, names of three references, and a 1-2-page synopsis of their current research interests and how these complement our overall research program.
The Department of Biology houses a strong group of research labs interested in regenerative and stem cell biology using a diverse array of animal models (e.g., spiny mice, salamanders, planarians, lampreys, zebrafish) and in vitro systems. Together, these labs create a vibrant atmosphere to pursue interdisciplinary projects across comparative genomics, developmental, regenerative and evolutionary biology.
The University of Kentucky is an Equal Opportunity Employer and encourages applications from veterans, individuals with disabilities, women, African Americans, and all minorities.
 (1 votes)
(1 votes) (No Ratings Yet)
(No Ratings Yet)