The Department of Biology at the University of Kentucky (UK) is seeking applications for the Thomas Hunt Morgan Postdoctoral Fellowship in the Seifert Lab. This fellowship provides exceptional PhDs an opportunity to conduct independent research in any aspect of regenerative biology. The Fellows are funded for two years through the College of Arts and Sciences and provided support to develop their own projects while they build external funding portfolios as a pathway toward independence. The Seifert lab uses an array of animal models to study complex tissue regeneration and maintains large, active breeding colonies of spiny mice (Acomys cahirinus) and salamanders at UK.
The Department of Biology houses a strong group of research labs interested in regenerative and stem cell biology using a diverse array of animal models (e.g., spiny mice, salamanders, planarians, lampreys, zebrafish) and in vitro systems. Together, these labs create a vibrant atmosphere to pursue interdisciplinary projects incorporating cutting edge science in regenerative, developmental, and evolutionary biology.
Review of applications will begin on a rolling basis and will continue until a candidate is selected. Candidates will have completed their Ph.D. prior to starting the position but need not have defended their dissertation prior to applying.
Interested applicants should apply online at http://ukjobs.uky.edu/postings/251988. Applicants should submit their application materials through the UK Jobs site (upload under Specific Request 1) as well as send a single pdf document to Ashley Seifert (awseifert@uky.edu). Application materials submitted through UK Jobs as well as submitted to Dr. Seifert should include a CV, names of three references, and a 2-page statement of proposed research interest. Informal inquiries by email are strongly encouraged. More information is available here: http://www.ashleyseifert.com/opportunities.html
In this episode exploring great ideas in genetics, we’re discovering our inner fish – finding out whether we really do go through a fishy phase in the womb, and looking at the legacy of Tiktaalik, the first fish to walk on land.
Born in 1834, Ernst Haeckel was a German zoologist with a flair for illustration – and a knack for creating incredibly detailed and widely shared scientific images. But do his infamous embryo drawings really show the true picture of early development?
Haeckel thought that we went through a ‘fish’ stage in the womb because our embryos appear to have gills during early development. Although his theory that ‘ontogeny recapitulates phylogeny’ has subsequently been shown to be incorrect, we now know there is a close connection between development and evolution, or ‘evo-devo’ as it’s sometimes known.
In short, our evolutionary history is written in our developmental genes, and it’s a history that we can trace right the way back to the very first vertebrates. The best example of this is Tiktaalik – our oldest ‘fishapod’ ancestor that forms the missing link between fish and land-dwelling tetrapods.
If you enjoy the show, please do rate and review on Apple podcasts and help to spread the word on social media. And you can always send feedback and suggestions for future episodes and guests to podcast@geneticsunzipped.com Follow us on Twitter – @geneticsunzip
Established by the British Society for Developmental Biology in 2014, The Gurdon/The Company of Biologists Summer Studentship scheme provides financial support to allow highly motivated undergraduate students an opportunity to engage in practical research during their summer vacation. Each year, ten successful applicants spend eight weeks in the research laboratories of their choices, and the feedback we receive is outstanding. You can read accounts from previous years here. If you’re interested in applying or hosting a student in 2020, applications need to be in by the end of March.
Our tenth and final report from the class of 2019 comes from Jake Cornwall Scoones who studied developmental mechanics in the fly embryo in Bénédicte Sanson’s lab at Cambridge.
Developing force inference strategies to analyse tissue-wide stress in the Drosophila germband
Morphogenesis is an integral aspect of embryonic development, shaping collections of cells into functional tissues. Developmental mechanics aims to unravel how forces across spatial scales, from cytoskeletal processes, to junctional dynamics and inter-tissue interactions, facilitate emergent transformations in embryonic form. Analysing mechanics across scales requires a systems-level approach, where quantifying tissue-wide stress and material properties is essential. Current methods for directly measuring forces in vivo have major short-comings: methods (e.g. laser ablation) are invasive, perturbing the stress field and prohibiting extensive sampling. Non-invasive force inference approaches are useful alternatives, using segmented images of fluorescently-labelled cells to infer relative tension across each cell membrane and pressure in every cell. They do this by fitting tissue-wide tension and pressure nets to these data via the assumption of force-balance.
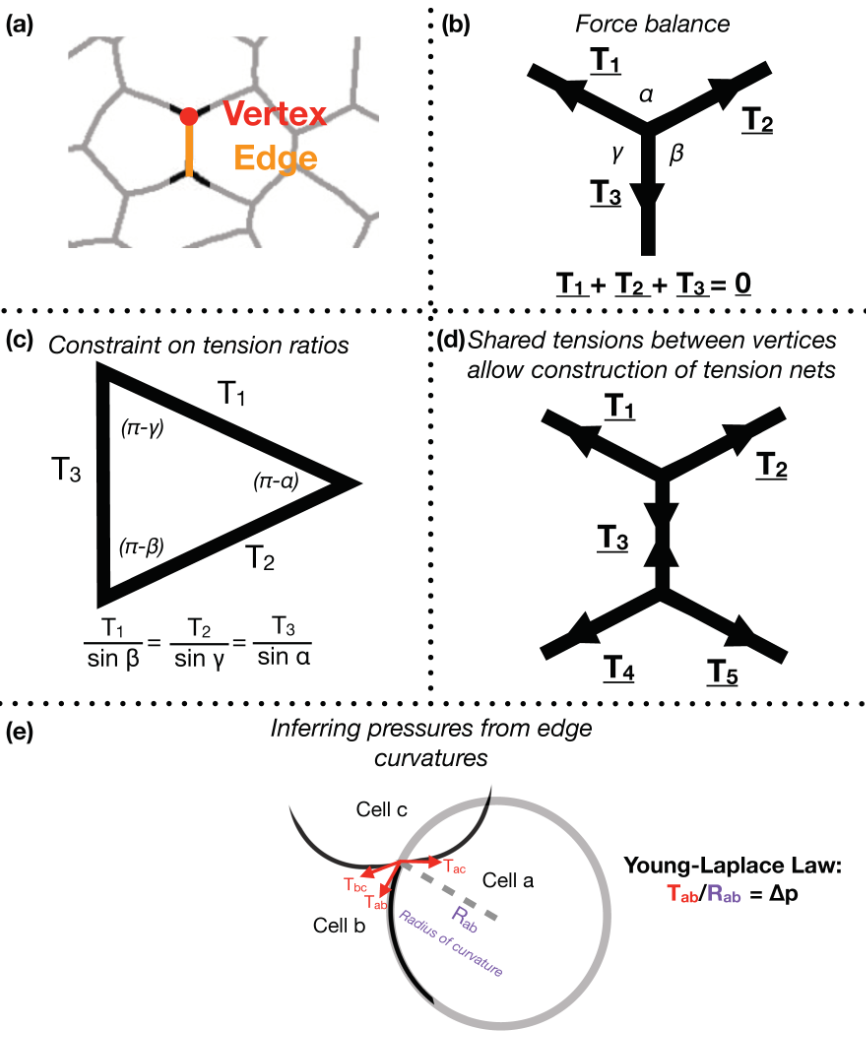
Focusing on Drosophila germ-band extension, a well-studied example of axis extension, I developed computational tools for non-invasive inference of stress distributions from microscopy data. These algorithms consider an epithelial sheet as a 2D curved-polygonal lattice, with edges demarcating cell membranes, and vertices defined where three cells meet (Fig. 1a). Using some simplifying physical assumptions, they use the geometries of edges at each vertex, as well as the curvatures at each edge, to infer relative pressures in every cell and relative tensions at every edge.
Figure 1
Despite the variation in computational implementation strategy, many existing force inference methods have a common underpinning logic. Specifically, they assume that the tensions at every vertex are balanced; in other words, that the system is in mechanical equilibrium (Fig. 1b). Thus given the angles that edges of a given vertex make with each other, as inferred from microscopy data, one can infer ratios of relative tensions (Fig. 1c). Further, they assume that tension is continuous across an edge, meaning relative tensions can be inferred tissue-wide (Fig. 1d). Cellular pressures can also be inferred. When edges are curved, it is assumed that the cell for which the edge is convex shows higher pressure. In fact, one can infer the pressure differential in terms of the inferred tension and the radius of curvature of that edge using the Young-Laplace law (Fig. 1e).
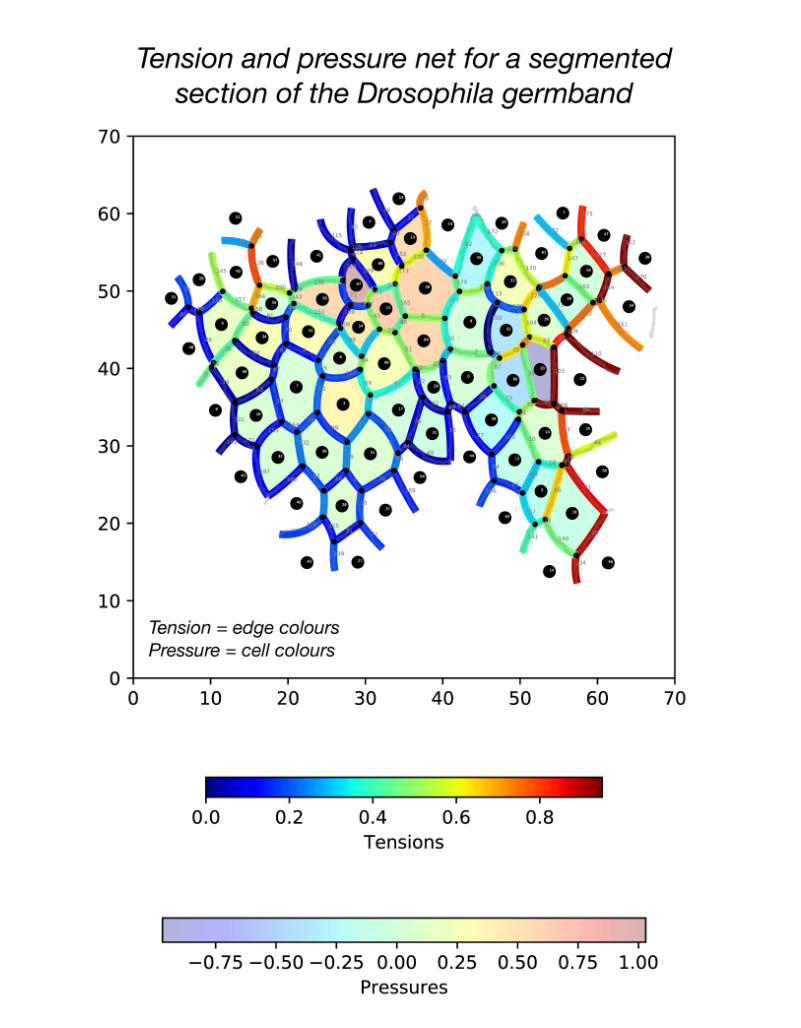
During my stay with the Sanson lab, I developed a pipeline in Python for force inference. The pipeline takes the many frames of segmented germband movies curated by the lab, uses one of several possible force inference strategies, and outputs the inferred tensions and pressures back to the lab’s wider computational workflow (Fig. 2). Given the wealth of data collected by the lab, these strategies now have immediate use in analysing global patterns of stress (indeed without the need for additional experiments), for example in characterising non-intuitive changes to stress distributions in key morphogenetic mutants (e.g. sidekick mutants, see Finegan et al 2019, PloS Biology, https://doi.org/10.1371/journal.pbio.3000522). Besides direct analyses of stress distributions, these inferences can now be used to parameterise physical models of the germband, across spatial scales. Beyond this, I have compared the various implementation strategies with regards to their concordance in predictions, and tested their relative capacities of dealing with noise in segmentation. I have also surveyed both the practical and conceptual challenges of these strategies with an intention of refining existing methodologies for faster and more accurate inference.
Figure 2
This project has allowed me to refine skills in programming, especially in visualisation, data processing and algorithm optimisation. I am now putting the algorithms together into a coherent and user-friendly package for use in the lab internally, but also, excitingly, to be released as an open-access resource. Further, working in Dr. Sanson’s lab also introduced me to key concepts in mechanics, situated in this biological context.
Given the highly interdisciplinary makeup of this project, supervised by Dr. Alex Nestor Bergmann (mathematician and physicist), Dr. Guy Blanchard (computational biologist), and Dr. Bénédicte Sanson (developmental biologist), I enjoyed how divergent perspectives and methodologies are brought together to solve this multi-scale problem. I endeavour to pursue similar interdisciplinary research in a PhD and beyond, carrying out both dry and wet components.
I would like to thank Dr. Alex Nestor Bergmann and Dr. Guy Blanchard for their fantastic supervision and the Sanson lab as a whole for their support and their fascinating and diverse lab meetings. I finally want to thank the BSDB for providing me the opportunity to carry out this research, and urge other undergraduates to apply to the Gurdon Studentship.
A Ph.D. position in molecular plant cell biology is available in the lab of Prof. Kay Schneitz, Dept. of Plant Developmental Biology, Technical University of Munich in Freising/Germany.
Plant cells are encapsulated by a semi-rigid and biochemically complex cell wall. Cell wall remodeling is central to cell growth as well as the response to biotic or abiotic stresses. The molecular mechanism monitoring cell wall integrity in plants is poorly understood. Recent data from our lab revealed that signaling mediated by the Arabidopsis receptor kinase STRUBBELIG (SUB), previously known for its function in controlling morphogenesis, plays a central role in this process (1-5). The successful candidate will investigate how SUB signaling controls the response to cell wall damage. Preferred starting date is spring/early summer 2020 but is negotiable. The lab is part of the Collaborative Research Centre SFB924 (sfb924.wzw.tum.de) and thus funding is at the usual TV-L E13/2 level. Requirements are e.g. a German masters (with a mark of 2.5 or better), a French DEA (a final average of 13 or more), or a masters thesis.
We are looking for a highly motivated scientist well-trained in molecular and cell biology with a strong interest in interdisciplinary work at the interface of plant cell and developmental biology and the response to stress. The person should have excellent problem-solving skills and be able to work independently. Fluency in English is a must. Freising is located about 35 km to the north of Munich. Munich is a lively, cosmopolitan city close to beautiful lakes and the Alps. For further information please contact Kay Schneitz (kay.schneitz@tum.de) and visit the webpage (plantdev.wzw.tum.de).
Please submit your application as a single PDF file by email to office.plantdev@wzw.tum.de. TUM is an equal opportunity employer. Applicants with disabilities are treated with preference given comparable qualifications. Position is open until filled.
For further information please contact:
Prof. Dr. Kay Schneitz
Plant Developmental Biology, School of Life Sciences, TUM, D-85354 Freising
Email: kay.schneitz@tum.de
URL: http://plantdev.wzw.tum.de
Dysregulated activity of cell surface proteolytic enzymes has a wide range of developmental and pathological consequences, but the underlying mechanisms are often poorly understood. A new Development paper uses mice to model a severe inherited form of enteropathy and the role of the serine protease matriptase in the disease’s progression. We caught up with first author Roman Szabo and his supervisor Thomas Bugge, Senior Investigator at the NIH National Institute of Dental and Craniofacial Research in Bethesda, Maryland, to find out more about the story.
Thomas (L) and Roman (R)
Thomas, can you give us your scientific biography and the questions your lab is trying to answer?
TB I did my PhD research at the European Molecular Biology Laboratory in Heidelberg working on nuclear receptors, and my postdoc at the University of Cincinnati studying fibrinolytic enzymes. I have been a Senior Investigator at the National Institute of Dental and Craniofacial Research, the National Institutes of Health, since 1999. Our laboratory studies how extracellular/pericellular proteases – a group of enzymes several hundred strong – signal to enable vertebrate embryonic development and maintain postnatal tissue homeostasis. We also study how these enzymes, when misregulated, cause human disease. This is a distinctly understudied area, and a fun research space to be in.
Roman, how did you come to work with Thomas, and what drives your research today?
RS As a graduate student I worked with yeasts and how they regulate their cell morphology based on environmental conditions. As the particular species of yeast I was working with typically thrives in a lipid- and protein-rich environment, its behaviour is in part regulated by a battery of proteolytic enzymes that it secretes into the medium in order to digest the proteins to use them as a source of nutrients. After defending my PhD, I was looking for a lab that would allow me to use at least some of that knowledge in experimental systems more related to human physiology. Attracted by both the reputation of the NIH as a leading research institution and Thomas’ impressive prior work in the field of extracellular proteolysis using genetically modified mouse models, I contacted him and it turned out he was just looking for a new postdoc. As I had never worked with mice before, it was probably a bit of a risk, on both our parts, but it was actually quite interesting, and in the end it seemed to have worked out well. And I was lucky that the lab had just started to work on a previously unknown family of proteolytic enzymes that turned out to be quite important for mammalian development and physiology. Learning about different ways in which these enzymes contribute to regulation of various biological processes has been really exciting and, I expect, will keep us busy for the foreseeable future.
How much do we understand about the role of cell-surface proteolytic enzymes in development?
RS & TB Still far too little. These are not the easiest enzymes to work with, at least biochemically. Insights into their developmental functions have been obtained predominantly through the use of reverse genetics in mice and fish, or through the clinical phenotyping of humans and animals with autosomal recessive mutations in their corresponding genes. These efforts have been successful in teasing out a diverse array of functions for individual cell-surface proteolytic enzymes, ranging from sperm maturation to placental development, formation of the feto-maternal interface, auditory system development, tight junction formation, epidermal barrier acquisition, hair growth and skin pigmentation. However, why cell-surface proteolytic enzymes are crucial for so many developmental processes is in large part waiting to be discovered.
And how did you come to work specifically on HAI-2 and matriptase?
RS & TB We came to work with matriptase quite accidentally while performing a screen for novel proteases putatively involved in skin wound healing. As it turned out, one of matriptase’s many developmental functions proved to be skin formation. Alas, we never got round to testing the function of matriptase in skin wound healing, but we have stayed with this fascinating and frustrating enzyme ever since. HAI-2 is structurally quite similar to the well-established matriptase inhibitor HAI-1, so it was an obvious candidate for a second matriptase inhibitor, and, indeed, turned out to be so.
Can you give us the key results of the paper in a paragraph?
RS & TB We provide evidence that intestinal failure in the syndromic form of the human disease congenital tufting enteropathy (CTE), which is caused by loss of HAI-2, may be due to matriptase hyperactivity. Specifically, we show that conditional ablation of matriptase from the mouse intestine prevents key features of the disease from developing. These include villous atrophy, luminal bleeding, loss of mucin-producing goblet cells, loss of defined crypt architecture and intestinal inflammation. The loss of the cell-cell junction proteins EpCAM and claudin 7 – a hallmark of CTE – was also prevented by the elimination of matriptase from the intestine. Consequently, the mice were able to gain weight and showed extended life-span. The study thus uncovers a potential therapeutic target for this devastating disease.
What do you think HAI-2 is doing, independently of matriptase, in later intestinal development?
RS & TB That is a very good question. HAI-2 is a serine protease inhibitor, so it is only natural to assume that it may need to regulate another serine protease expressed later in intestinal development. Unfortunately, HAI-2 is quite promiscuous, at least in the test tube, where it can inhibit a large number of serine proteases. So it may take a while to track down the culprit.
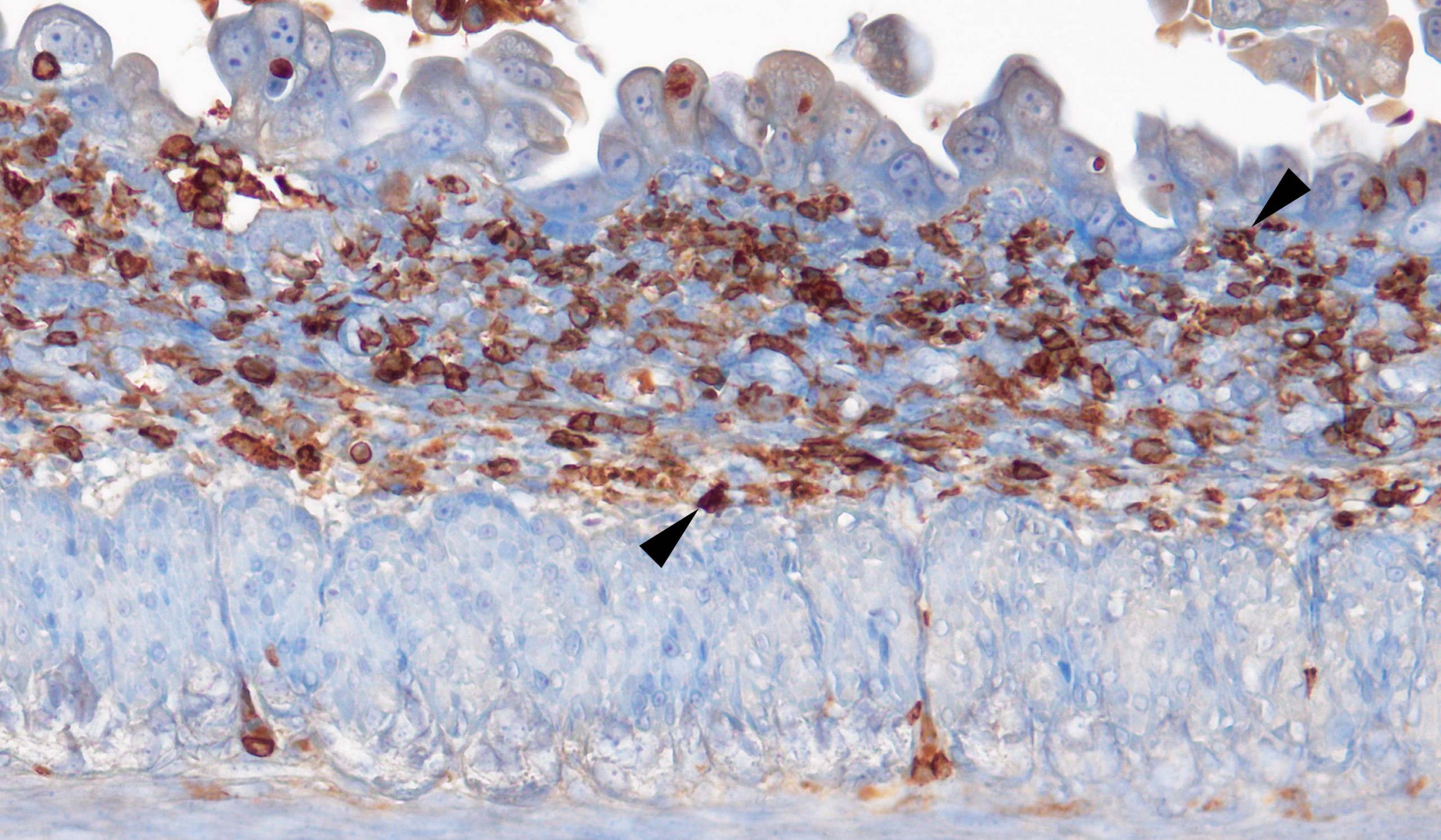
Staining for CD45-positive immune cells (brown) recruited to the site of matriptase-driven intestinal tissue damage.
How might matriptase be targeted therapeutically for CTE?
RS & TB Small molecule active-site inhibitors with selectivity for matriptase would be an obvious choice. Especially inhibitors with poor intestinal absorption that are less likely to affect important functions of matriptase in other parts of the body.
When doing the research, did you have any particular result or eureka moment that has stuck with you?
RS In our research, we rely heavily on designing new genetically modified mouse strains. It can take many months, even years, just to generate animals with the desired gene combination. So after all that work, it is always very exciting for me to see a new phenotype, to be the first person who knows that a certain gene does something really important. In this particular project, it was generating the first HAI-2-deficient mouse, in which elimination of matriptase dramatically increased life-span, and then when the analysis of the tissues from these mice showed that we have indeed prevented the intestinal damage caused by loss of the inhibitor.
It is always very exciting for me to see a new phenotype, to be the first person who knows that a certain gene does something really important
And what about the flipside: any moments of frustration or despair?
RS Not that much on this project, which turned out to be very straightforward. But we did have our share of frustration in the past. Before the invention of new gene targeting technologies, such as CRISPR/Cas, generation of genetically modified mice relied on introducing desired mutations into mouse embryonic stem cells, a process that was inherently ineffective, while at the same time slow and labour intensive. We once spent close to 2 years trying to generate one specific mouse strain, using every tool available to us at the time, before finally having to admit defeat. Similarly, we still lack tools to properly analyse many of the molecules we work with, including matriptase and HAI-2, in living tissues. It can be quite frustrating to have a promising hypothesis and not being able to find a way of testing it.
So what next for you after this paper?
RS Now that we have shown a connection between matriptase activity and intestinal failure in our mice, we would like to understand exactly how matriptase is causing all that damage. There is some evidence that it may do so by cleaving an important structural protein called EpCAM that is frequently lost in patients with CTE. We have already initiated experiments that would help us test whether that is indeed the case. In addition, the function of matriptase and HAI-2 in development is not limited to intestines, and we would like to develop new tools that would help us better analyse both physiological and pathological roles of these two important proteins. And besides my own projects I also have the privilege of organising the next meeting focusing on proteolytic enzymes, the class of proteins matriptase belongs to, which traditionally brings together leading scientists from all over the world to discuss latest advances in the field. It should be an exciting experience.
Where will this work take the Bugge lab?
TB Hopefully one small step further towards understanding the good and the bad side of an enigmatic enzyme, matriptase.
Finally, let’s move outside the lab – what do you like to do in your spare time in Bethesda?
TB Spend time with my wife and enjoy the beautiful nature we are surrounded by.
RS I am a very outdoorsy person so whenever time and weather permit, I like to go hiking, bird watching, wildlife photographing or fossil hunting. Fortunately, there are plenty of opportunities for all that here in the state of Maryland, from the Appalachian mountains to the Atlantic coastline. After a week of work at the bench and behind a computer, it is great to unwind and spend some time with my family going to a park or a beach. My daughter enjoys everything from nature to history and arts, so having a unique collection of national museums in nearby Washington, D.C., also helps.
NEUcrest is a four-year project, funded by the European Union Horizon 2020 Programme. The neural crest is an essential stem cell population of the vertebrate embryos. The project focuses on integrating academic, clinical and industrial research for a better understanding of neural crest development and neural crest related diseases. The NEUcrest network comprises 20 partners in academia, industry and hospitals from seven European countries.
Projects are available in the following labs and companies.
Applicants are encouraged to apply to more than one project if they are interested. Please note some different deadlines apply:
ESR 5, ESR 6 supervised by Grant Wheeler (University of East Anglia, Norwich, UK)
Training for transverse skills in outreach and industrial managements are deeply embedded in the programme. The NEUcrest ITN and PhD project is due to start in January 2020. Studentships can start anytime from 1 January to 1 April 2020. Candidate Specification: First degree or Masters in Biological Sciences, Cell Biology, Genetics and Molecular Biology. Mobility requirement: EU applicants are eligible to apply to all positions, non-EU applicants are eligible to a subset of the positions. Applicants must not have been based in the country of desired Ph.D. position for more than 12 months in the last 3 years prior to recruitment.
Please note that in order to demonstrate fair equal recruitment and to provide statistical data on the recruitment for MSCA program NEUcrest management team may retain the following personal data of all applicants: full name, gender, nationality, copy of the CV.
The data will be preserved in the internal repository of Institut Curie till maximum up to 5 years after the termination of the NEUcrest grant. The security of the data is provided and guaranteed by the centre for information processing of Institut Curie.
By applying for the advertised positions the applicant automatically gives the authorization to store his/her personal data. The NEUcrest consortium will not share personal data with third parties outside the consortium; however, data may be transmitted to bodies responsible for monitoring, inspection or regulatory tasks under EU law, e.g. the European Anti-Fraud Office (OLAF) or the teams performing administrative investigations.
The applicant may refuse, without having to give any explanations, the preservation of the data. In this case he/she needs to inform about it the management team of NEUcrest consortium upon submitting the application or sending a request at daria.barsuk@curie.fr or neucrest@gmail.com.
A postdoctoral position is opened in the laboratory of Dr. Julien Royet, at Aix- Marseille University, France. The lab is part of the IBDM Institute, providing a highly rich and international scientific environment, and is integrated into the Centuri excellence network.
Our team is interested in understanding the mechanisms by which bacteria are interacting with the nervous system of their eukaryotic host and how this interaction affects their behavior. We have recently shown that some neurons of the Drosophila central nervous system are expressing proteins that sense some bacteria metabolites. Using genetics and imaging techniques we have demonstrated that the direct sensing of gut-bacteria derived metabolites can directly impact the activity of theses neurons and, as a consequence, of that of their host (Kurz et al, eLife, 2017; Charroux et al, Cell Host and Microbes, 2018, Masuzzo et al, eLife, 2019). The postdoctoral project will aim at further dissecting the molecular and functional picture of these bacteria-neuron interactions.
Applicants must have completed a PhD thesis in the neurobiology field using the Drosophila model. They should be highly motivated, with good interpersonal and communication skills. Fluency in English is mandatory but ability to speak French is not required. Projects are funded by a grant from the FRM (Equipe FRM) for 2 years. Salary will depend on previous experience.
Applications should contain a CV, a letter of motivation with a description of research accomplishments and the contact information of two references to Julien.royet@univ-amu.fr.
The Bray lab has an opening for two Research Associates. Our collaborative, multidisciplinary team is investigating Notch regulatory networks in development and their relevance to disease. One project will focus on epigenetic mechanisms that remodel enhancers during cell fate transitions to programme the correct response to signalling (for example to maintain stem cells or promote differentiation depending on the context) and that enable enhancers to select the correct promoter. The other project will investigate how Notch signals are decoded in real time in vivo and will involve live-imaging of transcription and of nuclear complexes, including single molecule tracking of transcription factors in living tissues. The projects build on exciting assays we have recently developed (Falo Sanjuan et al., 2019, Dev Cell 50:411-425.e8; Gomez-Lamarca et al., 2018, Dev Cell 44:611-623.e7) and will involve generating new tools, including optogenetics, through genome editing and other approaches.
We are looking to recruit highly motivated scientists with strong investigative skills. You should have a PhD (or soon to be awarded your PhD) and experience in one or more of the following, molecular biology, epigenetics, developmental biology, cell biology, Drosophila genetics. Experience of genome-wide transcriptional analysis would be an advantage for one position and of advanced imaging and image analysis for the other.
A 2-year postdoctoral position in developmental biology is available in the group of Grigory Genikhovich at the Department of Neurosciences and Developmental Biology of the University of Vienna. The successful candidate will work on an Austrian Science Foundation-supported project “BMP shuttling and the evolution of animal bilaterality” performed in collaboration with the group of Prof. Dr. Siegfried Roth at the University of Cologne. The work will involve comparing the shuttling capacity of a range of cnidarian and bilaterian chordin proteins using Drosophila S2 cell line as well as Drosophila embryos as test models. The goal is to evaluate the role of Chordin-dependent shuttling of BMP ligands in the evolution of bilateral body symmetry in animals.
The successful candidate must hold a PhD in biology or a related field and be first or co-first author on at least two peer-reviewed publications. The position requires experience in Drosophila genetics and knowledge of the standard molecular biology techniques, as well as excellent oral and written English skills. The postdoc is expected to perform part of the project in the Roth lab at the University of Cologne.
How to apply:
Please send a single pdf file containing a cover letter, CV and contact information of three referees to grigory.genikhovich@univie.ac.at before 20.02.2002
 (No Ratings Yet)
(No Ratings Yet) In this episode exploring great ideas in genetics, we’re discovering our inner fish – finding out whether we really do go through a fishy phase in the womb, and looking at the legacy of Tiktaalik, the first fish to walk on land.
In this episode exploring great ideas in genetics, we’re discovering our inner fish – finding out whether we really do go through a fishy phase in the womb, and looking at the legacy of Tiktaalik, the first fish to walk on land. 

 (1 votes)
(1 votes)
 _____________________________________________________________________
_____________________________________________________________________