NSF and AHA funded post-doctoral position available in the Davidson lab at Swarthmore College.
Searching for a post-doc who is passionate about both teaching and research. We are studying the interplay between division and inductive signaling. In particular, we are exploring how signaling receptors are trafficked in dividing cells to generate asymmetric induction of a cardiac progenitor lineage. We are also using comparative genomics to explore the evolutionary constraints that shape gene regulatory networks. We study these questions in the invertebrate chordate, Ciona robusta. Ciona embryos consists of extremely low cell numbers allowing high resolution analysis of intra-cellular dynamics in intact embryos. The ease of generating transgenic Ciona embryos make this an excellent model organism for undergraduate research. We have recently initiated a collaboration with Danelle Devenport at Princeton focused on similar processes in cultured mouse epithelial cells. This is a great opportunity for a post-doc with an interest in undergraduate teaching and research. My former post-doc recently started a tenure-track position at a small liberal arts college and it was clear that the combination of a strong research record along with a demonstrated commitment to undergraduate mentoring and teaching made her a strong candidate for this very competitive career track.
Applicants should have a PhD (or be close to completing one) in a relevant subject area. Excellent communication skills and a commitment to undergraduate mentoring are essential.
To apply or if you have questions about the position – please send your CV and a cover letter describing your interest to Bradley Davidson at bdavids1@swarthmore.edu. I will be in touch with instructions for submitting a formal application.
Established by the British Society for Developmental Biology in 2014, The Gurdon/The Company of Biologists Summer Studentship scheme provides financial support to allow highly motivated undergraduate students an opportunity to engage in practical research during their summer vacation. Each year, ten successful applicants spend eight weeks in the research laboratories of their choices, and the feedback we receive is outstanding. You can read accounts from previous years here. If you’re interested in applying or hosting a student in 2020, applications need to be in by the end of March.
Our sixth report from the class of 2019 comes from Matyas Bubna (QMUL) who studied frog neural crest in Roberto Mayor’s lab at UCL.
Visualising neural crest induction, migration and differentiation in Xenopus
Throughout my undergraduate studies I have become increasingly captivated by the intricacy and elegance of animal development. Especially interesting to me is how processes such as morphogenesis, tissue patterning or cell migration, which can appear incomprehensible, emerge from relatively simple interactions at the molecular level. Observing how a single cell transforms into a complex organism is a unique and thought-provoking experience. I am grateful to the BSDB for the studentship allowing me to explore this field. I would strongly recommend the Gurdon studentship to anyone interested in topics ranging from evolution, epigenetics, cell signalling, cancer, to stem cells and regeneration – all of these processes may be elucidated by taking a developmental point of view.
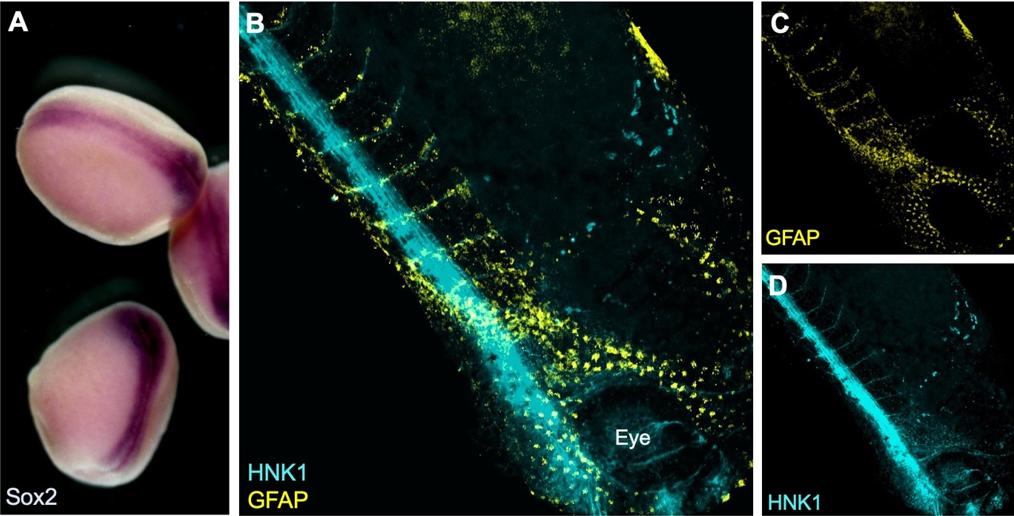
During the summer studentship I have learnt to work with Xenopus laevis embryos and keep track of their development. The aim of my project was to optimise techniques for visualising the neural crest with other tissues relevant to its induction, migration and differentiation. To do this I used two well established techniques in developmental biology: in situ hybridisation and immunofluorescence (see Figure 1).
Figure 1: A) Colorimetric in situ hybridisation with neural plate marker Sox2 in late neurula stage Xenopus embryos. B-D) Whole-mount fluorescent immunostaining for neuronal marker HNK1 and glial marker GFAP in a tailbud (Neiuwkoop and Faber stage 31) embryo.
Why study neural crest cells?
The neural crest is a fascinating population of cells unique to vertebrates, which is induced at the neural plate border and following neurulation delaminates and migrates away from the neural tube. A subset of these cells migrates into head regions where it gives rise to a variety of tissues including bones, cartilage, as well as neurons. This cranial neural crest also migrates into the branchial (or pharyngeal) arches and contributes in a major part to the craniofacial skeleton. This has undergone major changes during vertebrate evolution. For instance, some of the jaw bones present in our common ancestors with reptiles have given rise to the middle ear bones of mammals (Santagati & Rijli, 2003). A possible explanation for this versatility of the neural crest is that its cells retain multipotency for longer than the three embryonic germ layers, prompting some to consider it a ‘fourth germ layer’ (Simoes-Costa & Bronner, 2015).
The diversity of neural crest-derived cells, which also includes melanocytes, Schwann cells, meninges or the cornea, makes it an important model of differentiation with a potential for therapeutic applications. It is also a great model for studying cell migration (Szabó & Mayor, 2018) and by proxy epithelial to mesenchymal transition in cancer metastasis. Recent work even attributed the ability to remove cellular debris from early neural tube to migrating neural crest cells, potentially through a macrophage-like mechanism (Zhu, et al., 2019).
Going further with established techniques
I have been testing and optimising techniques to visualise proteins and gene expression that may help elucidate how the neural crest gets induced and how it migrates through the embryo. For example, I have image the neural crest with nearby mesoderm, which is required during induction, using a combination of fluorescent in situ hybridisation (FISH) and immunostaining. In order to visualise expression of two different genes within the embryo a double in situ hybridisation (ISH) can be used. This is especially important in Xenopus, as no antibody is known to efficiently and exclusively label the neural crest. Although colorimetric ISH is easier and does not require clearing of the tissues, it doesn’t enable exploring the 3D structure by imaging the whole embryo at once. I have used confocal and multiphoton microscopy to analyse the embryos (see Figure 2). Unfortunately, it seems that using two RNA probes at the same time reduces signal intensity and although I have tried to optimise the signal amplification reaction and bleaching to neutralise endogenous peroxidase activity, I have not been able to reduce the background.
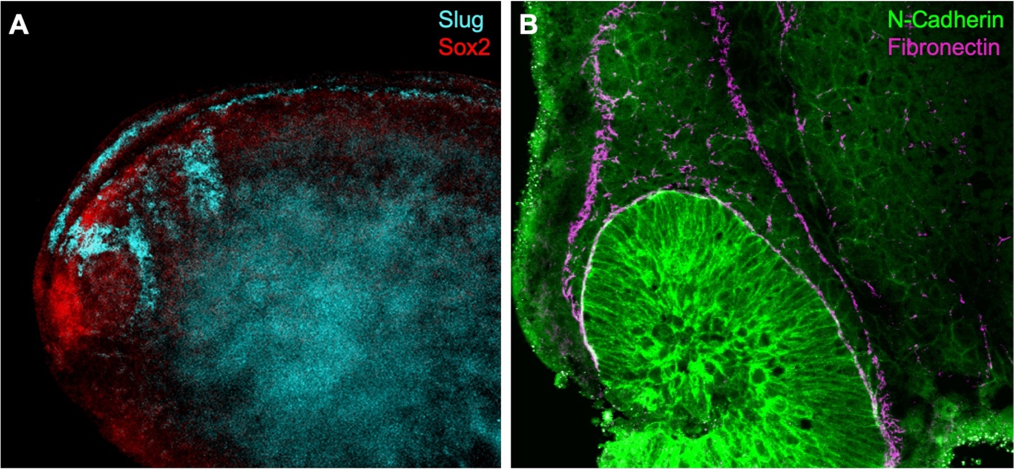
Figure 2: A) Fluorescent in situ hybridisation using RNA probes for Slug (neural crest) and Sox2 (neural plate) in a whole-mount stage 22 Xenopus embryo. B) Dissected embryo immunostaining shows N-Cadherin expression in migratory neural crest. Fibronectin outlines the streams of cranial neural crest.
One possibility is to delimit the neural crest using an antibody for fibronectin, an extracellular matrix component that encourages neural crest migration. Using double immunostaining (Figure 2B) I was able to confirm migratory neural crest cells express N-Cadherin near the optic vesicle as fibronectin outlines the neural crest streams (Scarpa, et al., 2015). Towards the end of my summer project, I have also manipulated cell contractility using drug treatments to observe changes in neural crest cell behaviour using several antibodies.
I thank Prof Roberto Mayor for supervising this summer project and Dr Adam Shellard for teaching me methods used in the lab. I have learnt a lot about how research is done and presented my results at a lab meeting in the final week and used this experience to transition from undergraduate study into my PhD.
References
Santagati, F. & Rijli, F. M., 2003. Cranial neural crest and the building of the vertebrate head. Nature Reviews Neuroscience, Issue 4, p. 806–818.
Scarpa, E. et al., 2015. Cadherin Switch during EMT in Neural Crest Cells Leads to Contact Inhibition of Locomotion via Repolarization of Forces. 34(4), pp. 421-434.
Simoes-Costa, M. & Bronner, M. E., 2015. Establishing neural crest identity: a gene regulatory recipe. Development, Volume 142, pp. 242-257.
Szabó, A. & Mayor, R., 2018. Mechanisms of Neural Crest Migration. Annual Review of Genetics , Volume 52, pp. 43-63.
Zhu, Y. et al., 2019. Migratory Neural Crest Cells Phagocytose Dead Cells in the Developing Nervous System. Cell, 179(1), pp. 74-89.
Established by the British Society for Developmental Biology in 2014, The Gurdon/The Company of Biologists Summer Studentship scheme provides financial support to allow highly motivated undergraduate students an opportunity to engage in practical research during their summer vacation. Each year, ten successful applicants spend eight weeks in the research laboratories of their choices, and the feedback we receive is outstanding. You can read accounts from previous years here. If you’re interested in applying or hosting a student in 2020, applications need to be in by the end of March.
Exploring the genetic control of microRNAs in Drosophila melanogaster
This summer, thanks to the BSDB Gurdon Studentship, I was able to work closely with a PhD student in the lab of Sarah Newbury at the Brighton and Sussex Medical School. Under their supervision I was able to get an applied, practical approach to lab work, much different from that experienced during my undergraduate course.
My project involved looking at the genetic control of microRNAs in Drosophila melanogaster, otherwise known as the common fruit fly. MicroRNAs are small-noncoding RNAs that participate in RNA silencing and regulation of gene expression. miRNAs are able to base-pair with their complementary target mRNA, and through this they are able to silence them, either by their subsequent degradation or prevention of their translation.
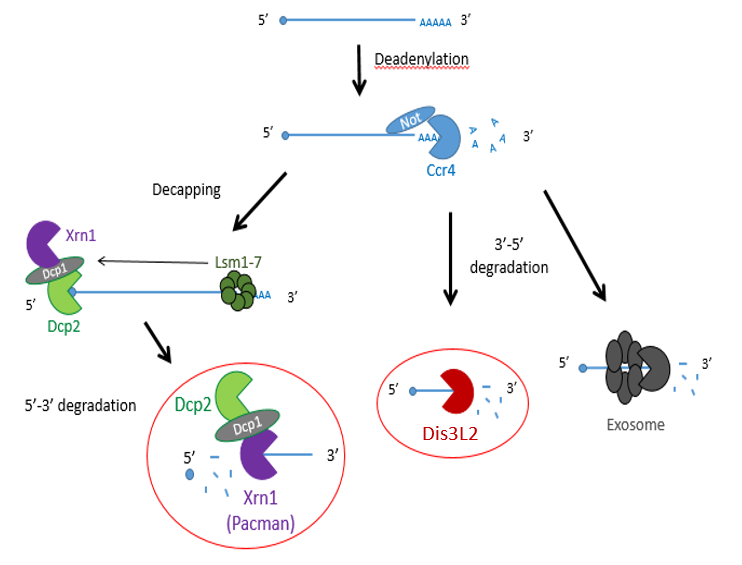
Previous studies have shown that exoribonucleases, known as Pacman (XRN1) and Dis3L2 degrade different microRNAs through the mRNA decay pathway, shown in Figure 1.
Figure 1 – The mechanisms of mRNA decay. Most mRNAs undergo decay through the deadenylation dependent pathway shown above. The poly(A) tail is first removed by a deadenylase (shown here as Ccr4-Not) and then proceeds down 3 possible routes: decapping by enzymes (shown as Dcp1/2) , followed by 5’ → 3’ decay by XRN1 (Pacman in Drosophila) or 3’ → 5’ decay by Dis3L2 or the exosome.
The project made use of wing imaginal discs, which are highly proliferative organs found in Drosophila that will eventually develop into the adult fly’s wings. Prior work in the lab had found that there were significant differences between the size of these discs in both pacman and dis3L2 mutants, with dis3L2 mutants having much larger discs (Towler et al. 2016), and pacman mutants having smaller ones (Waldron et al. 2015). Both exoribonucleases are conserved to humans and their defects have shown to be significant in human disease as well. For example, mutations of DIS3L2 in humans have been linked to Perlman syndrome, an overgrowth syndrome which presents as organomegaly and is associated to a high risk of developing Wilm’s tumours (Astuti et al. 2012). Together this suggested that there could be an involvement of these exoribonucleases in the control and regulation of cell proliferation and apoptosis within these discs, potentially through their degradation activity. Therefore, the aim of my project was to study the difference between levels of specific miRNAs (which were extracted from these WIDs) in both pacman and dis3L2 mutants, compared to their respective control wildtype.
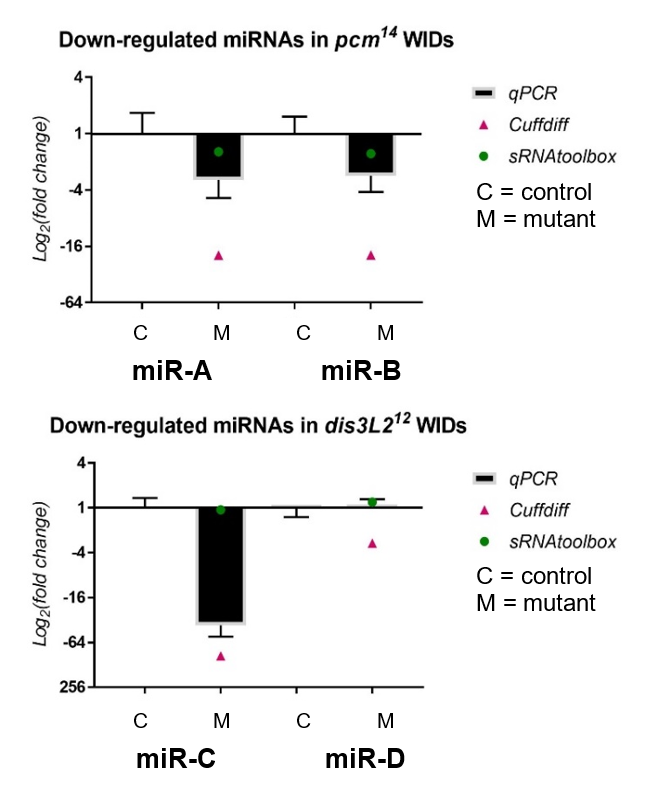
My results, shown in Figure 2, were obtained through the use of qPCR (quantitative or real-time PCR). Also shown on Figure 2 are two other datasets from previous high-throughput RNA sequencing experiments, known as Cuffdiff and sRNAtoolbox.
Figure 2 – The fold change of miRNAs downregulated in both dis3L2 and pacman (pcm) mutants normalized to their controls, obtained through my qPCR and previous RNA-seq methods.
Looking at my data, the pacman mutants showed that there was a negative fold change in the levels of two miRNAs, miR-A and miR-B when normalized to their wildtype’s levels. This indicated that the two miRNAs are downreguated in the pacman mutants. This confirmed the results obtained from the RNA sequencing, as they both showed that the miRNAs were downregulated as well.
Results from the dis3L2 mutants were less definite. For one of the miRNAs, miR-C, there was a negative fold change when normalized to the wildtype’s level. Again, this confirmed results obtained from the RNA-seq, but my data, and data from Cuffdiff, suggested a much stronger downregulation than that from sRNAtoolbox. For the other miRNA, miR-D, qPCR showed that there was no significant fold change in the mutants. This agrees with results from one of the RNA-seq, but not the other. Data from Cuffdiff showed a slight negative fold change of miR-D, suggesting it is downregulated in the mutants. However, my data, and that from sRNAtoolbox, suggested there is no change between the wildtypes and mutants.
Further areas of my project involved looking at the difference in volume of the abdomen and ovaries in flies with a defect in the gene of a TUTase known as Tailor, to see whether it was involved in controlling their size. TUTases, or Terminal Uridylyl Transferases, are enzymes that uridylate miRNAs, potentially serving as a signal for exoribonuclease-mediated degradation such as Dis3L2. This involved me freezing flies in liquid nitrogen, then photographing and dissecting out their ovaries. The limited number of ovaries measured suggested that there were no significant differences in the sizes between the wildtype and mutants.
In addition to this, I also looked at the differences in sizes of wings in Tailor mutant and double mutants of Tailor and Dis3L2 compared to wild types. This included dissecting wings and mounting them on a slide then taking measurements using computer software and a microscope. We hypothesised that the wings from the Tailor mutants would be the same size as the wildtype but the Tailor/Dis3L2 double mutants would have larger wings than the wildtype. My results here were too limited to draw a reliable conclusion but indicated there was no difference in wing sizes between the mutants and wildtype.
Overall, my time in the lab has proved highly valuable and interesting. Learning about the techniques in lectures and actually performing them yourself in a lab are completely different experiences and I am really grateful. I feel it was really important for me to experience the fly work alongside the molecular work as this has helped me to familiarize myself with numerous practises and therefore determine what is most interesting and suitable for me in the future. But not only did I learn how to perform these techniques, I was also introduced into the world of Drosophila, ranging from learning about their genome, recognising their phenotypes, and performing my own genetic crosses to learning their general upkeep and how to do simple things such as egglays or anesthetizing the flies. This experience for me has really increased my interest in developmental biology. Being able to produce my own data and learning to work independently in a lab have also strengthened my desire to do a PhD and I’m very appreciative that I got to experience something not many undergraduate students will. On top of all this, being inside a lab with such a friendly and welcoming group of people has made it all that more enjoyable.
A huge thank you to everyone in the lab for their support and assistance, and to the BSDB for making this opportunity possible through the Gurdon Studentship.
Sources
Towler BP, Jones CI, Harper KL, Waldron JA, Newbury SF. 2016. A novel role for the 3′-5′ exoribonuclease Dis3L2 in controlling cell proliferation and tissue growth. RNA biology 13: 1286-1299.
Waldron JA, Jones CI, Towler BP, Pashler AL, Grima DP, Hebbes S, Crossman SH, Zabolotskaya MV, Newbury SF. 2015. Xrn1/Pacman affects apoptosis and regulates expression of hid and reaper. Biology open 4: 649-660.
Astuti D, Morris MR, Cooper WN, Staals RH, Wake NC, Fews GA, Gill H, Gentle D, Shuib S, Ricketts CJ et al. 2012. Germline mutations in DIS3L2 cause the Perlman syndrome of overgrowth and Wilms tumor susceptibility. Nature genetics 44: 277-284.
The Koenig Lab at Harvard University is currently accepting applications to hire a Research Assistant II. Research in the lab will focus on the evolution and development of visual systems in the sea anemone Nematostella vectensis and the cephalopod Doryteuthis pealeii. The candidate will be pursuing questions using a combination of developmental and genetic approaches in the field and the laboratory.
Performs laboratory experiments using standard molecular biology techniques and advanced microscopy methods.
In collaboration with senior lab personnel, administers the implementation of a genetic screen in the lab and the general care of the Nematostella vectensis colony.
Processes, organizes, and summarizes data using scientific, word processing, statistical software applications, and the Adobe creative package.
Participates in weekly lab meetings and journal clubs.
Collects project data.
Assists in the design of laboratory experiments, techniques, and protocols involved in developmental biology and genetics.
May instruct and supervise other staff and students in basic laboratory techniques including standard molecular biology techniques, developmental techniques, and animal care.
Performs related lab duties such as maintaining and cleaning equipment, maintaining the Nematostella colony and assisting in the acquisition of squid, and ordering supplies.
The John Harvard Fellows Program and the surrounding community at Harvard provides a vibrant, interdisciplinary research environment with close links to the top academic and industrial institutions across the Boston area and provides the potential for your contributions to be used and recognized worldwide.
Basic Qualifications
A college background in molecular biology or evolutionary biology, or equivalent of education plus relevant work experience.
One or more years of related work experience with embryos and molecular developmental techniques. Demonstrated abilities in laboratory techniques are required.
Additional Qualifications
A college degree (BS/BA) in molecular biology or evolutionary biology is preferred.
Must be able to work independently but also work in close collaboration with other members of the lab
Must be detailed oriented
Exposure to molecular biology and Gibson assembly protocols preferred.
Exposure to embryo injections, transgenesis and CRISPR Cas9 protocols preferred.
Exposure to applicable computer languages including Python, R and Unix is preferred.
Experience working in ImageJ, Photoshop and Illustrator preferred.
Exposure to confocal microscopy is preferred.
May be required to lift lab equipment and seawater up to 50 lbs
Must be willing and able to work with animals and their embryos
We invite individuals with diverse backgrounds, experiences, and abilities to be a part of our community.
Established by the British Society for Developmental Biology in 2014, The Gurdon/The Company of Biologists Summer Studentship scheme provides financial support to allow highly motivated undergraduate students an opportunity to engage in practical research during their summer vacation. Each year, ten successful applicants spend eight weeks in the research laboratories of their choices, and the feedback we receive is outstanding. You can read accounts from previous years here. If you’re interested in applying or hosting a student in 2020, applications need to be in by the end of March.
How to get radial: Unlocking the mechanisms for symmetry establishment across plant organs
The development of multicellular organisms depends on correct establishment of symmetry both at the whole-body scale and within individual tissues and organs. Setting up planes of symmetry must rely on communication between distant cells within the organism, presumably via mobile morphogenic signals. Although symmetry in nature has fascinated scientists for centuries, it is only now that molecular data to unravel mechanisms for symmetry establishment are beginning to emerge.
Over the summer, I had the opportunity of conducting a research project at the John Innes Centre (JIC) working under the supervision of Dr Laila Moubayidin, whilst participating in the amazing JIC/TSL/EI International Undergraduate Summer School.
Dr Moubayidin’s research looks at identifying a conserved “core machinery” necessary and sufficient to control symmetry establishment across plant organs. She is elucidating the underlying process of symmetry establishment at the cellullar, molecular and genetic level.
Unlocking the molecular mechanisms that underpin this regulation holds potential for understanding the processes that allow plant organs to reach their perfectly-optimized shape and function.
The project I worked on focused on gynoecium (the plant female reproductive structure) development, using Arabidopsis thaliana as a model system.
The gynoecium forms in the centre of the flower and it is derived from the fusion of two carpels. The gynoecium includes the ovary, which displays bilateral symmetry, and the style, at its apex, characterized by radial symmetry (Figure 1). The ovary is important for seeds production and the style is important for fertilization.
Figure 1: Scanning Electron Microscopy picture of A.thaliana WT gynoecium. Red arrow indicates the style, radially symmetric; white arrow indicates the ovary, bilaterally symmetric.
During its development, the distal end of the gynoecium becomes radially symmetric via a switch from bilateral to radial symmetry. Symmetry transitions are common during embryogenesis in all multicellular organisms. In most cases, the transition is from radial to bilateral symmetry, which is controlled by Hox and decapentaplegic genes in animals. The A.thaliana gynoecium, instead, is the only molecularly documented example of a developing structure that reprograms its development over time to achieve a bilateral-to-radial symmetry transition.
Growing evidence shows that coherent organ growth and symmetry establishment during organogenesis are influenced by post-translational modifications of specific nuclear and cytoplasmatic proteins. My project focused on the role of O-linked N-acetylglucosamine modification during gynoecium development. O-GlcNAcylation is a post-translational modification that consists of a single O-linked N-acetylglucosamine attached to either a serine or a threonine residue. It has been extensively studied in animals, where it regulates a wide range of developmental and metabolic processes.
O-GlcNAcylation is dynamically controlled by two enzymes: an O-GlcNAc transferase (OGT) and an O-GlcNAcase (OGA), which add and remove O-GlcNAc, respectively. O-GlcNAcylation occurs in the cytoplasm, nucleus and mithocondria, and it is implicated in cellular processes, including transcription, translation, signal transduction, nuclear pore function, epigenetic regulation and proteosomal degradation. Altered levels of protein O-GlcNAcylation in animals have been associated with neurodegeneration, diabetes, cardiovascular diseases and cancer.
Arabidopsis thaliana has two putative OGTs: SPINDLY (SPY) and SECRET AGENT (SEC) whose role is essential for plant development, since spy;sec double mutant is embryionically lethal, similar to the OGT knockout mutant in animals.
Exploring this interesting and at the same time quite complicated topic, gave me the chance of learning and mastering a wide range of molecular biology techniques, such as cloning and Yeast-two-Hybrid. It was amazing to experience practical methods and protocols I had studied during university courses in Rome and to understand how much work, attention to detail and care there is behind even repetitive actions (Figure 2).
Figure 2: Working with transformed yeast plates in very sterile conditions
Using the state-of-the-art JIC microscopy facilities, I had the opportunity to look at wild-type and mutant Arabidopsis gynoecia at different developmental stages using Scanning Electron Mycroscopy (SEM). Fixing samples for SEM analysis, dissecting flowers and using machines such as the Critical Point Dryer and the Gold Sputter Coater was a challenging experience, but the final result was worth the hard work. I was astonished whilst I was conducting SEM analysis as it allows observation of samples at incredible magnifications, providing details of a single cell surface (Figure 3).
Figure 3: Observation of gynoecia at the Scanning Electron Microscope
At the end of this experience, I felt extremely grateful for several reasons.
Firstly, I greatly enhanced my knowledge of plant developmental biology. As a “Sapienza School for Advanced Studies” student, in the first year of university, I was keen to work on plants during my Research Project, so I applied to the International Undergraduate Summer School at the JIC. After being selected, I was urged by Dr Moubayidin to apply for a Gurdon/The Company of Biologists Summer Studentship offered by the British Society for Developmental Biology (BSDB). Moreover, working side-by-side an experienced, young principal investigator, such as Dr Moubayidin, has been a rare opportunity and a great inspiration for my future career. Dr Moubayidin has been recently appointed a prestigious Royal Society University Research Fellowship, therefore it has been an exciting time to be part of her lab, experiencing how a research group is built and the importance of teamwork.
Lastly, this summer placement has given me great insight into research career paths and has made me even more aware of how incredibly surprising and stimulating daily lab life can be. Each workshop organized by the JIC Summer School and every seminar held at the JIC has been a great chance for me to learn about new topics, uncovering the research world of an international centre and the beauty of working in a collaborative and stimulating atmosphere.
Altogether, this unexpected chain of events has lead me to immensely broaden my scientific and transferable skills, as well as my perspectives, not only as a scientist, but also as a “world citizen”. This Summer School experience, set in the incredibly positive environment of the JIC, has reaffirmed my desire of pursuing a career in science, despite all the hard work it actually requires. I wholeheartedly recommend the Gurdon/BSDB Summer Studentship and the possibility to carry on research abroad during university studies, visiting new countries and encountering new cultures. I believe every undergraduate student should be made aware of unique opportunities, like this, available to them.
References
Moubayidin L., Østergaard L., Dynamic control of auxin distribution imposes a bilateral-to-radial symmetry switch during gynoecium development, Current Biology 24, 2743-2748, (2014).
Moubayidin L., Østergaard L., Symmetry matters, New Phytologist (2015).
Shou-Ling Xu, Robert J. Chalkley, Jason C. Maynard, Wenfei Wang, Weimin Ni, Xiaoyue Jiang, Kihye Shin, Ling Cheng, Dasha Savage, Andreas F. R. Hühmer, Alma L. Burlingame, and Zhi-Yong Wang, Proteomic analysis reveals O-GlcNAc modification on proteins with key regulatory functions in Arabidopsis, PNAS 114 (8) E1536-E1543, (2017)
Established by the British Society for Developmental Biology in 2014, The Gurdon/The Company of Biologists Summer Studentship scheme provides financial support to allow highly motivated undergraduate students an opportunity to engage in practical research during their summer vacation. Each year, ten successful applicants spend eight weeks in the research laboratories of their choices, and the feedback we receive is outstanding. You can read accounts from previous years here. If you’re interested in applying or hosting a student in 2020, applications need to be in by the end of March.
Our third report from the class of 2019 comes from Isabel Swinburn (University of Birmingham) who studied zebrafish muscle development with Robert Knight’s lab at King’s College London.
Throughout my Biological Sciences degree, I came to realise that I want to pursue a career in medicine that combines clinical practice with research. After having thoroughly enjoyed my final year research project that focused on bacterial genetics, I decided to apply for a studentship in a biomedical research lab. This was with the hope that I’d be able to apply some of the experimental techniques I’d already learnt to understand complex processes in living organisms. Having a good grounding in organismal biology will be useful when applying a research perspective to human systems.
I was very fortunate to complete my Gurdon/The Company of Biologists Summer Studentship under the supervision of Dr. Robert Knight at King’s College London. His group uses zebrafish as a model to explore the molecular control of muscle repair and the regulation of muscle stem cells, otherwise known as satellite cells. These cells are characterised by their expression of the pax7b transcription factor (1). The zebrafish is a suitable model organism for their investigation, as it has a well-defined genome and is transparent in its larval stages. Therefore, genetic manipulations that affect cell and tissue dynamics can be readily visualised. Their work particularly appealed to me, as I am very interested in the use of reverse genetics approaches to investigate genetic diseases and how they arise due to the disruption of development.
Background to project:
The transmembrane Ret tyrosine kinase receptor is activated by the binding of Glial-derived neurotrophic factor family ligands (GFL) to Glial-derived neurotrophic factor receptors (Gfra), as this interaction stimulates Ret dimerisation and subsequently, its activation. Ret signalling has an important role in the development of the enteric nervous system and hepatic tissue, but its role in muscle development is poorly understood. Ret signalling has been implicated in facioscapulohumeral muscular dystrophy, which manifests as the weakening of the facial muscles in its initial stages (2). The binding of artemin2 (artn2), a GFL, to Grfa3 is an interaction required for the activation of Ret related to the development of the cranial muscles in zebrafish (3). My project set out to investigate the role and level of importance of Ret signalling in the development of cranial muscle satellite cells.
Experiments
1) Is an activating ligand for the Ret receptor able to alter cranial muscle development?
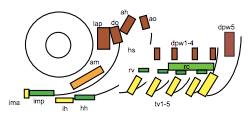
To test this hypothesis, I performed MF20 immunolabelling on zebrafish embryos exhibiting heat-shock inducible artn2 overexpression, as well as controls. Diaminobenzidine (DAB) staining was used to detect the signal from the antibodies, which made myofibres appear orange-brown (Figure 2a). The embryos were fixed and imaged using bright-field microscopy. The sizes of a subgroup of cranial muscles were measured and compared between the experimental and control samples (Figure 1). There did not appear to be a significant difference in muscle size between the two sample groups, suggesting that the activating ligand for the Ret receptor cannot alter cranial muscle development.
Figure 1: Schematic left-side view of the zebrafish cranial muscles. The muscles of interest in my investigations were the lap (levator arcus palatine), do (dilator opercula), ah (adductor hyoideus) and ao (adductor opercula). These muscles were selected due to their flat shape, making them easy to analyse (4).
2) Is an activating ligand for the Ret receptor able to alter cranial muscle satellite cell formation?
To test this hypothesis, a transgenic line was used, in which pax7b-expressing cells were labelled with EGFP and artn2 was overexpressed. Embryos with this genotype, along with GFP+ embryos with the wild type artn2 genotype, were fixed and imaged using confocal microscopy. The number of GFP+ myofibres and myoblasts within the muscles of interest were quantified and compared between the experimental and control samples. I was unable to discriminate a significant difference, due to the control sample size being too small to be able to perform statistical tests. Therefore, more wild-type embryos need to be imaged and analysed to confirm whether artn2 can alter cranial muscle satellite cell formation.
3) How important are satellite cells for cranial muscle development?
To answer this question, I ablated pax7b-expressing cells during development. A GAL4;UAS system was used, in which the pax7b promoter induced expression of nitroreductase (NTR). Embryos which carried this system were treated with metronidazole, which reacts with NTR to produce a cytotoxic compound. The muscles of these embryos, as well as controls, underwent MF20 immunolabelling and these signals were detected using either DAB or tyramide FITC, to which a green fluorophore is conjugated. Depending on the detection method, the embryos were imaged using bright-field or confocal microscopy, respectively, and the myofibres in each image were quantified (Figure 2). Analysis of the bright-field images suggested a difference in the number of myofibres within the cranial muscles of the experimental and control sample groups. However, quantification analysis of the confocal images did not reveal a significant difference. Therefore, further experiments must be carried out to confirm the importance of satellite cells for cranial muscle development.
Figure 2: MF20 immunostaining of zebrafish embryos after the ablation of their pax7b-expressing cells during development. a) Confocal image of an embryo in which the MF20 signal was detected with tyramide FITC. b) Bright-field image of an embryo in which the MF20 signal was detected with DAB
Outcomes:
At university, I had only used optical microscopy in practical sessions. However, following my time in the Knight lab, I now feel confident in using a much wider range of more advanced microscopy techniques to gather experimental data. I also appreciate much more the importance of analysing the data – quantifying cells and myofibres within images seemed like a simple task, but it took me weeks! Unlike at university, the data I collected whilst working in the lab was raw and unclean and strict parameters had to be set for its analysis. Although the task was daunting and sometimes difficult, it was very gratifying when the numbers and statistical test results allowed me to test the hypotheses generated and realise that I had made a scientific discovery.
I would like to say thank you to Robert for allowing me to work under his supervision, and to the British Society for Developmental Biology for providing the financial backing to enable me to do so. A special thank you also goes to the rest of the Knight group for all of their support and encouragement throughout. Despite my focus being primarily on a career in medicine, this experience has reinforced my desire to continue to contribute to the type of evidence that will drive medical practice. I would recommend anyone considering a career in biological research to apply for this programme.
References
Pipalia, T. G. et al., 2016. Cellular dynamics of regeneration reveals role of two distinct Pax7b stem cell populations in larval zebrafish muscle repair. Diseases Models & Mechanisms, 9(6), pp. 671-684.
Moyle, L. A. et al., 2016. Ret function in muscle stem cells points to tyrosine kinase inhibitor therapy for facioscapulohumeral muscular dystrophy. eLife, e11405.
Knight, R. D. et al., 2011. Ret signalling integrates a craniofacial muscle module during development. Development, 138(10), pp. 2105-2024.
Schilling, T. F. & Kimmel, C.B., 1997. Musculoskeletal patterning in the pharyngeal segments of the zebrafish embryo. Development, Volume 124, pp. 2945-2960.
The Department of Systemic Cell Biology (Prof. Dr. Bastiaens) at the Max Planck Institute for Molecular Physiology in Dortmund is offering a postdoctoral position (developmental biology/cell biology) to work in an interdisciplinary team of cell biologists and physicists studying mechanisms of tumorigenesis in intestinal organoids.
We are investigating fundamental mechanisms of oncoprotein induced tumor development by combining chemically induced optogenetic tools with advanced molecular imaging approaches that report on the dynamic states of signalling networks in developing multi-cellular assemblies.
We are looking for an enthusiastic, inquisitive and creative individual with a degree in life sciences and a PhD in cell or developmental biology and a strong background in quantitative microscopy to join our team that is interested in understanding how bidirectional cellular communication impacts cell fates in three-dimensional cellular assemblies.
The position is available immediately and is limited to 2 years with the possibility of prolongation. Financial compensation will be carried out according to the public service remuneration scheme TVöD.
The Max-Planck-Society is committed to increasing the number of individuals with disabilities in its workforce and therefore encourages applications from such qualified individuals.
The Max-Planck-Society strives for gender equality and diversity within its workforce.
Furthermore, the Max Planck Society seeks to increase the number of women in those areas where they are underrepresented and therefore explicitly encourages women to apply.
Applications should be sent to the hands of the scientific coordinator of the department, Dr. Astrid Krämer, email: astrid.kraemer[at]mpi-dortmund.mpg.de in digital form (e.g. as PDF). In your application, include a cover letter, recent CV, and copies of your certificates and two names of reference. Please submit all documents in a single application until February 9th, 2020.
What happens when an innocent genetic test reveals hidden secrets?
It’s been impossible to ignore the rise in direct-to-consumer and medical genetic testing over the past few years. And as the cost of whole genome sequencing falls – and the potential personal, health and financial value of genomic data rises – this trend is only likely to continue.
But do people really realise what they’re signing up for when they spit into a tube or squirt out a blood sample?
As we head into the next decade, ethical issues like informed consent and privacy for genomic testing and research are becoming impossible to ignore – especially as your genetic information doesn’t just belong to you but is also shared with your blood relatives.
When Jack Nunn started doing a PhD at La Trobe University in Melbourne, Australia, looking at how to involve people in genomics research, the most obvious place to start was with his own family. But he could never have predicted the secret that would be revealed once they started looking into their genes.
We also speak with ethics research and genetic counsellor Anna Middleton, to find out more about the ethical issues around the fast-changing field of consumer and medical genomics.
If you enjoy the show, please do rate and review on Apple podcasts and help to spread the word on social media. And you can always send feedback and suggestions for future episodes and guests to podcast@geneticsunzipped.com Follow us on Twitter – @geneticsunzip
A 4-year PhD position is available in the lab of Patrick Tschopp at the University of Basel, Switzerland, to study the ontogenetic basis and developmental plasticity of dietary adaptations in vertebrates, with a focus on African cichlid fishes. This project is part of a Sinergia grant by the Swiss National Science Foundation (SNF) to H. Kaessmann (UHeidelberg), M. Clauss (UZurich), P. Tschopp (UBasel) & W. Salzburger (UBasel).
We are looking for a highly motivated candidate with strong interests in developmental biology, single-cell functional genomics and bioinformatic analyses, as well as experimental work with fish. We offer a highly interactive, stimulating and interdisciplinary research environment, state-of-the-art research infrastructure, and a competitive salary.
The University of Basel (www.unibas.ch) is the oldest university in Switzerland, located in one of Europe’s most important life science hubs at the border between Switzerland, France and Germany. The Tschopp lab (www.evolution.unibas.ch/tschopp/research/) studies the gene regulatory mechanisms of cell type specification and evolution in vertebrates.
Applications should include a motivation letter, a CV, a list of publications, a statement about research interests, as well as the names and contact details of at least two referees. Applications (in the form of a single .pdf file) should be sent to Patrick Tschopp (patrick.tschopp@unibas.ch); the deadline is February 7th, 2020.
A 4-years PostDoc position is available in the labs of Patrick Tschopp and Walter Salzburger at the University of Basel, Switzerland, to study the molecular and ontogenetic basis of dietary adaptations in vertebrates at the macro- and micro-evolutionary scale. This project is part of a Sinergia grant by the Swiss National Science Foundation (SNF) to H. Kaessmann (UHeidelberg), M. Clauss (UZurich), P. Tschopp (UBasel) & W. Salzburger (UBasel).
Through a timely combination of state-of-the-art genomics technologies and a broad phylogenetic sampling, we will study the molecular and developmental underpinnings of dietary adaptations in vertebrates.
We are looking for a highly motivated young scientist, who holds a PhD in biology or related fields, and has a strong background in bioinformatics, single cell/transcriptome/genome analyses, as well as in evolutionary and/or developmental biology. She/he should have experience in supervising and project management. We offer a highly interactive, stimulating and interdisciplinary research environment, state-of-the-art technology platforms and research infrastructure, attractive employment conditions with a competitive salary, and career development opportunities.
The University of Basel (www.unibas.ch) is the oldest university in Switzerland, located in one of Europe’s most important life science hubs at the border between Switzerland, France and Germany. The Tschopp lab (www.evolution.unibas.ch/tschopp/research/) studies the gene regulatory mechanisms of cell type specification and evolution in vertebrates. The lab of Walter Salzburger (www.salzburgerlab.org) studies the dynamics of adaptation and organismal diversification in cichlids from African Lake Tanganyika and other exceptional groups of fishes.
Applications should include a motivation letter, a CV, a list of publications, a statement about research interests, as well as the names and contact details of at least two referees. Applications (in the form of a single .pdf file) should be sent to Patrick Tschopp (patrick.tschopp@unibas.ch) and Walter Salzburger (walter.salzburger@unibas.ch) until February 7th, 2020.
 (No Ratings Yet)
(No Ratings Yet)
 (5 votes)
(5 votes)