Journal of Cell Science and its publisher, The Company of Biologists, are seeking to appoint a Community Manager, based in Cambridge, UK, to run a new microscopy resource website.
An extensive consultation told us that the microscopy community would welcome a trusted, curated and centralised site to connect people, resources and information. Our exciting new initiative is therefore intended to create a platform for the microscopy community to share news and techniques, discuss issues relevant to the field and read about the latest research and events.
We are now looking for an enthusiastic and motivated person to join us to develop, launch and maintain this site, which will be hosted by Journal of Cell Science but will be relevant to all of the Company of Biologists’ journals.
Core responsibilities of the position include:
Creating and commissioning content, including writing posts and soliciting content from the academic community, societies, companies and other organisations.
Providing user support and ensuring site functionality on a day-to-day basis.
Providing creative and practical input into the design and development of the site.
Maintaining and developing the site’s presence on social networking sites such as Facebook and Twitter.
Developing and managing sponsorship, ads and commercial relationships.
Representing Journal of Cell Science and the resource site at international conferences.
Applicants should have a PhD with research experience in a relevant scientific field and extensive imaging experience – ideally involving the application or development of new methods for microscopy or image analysis. The successful candidate will have proven social media skills and a clear understanding of the online environment as it applies to scientists. Applicants should have excellent writing and communication skills, and strong interpersonal and networking abilities – both online and in person. Experience with additional media, such as video or podcasting, and an eye for aesthetics, would be an advantage. We are looking for an individual with fresh ideas and a willingness to learn new skills, and who will contribute broadly to the Company’s activities.
This is an exciting opportunity to develop a hub for the microscopy community – in a similar vein to the Company’s established community site for developmental biologists, the Node – and to engage with relevant people at all levels: academics, developers, facilities, institutes and companies. The Community Manager will work alongside an experienced in-house team, including the Executive Editor of Journal of Cell Science, as well as with the journal’s international team of academic editors. Additional responsibilities may be provided for the right candidate. The Company of Biologists is based in attractive modern offices on the outskirts of Cambridge, UK.
The Company of Biologists (biologists.com) exists to support biologists and inspire advances in biology. At the heart of what we do are our five specialist journals – Development, Journal of Cell Science, Journal of Experimental Biology, Disease Models & Mechanisms and Biology Open – two of them fully open access. All are edited by expert researchers in the field, and all articles are subjected to rigorous peer review. We take great pride in the experience of our editorial team and the quality of the work we publish. We believe that the profits from publishing the hard work of biologists should support scientific discovery and help develop future scientists. Our grants help support societies, meetings and individuals. Our workshops and meetings give the opportunity to network and collaborate.
Applicants should send a CV along with a covering letter that summarises their relevant experience, and in particular their specific microscopy/imaging expertise, any links to online activities), current salary, and why they are enthusiastic about this opportunity.
Applications and informal queries should be sent by email to hr@biologists.com.
We may request written tests in advance of any interview.
Job Title: Post-doctoral fellow. Position is currently recruiting. Minimum of 2-year commitment is required.
Job Description
The Liao Laboratory at the Center for Regenerative Medicine at Massachusetts General Hospital in affiliation with the Division of Plastic Surgery and the Harvard Stem Cell Institute is seeking a highly motivated research fellow interested in craniofacial developmental biology and genetics. We are currently recruiting 2 post-doctoral fellows to join our team of 10 investigators. One post-doc position is seeking a candidate with experience in iPSC derivation and cell model work. The second post-doc position is seeking a candidate with experience in zebrafish or mouse models. We have a pipeline of studying human subjects with rare craniofacial conditions, where we apply human iPSC, mouse or zebrafish models to dissect the developmental genetic basis of craniofacial malformations.
The fellow will join a collaborative group of scientists who study the basic and translational biology of facial morphogenesis, cranial neural crest specification and development. In this instance, the fellow’s work will have a direct bearing on discovery of drugs to mitigate malformation phenotypes and functional genomics of human candidate genes implicated in orofacial clefts. We are inherently interested in the basic biology of neural crest cell specification, migration, differentiation and potential for regeneration. Integrated approaches of classical developmental and modern stem cell biology are used to explore human tissue regeneration and disease. We apply the latest technology in zebrafish genetics with routine use of CRISPR, iPSC, single cell RNAseq, Cut&Run transcriptional analysis, cell transplants, FACS, live confocal labeling and imaging, transgenics, chemical screening, and many other experimental approaches. Our laboratory has a dedicated zebrafish facility outfitted with the latest imaging, injection and cell manipulation instruments. Researchers work in open format laboratory space that foster interactions with other stellar researchers working in the Center, with ample opportunities to participate in scientific seminars and professional development.
The current lab members include 3 post-doc fellows, 3 graduate students, 3 technicians, 1 undergraduate and 1 high school student. The laboratory has stable funding with 4 extramural grants. There will be ample opportunity for eligible candidate to compete for independent funding.
Job Requirements
The candidate should have PhD or an MD/PhD in a relevant field or equivalent training. A strong experimental foundation in iPSC derivation, iPSC analysis, cell culture, animal models are mandatory. Additional experience with mouse genetics, development, cell, developmental, are desirable. An ancillary knowledge of genomics or systems biology is welcome. Outstanding oral and written communication skills are required.
Please email your inquiry, current CV, statement of interest and career goals, to Eric Liao, cliao@partners.org
Employing Department and Institution:Center for Regenerative Medicine, Massachusetts General Hospital.
Welcome to our monthly trawl for developmental biology (and related) preprints.
This month was notable for a preponderance of plant development preprints, many molecular maps (supported by single cell sequencing), a hearty helping of human development and a multiplicity of (Drosophila) melanogaster mechanics.
The preprints were hosted on bioRxiv, PeerJ, andarXiv. Let us know if we missed anything, and use these links to get to the section you want:
Osteocytes remodel bone by TGF-β-induced YAP/TAZ signaling
Christopher D. Kegelman, Jennifer C. Coulombe, Kelsey M. Jordan, Daniel J. Horan, Ling Qin, Alexander G. Robling, Virginia. L Ferguson, Teresita M. Bellido, Joel D. Boerckel
Early human embryos from Meistermann, et al.’s preprint
Spatio-temporal analysis of human preimplantation development reveals dynamics of epiblast and trophectoderm
Dimitri Meistermann, Sophie Loubersac, Arnaud Reignier, Julie Firmin, Valentin Francois Campion, Stéphanie Kilens, Yohann Lelièvre, Jenna Lammers, Magalie Feyeux, Phillipe Hulin, Steven Nedellec, Betty Bretin, Simon Covin, Gael Castel, Audrey Bihouée, Magali Soumillon, Tarjei Mikkelsen, Paul Barrière, Jérémie Bourdon, Thomas Fréour, Laurent David
Control of glioblastoma differentiated-to-stem cell reprogramming by IRE1α/XBP1s signaling.
Dimitrios Doultsinos, Mari McMahon, Konstantinos Voutetakis, Joanna Obacz, Raphael Pineau, Florence Jouan, Pierre-Jean Le Reste, Akram Obiedat, Juhi Samal, John B Patterson, Quinping Zheng, Afshin Samali, Abhay Pandit, Boaz Tirosh, Aristotelis Chatziioannou, Eric Chevet, Tony Avril
Redox potential defines functional states of adult hippocampal stem cells
Vijay S Adusumilli, Tara L Walker, Rupert W Overall, Gesa M Klatt, Salma A Zeidan, Tim J Fischer, Sara Zocher, Alex M Sykes, Susanne Reinhardt, Andreas Dahl, Dilyana G Kirova, Jörg Mansfeld, Annette E Rünker, Gerd Kempermann
ELIMÄKI locus is required for mechanosensing and proprioception in birch trees
Juan Alonso-Serra, Xueping Shi, Alexis Peaucelle, Pasi Rastas, Matthieu Bourdon, Juha Immanen, Junko Takahashi, Hanna Koivula, Gugan Eswaran, Sampo Muranen, Hanna Help-Rinta-Rahko, Olli-Pekka Smolander, Chang Su, Omid Safronov, Lorenz Gerber, Jarkko Salojärvi, Risto Hagqvist, Ari-Pekka Mähonen, Kaisa Nieminen, Ykä Helariutta
Brassica fruits from Stephenson, et al.’s preprint
Mapping and dynamics of regulatory DNA in maturing seeds
Alessandra M Sullivan, Andrej A Arsovski, Agnieszka Thompson, Richard Sandstrom, Robert E Thurman, Shane Neph, Audra K Johnson, Shawn T Sullivan, Peter J Sabo, Fidencio V Neri III, Molly Weaver, Morgan Diegel, Jennifer L Nemhauser, John A Stamatoyannopoulos, Kerry L Bubb, Christine Queitsch
Fog signaling has diverse roles in epithelial morphogenesis in insects
Nadine Frey, Matthew A. Benton, Rodrigo Nunes da Fonseca, Cornelia von Levetzow, Dominik Stappert, Muhammad Salim Hakeemi, Kai H. Conrads, Matthias Pechmann, Kristen A. Panfilio, Jeremy A. Lynch, Siegfried Roth
Two Y chromosome-encoded genes determine sex in kiwifruit
Takashi Akagi, Sarah M. Pilkington, Erika Varkonyi-Gasic, Isabelle M. Henry, Shigeo S. Sugano, Minori Sonoda, Alana Firl, Mark A. McNeilage, Mikaela J. Douglas, Tianchi Wang, Ria Rebstock, Charlotte Voogd, Paul Datson, Andrew C. Allan, Kenji Beppu, Ikuo Kataoka, Ryutaro Tao
Enabling large-scale genome editing by reducing DNA nicking
Cory J. Smith, Oscar Castanon, Khaled Said, Verena Volf, Parastoo Khoshakhlagh, Amanda Hornick, Raphael Ferreira, Chun-Ting Wu, Marc Güell, Shilpa Garg, Hannu Myllykallio, George M. Church
A simple and efficient CRISPR technique for protein tagging
Fanning Zeng, Valerie Beck, Sven Schuierer, Isabelle Garnier, Carole Manneville, Claudia Agarinis, Lapo Morelli, Lisa Quinn, Judith Knehr, Guglielmo Roma, Frederic Bassilana, Mark Nash
A Pandas complex adapted for piRNA-guided transposon silencing
Kang Zhao, Sha Cheng, Na Miao, Ping Xu, Xiaohua Lu, Ming Wang, Yuhan Zhang, Xun Yuan, Weiwei Liu, Xin Lu, Xuan Ouyang, Peng Zhou, Jiaqi Gu, Yiqun Zhang, Ding Qiu, Shan Wang, Zhaohui Jin, Youzhong Wan, Jinbiao Ma, Ying Huang, Yang Yu
A reference map of the human protein interactome
Katja Luck, Dae-Kyum Kim, Luke Lambourne, Kerstin Spirohn, Bridget E. Begg, Wenting Bian, Ruth Brignall, Tiziana Cafarelli, Francisco J. Campos-Laborie, Benoit Charloteaux, Dongsic Choi, Atina G. Cote, Meaghan Daley, Steven Deimling, Alice Desbuleux, Amélie Dricot, Marinella Gebbia, Madeleine F. Hardy, Nishka Kishore, Jennifer J. Knapp, István A. Kovács, Irma Lemmens, Miles W. Mee, Joseph C. Mellor, Carl Pollis, Carles Pons, Aaron D. Richardson, Sadie Schlabach, Bridget Teeking, Anupama Yadav, Mariana Babor, Dawit Balcha, Omer Basha, Christian Bowman-Colin, Suet-Feung Chin, Soon Gang Choi, Claudia Colabella, Georges Coppin, Cassandra D’Amata, David De Ridder, Steffi De Rouck, Miquel Duran-Frigola, Hanane Ennajdaoui, Florian Goebels, Liana Goehring, Anjali Gopal, Ghazal Haddad, Elodie Hatchi, Mohamed Helmy, Yves Jacob, Yoseph Kassa, Serena Landini, Roujia Li, Natascha van Lieshout, Andrew MacWilliams, Dylan Markey, Joseph N. Paulson, Sudharshan Rangarajan, John Rasla, Ashyad Rayhan, Thomas Rolland, Adriana San-Miguel, Yun Shen, Dayag Sheykhkarimli, Gloria M. Sheynkman, Eyal Simonovsky, Murat Taşan, Alexander Tejeda, Jean-Claude Twizere, Yang Wang, Robert J. Weatheritt, Jochen Weile, Yu Xia, Xinping Yang, Esti Yeger-Lotem, Quan Zhong, Patrick Aloy, Gary D. Bader, Javier De Las Rivas, Suzanne Gaudet, Tong Hao, Janusz Rak, Jan Tavernier, Vincent Tropepe, David E. Hill, Marc Vidal, Frederick P. Roth, Michael A. Calderwood
Dendritic spines from Chakraborty, et al.’s preprint
Light-sheet microscopy with isotropic, sub-micron resolution and solvent-independent large-scale imaging
Tonmoy Chakraborty, Meghan Driscoll, Malea Murphy, Philippe Roudot, Bo-Jui Chang, Saumya Vora, Wen Mai Wong, Cara Nielson, Hua Zhang, Vladimir Zhemkov, Chitkale Hiremath, Estanislao Daniel De La Cruz, Ilya Bezprozvanny, Hu Zhao, Raju Tomer, Rainer Heintzmann, Julian Meeks, Denise Marciano, Sean Morrison, Gaudenz Danuser, Kevin M. Dean, Reto Fiolka
3D Multicolor Nanoscopy at 10,000 Cells a Day
Andrew E S Barentine, Yu Lin, Miao Liu, Phylicia Kidd, Leonhard Balduf, Michael R Grace, Siyuan Wang, Joerg Bewersdorf, David Baddeley
The ELIXIR Core Data Resources: fundamental infrastructure for the life sciences
Rachel Drysdale, Charles E Cook, Robert Petryszak, Vivienne Baillie Gerritsen, Mary Barlow, Elisabeth Gasteiger, Franziska Gruhl, Juergen Haas, Jeremy Lanfear, Rodrigo Lopez, Nicole Redaschi, Heinz Stockinger, Daniel Teixeira, Aravind Venkatesan, ELIXIR Core Data Resource Forum, Niklas Blomberg, Christine Durinx, Jo McEntyre
We are seeking an outstanding colleague to join collaborative research and provide bioinformatics expertise. Candidate will collaborate with members of the Center of Regenerative Medicine to analyze and interpret primary sequencing data (ChIP-seq, RNA-seq, genome/exome sequencing) by utilizing state of the art bioinformatics tools. This position assists in developing and conducting research projects, including experimental design, data analysis, and documentation of experiment results.
PRIMARY DUTIES AND RESPONSIBILITIES:
Designs, develops, and implements:
Algorithms and computer software for analyzing high-throughput, massively parallel genomic data sets.
Relational databases (SQL/NoSQL) and web server maintenance.
Familiar with biological data storage and format, including genomics data search, query, verification, and submission.
Independent research projects, including design of research protocols and development of procedures for the collection, verification, and management of data.
Performs integrative analysis of high-throughput biological data and writes interpretative reports.
Trains other researchers on the daily use of bioinformatics software and biology databases.
Assists with grant preparation and reporting of methods, data, and results.
REQUIRED QUALIFICATIONS:
Bachelor’s degree in computer science, bioinformatics, biostatistics, or related field plus 4 years of experience; OR master’s degree in computer science, bioinformatics, biostatistics, or related field plus 2 years of experience; OR combined education and experience of 9 years.
PREFERRED QUALIFICATIONS:
Master’s degree or Ph.D. in computer science or related field highly preferred.
Knowledge of developmental biology and/or regenerative medicine would also be a plus.
Experience with single cell sequencing and epigenomics highly preferred.
Proficiency with Linux system, common programing languages (e.g., R, Python, Perl, PHP), and bioinformatics analysis tools.
Excellent writing, communication and interpersonal skills.
The hiring range for this position is $53,830 – $72,675 annually. This position is eligible for full-time benefits. Please visit our website at http://hr.wustl.edu to view a summary of benefits.
VISIT https://jobs.wustl.edu and search job ID 43692 to apply.
We seek highly motivated post-doctoral Fellows interested in cell signaling at the intersection of development, stem cells and cancer. Candidates will join the group of Dr. Andres Lebensohn, an Earl Stadtman Principal Investigator in the Laboratory of Cellular and Molecular Biology (LCMB) at the Center for Cancer Research (CCR) of the National Cancer Institute (NCI), National Institutes of Health (NIH). We combine functional genomics, CRISPR/Cas9-mediated genome editing, cell biology and biochemistry to study how signaling pathways are used, reused and repurposed to drive the myriad different cellular processes that give rise to tissues and organs during embryonic development, and maintain them in adult life. A major focus of the lab is on WNT/R-spondin signaling, a fundamental pathway that orchestrates embryonic patterning and morphogenesis, and promotes stem cell self-renewal and tissue regeneration. We recently discovered a new mode of signaling by R-spondins that does not require LGRs, thought to be the main R-spondin receptors (https://elifesciences.org/articles/33126). We seek a Fellow who will use mouse genetics to elucidate the physiological functions of LGR-independent signaling by R-spondins during embryonic development and in stem cells. Descriptions of this and other research projects in the lab can be found at https://ccr.cancer.gov/Laboratory-of-Cellular-and-Molecular-Biology/andres-m-lebensohn.
About the Center for Cancer Research and the Laboratory of Cellular and Molecular Biology:
The CCR is the largest division of the NCI intramural research program, comprising nearly 250 basic and clinical research groups. In the words of Dr. Tom Misteli, director of the CCR, “Our scientists enjoy complete intellectual freedom and are expected to creatively and innovatively explore the most important questions in the field of cancer research and treatment.” The LCMB (https://ccr.cancer.gov/Laboratory-of-Cellular-and-Molecular-Biology) is located at the NIH Bethesda campus within a vibrant biomedical research community. We have access to state-of-the art research facilities and close links to the NIH Clinical Center, one of the leading clinical research hospitals in the world. The open lab space, collegial atmosphere and joint weekly seminars shared by the seven groups in the LCMB give Fellows a strong support network and plenty of opportunities to present and discuss their research with close peers. A short metro ride to Washington D.C. and the surrounding areas provides Fellows and their families access to numerous free museums (https://washington.org/free-things-to-do), excellent culinary choices and many outdoor activities.
Qualifications:
Candidates must have completed a Ph.D. and should be recent graduates or have less than 3 years of post-doctoral experience by the desired start date. Candidates with backgrounds in developmental, stem cell or cancer biology are encouraged to apply. Experience in mouse genetics is required for this project, and experience with organoid models is welcome but not required. Fellows will have exceptional stipend support for the entire duration of their post-doctorate, funds for travel to scientific conferences, and full healthcare benefits.
How to Apply:
Please send a cover letter describing your research accomplishments and future interests, including your specific interest in my lab, a CV with bibliography, and contact information for three references to andres.lebensohn@nih.gov. Please use the following email subject: “Application for post-doc position in WNT/R-spondin signaling”
As Lewis Wolpert put it (Wolpert, PLoS Biology 2010), if you extend your two arms, you will likely find that they match in length by better than 0.2%, though they do not seem to directly communicate during their development and growth. Similarly, flowers in an individual plant are likely to be almost identical in size and shape. Such precision, or robustness, in organ size might seem trivial when conceptualizing observations as reflecting a ‘developmental program’ with series of well-defined steps and ‘decisions’, but it is less so when thinking of the myriad of cells that form an organ and that divide and grow asynchronously (Figure 1). This raises the question of how organs and organisms reach well-defined sizes and forms, in the face of considerable spatial and temporal fluctuations of cell growth and deformation during morphogenesis (Hong et al. 2018, Uyttewaal et al. 2012, Kierzkowski et al. 2012, Hipfner et al. 2004).
Figure 1. Cell growth is heterogeneous in space and time. Example of a sepal (green leaf-like organ that protects a flower before it opens) from the model plant Arabidopsis thaliana. The color scale corresponds to growth rates (high in red, low in blue).
This question might find a very simple answer. As an organ grows larger and larger, an individual cell has less and less influence on overall size and shape of the organ; accordingly, precision of organ size would increase with the number of its cells. This is somehow like taking many independent measurements of an experimental parameter, the precision of the mean value of the parameter increases with the number of measurements, or, equivalently, the standard error of the mean (SEM) decreases with the number of measurements according to the law of large numbers. In an organ, if cells expanded randomly and independently from each other, then organ growth rate, which combines growth of all cells, would become increasingly precise as the number of cells increases (Hong et al. 2018). Nevertheless, this scenario cannot be fully true because cells communicate with each other and, therefore, are not necessarily independent. This led us to investigate how growth fluctuates according to the size of the region of interest, from cell to organ, so as to understand how precision of organ size can be achieved (Fruleux & Boudaoud, 2019). Interestingly, an independent and complementary study obtained results that are similar to ours (Damavandi & Lubensky, 2019).
A key question is whether there are mechanisms that enhance robustness of organ shape. If growth fluctuates between cells, those growing relatively fast will tend to push their neighbors, generating forces within the tissues. More generally, any tissue with heterogeneous growth builds up internal forces (internal stress) due to conflicts between cells or groups of cells. Such internal forces might be instructive: They could be sensed and used by cells to regulate their growth (Shraiman, 2005). This can be pictured by thinking of a group of blindfolded persons, each holding the hands of two neighbors, all involved in a round dance. Each dancer moves at its own natural pace and so pace fluctuates from dancer to dancer; as a consequence each dancer feels forces from the hands of the neighbors. Can the dancers use information provided by these forces to circle at the same speed?
Cells within a tissue can sense and respond to forces. It was first hypothesized that proliferation is affected by such forces, with cells under mechanical compression or tension reducing or increasing their division rates, respectively (Shraiman, 2005), which has received indirect and direct support in developing fruit fly and zebrafish (LeGoff et al., 2013; Campinho et al. 2013; Mao et al., 2013; Pan et al., 2016). The cytoskeleton may also respond to forces: actomyosin cables are reinforced by mechanical tension in the wing imaginal disk of Drosophila (LeGoff et al., 2013; Duda et al., 2017). The synthesis of the extracellular matrix (ECM) might also be modulated in response to mechanical signals. In plant cells, growth is driven by the cell’s inner pressure and restrained by the cell wall (the equivalent of the ECM) that encloses cells. The deposition of cellulose fibers, the main load-bearing component of the plant cell wall, depends on wall tension (Hamant et al. 2008, Hervieux et al., 2017), likely leading to reinforcing the cell wall in the direction of maximal tension (Sampathkumar et al., 2014). We aimed to incorporate all these responses to mechanical signals in a unifying framework, so as to be able to reach conclusions that may be relevant to robustness of morphogenesis in many developmental systems.
We described a tissue as a two-dimensional (2D) absract continuous medium, so as to make the calculations tractable and the conclusions general; a 2D model captures all main processes at play (notably directional processes such as those related to cytoskeleton), while a continuous model enables to disregard the details of cell arrangements and describe them at a supracellular level. We assumed the medium to be viscous, which accounts for the long-term relaxation and remodeling of cell junctions, cytoskeleton, ECM, or cell wall. We assumed the medium to be anisotropic; anisotropy is quantified with a mathematical object known as nematic tensor, which has been extensively used to measure fibril orientation and degree of orientation in cells and tissues (Boudaoud et al., 2014). We describe in the model the dynamics of density (density of cells, of cytoskeleton, of ECM, or of cell wall) and nematic tensor, that vary in time and space due to tissue expansion and deformation and to synthesis; for instance, ECM density decreases if tissue expands without synthesis, increases if cells exocytose ECM components, while ECM fibers align passively if the tissue is stretched in one direction.
Up to this point, all assumptions are rather straightforward. Our key additional assumption is that synthesis is modulated according to mechanical stress, reflecting the evidence mentioned above: density is modulated (positively or negatively) in response to level of mechanical stress and anisotropy is modulated (positively or negatively) in response to directional mechanical stress; the corresponding response parameters (which may be positive or negative) are the main parameters of our model. Positive response to stress levels describes putative increase in ECM / cell wall synthesis, and subsequent reduction in expansion, in response to tensional stress; negative response to stress levels describes reduction in proliferation due to compression. Positive response to directional stress corresponds to cytoskeleton / cellulose fibers reinforcement under tension; negative respond to directional stress corresponds to cell intercalation under directional stretching. Accordingly, the sign of the response (positive or negative) allows us to account for biological diversity in mechanical responses.
Figure 2. Response to a localized perturbation.Snapshots of the predicted dynamics following a perturbation of tissue density in the form of a disk at time t0. The density is color-coded (white corresponds to no deviation from average density and dark red to large deviations). Tissue anisotropy is indicated by the small lines: the stiffest tissue direction corresponds to line orientation and the degree of anisotropy to line length.
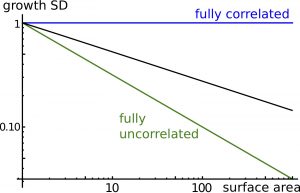
We first examined tissue dynamics following a perturbation in tissue density of a small disk-shaped region (Figure 2), which could correspond to a transient depolymerization of cytoskeleton or a transient reduction in ECM / cell wall synthesis The model predicts the expansion of this region due to tissue growth and the relaxation of density to its average value due to tissue response to mechanical stress; anisotropy orients circumferentially around the perturbed region, like in wound healing. We then considered the effects of a stochastic component to synthesis, reflecting temporal fluctuations of cellular dynamics. The model predicts that cells (more rigorously regions as the model is continuous) are correlated to each other at long distances (Figure 3): Cells do not grow independently (although they synthesize materials independently) and growth fluctuations in one cell remain correlated with growth fluctuations in a cell far apart. Long-range correlations are characterized by an exponent indicating how fluctuations of an element of the tissue scale with its size (Figure 3). Thus individual cells may have a stronger impact on organs size than if they were fully independent.
Figure 3. Long-range correlations. Cell-to-cell correlations quantify the degree of spatial coordination between cell fluctuations. The plot shows relative fluctuations in areal expansion of the tissue (growth SD) as a function of size of the region of interest (surface area). If cells wall fully correlated, fluctuations in areal expansion would not depend on size (exponent 0, blue line) and if cells wall fully uncorrelated, fluctuations in areal size would quickly decrease with size (exponent -0.5, green line). In general, the model predicts an intermediate behavior (with exponent ranging between -0.5 and 0, dark line), known as long-range correlations.
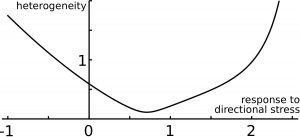
Finally, we examined the precision of organ shape and size. Positive response to mechanical stress levels (tissue reinforcement in response to tension) makes organ size less precise than with negative response (tissue softening in response to tension). Interestingly, increasing response to directional stress has a non-intuitive effect on organ precision (Figure 4): first the precision of organ size is enhanced, but then precision reaches a minimum, and then it increases for higher response levels. Thus, the model predicts an optimal response to directional mechanical stress for robustness of organ shape and size (Figure 4). This can be qualitatively understood by getting back to the dance picture. The round dance could be broken if the blindfolded dancers respond too weakly or too strongly to the forces they perceive from their neighbors; accordingly, an optimal response to forces would ensure a harmonious dance.
Figure 4.Precision of size and shape. The predicted ‘error’ on organ size (heterogeneity) is shown as a function of the level of response to directional mechanical stress, and features an optimal response level.
Long-range correlations of growth fluctuations are a crucial prediction of our analysis since it is strongly related to how cellular fluctuations impact robustness of morphogenesis. We therefore believe that correlations need to be measured at many scales in different experimental systems, in various genetic backgrounds, and in absence and presence of transient genetic or chemical perturbations. This would enable to disentangle the contributions of cell proliferation, cytoskeleton, ECM, and cell wall to the control of robustness. In addition, we speculate that long-range correlations could be involved in absolute size control; additional work would be required to investigate how this could occur. Finally, we note that we focused here on precision and robustness of morphogenesis, starting from the examples of bilateral symmetry in animals and flowers in plants; but this does not always need to be the case. For instance, plant leaves and roots are phenotypically highly plastic to adapt to environmental constraints. Tumor expansion and metastasis are likely favored by heterogeneous morphogenesis. Altogether, our work sheds light on the importance of considering fluctuations in understanding developmental processes.
References
Boudaoud, A., Burian, A., Borowska-Wykręt, D., Uyttewaal, M., Wrzalik, R., Kwiatkowska, D., & Hamant, O. (2014). FibrilTool, an ImageJ plug-in to quantify fibrillar structures in raw microscopy images. Nature protocols, 9(2), 457. https://www.ncbi.nlm.nih.gov/pubmed/24481272
Campinho, P., Behrndt, M., Ranft, J., Risler, T., Minc, N., & Heisenberg, C. P. (2013). Tension-oriented cell divisions limit anisotropic tissue tension in epithelial spreading during zebrafish epiboly. Nature cell biology, 15(12), 1405. https://www.ncbi.nlm.nih.gov/pubmed/24212092
Damavandi, O. K., & Lubensky, D. K. (2019). Statistics of noisy growth with mechanical feedback in elastic tissues. Proceedings of the National Academy of Sciences, 116(12), 5350-5355. https://www.ncbi.nlm.nih.gov/pubmed/30819899
Duda, M., Kirkland, N. J., Khalilgharibi, N., Tozluoglu, M., Yuen, A. C., Carpi, N., … & Mao, Y. (2019). Polarization of myosin II refines tissue material properties to buffer mechanical stress. Developmental cell, 48(2), 245-260. https://www.ncbi.nlm.nih.gov/pubmed/30695698
Fruleux, A., & Boudaoud, A. (2019). Modulation of tissue growth heterogeneity by responses to mechanical stress. Proceedings of the National Academy of Sciences, 116(6), 1940-1945. https://www.ncbi.nlm.nih.gov/pubmed/30674660
Hamant, O., Heisler, M. G., Jönsson, H., Krupinski, P., Uyttewaal, M., Bokov, P., … & Couder, Y. (2008). Developmental patterning by mechanical signals in Arabidopsis. science,322(5908), 1650-1655. https://www.ncbi.nlm.nih.gov/pubmed/19074340
Hervieux, N., Tsugawa, S., Fruleux, A., Dumond, M., Routier-Kierzkowska, A. L., Komatsuzaki, T., … & Hamant, O. (2017). Mechanical shielding of rapidly growing cells buffers growth heterogeneity and contributes to organ shape reproducibility. Current Biology, 27(22), 3468-3479. https://www.ncbi.nlm.nih.gov/pubmed/29129534
Hong, L., Dumond, M., Zhu, M., Tsugawa, S., Li, C. B., Boudaoud, A., … & Roeder, A. H. (2018). Heterogeneity and robustness in plant morphogenesis: from cells to organs. Annual review of plant biology, 69, 469-495. https://www.ncbi.nlm.nih.gov/pubmed/29505739
Hipfner, D. R., & Cohen, S. M. (2004). Connecting proliferation and apoptosis in development and disease. Nature Reviews Molecular Cell Biology, 5(10), 805. https://www.ncbi.nlm.nih.gov/pubmed/15459661
Kierzkowski, D., Nakayama, N., Routier-Kierzkowska, A. L., Weber, A., Bayer, E., Schorderet, M., … & Smith, R. S. (2012). Elastic domains regulate growth and organogenesis in the plant shoot apical meristem. Science, 335(6072), 1096-1099. https://www.ncbi.nlm.nih.gov/pubmed/22383847
LeGoff, L., Rouault, H., & Lecuit, T. (2013). A global pattern of mechanical stress polarizes cell divisions and cell shape in the growing Drosophila wing disc. Development,140(19), 4051-4059. https://www.ncbi.nlm.nih.gov/pubmed/24046320
Mao, Y., Tournier, A. L., Hoppe, A., Kester, L., Thompson, B. J., & Tapon, N. (2013). Differential proliferation rates generate patterns of mechanical tension that orient tissue growth. The EMBO journal, 32(21), 2790-2803. https://www.ncbi.nlm.nih.gov/pubmed/24022370
Pan, Y., Heemskerk, I., Ibar, C., Shraiman, B. I., & Irvine, K. D. (2016). Differential growth triggers mechanical feedback that elevates Hippo signaling. Proceedings of the National Academy of Sciences, 113(45), E6974-E6983. https://www.ncbi.nlm.nih.gov/pubmed/27791172
Sampathkumar, A., Krupinski, P., Wightman, R., Milani, P., Berquand, A., Boudaoud, A., … & Meyerowitz, E. M. (2014). Subcellular and supracellular mechanical stress prescribes cytoskeleton behavior in Arabidopsis cotyledon pavement cells. Elife, 3, e01967. https://www.ncbi.nlm.nih.gov/pubmed/24740969
Shraiman, B. I. (2005). Mechanical feedback as a possible regulator of tissue growth. Proceedings of the National Academy of Sciences, 102(9), 3318-3323. https://www.ncbi.nlm.nih.gov/pubmed/15728365
Uyttewaal, M., Burian, A., Alim, K., Landrein, B., Borowska-Wykręt, D., Dedieu, A., … & Kwiatkowska, D. (2012). Mechanical stress acts via katanin to amplify differences in growth rate between adjacent cells in Arabidopsis. Cell, 149(2), 439-451. https://www.ncbi.nlm.nih.gov/pubmed/22500806
There is a vacancy for a 4-year PhD student position at the Sars International Centre for Marine Molecular Biology in the research group of Dr. Pawel Burkhardt (www.sars.no). The position is funded by Sars Centre core funding. The Sars Centre belongs to the University of Bergen and is partner of the European Molecular Biology Laboratory (EMBL) (www.embl.de). The place of work will be at the Sars Centre. The starting date is flexible, but should be no later than 01 October 2019.
About the project/work tasks:
The goal of the Burkhardt group is to reconstruct the evolutionary origin of synapses and neurons. The group is particularly interested in studying synaptic protein homologs in choanoflagellates, sponges and ctenophores. We are looking for a highly self-motivated and enthusiastic PhD student with interests in evolutionary biology, neurobiology or cell biology. The successful candidate will undertake research with the possibility to use a variety of techniques, ranging from various biochemical methods to X-ray crystallography, super resolution immunofluorescence and electron microscopy, to study synaptic protein homologs in choanoflagellates and sponges (for examples see: Burkhardt et al, 2011 PNAS; Burkhardt et al, 2014 Mol Bio Evo; Bhattacharyya et al, 2016 eLife; Laundon et al, 2019 PLoS Biology). The PhD project will focus on when protein signalling complexes required for synaptic activity first evolved and how they functioned at a molecular level.
Qualifications and personal qualities:
The applicant must hold a master’s degree or the equivalent or must have submitted his/her master’s thesis for assessment prior to the application deadline. It is a condition of employment that the master’s degree has been awarded.
Routine experience in standard molecular and cellular biology techniques is required
Practical experience in a variety of biochemical techniques (e.g. protein expression & purification, Co-IPs, mass spec) and with different fluorescence imaging techniques is highly desirable
Specific experience with choanoflagellates is beneficial, but not essential.
The ability to work both independently and to cooperate with others and a high motivation/enthusiasm is essential.
Proficiency in both written and oral English
About the PhD position:
The duration of the PhD position is 4 years, of which 25 per cent of the time each year comprises required duties associated with research, teaching and dissemination of results. The employment period may be reduced if you have previously been employed in a recruitment position.
About the research training:
As a PhD Candidate, you must participate in an approved educational programme for a PhD degree within a period of 4 years. A final plan for the implementation of the research training must be approved by the faculty within three months after you have commenced in the position. It is a condition that you satisfy the enrolment requirements for the PhD programme at the University of Bergen.
We can offer:
A professional, challenging and international working environment.
Well-equipped, modern laboratories and facilities
Salary at pay grade 51 (Code 1017/Pay range 20, alternative 9) in the state salary scale. Currently equal to NOK 449.400. Further promotions are made according to qualifications and length of service in the position
Enrolment in the Norwegian Public Service Pension Fund.
A position in an inclusive workplace (IA enterprise).
A brief account of the applicant’s research interests and motivation for applying for the position
The names and contact information for two reference persons. One of these must be the main advisor for the master’s thesis or equivalent thesis
CV
Transcripts and diplomas showing completion of the bachelor’s and master’s degrees, or official confirmation that the master’s thesis has been submitted
Relevant certificates/references
List of published papers and pre-prints or other relevant scientific work.
Please note that the entire application and relevant appendices with certified translations into English or a Scandinavian language must be uploaded at Jobbnorge. Applications submitted without required information or applications sent as e-mails will not be considered. Only submitted documents will be subjected to an expert assessment.
Detailed information about the position can be obtained by contacting: Group Leader Pawel Burkhardt, phone: +47 906 48 539, email: Pawel.Burkhardt@uib.no
A postdoctoral position is available in the Laboratory of Gareth Fraser (www.fraser-lab.net) at the Department of Biology, University of Florida. Our lab is focused on a range of research projects based around the central theme of vertebrate evolutionary developmental biology – they include tooth development and regeneration, craniofacial diversity, EvoDevOmics, and skin appendage evolution and development.
We are looking for a motivated and creative scientist to continue to develop the shark and ray as comparative evodevo models for a range of research projects. The primary focus of this position will be stem cell mediated tooth regeneration in sharks and rays, however we do encourage development of the candidates own creative research ideas and interests, so there is potential here to develop a project that suits the candidate in line with the focal themes of the lab. The salary is competitive and commensurate with qualifications and experience, and includes a full benefits package. The applicant selected for this position would begin the beginning of the Fall 2019 term. The anticipated date would be August 16, 2019.
Qualifications
Candidates should have an interest in evolutionary developmental biology, and vertebrate diversity. Knowledge of developmental biology techniques, RNAseq analyses or stem/regenerative biology is desirable.
Application Instructions
For full consideration, applications must be submitted online at http://apply.interfolio.com/62577 and must include: (1) a letter of application summarizing the applicant’s qualifications and interests, (2) a complete curriculum vitae, (3) a statement on research goals, and (4) ) a list of at least three references. After initial review, applicants who are chosen to receive further consideration will be asked to request confidential letters of recommendation from the references. If you would like further information about the position please contact Gareth Fraser. Email: g.fraser@ufl.edu.
Application deadline is May 29, 2019.
All candidates for employment are subject to a pre-employment screening which includes a review of criminal records, reference checks, and verification of education.
The selected candidate will be required to provide an official transcript to the hiring department upon hire. A transcript will not be considered “official” if a designation of “Issued to Student” is visible. Degrees earned from an educational institution outside of the United States require evaluation by a professional credentialing service provider approved by the National Association of Credential Evaluation Services (NACES), which can be found at http://www.naces.org/.
The University of Florida is an equal opportunity institution dedicated to building a broadly diverse and inclusive faculty and staff. Searches are conducted in accordance with Florida’s Sunshine Law. If an accommodation due to disability is needed in order to apply for this position, please call (352) 392-2477 or the Florida Relay System at (800) 955-8771 (TDD).
The Department particularly welcomes applicants who can contribute to a diverse and inclusive environment through their scholarship, teaching, mentoring, and professional service. The university and greater Gainesville communities enjoy a diversity of cultural events, restaurants, year-round outdoor recreational activities, and social opportunities.
Applications are invited from highly motivated individuals who are interested in fundamental mechanisms of neurodevelopment and neurogenetic disorders. The focus of the project is to understand neural developmental and neurological phenotypes in mouse models ofTuberous Sclerosis Complex.The fellowship is in the laboratories of Sara Wilson at the Department of Integrative Medical Biology (IMB), and Leif Carlsson at Umeå Center for Molecular Medicine (UCMM) both laboratories at Umeå University, Sweden. The facilities provide an interactive modern environmentwith easy access to good core facilities. The fellowship is administratively placedat IMB, which is an interdisciplinary department focusing on questions in basic and medical sciences. The fellowshipis funded for two years and is available immediately. The working ‘day to day’ language in the laboratories is English.
Requirements:Individuals with a background in mouse genetics and handling, developmental biology, neuroscience, molecular and/or cell biology with a keen interest in developmental neuroscienceare encouraged to apply. Experience with rodent motor circuits or rodent motor or seizure behavior analysis is a big advantage.Technical experience with imaging, molecular biology, immunohistochemistry,in situhybridization, vertebrate embryonic model systemsare required. Technical experience with mouse embryo electroporation and /or neurite outgrowth/migration assayswill also be positively evaluated. The successful candidate will have or about to receive a Ph.D.in a relevant discipline, have good communication skills and be proficient in written and spoken English. The most successful candidatewill have ahigh level of motivation, be organized, rigorous and have the ability to work both independently and within a team.
Application:Please submit your application (reference 2019SW7) by 31stMay 2019 to sara.wilson@umu.seby sending the following documents as a single pdf file:
A short cover letter (not more than 1 page) to include a description of your research experience and suitability for the position.
Curriculum Vitaeincluding: publication list, technical expertise, names and contact information for three referees.
Informal enquiries may be directed to Dr. Sara Wilson (sara.wilson@umu.se) or Prof. Leif Carlsson (leif.carlsson@umu.se).
How can a genome’s cis-regulatory potential drive robust gene expression throughout embryonic development? Do topologically associating domains (TADs) truly contribute to gene expression by spatially constraining the regulatory actions of cis-sequences? And might phase separation by multi-valent proteins also be involved in the regulation of genes and genomic regions? The recent Company of Biologists Workshop on “Chromatin-based regulation of development”, organized by Benoit Bruneau (Gladstone Institutes, UCSF) and Joanna Wysocka (Stanford University), was the perfect playground to discuss important questions like these, and to stimulate interactions between early career and established scientists. Why was this Workshop so great to me (currently a postdoc in the lab of François Spitz, Institut Pasteur, France)? And why should you – a young investigator – apply to attend a future one? (more…)
 (No Ratings Yet)
(No Ratings Yet)











 (1 votes)
(1 votes)




 (5 votes)
(5 votes)