3 year-POSTDOC + Starting grant (OPEN-CALL BEATRIU de PINOS program) to join our lab on EvoDevoGenomics in Barcelona
We are seeking candidates to join our lab to study our favorite chordate model Oikopleura dioica, in which we are currently interested in heart and muscle development, tail elongation and the impact of gene loss on the evolution of gene regulatory signalling networks. Click here for a tour “A day in our lab” posted in The Node
We have also engaged a new EcoEvoDevo line investigating if the developmental mechanisms of marine embryos are ready to respond to climate change, including biotoxins derived from algal blooms. Click here for a tour on this new EcoEvoDevo adventure.
Our approaches include RNAseq, CRISPR, RNAi, Fluorescent-Microscopy
DEADLINE call: March 4th 2019 (contact for enquiries as soon as possible canestro@ub.edu)
REQUIREMENT: to have defended the PhD within the period January 1st 2011 – December 31st 2016
DURATION: 3 years: starting not later than February 2020
FUNDING: 132.300€ total gross salary for 3 years + 12.000€ research funds
CONTACT: Interested candidates, please send an email to Cristian Cañestro (canestro@ub.edu), including a brief letter of interest, a brief CV, including list of publications with their impact, and technical skills for post-doc applications, and official scores for doctoral candidates, all together in ONE single pdf file.
The Company of Biologists, as well as publishing Development and four other journals and offering travel and conference grants, runs a successful series of Workshops. The Workshops provide leading experts and early career scientists from a diverse range of scientific backgrounds with a stimulating environment for the cross-fertilisation of interdisciplinary ideas. The programmes are carefully developed and are intended to champion the novel techniques and innovations that will underpin important scientific advances.
The Workshop Committee are currently seeking proposals for Workshops to be held during 2021.The deadline for applications is 31 May 2019.
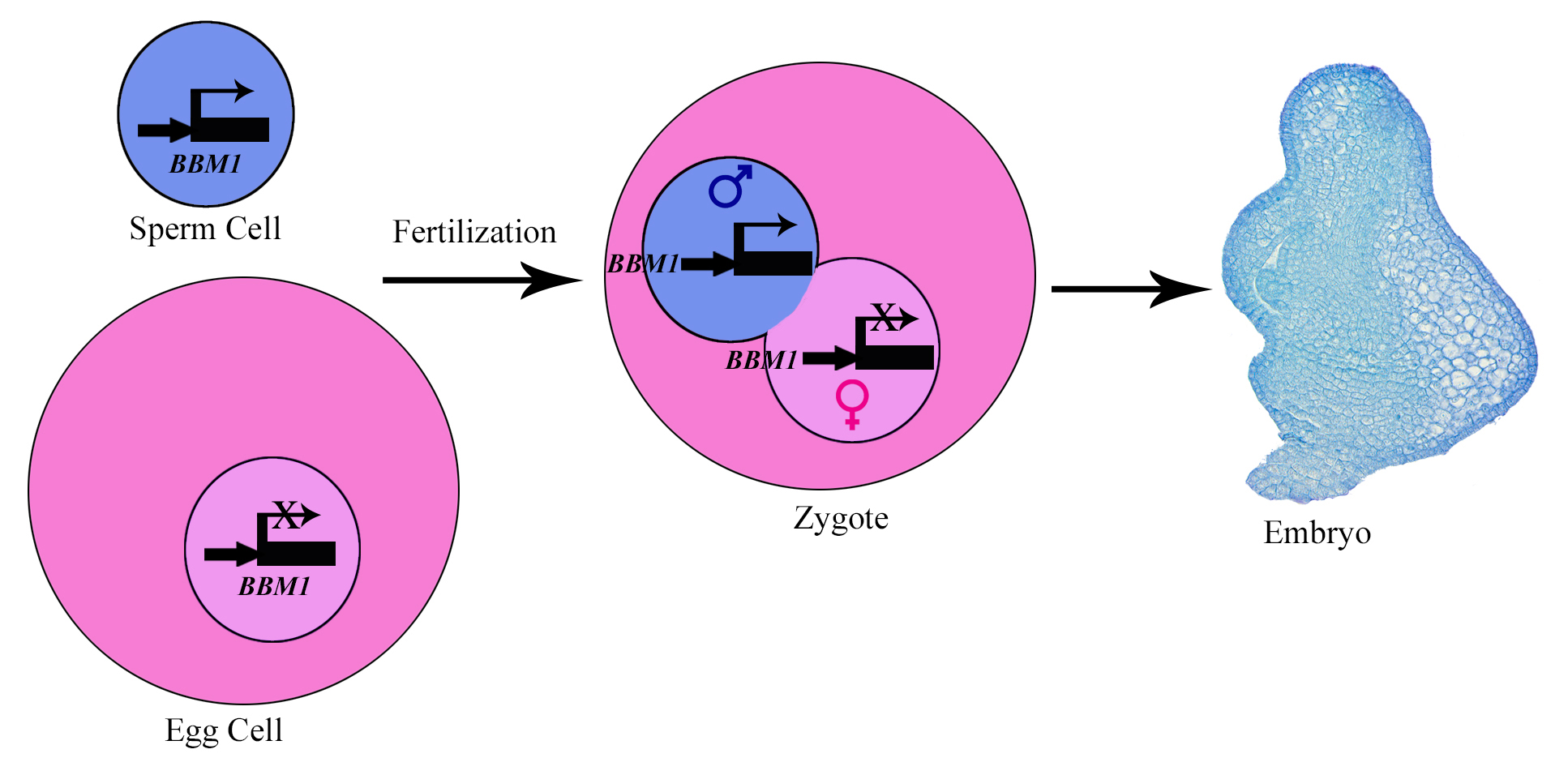
For sexually reproducing organisms, the diploid life cycle starts with the fusion of a sperm cell with an egg cell. This process, known as fertilization, results in the formation of a zygote, the first diploid cell from which a multicellular embryo develops. In animals, embryo initiation has been shown to be under maternal control, driven by the gene products stored in the egg cytoplasm. For example, in the absence of zygotic transcription, Caenorhabditis elegans zygotes can progress to the 100 cell stage before arresting1. This raises an important question: if the maternal gene products are able to drive embryo development, why do egg cells depend on sperm cell fertilization for embryo initiation? Why is there a fertilization block in sexual organisms?
This work was started in the Sundaresan lab at UC Davis to understand embryo initiation in rice, particularly the role of BABY BOOM (BBM) genes in it. BBM genes belong to AP2 family of transcription factors. Our previous study showed that the expression of three BBM genes is induced in zygotes after fertilization2. Among these, BBM1 was specifically expressed from the male allele (Fig. 1). Although this was an important observation, it needed further confirmation. We attempted confocal imaging of intact zygotes expressing BBM-GFP, but when BBM-GFP plants were used as either the male or the female parent, it did not work: rice carpels are thick and green, so the GFP signal was impossible to view through this tissue. Isolating zygotes is possible in rice and had been done by our team previously, but it is a tedious process which involves manual dissection. Isolating and imaging a statistically significant number of zygotes for our purpose would have probably taken us a year or more, so we decided to use antibodies against GFP to detect the BBM1-GFP expression in zygotes. However, this created another challenge- is immunohistochemistry sensitive enough to detect the expression in a single nucleus? It did work, and it worked well!
Figure 1. Mechanism of embryo initiation in rice. BBM1, an AP-2 domain transcription factor is expressed in the sperm cell but not in the egg cell. After fertilization, its expression from the male genome activates the embryo initiation program in the zygote. Its expression in the later stages of embryo development is required for embryo organ formation.
We could now confirm that BBM1 is expressed only from the male allele in zygotes, immediately after fertilization (Fig. 1). We also found that ectopic overexpression of BBM1 induces somatic embryogenesis in heterologous tissues like leaves. The latter observation, combined with male-specific expression of BBM1 in the early zygotes, lead us to hypothesize that BBM1 expression probably initiates embryogenesis after fertilization. To test this hypothesis, we drove the expression of BBM1 in egg cells using an Arabidopsis egg cell promoter3. At that time (back in 2014), we were not even sure if this promoter was going to work in rice. Egg cell expression of BBM1 resulted in embryo formation without the need for fertilization, a process known as parthenogenesis. Thus, it turns out that the sperm cell transmitted BBM1 initiates embryogenesis after fertilization (Fig.1). BBM-like genes were first discovered in Brassica microspore cultures4. Although, BBM-like genes from Arabidopsis and Brassica have been shown to induce somatic embryogenesis4, their role is zygotic embryogenesis is not known because their loss-of-function mutants do not show any embryonic phenotypes.
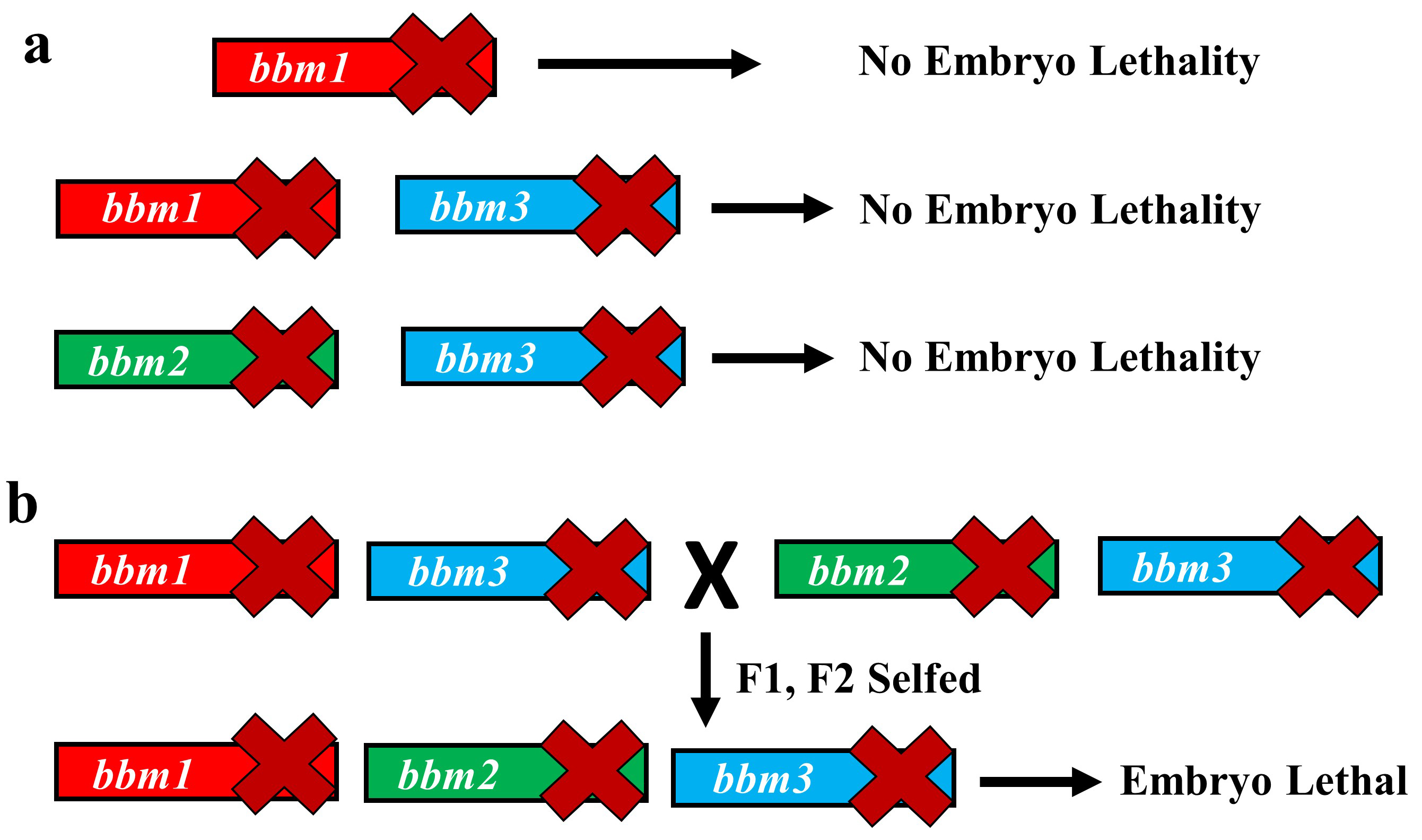
The variety of rice that we use for our experiments, Kitaake, has one of the shortest generation times among different experimental varieties of rice (still about four months!). However, generating new transgenic lines from tissue culture can take up to 8 months. So, using rice for studying genetics can be a lengthy affair. However, we decided to take this long path and study the genetics of BBM genes in rice. The vectors for CRISPR-Cas9 gene editing were provided by Dr. Bing Yang from Iowa State University. We created a single knockout mutant of BBM1, but it did not show any phenotype (Fig. 2a). The double mutant of BBM1 and BBM3 (bbm1 bbm3) did not show any embryonic phenotype either (Fig 2a). An attempt to create a triple mutant using a single CRISPR-Cas9 construct was unsuccessful as the triple knockout construct would not regenerate plants in the tissue culture. This was a setback; however, it also meant that triple mutant is probably embryo lethal. To work around this, we created another double mutant of BBM2 and BBM3 (bbm2 bbm3), crossed it with bbm1bbm3 double mutant and selfed the progeny for the next two generation (Fig. 2b). The triple mutant was indeed embryo lethal (Fig. 2b). But instead of observing a typical 25% lethality (expected from Mendelian genetics), because the mother plant was segregating only for BBM1 (BBM1/bbm1 bbm2/bbm2 bbm3/bbm3), there was a 36% lethality. This was found to be linked to the male transmission of BBM1 from the sperm cell (Fig. 1). This proves that a functional copy of BBM1 from the sperm cell is essential for embryo initiation in rice. This is a novel mechanism that explains why the egg cell (at least in rice) depends on sperm cell fertilization to initiate embryo development.
Figure 2. Schematic showing constructs for mutants and redundancy in BBM1, BBM2 and BBM3 genes. (a) bbm1 mutant alone, or in combination with bbm3 have no embryo phenotypes. Also, loss-of-function bbm2 bbm3 double mutant shows no aberrant embryonic phenotype. (b) bbm1 bbm3 and bbm2 bbm3 double mutants were crossed to create bbm1 bbm2 bbm3 triple knockout mutant, which was embryo lethal.
This work was started purely as a development biology project, but we soon realized this understanding of the basic mechanism of embryo initiation can have agricultural applications. The first application is the conversion of egg cell directly into an embryo without fertilization: it meant we could generate haploid plants. Haploids have only one set of chromosomes, in this case maternal. This makes them efficient agricultural breeding tools as homozygous lines can be produced in one generation after chromosome doubling, bypassing the several generations it takes by inbreeding procedures. The technique we developed does not involve the tedious and laborious tissue culture techniques used in microspore cultures or rescuing and culturing haploid embryos in subsequent generations as seen in wide crossing procedures. The haploids can be grown simply from functional seeds (Fig. 3a).
Figure 3. Haploids and synthetic apomicts in rice. (a) Two segregating sibling plants from BBM1 egg cell expressing mother. The haploid plant is a result of parthenogenesis and diploid plant of sexual reproduction. (b) Progenies from a diploid synthetic apomixis mother. Diploid plant develops from the parthenogenesis of diploid egg cell created by MiMe and hence maternal clone. Tetraploid sibling develops sexually from the fusion of diploid gametes.
The second application is synthetic apomixis by which an unreduced diploid egg cell is converted into a maternal clone, allowing for the maintenance of hybrid vigor. Hybrid vigor, or heterosis refers to the increase in yield, growth or other quantitative characteristics in F1 hybrids, compared to parental inbred lines. However, the genetic combinations that lead to this vigor in F1 hybrids, segregate in the F2 generation due to sexual reproduction and thus resulting in loss of vigor. For this reason, farmers need to buy hybrid seeds, every sowing season. Therefore, a hybrid crop that could self-reproduce through seeds while maintaining the parental heterozygosity would solve this problem.
During sexual reproduction, meiosis results in recombination and segregation of genetic traits, and fertilization creates new genetic combinations. MiMe (mitosis instead meiosis), is a genetic approach that skips meiosis and converts meiotic cell division into a mitotic like division5. This approach was developed by our collaborator, Raphael Mercier’s group at INRA France. We combined MiMe with our BBM1 induced parthenogenesis system. The two approaches together produced progenies which have the same genetic constitution as that of the mother plant. In other words, the progenies are genetic clones of the mother plant (Fig. 3b). The MiMe produces diploid egg cells which parthenogenetically develop into embryos due to BBM1 expression. The clonal nature of progeny and mother plant was confirmed by whole genome sequencing (Dr. Debra Skinner analyzed the sequences). The endosperm that developed was, however, sexual. Since the gametes are diploid, the endosperm in progenies is hexaploid instead of the usual triploid. This 6X endosperm increases the seed size (Fig. 4a). Thus, this synthetic apomixis approach not only results in clonal propagation but also increases the seed size and hence the yield. This mode of apomixis is seen in some naturally apomictic plants like Boechera6 and others. Engineering apomixis in crop plants ensures fixation of hybrid vigor and stabilization of superior heterozygous genotypes. Also, the yield, quality and exchange of vegetatively propagated true seed crops can been improved by introgression of apomixis (hence seed propagation) as the accumulation of somatic mutations, viruses, and other pathogens over successive generations can be avoided. The successful engineering of apomixis in crop plants is a significant step towards achieving the food security for growing world population.
Figure 4. Seed size in synthetic apomixis plants. (a) Seeds from different genotypes having different ploidy for embryo (em) and endosperm (en). The seed size increases in synthetic apomictic pants due to increase in endosperm ploidy (6X). 1, wild-type; 2, haploid synthetic apomictic seed; 3, control MiMe; 4, diploid apomictic seed; and 5, tetraploid apomictic seed. (b) Genotyping team at Sundar lab, UC Davis.
A major challenge we faced during this study was to be able to genotype the thousands of rice plants used in this study (Fig. 4b). The plants needed to be genotyped for T-DNA insertions, copy number determination and CRISPR-Cas9 mutation analysis (quite often for three genes in the same plant). We highly appreciate the helping hand from Bao Nguyen (now at UC Santa Cruz) and Alina Yalda. Preparing samples for flow cytometry for ploidy determination was another time-consuming procedure. However, the final outcome of being able to decipher the mechanism of sexual reproduction in rice and utilization of this knowledge to make it reproduce asexually, made it all worth the effort!
Full Time one year employment contract (renewable for three years)
Location: University of Cyprus
The University of Cyprus (www.ucy.ac.cy) invites applications for one full time postdoctoral associate to work in the Laboratory of Cell and Developmental Biology (http://xeno.biol.ucy.ac.cy).
DUTIES AND RESPONSIBILITIES
The researcher will be part of a team working on a highly ambitious Research Promotion Foundation Strategic Infrastructure grant that aims to establish super-resolution microscopy in Cyprus and explore questions related to how cells sense and respond to mechanical stimuli. These will be addressed using cutting edge instrumentation and methodologies both in vitro and in vivo. The team will go on to employ the principles revealed by these studies for the development of novel therapeutic approaches for the treatment of cancer and metastatic disease, biomedical applications and in regenerative medicine. The post-doctoral associate will also be responsible for supervision of PhD and MSc candidates participating in the project, together with the group leader.
REQUIRED QUALIFICATIONS
PhD Degree in Cell or Molecular Biology or related area
An excellent research and academic record
Previous laboratory experience and knowledge of basic molecular and cellular biology techniques
Research experience in the areas of cell biology and/or mechanobiology
Ambition, enthusiasm and motivation, with an interest in a long-term career in research
Good publication record
APPLICATION PROCEDURE
All applications must include:
A cover letter, that describes how the applicant meets both the selection criteria and their motivation
Curriculum Vitae, including description of past research experience
Name and contact details of at least two referees
Qualified applicants will be invited for a personal interview
TERMS OF EMPLOYMENT
The position will be funded through the grant INFRASTRUCTURES/1216/0060 andinitiated on a year-long contract, renewable annually for 3 years. It will include a competitive compensation package, depending on the candidate’s qualifications and experience. The monthly salary is set between €2200- €2700, depending on the candidate’s qualifications and experience. Employee and employer contributions will be deducted from the above amount. 13th Salary bonus or medical insurance coverage are not provided, but the candidate can be included in the UCY health plan if they choose to do so.
Contact information
Applications must be emailed to skourip@ucy.ac.cy by 5th February 2019, with the title “Postdoc position-MRU-2019”.
For more information please contact the coordinator of the project Dr. Paris A. Skourides tel.: +357-22892895 or email: skourip@ucy.ac.cy.
In this post, I share how I learned about the Embryology course, what made me apply and what I brought back from it.
Nerdy embryology t-shirts
A couple of months ago, I was attending a scientific conference on cell fate in Roscoff, northern France. While waiting in line at the coffee break, I hear someone commenting on the t-shirt that I was wearing:
«I assume you went to the Embryology course in Woods Hole this year! You know, many people here have one of these…»
That is how I met Dominique Bergmann, plant developmental biologist at Stanford University, as she was noticing the many names and embryos on the back of my 2018 Embryology Course t-shirt. «I can’t find mine anymore, but in my case that’s from many many years ago», she later added.
Back of the Embryology 2018 course t-shirt. The names of all people involved in the course are included. The font used has been generated off of Paul Gerald Sanchez’s own handwriting. Embryo drawings from Ashley Rasys.
Despite of the years though, many people have been meticulously collecting those t-shirts. In fact, some of the scientists who I met this summer would have collections spanning decades. I remember Andres Collazo talking about how he strived to keep them in perfect conditions in order to showcase them every year, once back to the Marine Biological Laboratory for Embryology. I now understand why: each of those t-shirts tells multiple stories. Stories of science of course, but also of people who decided, that year, to spend six weeks sharing their passion for developmental biology.
A historical subject
During my undergraduate years I was lucky enough to find this passion. I studied biotechnology at the University of Palermo, in my italian hometown. Among the many courses, I still remember very clearly the only developmental biology class, taught by Ida Albanese. There I heard for the first time about developmental biology’s questions and approaches. Ida’s lectures often had a historical perspective, blending the experimental elegance of classical embryology with the latest molecular and biophysical characterisation of embryonic events. I was hooked, simply amazed by the progression of intertwined processes through which diversity and complexity are generated from a single fertilised egg.
At the end of the course Ida told us about Alberto Monroy, after whom our department was named. Monroy had a key role in shaping the field of developmental biology in southern Italy, having spent his career between Palermo, the Stazione Zoologica in Naples and the MBL. There he used to go every summer to conduct research and interact with the students. To honour him, the University and some private donors had established the A. Monroy Fellowship, which once a year funds the participation of an Italian student to the prestigious Embryology course. I googled it to see what the course was about. I said to myself, one day I’d like to be part of this.
Four years later I had completed my masters degree and joined Jacqueline Tabler‘s lab for my PhD. That moment, the beginning of a new challenging project, would have been perfect to take the course. With the support of my supervisor, I applied. I was performing my first experiments in the lab when I got the results. I felt extremely happy to accept the scholarship and prepare for a summer in Woods Hole, looking forward to making it part of my story as well.
Expectation vs reality
When I first visited the Embryology course website, I was struck by some sentences on the application form.
We would like to emphasize to applicants that MBL discovery courses are extremely rigorous […] There is little time for much else but course work.
I wondered what a samurai training course for developmental biologists looked like, and feared that would have been exhausting. Such fear was unfounded.
The first day of the course started with an introduction by the course directors, Dave Sherwood and Rich Schneider. They then suggested to start getting to know each other with a game. In front of the posters that we brought, each one of us had to talk about his own research to another student in the course. Afterwards, all back in the same break room, students would tell the entire class about the research of their fellow. Everybody’s enthusiasm for science filled the room in a way I had never witnessed before: we were all sharing what we truly loved about our projects.
That was only the beginning of a six-weeks journey of scientific discovery, creativity and fun. We would learn about a new field and organism everyday by some of the world experts on those topics. We would then have intense but informal Q&A sessions with them, during which you had to ask something and could ask anything.
Lab work was on in the afternoon. At the microscope, injecting and dissecting embryos, running reactions or just observing faculty and TAs performing their favourite experiments. We would be taught how to craft pipettes out of glass capillaries, in order to graft a tiny piece of tissue from an embryo to another. At a certain point of the night we would stop, as Paul Trainor put his forceps back into the drawer and started making amazing cocktails for us. Having fun in and out of the lab, trying new and crazy experiments, some of us even got authorship in a paper. That’s Woods Hole.
One of my favourite pictures from Embryology 2018. It’s 1AM and we’re going through the Xenopus module. Ray Keller discusses life and science with Anneke after having taught her how to make a Keller explant. Melvin is at his bench, still working on an experiment. Under the scope, Xenopus embryos are performing gastrulation.
Past, present and future of developmental biology
Many say that taking the Embryology course changes the life of its students. In fact, right there some people found the research question or organism they would later base their career on. Others ended up meeting their future postdoc advisor among the faculty. I’ve also read of students who got rid of their fears thanks to the experiences made at Embryology.
Personally, I think taking the Embryology course really changes your life, but in a more subtle way. For one summer, one hundred people with a deep interest in developmental biology meet up. All at different stages of their career, they represent the past, the present and the future of developmental biology. So many diverse expertises, experiences and personalities converge to that lab and those lecture rooms. Even if many of us work in different fields or in different parts of the globe, I’m sure the connections that we established will last long. I’m excited and curious to see the many unexpected paths they will put us on.
You don’t take the Embryology course just to learn, you become part of a community of scientists in love with development. So, like Andres, I’ll make sure not to wear off my embryology t-shirt too much. And like Dominique, I’ll be looking out for people wearing those t-shirts with embryos on the front and embryologists on the back.
IBENS develops advanced and original studies aiming at discovering basic mechanisms and principles that underlie biological processes. Located in the heart of Paris, IBENS hosts 28 research teams organized into four main research axes: Developmental Biology, Neuroscience, Functional Genomics and Ecology & Evolution.
The Developmental Biology section will recruit one or two group leaders at junior and/or senior levels. The Neuroscience section seeks one junior group leader. We are looking for outstanding candidates developing ambitious research programs with the potential to interact with and complement the existing strengths of IBENS.
A poster with full details of the announcement can be found here
A 3-year postdoctoral position is available at the UCL Ear Institute (London, UK). This cell biology project will study the role of protein trafficking in cell polarity, cilium function and cytoskeletal organisation in the inner ear.
The postdoc will use imaging (super-resolution confocal, time-lapse microscopy, electron microscopy), molecular and biochemical techniques. They will work independently under the guidance of Dan Jagger (@AuditoryNerves), Nico Daudet (@daudet_n) and Andrew Forge. The project involves a collaboration with Mireille Montcouquiol (@PlanarPolarityT; Bordeaux, France).
I had heard legendary stories about the MBL and the Embryology course in the months leading up to it. Stories about the collaborations of minds that wouldn’t normally meet in the same lab, discoveries that could only have occurred at the MBL, it’s what made me apply. But once I was accepted the nervousness kicked in. The other ubiquitous tales are of the incredibly long days and equally long nights in the lab. I’m not a night owl, I’m an eight-hours-a-night sleeper and I was convinced I would be the first and go to bed every night. I worried that I wouldn’t be able to keep up with my classmates-valid, reasonable concerns I initially thought.
Boy was I wrong.
The atmosphere at the Embryology course is electric. I felt like I was buzzing for six weeks straight and didn’t want to, almost couldn’t, leave the lab. To be surrounded by incredible classmates that get just as excited about the idea of an impossible experiment as me, egged on by brilliant, hilarious and endlessly supportive faculty every night? Why would you go to bed?
I remember a night when I absolutely HAD to image primitive streak ingression in the chick embryo with mosaic labelled cells. Andrea Streit and Claudio Stern sat with me and coached me through flipping a paper-thin stage-three chick embryo and poking it delicately with a dye loaded needle before imaging it for 16 hours. I can’t describe how excited I was waking up at 8am to turn off the microscope before lecture. Only to miss breakfast because I was watching my movie over and over again as I watched labelled cells stream into the embryo. It’s also when I realized how amazing my classmates were. That morning, I walked bleary eyed into lecture and was promptly handed a bagel and a steaming cup of coffee.
I learned to trust my hands during the Embryology course. I had never dissected anything before and I found that I love it. I dissected anything I could get my hands on throughout the summer, culminating in a few of us dissecting ovaries out of every arthropod species we had on hand. We imaged structures that had only been described in the early parts of the 20th century.
Vanessa cardui egg chamber stained with DAPI and Phalloidin
It’s also where I learned to be fearless. The lack of adverse consequences and the adventurous atmosphere meant that I got sea urchin embryos drunk to see how it affected their skeleton, I transplanted cells into zebrafish embryos and I tried to image tardigrade gastrulation. We used Walmart items and state-of-the-art microscopes in the same experiments! I can confidently say that 75% of everything I tried failed, but that was part of the fun. Our class motto ended up being along of the lines of “in DAPI we trust”.
The course tested my intellectual endurance like nothing has before. There is NO pressure to produce or perform, no judgement, no deadlines. You have to find your own drive to learn and try and fail and try and fail and sometimes succeed. Thankfully, its not hard to find that drive. At least six lectures a week from the best minds in the field, who then join you in lab and encourage you to answer impossible unanswered questions. Faculty that are just as willing to give you moral support at the microscope as they are to come have a drink with you at the local pub. Faculty that agree to an hour-long “sweatbox” Q&A session after a two hour talk so we can pepper them with every question under the sun. How can you resist?
Every time it got too hard or too overwhelming, there was always someone to get pie with at Pie in the Sky, always someone willing to take a quick dip into the ocean and always someone willing to “hunt for lightning bug embryos” on a walk by the ocean. The science makes the Embryology course amazing and enriching, the people make it incredible and special. Last summer I learned what I wanted to do and be as a scientist but I also made friends for life #embryology2018.
We enacted convergent extension during the Woods Hole 4th of July parade.
The Santa Cruz Developmental Biology Conference happens every couple of years in UCSC’s beautiful campus, and is seen as a kind of West Coast Gordon Conference for the field. In August a hundred or so developmental biologists gathered for the 2018 version, organised by Bin Chen, Natasza Kurpios and Ed Munro.
It was a first trip to California for one of us (Aidan, who came representing the Node and Development), and a short trip from the office for another (Jordan, whoseUCSC lab works on gene regulation and evolution in C. elegans and parasitic nematodes). Aidan got a chance to visit the town of Santa Cruz before the conference – Blue Oyster Cult were playing on the beach, the boardwalk was full of families, pelicans were diving, sea otters and seals cavorting, all as a bank of freezing fog rolled in – quite a scene! He also got a chance to explore the campus with its giant redwoods, roving turkeys and repeatedly breath-taking view over Monterey Bay.
The conference’s tagline, ‘Old and new: modern answers to enduring questions in development’, was introduced beautifully by the opening three keynotes. Judith Kimble gave a fantastic career retrospective on her quest to understand how embryos get in and out of totipotent states. She described the 27 year search for Notch targets that maintained a naive stem cell fate, and how it lead them to two redundant low complexity proteins with no domains and no clear homologs in other species. This lack of conservation or functional domains dissuades many of us from working on such factors, but the importance of such proteins in development was a theme of the meeting, popping up in several talks.
Eric Wieschaus explained how genetically determined cell fates are translated into local cell behaviors, discussing how myosin tension is a regulator of morphogenetic sequences. He also nicely touched upon his scientific philosophy of reducing his work to the simplest model, seeing what it lacked, and then refining his model. Good lesson for trainees!
Vilaiwan Fernandes, who has just startedher own lab in UCL with a Wellcome Trust Sir Henry Dale Fellowship, won the SCDB Young Investigator Award. Her award lecture covered her time as a postdoc in Claude Desplan’s lab, where she worked on the establishment of retinotopy in the fly brain. She found that glia provide a key link between photoreceptors in the retina and target cells of the optic lobe. The differentiation of both types of cells needs to be tightly coordinated, and Vilaiwan found that glia are crucial to this process by forming a signalling relay, in part using insulin. Aidan got to interview Vil over a beer at a poster session – you can hear all about her research career and plans for the future in the associated interview.
~
The next two days provided repeated demonstrations of why developmental biology is such an exciting field at the moment, as new tools unlock fundamental questions that have kept the field busy for decades. There were so many amazing talks, and we will cover select ones to give readers a flavor of the meeting.
Dominique Bergmann introduced the problem of how cells and tissues establish polarity. Plants, which evolved multicellularity independently of animals, do things quite differently, not just in terms of the components involved but also in terms of how processes like symmetry breaking are achieved. She shared wonderful, long-term videos of asymmetric divisions in the plant stomatal lineage, and explored the reasons why can the establishment of polarity can take hours in plants compared minutes in something like a C. elegans embryo. This talk was another place in which small proteins of low complexity, no domains or conservation played a starring role.
Sally Horne-Badovinac asked how cells coordinate their migration with one another when moving collectively as a sheet. Follicle cells of the Drosophila ovary migrate around the growing egg chamber using basement membrane as a substrate, and this migration leads to the rotation of the egg chambers in a process thought to promote their elongation (it’s just such a bonkers way of sculpting a tissue, evolution really getting creative). Sally showed that the coordinated migration of follicle cells is regulated by the same molecules used in axon guidance, an example of how neurons and epithelia are closer to one another than we might usually think.
Shuonan He, a graduate student in Matt Gibson’s lab who also won the People’s Choice Award for his terrific poster, addressed the role of Hox genes in Cnidaria, which lack a true A-P axis and therefore provide a bit of a conundrum for those of us who think about Hox genes only in relation to A-P patterning. To tackle this problem, Shuonan utilised shRNAs knockdown and CRISPR/Cas9 knockouts to test Hox functional roles, and, with beautiful pictures of Nematostella cross sections, showed that Hox genes control tissue segmentation and tentacle patterning. It was a wonderful evo-devo story, hastened by the use of modern gene editing tools.
Taking a break from the multicellular, Laura Landweber took us somewhere truly strange – the astounding genome rearrangements that occur in the ciliate Oxytricha trifallax during its sexual reproduction. Oxytrichia has two genomes, one of which comprises over 16,000 chromosomes, most of which encodes a single gene; this ‘macronuclear’ genome has to be made anew from a micronuclear precursor during sexual reproduction, in a process of lncRNA-regulated genomic acrobatics that made many heads spin in the audience. It’s an incredibly powerful system to investigate genome fidelity across generations, and it was wonderful to hear about this in a development meeting.
Another major theme of the meeting was the power of single molecule approaches. Jeff Farrell and Sean Megason both presented their single cell RNA-seq (scRNA-seq) approaches to map zebrafish developmental trajectory. Dozens of distinct cell populations could be found by 12 hours of development. This work promises to answer major questions in developmental biology, such as how do cell states change over time and what paths can cells take through development?
Alejandro Sanchez-Alvarado identified the elusive planarian neoblast — the stem cell that allows for the amazing regenerative capacity of this animal — using scRNA-seq. Stunningly, a single neoblast could rescue viability and restore all cells in a lethally irradiated animals, which would otherwise dissolve into goop. Fun fact he shared: the strain of planarian they work on has lost sexual reproduction through a single translocation and now reproduce by anchoring their tail, crawling away until a fragment of their tail rips off, after which both halves generate entire new animals. Kristy Red-Horse used scRNA-seq to understand cell dynamics during coronary development, revealing that there is a gradual, overlapping transition of veins to arteries. Long Cai described sequential fluorescence in situ hybridization (seqFISH), which allowed single cell, single molecule detection of hundreds of transcription factor mRNAs or over 10,000 genomic loci by intron seqFISH.
There were also numerous talks using other powerful new techniques. Ari Pani explored Wnt signaling in C. elegans, providing evidence that a particular Wnt was using a signaling gradient, not a contact-based mechanism such as cytonemes. He used a clever MorphoTrap to disrupt the gradient without altering receptor or ligand levels and found Wnt diffusion was essential for neuroblast migration. Maria Barna used cutting edge mass spec to explore ribosome diversity, providing evidence for ribosomal proteins that associate only with a subset of ribosomes and allow translation of specific mRNAs
Like potato chips in sandwiches, the final keynote pairing of a talks on the morphogenetic Hox clock and evolution of forest mouse tail length was unexpectedly delicious. Denis Duboule gave an energetic, engaging broad overview of his work on Hox gene topologically associating domains. He also discussed the curious case of Hoxd13, which acts as a dominant negative to inhibit the other Hox proteins and seems to “mark the end of things, like digits”. Hopi Hoekstra’s keynote to close out the meeting started off exploring difference in forest mouse morphology, first observed by Osgood in 1909. Forest mice had longer tails due to both longer and more vertebrae, and these long tails were important for balance in lab tests. QTL mapping found three QTLs for length of longest vertebra, three for number of vertebra. And circling back beautifully to the first talk, these QTLs involved the Hox genes Duboule discussed in his talk. This session was a beautiful example of how the mechanisms we discover in the lab function in nature.
~
On the final night we had dinner outside and a final chance to discuss old questions and new techniques under the redwoods, those giants that miraculously, like all of us, started life as a single, tiny cell. The fog rolled in from the Pacific to envelop campus and left some of us lost in search of the bar, but we got there in the end (thanks to the student volunteers for supplying the drinks and acting as deeply knowledgeable beer sommeliers!).
The next Santa Cruz meeting is scheduled for 2020 – we would urge anyone keen on a smallish, diverse and relaxed development meeting in a stunning location to apply.
Day 3 #SCDB2018 and I get to kick off the session under the perpetual vigilance of @ucsc Sequoia sempervirens, the sole living species of the genus Sequoia in the cypress family Cupressaceae. @ScienceStowerspic.twitter.com/SMyP16eI6M
At the meeting I was lucky enough to interview three amazing scientists, one of whom was Vilaiwan Fernandes, winner of the SCDB Young Investigator Award for her outstanding research achievements, who has just started her own lab in UCL. (Look out for interviews with Judith Kimble and Cassandra Extavour in future issues of Development.) I sat with Vilaiwan in a noisy and beery evening poster session and chatted about her career so far and plans for her new lab.
When did you first get in to science?
I grew up in Bombay (now Mumbai) in India, and I guess I was interested in science in school because it involved less memorisation than other subjects. I learned to scuba dive, which got me interested in conservation biology and I began volunteering with an organisation that held public education snorkelling camps. I tried to continue that when my family emigrated to Canada, and studied marine biology in my undergraduate. There’s a marine station on the west coast of Vancouver Island called Bamfield Marine Sciences Center, and I spent a lot of time there. Every summer in my undergraduate I would either do a course there or work as a research assistant on some project or other. There’s lots of amazing intertidal life there, particularly invertebrates, and we were lucky enough to explore tiny uninhabited islands and set up quadrats to see what we could find!
How do you go from that into something resembling developmental biology?
I was never interested in cell biology or development, because I liked being outdoors. At the end of my undergraduate I had a summer job but started panicking because I had no plan for once it was up. So I applied to a ton of random things, including an apprenticeship in Friday Harbor Labs in Washington, called ‘Computational Modelling of Biological Networks’. I had also studied math in my undergraduate, and assumed the apprenticeship was about computationally modelling ecological networks, which was wrong: I got there and found it was about gene networks. So that’s sort of how I got pulled in to the field – by accident.
I worked with an amazing postdoc, Kerry Kim, to try to understand the evolutionary benefit of being a haploid or a diploid that reproduces either asexually or sexually. We ran genetic algorithms to evolve populations under these different conditions for several generations. It was cool because we were able to represent haploidy and diploidy (and recombination) in a much more realistic way than other models had at the time. It was a purely computational project but a lot of fun and one I’m still very proud of.
And you then transitioned back again from dry to wet biology?
The apprenticeship was based in the NIGMS Center for Cell Dynamics, which was run by Garry Odell. His wife Victoria Foe and other amazing scientists like Ed Munro and George VonDassow also worked there. Garry was brilliant and encouraged me to try my hand at experiments. He had an engineering background and was really interested in size control and scaling. He convinced me to transplant nuclei between different sized Drosophila species just before cellularisation. The goal was to see whether the foreign nuclei would form cells staying true to the size of their own species or if they would switch to the size of the host species. Unfortunately, we didn’t finish this project as I left to start grad school, but it was the best introduction to experiments!
It sounds quite fiddly as an introduction to experimental work!
It was, but Victoria, who trained me, has amazing hands and she just taught me to be patient. It made me opt for lab work over computational biology in grad school, and I applied to McGill and joined Ehab Abouheif’s lab. Quebec didn’t suit me for a number of reasons and I ended up dropping out of that PhD programme and moving back to the West coast, where I re-enrolled in a grad student in a fly lab – Esther Verheyen’s in Vancouver.
My initial project with Esther was to work on Nemo, a rather promiscuous protein kinase. Thankfully, Esther convinced me to do a screen for morphogenetic furrow progression defects in the eye disc. That’s where we pulled out a kinase called Misshapen, which stood out because it sped up furrow progression in contrast to all our other hits. At that time, Sally Horne-Badovinac’s lab had just published Misshapen’s regulation of integrins in the follicle cells. I met her postdoc Lindsay Lewellyn at a conference, and following that we decided to test the role of integrins in the eye. Lindsay was super cool, and ended up on my paper! I guess the main point of that paper was to link the signalling pathways that affect transcription in the eye with the non-transcriptional, post-translational events, like cytoskeletal rearrangements and changes in adhesion, which are important in the morphogenetic furrow.
And then you go to a New York for your postdoc with Claude Desplan – I guess by that point you’d fallen for eye development?
Not quite – but every time I say I’m not going to do something, I end up doing that thing. I thought wanted to switch models and move away from neurobiology, but then I met Claude at a Gordon Conference and he suggested that I apply to his lab. The atmosphere in his lab was great, and I was lucky to get accepted. Claude is always, always excited about science and I could not have been in a better place.
My project was initially to try to understand how to make individual cell fates in the developing visual system, in particular how to generate cellular diversity in the lamina. When I started working on it we realised we didn’t actually know why neurons differentiate in the particular pattern that they do, and it seemed that this pattern was key to why they adopted different fates. Seeing how well glial cell processes entering the lamina correlated with the front of neuronal differentiation made us suspect glial involvement. You can read about where that story took us in my Science paper from last year.
So the next step was to look for jobs – you’d been a postdoc for a relatively short time, so what made you think about being a PI?
I had a lot of leads that I really wanted to follow up on, but felt that I wouldn’t be able to do all of it myself. I’m very excited to pursue all of them and see where they go. And Claude was very supportive.
When I applied for the Wellcome Trust Henry Dale Fellowships and got selected for an interview, UCL was very good at organising mock interviews – they ended up being more terrifying than the actual interviews! My very first mock interview was a complete disaster, and I cried for several hours afterwards, but I’m told that’s normal and I suppose it did the trick.
What are your plans for the initial years of your lab? Are you going to stick with glia?
Perhaps even more than glial involvement I want to focus more broadly on exogenous cues that affect neurons – the field has been very focused on neurons talking to neurons, and neuroblasts being intrinsically patterned, without really looking too much at exogenous cues. For now, I’m focusing on signals from glia.
In the long-term, one of the larger projects will use single cell sequencing of glia during development, with the goal of identifying secreted and membrane-associated factors that affect neuronal development. In theory, this approach should identify instances like the one we showed in our paper, where glia express insulins to induce lamina neuronal differentiation.
Finally, congratulations on your SCDB Young Investigator Award! How did you feel when you found out you’d won it?
It really was a huge honour to receive the award – I was very surprised and didn’t expect it!
At the very end of my talk I thanked the couple Garry [Odell] and Victoria [Foe] with whom I had started my research career in Washington. Garry died earlier this year, but I know he would have been really proud of me for this. I visited him in hospital before I went to my Henry Dale interview. He was adorable – he was there sick in bed but said ‘We have to practice your interview, get your slides out!’ He and Victoria had a can-do attitude in the lab – looking back at those early days I think I we were crazy to think we could transplant nuclei between species! But at the time, I didn’t think twice as to whether it was possible or hard. Gary was great for that, and was such a strong influence on my life and my science.

 (No Ratings Yet)
(No Ratings Yet)


 (4 votes)
(4 votes)