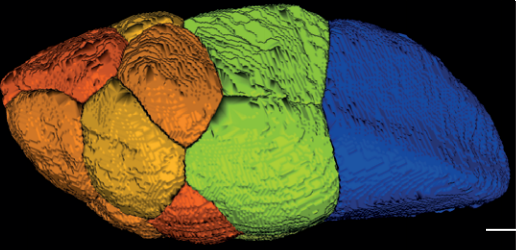
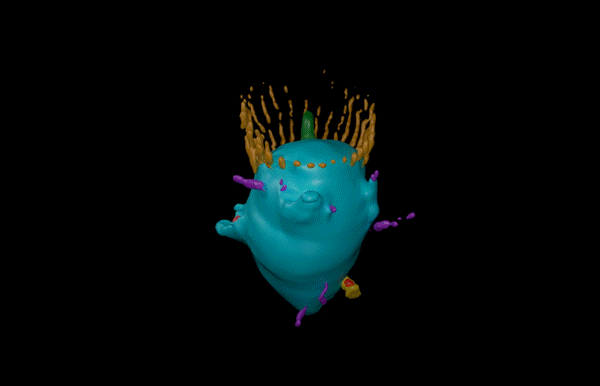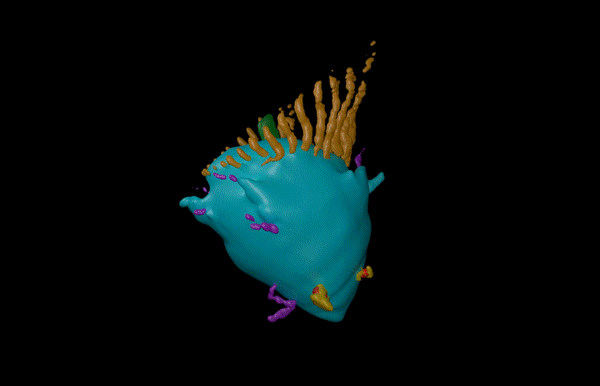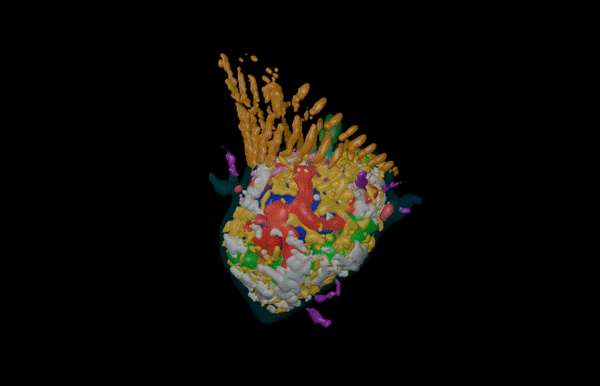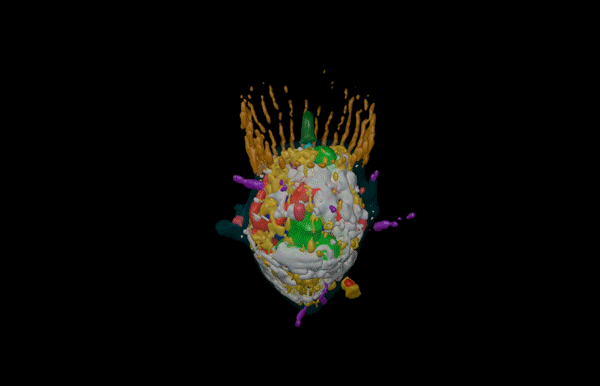
Hatchling catshark CT scan, scanned by Kyle Martin and Amin Garbout at The Imaging and Analysis Centre, NHM, London
Understanding how complex biological patterns arise is a long standing and fascinating area of scientific research. The patterning, or spatial arrangement, of vertebrate skin appendages (such as feathers, hair and scales) has enabled diverse adaptations, allowing animals to both survive and thrive in varied and challenging environments. Such adaptations include temperature control of mammalian hair1 and drag reduction of shark scales2. In our recent paper published in Science Advances, we examine how such patterning is regulated throughout development of the shark3.
Alan Turing was a remarkably influential scientist, renowned for his pivotal role as a code breaker during the Second World War. He is also considered the forefather of the modern computer. In 1952, two years before his tragic death following prosecution under the homophobic law of ‘Gross Indecency’, Turing wrote a mathematical model describing how interactions between diffusing chemicals known as morphogens can produce patterns4,5. These morphogens include a short-range, self-promoting activator which also promotes a long-range inhibitor. Turing demonstrated that when his model is appropriately tuned, stable patterns can arise.
Since its publication, there has been a surge of theoretical and experimental research discussing Turing’s model. In fact, his seminal paper has now been cited almost 12000 times4, and experimental work has suggested that this system controls the patterning of both mouse hair and bird feathers6,7. These animals – which belong to a group of vertebrates known as tetrapods – are both classic model species for studying developmental biology. However, until recently, the role of Turing’s system in the skin appendage patterning of vertebrates that arose prior to tetrapods has been poorly understood.
Hatchling catshark denticle SEM – Rory Cooper
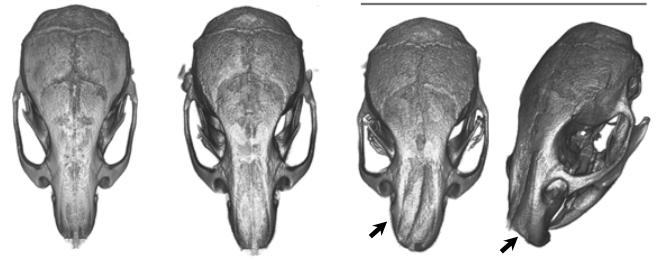
Sharks belong to a group of vertebrates known as cartilaginous fishes, which branched from most other jawed vertebrates before tetrapods diversified. They possess scale-like skin appendages known as denticles, which have been observed in the fossil record as long as 450 million years ago8. Denticles are made from dentine and enamel-like materials, similar to our own teeth. In our recent study, we demonstrate that Turing’s patterning system can explain the arrangement of denticles in an emerging model shark3 – the small-spotted catshark (Scyliorhinus canicula).
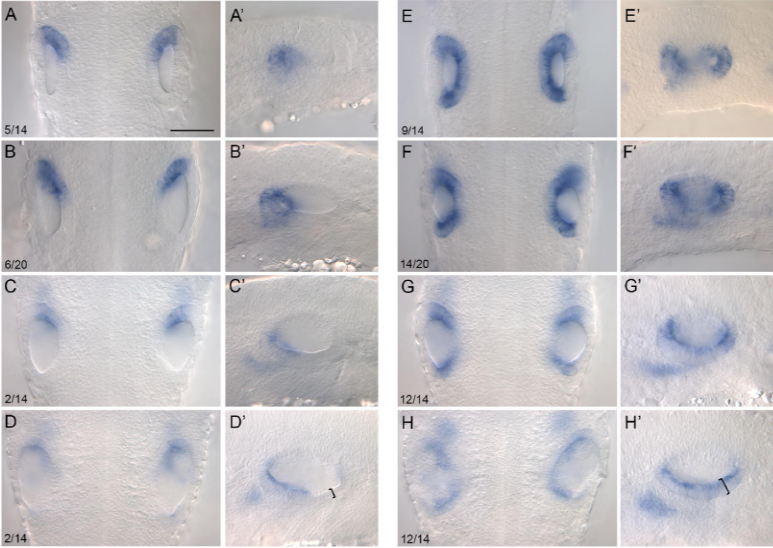
We first used computer modelling based on Turing’s equations to demonstrate that denticle patterning is consistent with his system. Next, we examined genes that act as morphogens during the Turing patterning of feathers, and revealed their expression is conserved throughout shark denticle patterning. This includes sonic hedgehog (shh) and fibroblast growth factors (FGFs) as activators, and bone morphogenetic proteins (BMPs) as inhibitors7. By inhibiting FGF signalling, we showed there is likely functional evolutionary conservation of these genes, meaning they appear to play the same roles in both feather and denticle development.
Clear and stained catshark denticles
Having found this evidence for Turing-like patterning of shark denticles, we went on to show that altering the parameters of the model can produce patterns with different densities of coverage. These patterns were reflective of arrangements observed in species of sharks and rays that are alive today, demonstrating that simple alterations to Turing’s model can explain the diversity of patterns observed in nature. Therefore, we provide a potential mechanism for how important adaptations, such as hydrodynamic drag reduction and defensive armour, have arisen in sharks. It is likely that Turing’s model is of widespread importance throughout the skin organ patterning of diverse vertebrate groups, all the way from sharks to mammals6,7.
The pattern of shark denticles is one important factor in achieving drag reduction. Another is the shape of individual denticles, which vary both within and between different shark species. Our next research goal is to examine the developmental mechanisms underlying the diversity of denticle shape. Together, this research will provide us with an insight into how important functional traits have arisen in the shark.
Catshark embryos (100 days post fertilisation)
Shark-inspired materials have already been created, with the aim of improving the efficiency of travel9. Potential applications include aeroplanes, boats and cars, which are all subject to drag, that is, resistance to the forward motion of an object. Understanding how both denticle patterning and shape contribute towards reducing drag may help to improve the effectiveness of these materials, leading to reduced energy consumption in a wide range of industries. This is an ever-important challenge in a time of depleting resources and climate crisis.
References
Ruxton, G. D. & Wilkinson, D. M. Avoidance of overheating and selection for both hair loss and bipedality in hominins. Proc. Natl. Acad. Sci.108, 20965–20969 (2011).
Dean, B. & Bhushan, B. Shark-skin surfaces for fluid-drag reduction in turbulent flow: a review. Philos. Trans. A. Math. Phys. Eng. Sci.368, 4775–806 (2010).
Cooper, R. L. et al. An ancient Turing-like patterning mechanism regulates skin denticle development in sharks. Sci. Adv. 4, 11, eaau5484 (2018).
Turing, A. M. The chemical basis of morphogenesis. Philos. Trans. R. Soc. Lond. B. Biol. Sci.237, 37–72 (1952).
Kondo, S. & Miura, T. Reaction-Diffusion Model as a Framework of Understanding Biological Pattern Formation. Science329, 1616–1620 (2010).
Sick, S., Reinker, S., Timmer, J. & Schlake, T. WNT and DKK Determine Hair Follicle Spacing Through a Reaction-Diffusion Mechanism. Science314, 1447–1450 (2006).
Jung, H. et al. Local Inhibitory Action of BMPs and Their Relationships with Activators in Feather Formation: Implications for Periodic Patterning. Dev. Biol.196, 11–23 (1998).
Sansom, I. J., Smith, M. M. & Smith, P. Scales of thelodont and shark-like fishes from the Ordovician of Colorado. Nature379, 628–630 (1996).
Domel, A. G. et al. Shark skin-inspired designs that improve aerodynamic performance. J. R. Soc. Interface15, (2018).
Two BBSRC/Wellcome Trust-funded positions at the University of Sheffield are available for candidates with a background in (i) cell and/or developmental biology and (ii) mathematical modelling to join an interdisciplinary team investigating the coordination of tissue patterning and growth using Drosophilaepithelial development as a model system.
The Strutt lab (http://www.shef.ac.uk/bms/research/strutt) is a world leader in studying the planar polarity signalling pathways that control coordinated cell polarisation in animal tissues. In this collaborative project with Dr Alexander Fletcher (http://alex-fletcher.staff.shef.ac.uk) we are now combining our biological expertise with mathematical modelling approaches, to build an integrated understanding of how tissue patterning and growth are coordinated to achieve consistent organ shape and size during animal development.
Informal enquiries may be directed to David Strutt (d.strutt@sheffield.ac.uk) or Alexander Fletcher (a.g.fletcher@sheffield.ac.uk). Formal applications should be made directly to the University of Sheffield (http://www.sheffield.ac.uk/jobs, Job Refs: UOS020789 and UOS020791) by no later than 4thDecember 2018.
Recent relevant publications:
Kursawe J, Baker RE, Fletcher AG (2018). Approximate Bayesian computation reveals the importance of repeated measurements for parameterising cell-based models of growing tissues. J Theor Biol. 443:66-81
Fisher KH, Strutt D, Fletcher AG (2017). Integrating planar polarity and tissue mechanics in computational models of epithelial morphogenesis. Curr Opin Sys Biol 5: 41-49.
Strutt H, Gamage J, Strutt D (2016). Robust asymmetric localization of planar polarity proteins is associated with organization into signalosome-like domains of variable stoichiometry. Cell Rep 17:2660-2671.
Hale R, Brittle A, Fisher KH, Monk NA, Strutt D (2015). Cellular interpretation of the long-range gradient of Four-jointed activity in the Drosophila wing. eLife 4:e05789.
Wells RE, Barry JD, Warrington SJ, Cuhlmann, S, Evans P, Huber W, Strutt D, Zeidler MP (2013). Control of tissue morphology by Fasciclin III-mediated intercellular adhesion. Development 140:3858-3868.
Brittle A, Thomas C, Strutt D (2012). Planar polarity specification through the asymmetric subcellular localisation of the atypical cadherins Fat and Dachsous. Curr Biol 22:907-914.
Every year our Institute (IST Austria) opens its doors to the public during an outreach event called Open Campus. Visitors can participate in demonstrations and guided tours of the labs while scientists explain their research. But how do you show a variety of different activities performed in the lab within a 20 min tour? We wanted to make it possible for people to take a look behind the scenes of week-long experiments in just 3 minutes… so we came up with an idea of a video: a glimpse into our everyday work with cells, embryos and datasets.
Making this type of video requires you to step out of your scientific comfort zone and to consider what the audience will want to see. It is a great way to engage the public in your research and, needless to say, you can finally show your family and friends what ‘being in a lab’ is all about.
After some brainstorming sessions, endless laughs (it is not easy to keep a straight face when you thought you had been recording for 5 minutes and you had forgotten to press the record button), lots of editing and, most importantly, scouring the internet to find an ideal soundtrack… We made a movie!
Since we received a lot of uplifting positive feedback, we would like to share our short movie with The Node Community as well. Welcome to Kicheva Lab and get inspired!
An engineer/technician position is available on the Marseille-Luminy campus to work on tumorigenesis using the chick embryo as a model organism. The candidate will work in the context of a collaborative project between the lab of Dr Cédric Maurange and Dr Marie- Claire Delfini at the Institute for Developmental Biology of Marseille (IBDM).
Funding is provided by the Agence Nationale de la Recherche (ANR) for 12 months.
We are seeking a highly-motivated candidate with some practical experience in molecular biology. A previous experience with chick embryos is desirable but not mandatory as the candidate will have the possibility to be trained.
The engineer/technician will generate DNA constructs to perform gain and loss of function experiments in the chick embryo via electroporation in order to initiate tumorigenesis. She/he will perform immunostainings or in-situ hybridization and use confocal microscopy to document the experiments.
Engineer or technician level will be recognized according to education (Bachelor +2-4 e. g. IUT, BTS, Diploma, Master).
The position is available starting from February/March 2019.
preLights, The Company of Biologist’s new preprint highlighting service, has now been running for more than eight months. At the heart of preLights is the community of early-career researchers who select and highlight interesting preprints in various fields.
As the service is building momentum, we are ready to grow our team of preLighters and are seeking early-career researchers (PhD students, postdocs, early-stage PIs) who are passionate about preprints and enjoy writing and communicating science. We welcome researchers from across the biological sciences and especially those with expertise in Neuroscience, Bioinformatics, Microbiology, Ecology, Plant biology, Biophysics, and Systems Biology.
To join our team of preLighters, please send your application to prelights@biologists.com by 1 December 2018.
In your application, please provide:
A short biography, telling us who you are and what you work on
A few sentences about why you are interested in joining our community
A preLights post highlighting a recent preprint of your choice (that is not older than 1 month)
We have a flexible format for preLights, but your post should aim to include:
A short and engaging ‘Tweetable’ summary of the preprint; background of the preprint; key findings of the preprint; what you like about this preprint/why you think the work is important; future directions and questions for the authors.
The post should reflect your personal opinion on the research in the preprint that you selected. Please also provide the URL link of the preprint. Your post should not exceed 1000 words.
To learn more about the ideas behind preLights, please read this introduction, or check out the interviews with current preLighters on their experience on our News page.
What’s in it for me?
This is a great opportunity for you to gain experience in science writing. You will get editing feedback from us and your peers and we aim to raise your profile as a trusted preprint selector and commentator. You will grow your professional network, and we are happy to support you by offering recommendation letters or in other ways.
But there is also a commitment; we expect you to select and highlight a preprint every one-or-two months.
We are aiming to have a diverse team and might not be able to accept all applicants, but are looking forward to welcoming our new preLighters.
Humankind has been researching and engineering for as long as we have existed. It was a matter of survival back then and it is still is nowadays. This long and involved process that spanned over several millennia has enabled civilisations to rise and fall. Thousands of years of science and scholarly traditions have led to the accumulation of an incommensurable amount of knowledge spread across various disciplines including mathematics, physics, chemistry and biology.
What is translational science?
The latin word “Scientia”means knowledge, but it is only recently that the concept of “translational science” has emerged.To understand the essence of this neologism the expression needs to be broken down into two definitions “translation” and “science”.
The mathematical definition of translation corresponds in geometry, to the process of moving something from one place to another. The second word science is defined as follows: the intellectual and practical activity encompassing the systematic study of the structure and behavior of the physical and natural world through observation and experiment.
The combination of these two definitions leads to the concept of translational science, which is the translation of fundamental scienceinto practical applications. A famous illustration of this is the serendipitous discovery of penicillin in 1928 by Alexander Fleming which broke open a new field in modern medicine.In other words, fundamental science is the engine that powers translational research. If for any particular reason, the engine stop running, then there is nothing left to be translated.
In order for science to be developed in an unbiased way it needs to be performed free from any interest, otherwise conflict arise and findings tend to matches expectation and not observations. This point is crucial and clearly is a roadblock for translational science which, by definition is developed to be applied in order to generate a useful application and potential profit.
Science is unique in the sense that it is not made to deliver a product; it is designed purely to generate knowledge. Obviously there are major directions in science, but there is not a pre-determined end point. Instead, each discovery leads to the next one and adds to our understanding of the world in which we live. It is an endless process that is really often convoluted. Taking advantage of a particular discovery to make an invention that will be useful in a specific context is a different process that cannot be assigned to fundamental science. It corresponds to an engineer perspective where a technical issue is solved by a technological advance. Research directions in fundamental science have to remain limitless, otherwise, the scope of discoveries, and therefore the range of potential applications, would be limited to a predefined scientific horizon.
A recent example of this, is the discovery of molecular scissors known as “crisp/rcas9” which is currently revolutionising biological and medical research. It is now possible to edit the genome of a living organism without complex procedure, this has opened up new research avenues and therapeutic options. Such a discovery was originally made by scientists working on understanding the basic molecular mechanism driving of viral infection in bacteria.
For practical and ethical reasons it is not sustainable for academic science to get engaged in products development.The lack of funding in academia for one part and the industry commitment and better ability to develop translational science leaves no doubt about role distribution.
Where things become blurry is that there is no clear demarcation defining where fundamental science does stop and where translational science is starting. There is not even aclear definition of what translational science is, this notion can vary between research fields. It is just a vague concept that is being abused since in essence every fundamental discovery is potentially translational, but in reality only a really low percentage will become translated. This confusion is mostly due to the time scale difference. While academia establishes project planned over a decade translational research projects span over a shorter period of time (a few years).
Why has translational science been so successful over the past few decades?
Translational science draws on the large amount of knowledge that has accumulated over the past century. Sadly, this wealth of basic findings is not endless. The accumulation of knowledge generated by fundamental science has suddenly started to down-size due to a major shift into translational science activity that mostly feeds on previous ground-breaking discoveries, but that does not generate any novel fundamental findings itself. Translational science owes its success in part to a high level of attention from the media. This has contributed to draw support from the public arena but this push by the media in a desperate search for a scientific buzz also comes with a risk. In fact there is a real threat for scientists of potentially losing credit in the long term if the community fails to deliver.
How does the system remain sustainable in the long-term?
It is critical to maintain the right balance between fundamental science, which constitutes the foundation of any progress, and translational science, which converts a discovery into a useful application. This equilibrium is hard to maintain simply because the rate by which these two sciences evolve are dissimilar. In the case of basic science, significant advances are relatively slow mostly due to the fact that science relies on serendipity and scientific wandering. Some unexpected paths have to be explored over decades to enable a ground-breaking discovery. Failure is an essential part of the discovery process. Further, basic science is often limited by available technology. Ancient concepts are revisited regularly due to the development of new technology that enables us to probe fundamental mechanisms in more depth.
By essence, observation and experimentation are slow processes that rely more and more on complex research tools. Since science is becoming increasingly specialised and dependent on cutting-edge technologies, the discovery process is becoming increasingly more challenging. For instance, one of the bottlenecks of modern science is managing the huge datasets generated by genomic research. In that particular case, the physiological interpretation of the data is one of the limiting step. For this particular reason, it will take time to bridge the gap between genomics approaches and personalised medicine for instance.
By contrast, translational science is evolving at a rapid pace, since it is being determined by a specific endpoint, and its proof of principle, feasibility and viability have been already established by fundamental science.
Where to draw the line between fundamental science and translational science?
This discrepancy has been masked until now by the fact that a lot of knowledge has been accumulated in fundamental science and translational science could draw from this gigantic gold mine. However shortages in option start to arise in particular industries, since basic knowledge is running out. For instance in the case of drug discovery, conventional molecular targets have been over-exploited and pharmaceutical industries and academia have fallen short in discovering new molecular mechanisms that would lead to alternative therapeutic avenues.
National research agencies are pushing hard to encourage translational science, but the way it is being developed is not optimum. Funding bodies are trying to impose a shift of fundamental science into translational science, instead of promoting more bridging strategies that would enable academics and industries to work in a complementary fashion. At the international level with the merciless competition for commercialisation, this strategic choice could cost even more than not investing into fundamental science. At the end of the day, any novel drug of technology that reaches the market will be used on a global scale, and the price of buying its patent will cost a lot more than the initial amount of money that would have been necessary to discover its principle.Governments and other funders must recognise the importance of having a thriving base of fundamental knowledge from which to translate, for both economic and health reasons.
Last but not least, an essential aspect of fundamental science is often forgotten, its main function, which is to generate knowledge. There is no direct dollar value for knowledge and expertise, however one of the industries directly benefiting from this output is the education sector. Translating knowledge into the education system is far more valuable in the long term than any drug that is being commercialised, and it is pretty daunting to envision a future where the engine of human progress would fall into decay.
Welcome to our monthly trawl for developmental biology (and related) preprints.
This month we decided to reinstate our Plant Development section after a Twitter chat, and as it happened October had a glut of preprints covering all aspects of plant development. You’ll also find lots of regeneration (kidneys, colons, eyes, axons and whole colonial tunicates), a typically diverse Evo-devo zoo, and a feast for fans of cell division in our Cell Biology section.
The preprints were hosted on bioRxiv, PeerJ, andarXiv. Let us know if we missed anything, and use these links to get to the section you want:
Surface tension determines tissue shape and growth kinetics
Sebastian Ehrig, Cécile M. Bidan, Alan West, Cornelius Jacobi, Karen Lam, Philip Kollmannsberger, Ansgar Petersen, Pavel Tomancak, Krishna Kommareddy, Franz Dieter Fischer, Peter Fratzl, John W. C. Dunlop
A segmented fly egg chamber, from Lamire, et al.’s preprint
Single-cell RNA-seq reveals that glioblastoma recapitulates normal brain development
Charles P. Couturier, Shamini Ayyadhury, Phuong U. Le, Jean Monlong, Gabriele Riva, Redouane Allache, Salma Baig, Xiaohua Yan, Mathieu Bourgey, Changseok Lee, Yu Chang David Wang, V. Wee Yong, Marie-Christine Guiot, Bratislav Misic, Jack Antel, Guillaume Bourque, Jiannis Ragoussis, Kevin Petrecca
Single-cell in situ transcriptomic map of astrocyte cortical layer diversity
Omer Ali Bayraktar, Theresa Bartels, Damon Polioudakis, Staffan Holmqvist, Lucile Ben Haim, Adam M.H. Young, Kirti Prakash, Alexander Brown, Mercedes F. Paredes, Riki Kawaguchi, John Stockley, Khalida Sabeur, Sandra M. Chang, Eric Huang, Peter Hutchinson, Erik M. Ullian, Daniel H. Geschwind, Giovanni Coppola, David H. Rowitch
CRISPR-Cas9 Screens Reveal Genes Regulating a G0-like State in Human Neural Progenitors
Heather Feldman, Chad Toledo, Sonali Arora, Pia Hoellerbauer, Philip Corrin, Lucas Carter, Megan Kufeld, Hamid Bolouri, Ryan Basom, Jeffrey Delrow, Joshua Meier, Feng Zhang, José McFaline-Figueroa, Cole Trapnell, Steven Pollard, Christopher Plaisier, PATRICK PADDISON
RNA-Seq in 296 phased trios provides a high resolution map of genomic imprinting
Bharati Jadhav, Ramin Monajemi, Kristina K. Gagalova, Daniel Ho, Harmen H.M. Draisma, Mark A. van de Wiel, Lude Franke, Bastiaan T. Heijmans, Joyce van Meurs, Rick Jansen, GoNL Consortium, BIOS Consortium, Peter A.C. ʼt Hoen, Andrew J. Sharp, Szymon M. Kiełbasa
Aged-senescent cells contribute to impaired heart regeneration
Fiona C Lewis-McDougall, Prashant J Ruchaya, Eva Domenjo-Vila, Tze Shin Teoh, Larissa Prata, Beverley J Cottle, James E Clark, Prakash P Punjabi, Wael Awad, Daniele Torella, Tamara Tchkonia, James L Kirkland, Georgina M Ellison-Hughes
PP4-dependent HDAC3 dephosphorylation discriminates between axonal regeneration and regenerative failure
Arnau Hervera, Luming Zhou, Ilaria Palmisano, Eilidh McLachlan, Guiping Kong, Thomas Haynes Hutson, Matt C Danzi, Vance P Lemmon, John L Bixby, Andreu Matamoros-Angles, Kirsi Forsberg, Francesco De Virgiliis, Dina P Matheos, Janine Kwapis, Marcelo A Wood, Radhika Puttagunta, Jose Antonio Del Rio, Simone Di Giovanni
Murine MPDZ-Linked Hydrocephalus is Caused by Hyperpermeability of the Choroid Plexus
Junning Yang, Claire Simonneau, Robert Kilker, Laura Oakley, Matthew Byrne, Zuzana Nichtova, Ioana Stefanescu, Fnu Pardeep-Kumar, Sushil Tripathi, Eric Londin, Pascale Saugier-Veber, Belinda Willard, Mathew Thakur, Stephen Pickup, Richard Smeyne, Arie Horowitz
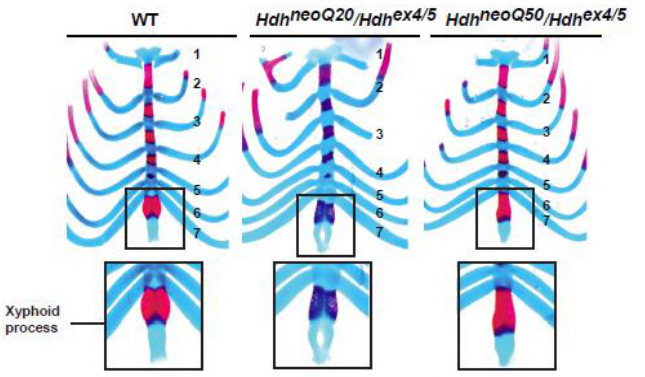
Hypomorphic mutation of the mouse Huntington’s disease gene orthologue
Vidya Murthy, Toma Tebaldi, Toshimi Yoshida, Serkan Erdin, Teresa Calzonetti, Ravi Vijayvargia, Takshashila Tripathi, Emanuela Kerschbamer, Ihn Sik Seong, Alessandro Quattrone, Michael E. Talkowski, James F. Gusella, Katia Georgopoulos, Marcy E. MacDonald, Marta Biagioli
Functional dissection of the ARGONAUTE7 promoter
J Steen Hoyer, Jose L Pruneda-Paz, Ghislain Breton, Mariah A Hassert, Emily E Holcomb, Halley Fowler, Kaylyn M Bauer, Jacob Mreen, Steve A Kay, James C Carrington
Root and shoot apical meristems from Vernoux, et al.’s preprint
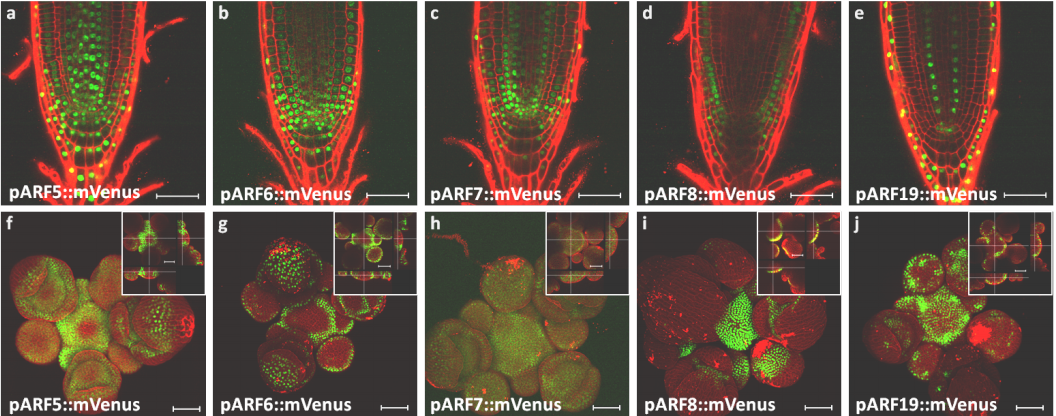
A network of transcriptional repressors mediates auxin response specificity
Teva Vernoux, Jekaterina Truskina, Jingyi Han, Carlos S Galvan-Ampudia, Stéphanie Lainé, Géraldine Brunoud, Silvana Porco, Anne-Maarit Bågman, Margot E Smit, Malcolm Bennett, François Roudier, Siobhan M Brady, Anthony Bishopp
Floral phenotypes in Arabibodpsis, from Taylor, et al.’s preprint
Programmed DNA elimination of germline development genes in songbirds
Cormac M. Kinsella, Francisco J. Ruiz-Ruano, Anne-Marie Dion-Côté, Alexander J. Charles, Toni I. Gossmann, Josefa Cabrero, Dennis Kappei, Nicola Hemmings, Mirre J. P. Simons, Juan P. M. Camacho, Wolfgang Forstmeier, Alexander Suh
The whale shark genome reveals how genomic and physiological properties scale with body size
Seung Gu Park, Victor Luria, Jessica A. Weber, Sungwon Jeon, Hak-Min Kim, Yeonsu Jeon, Youngjune Bhak, Jehun Jun, Sang Wha Kim, Won Hee Hong, Semin Lee, Yun Sung Cho, Amir Karger, John W. Cain, Andrea Manica, Soonok Kim, Jae-Hoon Kim, Jeremy S. Edwards, Jong Bhak, George M. Church
NanoJ: a high-performance open-source super-resolution microscopy toolbox
Romain Laine, Kalina Tosheva, Nils Gustafsson, Robert D. M. Gray, Pedro Almada, David Albrecht, Gabriel T. Risa, Fredrik Hurtig, Ann-Christin Lindås, Buzz Baum, Jason Mercer, Christophe Leterrier, Pedro M. Pereira, Siân Culley, Ricardo Henriques
Robust and sensitive GFP-based cGMP sensor for real time imaging in intact Caenorhabditis elegans
Sarah Woldemariam, Jatin Nagpal, Joy Li, Martin Schneider Schneider, Raakhee Shankar, Mary Futey, Aruna Varshney, Kristine Andersen, Benjamin Barsi-Rhyne, Alan Tran, Wagner Steuer Costa, Chantal Brueggemann, Scott Hamilton, Denise Ferkey, Miri VanHoven, Alexander Gottschalk, Noelle L’Etoile
We are looking for: Enthusiastic researchers with a BSc or Master Degree in biomedical sciences with interest in Developmental Neurobiology. Good academic records are required, as well as good spoken and written command of English.
We offer: A highly multidisciplinary and competitive training programme in biomedical research. Access to state-of-the-art infrastructures.
The selected candidate will investigate growth of the central nervous system and associated primary microcephaly, with the aim to understand the mechanisms that control cell numbers and brain size at birth. We recently demonstrated that the activity of classical growth factors such as Sonic hedgehog and BMPs is required for the expansion of the pool of neural progenitors by maintaining symmetric divisions. We are now combining high resolution imaging and data from transcriptomics and functional genetics, to describe the mechanisms downstream these growth factors that regulate neural stem cell maintenance.
Those interested please send CV, a cover letter justifying the interest of the applicant in the project, and the names of two referees toemgbmc@ibmb.csic.es
The Royal Society is organising the upcoming Single cell ecology meeting on 10-11 December 2018 in London, UK, on behalf of Professor Thomas Richards, Dr Ramon Massana and Professor Neil Hall.
This will be an interdisciplinary meeting to explore the use of single cell technologies to understand the function, diversity and interactions of microbes. This meeting aims to bring together physicists who manipulate cells; microbiologists who seek to understand the nature of microbial communities; genomicists who are developing new approaches to study individual cells; and evolutionary biologists who are trying to sample microbes and understand where they branch on the tree of life.
This two-day symposium will showcase the best in developmental biology across the life course. From embryogenesis through to ageing, areas of current excitement in the field will be highlighted by plenary talks from 15 internationally renowned speakers, along with selected short talks from abstracts.
The symposium will honour the memory of Rosa Beddington, a leading UK embryologist who was Head of the Division of Mammalian Development at the MRC National Institute for Medical Research from 1993-2001. Many of Rosa’s colleagues and lab alumni will be attending the symposium.
We invite submissions for short talks (15 minutes) from early-career researchers (PhD students, postdocs and recently-established PIs). Please submit your abstracts to events@crick.ac.uk by 1 December 2018. Abstracts should be no more than 500 words including the title, authors and institution information. The presenter for short talks must have registered their attendance to the conference by 1 December 2018. Please ensure the presenter’s name is underlined on your submission.
We look forward to seeing you at the Crick in February,
The Organising Committee (James Briscoe, Alex Gould, Rita Sousa-Nunes and Jean-Paul Vincent)



 (5 votes)
(5 votes)
 (No Ratings Yet)
(No Ratings Yet)