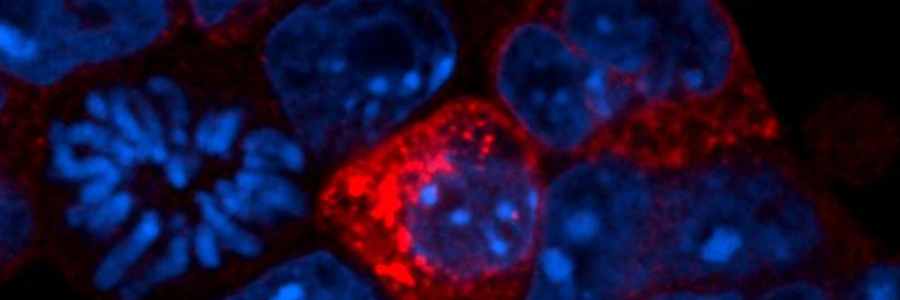
Production of proteins from transposons (red). DNA shown in blue.
Molecules called endosiRNAs help us avoid genetic chaos, according to a new study from a team at the Babraham Institute. Much of the human genome contains pieces of DNA called transposons, a form of genetic parasite. When active, transposons can damage genes so it is important to keep them inactive. At a certain point early in the human life cycle controlling transposons is particularly difficult. This latest research, published in Cell Stem Cell reveals how endosiRNAs keep our genes safe during this vulnerable stage.
Transposons, also called transposable elements, are ancient viruses that have become a permanent part of our genes. Around half of the human genome is made of transposons, many are damaged, but some can become active. Active transposons can be harmful because they move about the genome. When transposons move they can damage genes, leading to genetic illnesses and playing a part in some cancers.
Chemical markers in DNA called methylations can keep transposons inactive. Cells often use methylations to inactivate pieces of DNA, whether they are genes or transposons. Yet, in each new generation most methylations are temporarily erased and renewed by a process called epigenetic reprogramming. This means that, during sperm and egg production there is a short time when methylations do not control transposon activity, leaving them free to damage genes and shuffle DNA.
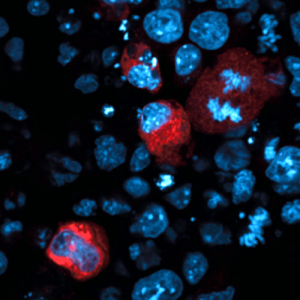
The new findings show that transposons become active when cells erase DNA methylation and they are shut down by the endosiRNA system. Just like active genes, active transposons produce messages in the form of RNA molecules, which have many similarities to DNA. The study reveals that cells can detect these transposon RNA messages and use them to create specific endogenous small interfering RNAs (endosiRNAs). The endosiRNAs then act like a trap allowing a protein called Argonaute2 (Ago2) to seek and destroy transposon messages before they cause any harm.
Speaking about the research, lead author on the paper, Dr Rebecca Berrens, said: “Epigenetic reprogramming plays a vital role in wiping the genome clean at the start of development, but it leaves our genes vulnerable. Understanding the arms race between our genes and transposon activity has been a long-running question in molecular biology. This is the first evidence that endosiRNAs moderate transposon activity during DNA demethylation. EndosiRNAs provide a first line of defence against transposons during epigenetic reprogramming.”
The effects of active transposons vary, often they have no effect, only occasionally will they alter an important gene. Yet, transposons can affect almost any gene, potentially leading to different kinds of genetic disease. Studying the control of transposons, adds to our understanding of the many ways that they can impact on human health.
This work highlights that a transposons often sit within genes and are read in the opposite direction to the surrounding gene. It is this arrangement that allows cells to identify RNA messages from transposons. RNA messages read from the same piece of DNA in opposite directions are complementary, meaning they can join to form a structure called double-stranded RNA (dsRNA), which initiates the creation of endosiRNAs.
Senior scientist on the paper, Professor Wolf Reik, Head of the Epigenetics Laboratory at the Babraham Institute, said: “Transposons make up a large part of our genome and keeping them under control is vital for survival. If left unchecked their ability to move around the genome could cause extensive genetic damage. Understanding transposons helps us to make sense of what happens when they become active and whether there is anything we can do to prevent it.”
Much of this research was carried out using embryonic stem cells grown in the lab, which had been genetically modified to lack DNA methylations. Natural epigenetic reprogramming happens in primordial germ cells, the cells that make sperm and eggs, but these are harder to study. The researchers used primordial germ cells to verify the key results from their study of stem cells.
Notes:
Publication Reference
Berrens, RV., Andrews, S., Spensberger, D., Santos, F., Dean, W., Gould, P., Sharif, J., Olova, N., Chandra, T., Koseki, H., von Meyenn, F., Reik, W.. An endosiRNA-based repression mechanism counteracts transposon activation during global DNA demethylation in embryonic stem cells. Cell Stem Cell
DOI: http://dx.doi.org/10.1016/j.stem.2017.10.004
Research Funding
This work and the researchers that contributed to it were generously supported by SNSF, Gates Cambridge Trust, BBSRC (Epigenetics Institute Strategic Programme Grant), Wellcome Trust, EU BLUEPRINT and EpiGeneSys
Image Credit
Credit: R. Berrens
Header – Image of fluorescently labelled embryonic stem cells. These cells have been modified to mimic the effects of epigenetic reprogramming. DNA in the cells is marked in blue. Red indicates cells that contain active transposons.
Embedded – Graphical abstract of the key findings from the paper. The opposing directions of a transposon and a gene result in double stranded RNA that can be used to produce endosiRNA to prevent transposon activation. Credit: Veronique Juvin
Animal Statement:
As a publicly funded research institute, the Babraham Institute is committed to engagement and transparency in all aspects of its research. Animals are only used in Babraham Institute research when their use is essential to address a specific scientific goal, which cannot be studied through other means. The main species used are laboratory strains of rodents, with limited numbers of other species. We do not house cats, dogs, horses or primates at the Babraham Research Campus for research purposes.
Samples of primordial germ cells were collected from C57Bl/6J transgenic mice at embryonic day 13.5 and 14.5. The use of animals in this study was performed in accordance with the Animal (Scientific Procedures) Act 1986, and regulated by the Babraham Institute Animal Welfare and Ethical Review Body (AWERB). Experiments were planned and designed in accordance with the 3Rs.
About the Babraham Institute:
The Babraham Institute receives strategic funding from the Biotechnology and Biological Sciences Research Council (BBSRC) through an Institute Core Capability Grant to undertake world-class life sciences research. Its goal is to generate new knowledge of biological mechanisms underpinning ageing, development and the maintenance of health. Research focuses on signalling, gene regulation and the impact of epigenetic regulation at different stages of life. By determining how the body reacts to dietary and environmental stimuli and manages microbial and viral interactions, we aim to improve wellbeing and support healthier ageing.
The Company of Biologists (biologists.com) is looking to recruit a Scientific Copy Editor to work across our portfolio of five life-science journals, with a particular focus on Journal of Experimental Biology. This full-time position is available for two years initially. The role entails copyediting articles to a high standard, compiling author corrections, overseeing the journal production process, and liaising with authors, academic editors, external production suppliers and in-house staff to ensure that articles are published in a timely and professional manner.
Candidates should have a degree (ideally a PhD) in a relevant scientific area, and previous copyediting experience is strongly preferred. Additional requirements include excellent literacy skills, high attention to detail, a diplomatic communication style, good interpersonal and IT skills, a flexible approach and the ability to work to tight deadlines.
The position represents a unique opportunity to gain experience on our highly successful life-science journals and offers an attractive salary and benefits. The position will be based in The Company of Biologists’ attractive modern offices on the outskirts of Cambridge, UK.
The Company of Biologists (biologists.com) exists to support biologists and inspire advances in biology. At the heart of what we do are our five specialist journals – Development, Journal of Cell Science, Journal of Experimental Biology, Disease Models & Mechanisms and Biology Open – two of them fully open access. All are edited by expert researchers in the field, and all articles are subjected to rigorous peer review. We take great pride in the experience of our editorial team and the quality of the work we publish. We believe that the profits from publishing the hard work of biologists should support scientific discovery and help develop future scientists. Our grants help support societies, meetings and individuals. Our workshops and meetings give the opportunity to network and collaborate.
Applicants should send a CV to recruitment@biologists.com, along with a covering letter that summarises their relevant experience, why they are enthusiastic about the role, and their current salary.
All applications should be received by 27 November, although late applications may still be considered.
SUPERVISORS: Dr Thomas Theil and Dr Pleasantine Mill
PROJECT SUMMARY
This project will dissect how cell signalling via the primary cilium coordinates the activity of neural stem cells during the development of the cerebral cortex. Proliferation and differentiation of cortical stem cells are tightly controlled processes. Changes in these parameters can have profound effects on cortical size and are thought to underlie cortical malformations in human disease and the expansion of the human cerebral cortex during evolution.
To investigate roles of primary cilia in cortical stem cell development, the project will employ Inpp5e mutant mice which display an elongated, folded cerebral cortex. Using a combination of in utero electroporation with super resolution and live imaging of primary cilia dynamics in the developing brain the project aims to understand how primary cilia control the signalling required to determine the balance between stem cell proliferation and neurogenesis and how cilia determine the asymmetric inheritance of cell fate determinants These analyses will be a vital step towards gaining a comprehensive understanding of how cilia coordinate the proliferation of cortical stem cells in health and in ciliopathies, human syndromes caused by defects in cilia structure and/or function.
STUDENT TRAINING
The PhD student will be closely integrated into the Theil and Mill research groups which have overlapping and complementing interests in cortical development and primary cilia. The student will benefit from the excellent research environment and the unique research infrastructure including state of the art animal and super-resolution imaging facilities at the Centre for Discovery Brain Sciences and at the Institute for Genetics and Molecular Medicine at the University of Edinburgh.
FUNDING NOTES
Applications for BBSRC EASTBIO studentships are invited from excellent UK students (and EU citizens if they meet UK Research Council residency criteria) with at least a BSc (Hons) 2.1 undergraduate degree.
Department/Location: Wellcome Trust – Medical Research Council Cambridge Stem Cell Institute
Studentships starting October 2018
Stem Cell Biology Stem cells are defined by the dual capacity to self-renew and to differentiate. These properties sustain homeostatic cell turnover in adult tissues and enable repair and regeneration throughout the lifetime of the organism. In contrast, pluripotent stem cells are generated in the laboratory from early embryos or by molecular reprogramming. They have the capacity to make any somatic cell type, including tissue stem cells.
Stem cell biology aims to identify and characterise which cells are true stem cells, and to elucidate the physiological, cellular and molecular mechanisms that govern self-renewal, fate specification and differentiation. This research should provide new foundations for biomedical discovery, biotechnological and biopharmaceutical exploitation, and clinical applications in regenerative medicine.
Cambridge Stem Cell Community
The University of Cambridge is exceptional in the depth and diversity of its research in Stem Cell Biology, and has a dynamic and interactive research community that is ranked amongst the foremost in the world. By bringing together members of both the Schools of Biology and Medicine, this four year PhD programme will enable you to take advantage of the strength and breadth of stem cell research available in Cambridge. Choose from over 50 participating host laboratories using a range of experimental approaches and organisms.
Programme Outline
During the first year students will:
Perform laboratory rotations in three different participating groups working on both basic and translational stem cell biology.
Study fundamental aspects of Stem Cell Biology through a series of teaching modules led by leaders in the field.
Learn a variety of techniques, such as advanced imaging, flow cytometry, and management of complex data sets.
Students are expected to choose a laboratory for their thesis research by June 2019, and will then write a research proposal to be assessed for the MRes Degree in Stem Cell Biology. This assessment will also be used to determine whether students continue on to a 3-year PhD.
Physical Biology of Stem Cells
Incorporated into the ‘Stem Cell Biology’ Programme, opportunities are available specifically for candidates with a Physical, Computational or Mathematical Sciences background, wanting to apply their training to aspects of Stem Cell Biology *.
Great inroads have been made towards understanding how stem cells generate tissue and sustain cell turnover, most of which have been made by studying the biochemistry of stem cells. Less is known of their function across scales – from molecules to tissue – or interaction with their physical environment. We aim to identify the importance of physical, chemical, mathematical and engineering considerations in stem cell functionality. This could include mathematical modelling, engineering controlled environments to control stem cell function, single molecule approaches to study molecular interactions, systems biology, or investigating stem cell’s response to forces in its environment.
Eligibility
We welcome applications from those who hold (or expect to receive) a relevant degree. You must have a passion for scientific research.
Stem Cell Biology and Medicine Programme (funding by the Wellcome Trust)
We welcome applications from EU and non-EU candidates. The Wellcome Trust provide full funding at the ‘Home/EU’ rate. Funding does not include overseas fees, so non-EU applicants will need to find alternative funding sources to cover these.
‘Physical Biology of Stem Cells’ Programme (funding by the Medical Research Council)
We welcome applications from UK/EU candidates, with a Physical Sciences, Mathematical or Computational Sciences background. *The Medical Research Council provide full funding for UK applicants only. Applicants from EU countries other than the UK, are generally eligible for a fees-only award. Please check your eligibility status at https://www.mrc.ac.uk/skills-careers/studentships/studentship-guidance/student-eligibility-requirements/ before applying.
Purdue University is seeking graduate students to train for high-tech science careers in basic plant sciences through the Center for Plant Biology.
Purdue has invested more than $20 million in plant sciences and hired ten new faculty members since 2015 to expand research and education in basic plant biology. Graduate students in the Center for Plant Biology (CPB) have access to an innovative curricula, diverse research opportunities, and world-class facilities and resources.
WORLD-CLASS FACILITIES AND RESOURCES
State-of-the-art plant imaging technology and expertise to support basic research in a broad range of subdisciplines. >>Learn more
DIVERSE RESEARCH OPPORTUNITIES
This community of 35 faculty members provides a dynamic training environment for graduate students to study a breadth of research areas. >>Learn more
Apply through the Purdue University Interdisciplinary Life Science Program (PULSe) by December 1 and specify Integrated Plant Sciences as your training group
All students admitted to our graduate program receive paid tuition and an assistantship for the first year, which allows students to complete core requirements and participate in laboratory rotations. Financial support for students in subsequent years comes from training grants, research grants secured by the major professor, or research and/or teaching assistantships. Given solid academic standing and lab performance, the level of support in subsequent years will meet or exceed the level of support provided in the students first year.
A new stem cell study conducted at the University of Copenhagen shows how we may increase the vital production of insulin in patients suffering from diabetes. The discovery helps to more efficiently at less cost make insulin-producing beta cells from human stem cells. Therefore, the research paves the way for more effective treatment of diabetes. The method may also prove significant to the treatment of a series of other diseases.
415 million people worldwide have been diagnosed with diabetes. And the number continues to rise. Common to all diabetes patients is that they lack the ability to produce sufficient amounts of insulin, which regulates the blood sugar in the body. This can lead to a number of complications and in many cases be potentially fatal.
A new study conducted at the University of Copenhagen, which has just been published in the internationally acclaimed journal Nature Cell Biology, shows how researchers using human stem cells can produce insulin-producing cells that in the future can be transplanted into diabetes patients.
‘By identifying the signals that instruct mouse progenitor cells to become cells that make tubes and later insulin-producing beta cells, we can transfer this knowledge to human stem cells to more robustly make beta cells, says Professor and Head of Department Henrik Semb from the Novo Nordisk Foundation Center for Stem Cell Biology at the Faculty of Health and Medical Sciences.
The Cells’ Development Depends on their Sense of Direction
The research group, which in addition to Henrik Semb consists of Ph.D. Zarah Löf-Öhlin and assistant professor Pia Nyeng, among others, originally set out to study how the body creates the complex piping systems that transport fluids and gasses in our organs.
They wanted to understand the machinery for instructing progenitor cells into their different destinies. To their surprise, the mechanism turned out to be simple. According to Assistant Professor Pia Nyeng, these processes are mainly controlled by the progenitors’ ability to tell up from down (the cells’ so-called polarity).
Hormone-producing cells (green and red) in the pancreas are formed in close contact with the piping system (blue)
‘It turns out that the same signal – the so-called epidermal growth factor (EGF) pathway – control both the formation of pipes and beta cells through polarity changes. Therefore, the development of pancreatic progenitor into beta cells depends on their orientation in the pipes. It is a really amazing and simple mechanism, and by affecting the progenitor cells’ so-called polarity we can control their conversion into beta cells’, says Pia Nyeng.
Exciting Potential for Diabetes Treatment
The study is mainly based on tests performed on mice, but the researchers decided to examine whether the same mechanism can be found in human cells.
’Zarah Löf-Öhlin discovered that the same cell maturation mechanism applies to the development of human cells. Now we can use this knowledge to more efficiently turn human stem cells into beta cells in the laboratory with the hope to use them to replace lost beta cells in patients suffering from diabetes’, says Henrik Semb.
The researchers expect regulation of cell polarity to be key to the development of many other human cell types, for example nerve cells. This may contribute to the development of stem cell therapy targeted at other diseases.
As we sat together at a farewell dinner after I graduated from Princeton University, my advisers asked me, “What got you into science?”. Although a simple, straightforward question on the surface, it sent me down memory lane and I found it incredibly difficult to provide a concise, one-line response.
I grew up in a rural settlement just outside the town of Udhampur which is situated in the Indian state of Jammu and Kashmir. A typical day involved school-time, helping my father and uncle at their grocery and stationary stores, playing street cricket, and planning activities around the daily power-cuts. Pursuing a bachelor’s degree was considered a distant dream and further education was unheard of in the area. Accordingly, I listened to the modest academic expectations from family and society, and ended up doing just well enough that no one ever complained. However, one fateful day in 6th grade forever changed this complacent attitude. During one of the lectures, the science teacher in the school called out one of the top students in the class who was siting besides me and also happened to be my close friend. She advised her, “Avoid the company of Yogesh. He is going to spoil you, and make you like him”. The teacher was correct in identifying me as neither academically sincere nor hard-working, and as I comprehended the truth in her words my face reddened with embarrassment. At the same time, I realized it as an opportunity for me to study hard and prove her wrong. Consequently, this had a substantial, even revolutionary, impact on me. For the first time I became serious about studying and generally succeeding academically. Fortunately, I found that I enjoyed studying and learning new scientific concepts. This was further reinforced by some exceptional teachers at school and family support at home. What began as a source of shame and embarrassment, slowly became point of pride! My interest in science continued to grow and I spent much of my teenage years solving interesting problems in physics and math.
Based on my performance in a certain exam during high school, I was assigned to major in chemical engineering for my undergraduate studies at IIT Gandhinagar in India. Here again, I experienced a crucial juncture in my sophomore year that made me realize how much I enjoyed research. Thanks to Prof. Narayanamurthy who taught us the first course in chemical engineering, I went from carefully planning a career pursuing an MBA after undergraduate studies to genuinely appreciating the nuances of chemical engineering. Importantly, I actively explored opportunities to be involved in research projects at my undergraduate institution. This, in turn, naturally led me to apply for PhD programs in chemical engineering.
I came to Princeton set on continuing my studies in chemical engineering; however, after taking a chemical reaction engineering course taught by my eventual adviser, Stanislav Shvartsman, I became intrigued by the chemistry inside living cells. He drew elegant parallels between reactions that happen in a chemical plant and in a living cell. This and subsequent interactions with another future adviser, Trudi Schüpbach, fostered a curiosity in biological questions. A lack of formal training in biology made the transition challenging, but it was also exciting to delve into a new field. Specifically, I was fascinated by how complex structures and functional forms emerge from elemental embryonic states. How are desirable properties such as precision, reproducibility, and robustness imparted to biological systems? Subsequently, my PhD focused on answering such fundamental developmental biology questions in the context of the early embryonic patterning in fruit flies (see here for details).
Together, fortuitous interventions bolstered by persistent hard work have led me to a place where I wake up every morning excited about going to lab and doing science. As I now embark on a foray into new biological research directions for my postdoctoral work, a diverse set of life and research experiences have taught me that nothing is impossible. Above all, one must follow their passion, work with inspiring and supportive mentors, and take risks.
Research Assistant in Developmental and Regenerative Medicine
Department of Physiology, Anatomy and Genetics & Weatherall Institute of Molecular Medicine, University of Oxford
Grade 6: £28,098 p.a.
Applications are invited for an experienced and meticulous Research Assistant to join the Cardiovascular Development, Repair and Regeneration team working under the direct supervision of Professor Paul Riley and Dr Filipa Simoes. The project is funded by the British Heart Foundation.
The aim of the project and team is to decipher the cellular and molecular mechanisms involved in the regenerating epicardium of the adult zebrafish heart. The Research Assistant will be responsible for carrying out a range of molecular and cell biological procedures, as well as assisting with experimental zebrafish work. Previous molecular biology and embryology experience is essential but relevant training in cardiovascular development and state of the art techniques will be provided where necessary. A range of different techniques will be used including CRISPR/Cas9 technology, Nanostring analysis, in situ hybridisation and multiplex hybridisation chain reaction, tissue cryosectioning, immunofluorescence microscopy, microinjection of zebrafish embryos, cell sorting, and management of wild type and genetically modified zebrafish lines. The post is ideally suited for a candidate with an interest in developing a career that involves working at the interface of Developmental Biology and Regenerative Medicine, with a strong background in the former.
This project is a collaboration between the Departments of Physiology Anatomy and Genetics, and the Weatherall Institute of Molecular Medicine.
Candidates must have a degree in a relevant field. Expertise in molecular biology techniques, histology, and imaging, with experience of animal models including husbandry and small animal surgery is preferable.
You will be based across two sites: the Sherrington Building, South Parks Road, Oxford, OX1 3PT and the Weatherall Institute of Molecular Medicine, University of Oxford, John Radcliffe Hospital, Oxford, OX3 9DS.
The position is offered until 31 December 2018. An early start date is preferable.
The closing date for applications is 12.00 noon on Wednesday 29 November 2017. Interviews will be held on Thursday 14 December 2017.
Position Summary: A postdoctoral position is available in the laboratory of Dr. Kristin Gribble in the Josephine Bay Paul Center for Comparative Molecular Biology and Evolution, to study the role of mitochondrial homeostasis in neurodegenerative disease. This project takes a multidisciplinary approach using phenotypic, transcriptomic, genetic, genomic, epigenetic, biochemical, and imaging methods in a new animal model system, the monogonont rotifer.
The Gribble lab focuses on understanding the cellular, epigenetic, and evolutionary mechanisms of aging. We are specifically investigating the genetic and epigenetic mechanisms of maternal effects in determining offspring health and lifespan. The successful applicant will have the opportunity for independent and novel research in an innovative new laboratory. For more information about the laboratory, please see our website, http://www.mbl.edu/jbpc/gribble/ or contact Dr. Gribble at kgribble@mbl.edu.
The position is available immediately, and is renewable annually depending upon progress.
Basic Qualifications: Applicants should posses a Ph.D. and/or M.D. in molecular biology, cell biology, biochemistry, genetics, or a related field. The ideal candidate will have a record of scientific rigor, productivity, and creativity; the ability to work both independently and as part of a team; and a strong publication record. Excellent oral and written communication skills are required. Highly motivated individuals with experience in other model systems are encouraged to apply.
Preferred Qualifications: The preferred applicant will have a background in mitochondrial biology, developmental biology, and/or neurobiology. Prior experience in confocal microscopy, RNAi, and/or bioinformatics is beneficial.
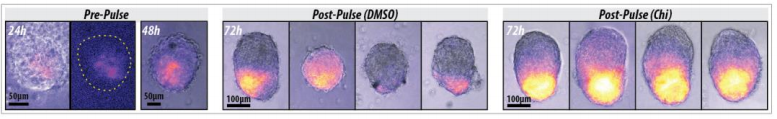
Embryonic patterning is dependent on the establishment of the anteroposterior (AP) and dorsoventral axes early in development. In mammals this occurs by a breaking of symmetry in the epiblast, however the molecular mechanisms controlling this process are still not fully understood. This week we feature a paper published in the latest issue of Development that models these patterning events in gastruloids. Two authors: David Turner and Peter Baillie-Johnson from the Martinez-Arias group at the University of Cambridge told us more.
David Turner and Peter Baillie-Johnson
David and Peter, can you give us your scientific biographies and the main questions the lab is trying to answer?
DT Our lab is primarily concerned with understanding the early decisions involved during mammalian development. Specifically, how the early mouse embryo patterns itself and specifies the body axes. We mainly use mouse embryonic stem cells as a model system to study these early developmental processes both in normal monolayer culture conditions (2D) and also using our gastruloid model system in 3D (embryonic organoids).
DT For quite some time I’ve been interested in cell signalling in general and how cell fate is determined, an interest which was initially sparked during my Pharmacology honours degree in Liverpool. Following in this vein, I took up a PhD (also in Liverpool) with Prof. Mike White (who’s now in Manchester) where I used single-cell fluorescence imaging to investigate the dynamics of NF-kB in response to low doses of the cytokine TNFa.
PBJ I studied Natural Sciences as an undergraduate at Cambridge from 2010-2013, specialising in Genetics in my final year. I started my PhD in October 2013 and handed in the final copies of my thesis in October 2017. During my PhD, I worked on developing the gastruloid system and applying it to the origin of the mammalian spinal cord. Since April this year, I’ve been working as postdoctoral research assistant to Professor Jenny Nichols, who has helped me shift my focus to earlier stages of mouse development, namely gastrulation.
Axial organisation of gastruloids from Figure 1, Turner et al, 2017
David and Peter, how did you both come to join the Martinez Arias lab?
DT Having finished my PhD in Liverpool, I saw a post-doc position open in Alfonso’s lab which was about using mouse embryonic stem cells as an in vitro model system to try and understand their properties and their differentiation potential. My background from my PhD was strongly in single-cell imaging and cell signalling so it was a perfect opportunity for me to pursue my interests.
PBJ I first joined Alfonso’s lab in 2012 as a summer student during my undergraduate course. At that time, the lab was more focused on the regulation of embryonic stem cell pluripotency and differentiation. I was really taken with this introduction to stem cell biology and felt warmly welcomed into the lab, so I applied to follow the work up in my final year research project. While awaiting the results of my finals, I ran into David and heard about the first gastruloid experiments, which naturally became the focus of my PhD.
Your paper addresses the question of axis establishment in the early mammalian embryo. What was known about the molecular control of polarity prior to your paper?
PBJ The textbook models of antero-posterior axis specification in the mouse describe an opposing arrangement of signals and their inhibitors emanating from the extra-embryonic tissues that surround the then radially-symmetric epiblast. These models describe how the signals in the future posterior are balanced in the future anterior by their corresponding secreted inhibitors. The cells of the epiblast are then restricted to undergo a localised EMT (i.e. the beginnings of the primitive streak) only in the future posterior region, while the anterior epiblast remains reserved for the anterior nervous system. The key feature of this model is that the asymmetry originates in the extraembryonic tissues, which then becomes conferred on the underlying epiblast. Following our study, it now seems as if this careful balance of signalling across the embryo might act to permit an intrinsic symmetry-breaking event in the future posterior, rather than actively instructing the process. It remains to be seen whether the lack of the anteriorly expressed inhibitors in the gastruloids fully explains the lack of anterior structures that we have observed.
Why did you use gastruloids and not an in vivo system for this project?
PBJ I think that the acquisition of antero-posterior polarity is a good example of a topic that has been well-described in the embryo through genetic experiments. I think the strength of the gastruloids as an experimental tool is in providing insights on development that would not be possible by looking at the embryo alone – in this case by looking at development in the absence of the extraembryonic tissues and the post-implantation mechanical microenvironment. This In vitro system also enables the possibility of identifying the sufficient components behind a genetic process by starting with a minimal set of interacting parts. This approach is quite different to genetic experiments, which are good at identifying the necessary components of developmental systems, but which can’t (easily) demonstrate sufficiency.
DT Echoing what Peter said, the gastruloids have a significant advantage over in vivo systems in that the developmental processes and events we’re interested in occur at a time in the embryo that’s difficult to access and manipulate experimentally. With our gastruloids, we’re able to ask very specific questions about subtle timings of signals in ways which are either very difficult or impossible to do in the embryo, such as pulses of signals at very precise temporal intervals. Compared with in vivo models, gastruloids are relatively inexpensive, easy to manipulate, and amenable to experimental perturbation. Also, because of this ability to recapitulate many of the early developmental processes (mentioned above), our system has the real potential to be used as a way to reduce or replace animals used for research in development, which is central to the aims of NC3Rs (National Centre for the Replacement, Refinement and Reduction of Animals in Research).
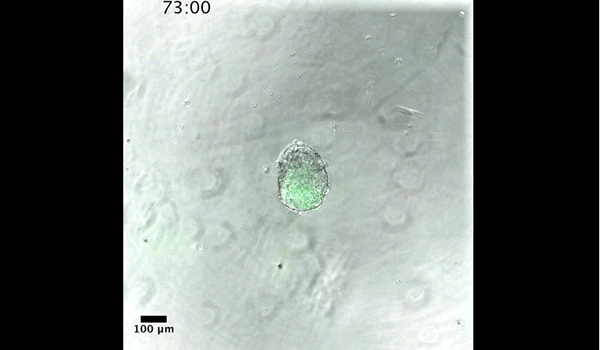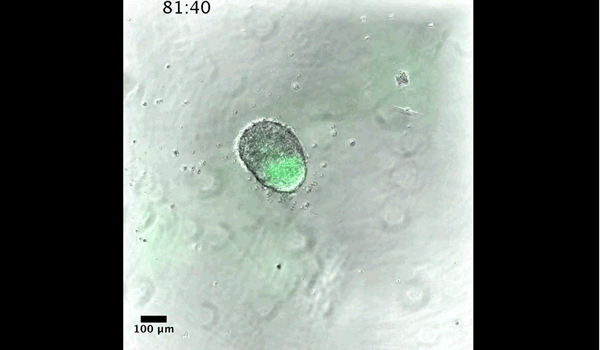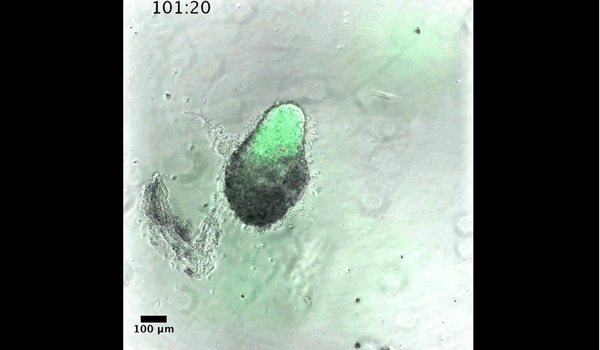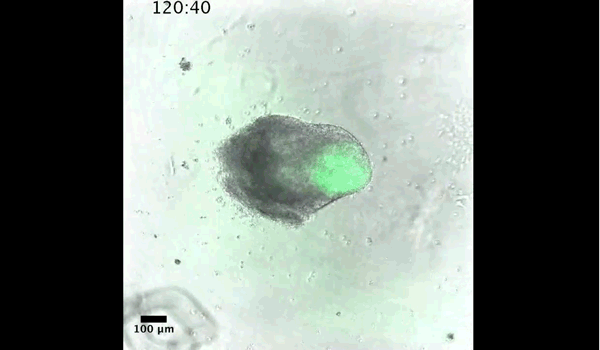
Gastruloids recapitulate early embryonic events, from figure 2, Turner et al, 2017
Can you give us the key results of the paper in a paragraph?
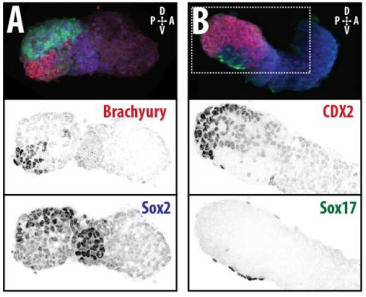
DT The main aim of the paper was to use gastruloids to study anteroposterior patterning. We found that gastruloids spontaneously break symmetry, polarise gene expression and undergo axial elongation in a robust and reproducible manner similar to the early embryo. One end of the gastruloid resembles the posterior region of the embryo, where Brachyury is up-regulated. Our quantitative analyses show that this is regulated by both Nodal and Wnt/b-Catenin signalling, and surprisingly with no detectable involvement of BMP signalling. Our most interesting finding is that the AP axis can form without any of the extraembryonic tissues which have been suggested to be important for this process. This lead us to hypothesise that the role of the extra-embryonic tissues in the embryo may not necessarily be to induce the AP axis, but to bias the intrinsic symmetry-breaking potential of the embryo.
Wnt signalling enhances gastruloid polarisation, from figure 3, Turner et al, 2017
To what extent does gastruloid patterning recapitulate embryonic development?
DT I think that the gastruloids can recapitulate many aspects of early post-implantation embryonic development in the mouse and at a similar timescale to the embryo. First and foremost is their ability to develop AP polarity with Brachyury expression at one end (which we have designated the posterior) and in the DV direction where Sox2 is directly opposite Sox17. Furthermore, they’re able to extrude cells from the extending, Brachyury-positive region, in a manner akin to gastrulation at the primitive streak (hence their name). Gastruloids do not, however, develop the pre-occipital tissues of the head and brain, so their antero-posterior axis probably only represents the post-occipital levels of the embryo. We think this is due to a lack of tissues that would protect this region from high levels of ‘posteriorising’ signals (such as the prechordal plate and anterior mesoderm). It remains to be seen exactly which axial levels are represented in these seemingly posterior tissues and what we need to do to expand this representation.
PBJ I think an interesting feature of gastruloid development is the time over which the events unfold, which in our standard cultures corresponds approximately to the five days after implantation. It’s striking how they always undergo the same progression of changes in gene expression and morphology in this time and this is, for me, a key reason for using them to investigate early developmental events such as gastrulation.
In addition to your own gastruloid research, other recent papers have described systems that aim to recapitulate early development in vitro. This has generated considerable media attention and has sometimes been described as ‘creating artificial life’. How do you feel about this description? More generally, do you think there are any ethical issues thrown up by research using so-called synthetic embryos?
PBJ I think that this is a sensitive issue that certainly demands careful ethical consideration – perhaps in a longer form than an interview. Although it’s a semantic point, I think that the description of “creating artificial life” is imprecise and unhelpful and I’d prefer to see “engineering developmental systems” used instead. I think that this emphasises that these systems are controlled approaches that can recreate features of embryonic development, but which might otherwise be limited in their developmental potential.
Many of the ethical considerations surrounding these experiments and ultimately the scope of the legislation that will regulate them hinge on how exactly we define an embryo. For example, if researchers constructed a set of tissues from specific cell types in the exact image of the early embryo, would we consider the two to be equivalent? Conversely, how would a structure that closely resembles a significant proportion of the embryonic body measure up to the embryo if the representation was not complete? Would this logic extend further to individual organs or organoids? I think that scientists and the public need to consider these issues if we are to determine how we will regulate this form of research alongside the existing framework for research with embryos (which may itself need to be revised). With a clearer definition in hand, it will be easier to approach the deeper questions of where to draw appropriate limits of research on mouse and human systems, whether engineered or otherwise. To paraphrase a commentary from Martin Pera et al.1, the extent to which researchers can recreate embryogenesis in vitro will come to define not only how experimentally useful these systems are, but also how much scrutiny they will attract.
When doing the research, did you have any particular result or eureka moment that has stuck with you?
DT I think one of the best moments was late in 2013 when Alfonso and I were watching a time-lapse movie on the microscope that Susanne van den Brink had just finished imaging (the lead author on the first gastruloid paper). This is where we saw, for the first time, cells being extruded from the elongating region of the gastruloids and realised how important and useful this system was going to be. A second important moment experimentally was during the early stages of optimising the gastruloid protocol, when we were trying to improve the reproducibility within each plate and between plates of gastruloids. We found that the simple act of adding a second wash of PBS improved all the stages of gastruloid formation: the aggregation, the patterning and the elongation, so this felt like an important breakthrough.
PBJ There have been a couple of those precious moments when you’ve seen something that no-one else has before, which have been real highlights for me. I think I was lucky to have worked on such an exploratory project for my PhD as those moments have really stayed with me. In general, however, the work has progressed incrementally but I’ve been consistently surprised by the level of autonomy that the gastruloids show as we’ve started to look more closely at their development.
Gif generated from supplementary movie 2: Live-imaging of gastruloid development. From Turner et al, 2017
And what about the flipside: any moments of frustration or despair?
DT We found, after quite a few frustrating early attempts, that the initial culture conditions are essential to ensure good formation of gastruloids, i.e. low passage numbers, consistent splitting ratios, plating density. It took a little while for us to realise this and to ensure that our stock flasks are treated in a consistent manner.
PBJ There were definitely frustrating times in the early stages of my PhD, when we hadn’t identified the key variables that could determine whether the cultures would thrive or fail. We also had moments of doubt as to whether our observations would prove to be new and useful biological insights, but our confidence in using gastruloids as an experimental tool has grown as we’ve learned more about their development.
What are your career plans following this work?
DT I was recently awarded a David Sainsbury Fellowship from the NC3Rs to use the gastruloid system to study left-right asymmetry during mammalian development, and for the next three years I’ll be working pretty much solidly on that!
PBJ I’m currently working as a postdoctoral Research Assistant to Professor Jenny Nichols, who is helping me to cut my teeth on mouse embryology. By studying the gastrulating mouse embryo first hand, I’m trying to determine how closely the process of “gastrulation” in the gastruloids measures up to that in the embryo. By doing so, I hope to determine whether gastruloids could be used as an experimental tool to dissect this complicated phase in the life of the embryo. My work is closely aligned to that of a Cambridge-based consortium that is investigating gastrulation through single cell genomics, so I hope that my work with the embryo will provide a reference for their findings and that the gastruloid system might offer a complementary approach in the future.
And what next for the Martinez Arias lab?
DT There are quite a few avenues of our research at the moment. One is to get more of a handle on what drives the elongation in gastruloids, whether it is a mechanism based purely on convergent extension or rapid cell growth, and whether the signals suggested to be involved during the axial extension of the embryo work in a similar manner in gastruloids. We’re also interested in seeing whether the gastruloid system is applicable to later stages of development and what its limitations might be; our ongoing collaboration with our co-authors in Matthias Lutolf’s lab at EPFL is an important part of this research.
Finally, what do you two like to do when you are not in the lab?
DT As little as possible to be honest, since free weekends and evenings are quite hard to come by when stem cell work is involved! Any free time I have I like to spend with my wife and my two children. Otherwise, I spend plenty of time reading and am currently working my way through Stephen King’s Dark Tower novels, which I can certainly recommend.
PBJ I love to be outdoors, so try to get out walking, running or cycling at the weekends. I’m also a keen cook and enjoy testing out new recipes on my friends and family.
 , also called transposable elements, are ancient viruses that have become a permanent part of our genes. Around half of the human genome is made of transposons, many are damaged, but some can become active. Active transposons can be harmful because they move about the genome. When transposons move they can damage genes, leading to genetic illnesses and playing a part in some cancers.
, also called transposable elements, are ancient viruses that have become a permanent part of our genes. Around half of the human genome is made of transposons, many are damaged, but some can become active. Active transposons can be harmful because they move about the genome. When transposons move they can damage genes, leading to genetic illnesses and playing a part in some cancers. (No Ratings Yet)
(No Ratings Yet)


 (22 votes)
(22 votes)
 (2 votes)
(2 votes)