Two Post-Doctoral positions are available immediately in the laboratory of Stephane Angers in the Leslie Dan Faculty of Pharmacy (www.angerslab.org), University of Toronto, to develop strategies to inhibit developmental signalling pathways in cancer.
Our laboratory is studying the genetic circuitry important for cancer development to identify vulnerabilities that could be harnessed for the development of new medicine. Our approach involves genome-wide CRISPR functional screens, which we perform in cancer cell lines and primary cells to identify context-dependent fitness genes important for cancer cell growth. Our recent work identified the Wnt receptor Frizzled-5 as being essential for the growth of a subset of pancreatic cancers (Steinhart et al, Nature Medicine 2017). In collaboration with the group of Dr. Sachdev Sidhu we are developing synthetic antibodies targeting Wnt components and other cell surface proteins involved in cancer progression.
We are looking for a highly motivated, self-directed postdoctoral fellow with strong team capabilities. Preference will be given to candidate that recently obtained their Ph.D (less than 12 months) and that have a keen interest to develop and independent research program in the areas of genomics, cell signaling and cancer biology. The ideal candidate will have a strong publication record as a first author in the field of cancer biology with expertise in either organoid models, mouse models, cell signaling or antibody development.
Please submit your application as a single PDF document to stephane.angers@angerslab.org with the following information: a cover letter, statement of interest, and CV with contact details for 3 referees.
The contribution of scientific research in shaping societies is increasingly significant. However, African researchers make up only around two per cent of the world’s academic research community. One of the central problems for African science is poor quality and quantity of research-based education. We believe that basic scientific research could help developing African nations, and we also believe that Drosophila melanogaster – the fruit fly – can be used as a powerful and inexpensive model system to scale-up and improve both post-graduate education and research output in Africa.
The “DrosAfrica” project (www.drosafrica.org) has the aim of training and establishing a connected community of African researchers with the knowledge to be able to use Drosophila as a model system to study biomedical problems. Historical evidence of the power of Drosophila as a research model comes from Spain in the 80’s (see this piece by Alfonso Martinez-Arias for background) where Drosophila transformed the scientific panorama when resources were limited. With this in mind, we have decided that the first aim of DrosAfrica should be to train well-established African scientists to use Drosophila as a model system to study human diseases.
To date, DrosAfrica has trained 57 scientists from many African countries including South Sudan, Egypt, Nigeria, Kenya and Uganda. These efforts have already paid dividends, as various DrosAfrica alumni and collaborators are using the fruit fly in their own labs, such as Profs. Abolaji and Adedeji, and Drs. Vicente-Crespo, Wuyep, and Nyanhom (Box 1). Critically, these scientists are already training the next generation, with multiple PhD and MSc students in their lab leading biomedical projects using the fruit fly as a model.
We believe DrosAfrica can make a substantial contribution in developing and advancing science for sustainable prosperity in Africa. The mission of DrosAfrica is two-fold. Firstly, to help establish a highly skilled community of researchers capable of using Drosophila as a model system to study biomedical problems. Secondly, to develop Drosophila biomedical units with high-quality research facilities that allow African researchers to train and run projects that will impact the biomedical sciences.
Participants and Faculty of the DrosAfrica2013 workshop at Kampala International University, Uganda.
The Workshop Approach
To achieve our goals, we have adopted an approach that centres on carrying out workshops with world-class researches training African scientists at host institutions. Through a collaboration with Professor Sadiq Yusuf, then at Kampala International University (KIU, Uganda), we organised the first DrosAfrica workshop in KIU for African scientists in 2013, followed by others in Uganda, Kenya and Nigeria. The aim of the workshops is to equip African scientists with all the knowledge and tools required to be able to use Drosophila to study biomedical problems. The workshops are organised to be highly practical and interactive, including hands-on laboratory experiments to compliment the lectures. The 20-25 workshop participants learn about the advantages and disadvantages of Drosophila for biomedical research, as well as how to set-up a Drosophila laboratory. Ultimately, and most importantly, the workshop helps participants to improve their critical thinking and to gain further experience using the scientific method.
Another key aspect of the workshops is to facilitate networking, especially among the African scientists that might be ready to implement in their own institutions the research approaches learnt in the workshop. The topic of the workshops is tailored to the research interest of the collaborating institution, and can range from insecticide resistance and host-pathogen interactions to cancer and neurodegeneration. To identify a host institution for a workshop we either directly contact a prospective institution that we think would benefit from our approach, or a scientist that knows about us (occasionally a previous workshop participant) invites us to organise a workshop where she/he works. After initial discussions by skype or phone, we visit the institution and make further arrangements for the workshop, topics and funding.
Our Thanks To
None of the work by DrosAfrica would be possible without the extremely generous help from various organisations and scientists. The Company of Biologists, who has funded various workshop expenses including the microscopes that are essential for Drosophila manipulation, constantly supports DrosAfrica. We would also like to thank the faculty that has helped us in our efforts over the years. The response has always been remarkable with everyone we approached agreeing to help us. We also thank KIU, The Cambridge-Africa Alborada Research Fund, the International Centre for Genetic, Engineering and Biotechnology (ICGEB), The World Academy of Sciences (TWAS), EMBO, and The Wellcome Trust for financial support. We are also thankful to trendinafrica.org, CamBioScience, the Department of Zoology and the University of Cambridge for support, and St John’s, Emmanuel and Pembroke Colleges (Cambridge, UK) for funds.
Anyone that reads this article is encouraged to visit our website (www.drosafrica.org), and think about ways in which they can help, from their own work and time to any advertisement and financial support.
ASANTE SANA
Box 1. Achievements by DrosAfrica alumni and collaborators
Prof. Amos O Abolaji co-organised and participated in a five-day course at the University of Jos (Nigeria) on the use of Drosophila in Experimental Medicine (2016). In all, about 40 participants attended the event. “My first encounter with this amazing model was during a postdoctoral training at the Federal University of Santa Maria, Brazil. Now in Ibadan, so far thirteen M.Sc. students have successfully used the fly for their projects. We now have a Drosophila lab that can conveniently accommodate 20 students.” Prof. Abolaji is hosting our next workshop at the University of Ibadan, July 2017 (http://ibadan2017.drosafrica.org)
In KIU (Ishaka, Uganda), under Dr. Marta Vicente-Crespo’s supervision (now in St Augustine International University (Kampala, Uganda), two BSc Pharmacy, and two MSc have completed their thesis using Drosophila to study various subjects from toxicity studies to epilepsy and the olfactory system. In addition, she currently supervises three PhD and two MSc students using Drosophila to investigate RNA decay, epilepsy and aging.
Dr. Ponchang Apollos Wuyep, Associate Professor of Applied Microbiology and Biotechnology, Head of Department, Department of Plant Science and Biotechnology, Faculty of Natural Sciences Building, University Of Jos, Nigeria. “My focus is fungal infectious studies. More specifically, to infect Drosophila melanogaster with various virulent Aspergillus sp and then screen for plant compounds that might help the fruitfly to fight the infection…maybe venture into antifungal drug screen. I am working with three students (one MSc, one BSc and one PhD). All these projects got inspired by DrosAfrica”. Dr Wuyep will teach in our 2017 workshop at the University of Ibadan.
Dr. Steven Nyanhom, Chairman of the Department of Biochemistry (Jomo Kenyatta University of Agriculture and Technology JKUAT, Nairobi) is currently using Drosophila in his research. He participated as Faculty in our last workshop in September 2016 at ICIPE, Nairobi.
Prof. Ahmed A. Adedeji. Habib Medical School, Islamic University In Uganda (IUIU), Kampala, Uganda. Currently using Drosophila in his research. He has repeatedly participated as Faculty in our workshops, and he will teach in our next workshop at the University of Ibadan, July 2017.
Mr. Temitope Etibor, former staff of KIU Western Campus, was accepted at the Integrated Biology and Biomedicine PhD program at the Institute Gulbenkian in Portugal. Etibor’s words show that the impact of the workshops goes way beyond the practical skills of working with flies: “The faculty of the DrosAfrica have been very good mentors and wonderful on a personal and career level. I have always been in touch with Martha Vicente-Crespo and Will Wood (members of Faculty) and they have helped me push my career forward in order to make me an excellent scientist. Through the many things I have learnt, I was able to apply for and successfully obtained an FCT PhD Scholarship in Portugal with the support of the aforementioned Faculty. I am so happy to be an Alumnus of the DrosAfrica initiative and I hope they keep receiving funds to aid the cause of research progress in Africa”.
Tokindo S. Okada (here referred to as TSO) was one of the leaders who steered developmental biology in new directions when this field was at its turning point around 1980. He also made invaluable contributions to the creation of a global forum for developmental biologists. He died on January 17, 2017, two weeks short of his 90th birthday; his name, Tokindo, stands for his birth on New Year’s Day of the Asian lunar calendar. Among developmental processes, two of his major interests were in the flexibility of the differentiated state, and in tissue organization from different cell types with different cell-adhesion properties. His own studies and those of researchers from his school created new directions for modern developmental biology in ways reflecting those interests.
TSO was born in Itami in Hyogo as a son of Rihei Okada, an owner of an old sake warehouse, collector/researcher of rare birds and authority on the Haiku poet Basho Matsuo. He thus grew up in a highly cultural environment. During his Konan High School days, he was exposed to contemporary developmental biology using amphibians, as conducted by Hiroshi Takaya. This inspired him to study developmental biology at Kyoto University. The most important elements of his undergraduate and graduate periods were his meeting and marriage to Ei Waki, who stimulated and supported TSO in all aspects throughout the rest of his life.
TSO received his PhD with a focus on tissue interactions in endodermal organogenesis in amphibians (Okada, 1960). Around this period, TSO, together with Ei Okada (by then his wife), visited the laboratories of Conrad H. Waddington at the University of Edinburgh, Department of Genetics (1957-1959), and of James Ebert at the Carnegie Institute of Washington in Baltimore (1964). These visits had a tremendous impact on his life. Studying with Waddington must have broadened TSO’s scope, and working with Ebert made him – in his own words – ‘learn from a wide perspective to organize people’.
As a result of these visits to British and American institutions, TSO was acquainted with leading developmental biologists, and he himself became highly recognized. This also promoted visits from many developmental biologists to TSO in Kyoto, resulting in the formation of an international forum centered around TSO, including Nicole Le Douarin, John Gurdon, Anne MacLaren, Lauri Saxén, Alberto Monroy, Walter Gehring, Volker Schmid, Aron Moscona, Jim Weston and others. TSO took advantage of this forum to help strengthen global liaisons among developmental biologists beyond personal ties. He served as the president of the International Society for Developmental Biologists (ISDB) from 1982 to 1986, and was awarded the Ross Harrison Prize in 1989 for his work on transdifferentiation. In the Asian sector, he also made a great effort to create China-Japan and India-Japan collaborations among developmental biologists.
His long friendships with John Gurdon and Nicole Le Douarin were particularly special. Gurdon first visited TSO in 1962 on his way back to Oxford from the USA via Japan; TSO had already told Ei then that John would eventually be awarded the Nobel Prize. Later, a student from the Okada school, Kazuto Kato, did a post-doc with Gurdon. TSO and his family spent a summer at Woods Hole with Nicole Le Douarin when she had started using chick-quail chimeras, which had a revolutionary impact on developmental biology using avian models (Le Douarin, 1973). TSO and Le Douarin developed a mutual respect and friendship. One of us (H.N.) was the first Japanese postdoctoral fellow (1978-80) to work at Le Douarin’s institute in Nogent-sur-Marne and many Japanese students followed, including Hirohiko Aoyama and Yoshiko Takahashi from TSO’s lab – the latter of whom united many of the ideas of Le Douarin and TSO (e.g. Sato et al., 2002).
TSO was promoted to a full professor at the Department of Zoology in Kyoto in 1967; during this period, he investigated organ reconstitution from dissociated kidney cells (Okada, 1965). This work could be considered a prototype of the currently popular organoid models. However, a real turning point for TSO came when he founded a new laboratory as a professor in the newly launched Department of Biophysics at Kyoto University in 1968. We were the first graduating class of the department. Japanese laboratories at the time were organized by a full professor, an associate professor and a few assistant professors, adopting a style analogous to that of German laboratories. TSO invited Goro Eguchi, who was working on lens regeneration in the newt iris, and Masatoshi Takeichi, who was then working on the lens, to join him as associate and assistant professors, respectively. TSO started investigating cell differentiation and flexibility of the differentiated state.
TSO was fond of the color contrast of dark green and red; his office furniture in the new Biophysics building bore this contrast. His green jacket was his trademark. He once owned a red Alfa Romeo. When TSO appeared at Le Douarin’s institute, they were very impressed by the color combination of his dress as it surpassed their expectations for a Japanese scholar. Thus, his life was rich in dandyism, and its combination with his clairvoyant science charmed his students and many other people.
In the classroom, TSO’s favorite teaching subjects strongly reflected his interests in topics such as transdetermination (e.g. the serial imaginal disc transplantation experiments of Ernst Hadorn; Hadorn, 1968) and tissue segregation (e.g. Malcolm Steinberg’s differential adhesiveness hypothesis; Steinberg, 1970). TSO also wrote many introductory books on developmental biology in Japanese for nonprofessionals, students and professional biologists. These were easy to read, inspiring and fascinating, and spoke of the beauty and mystery of developmental processes. They were rich in new and forward-looking conceptual frameworks. Of course, the flexibility of differentiation and cell-cell interactions for organogenesis always formed the basis of his books.
Inspired by his books, many talented students gathered at the TSO lab. The 10-year period from 1975 to 1984 was the highlight of Okada’s group, not only because of scientific productivity but also in terms of training the next generation of developmental biologists to develop their own unique characters; this became referred to as the ‘Kyoto School of Developmental Biology’. Although only chicken and mouse embryos, and some amphibians, were used in the TSO lab, his broad interests also encompassed areas as diverse as plant development. Graduating students went on to use various organisms in their subsequent careers: cats (Masami Watanabe), zebrafish (Kohei Hatta), medaka (late Kenjiro Ozato), newts (Mitsumasa Okamoto and Shin-ichi Abe), Drosophila (Shigeo Hayashi and Akinao Nose), butterflies (Kazuo Watanabe), nematodes (Kazuya Nomura and Shin Takagi), oligochaetes (Chikako Yoshida-Noro), cellular slime molds (Hideko Urushihara) and Arabidopsis (Koji Goto). Some of his students turned to cell biology (Yasuhiko Tsunematsu, Kei Takahashi, Masamichi Ueda, Kenji Ueda, Kenji Okazaki, Yasuji Ueda, Yasuaki Shirayoshi and Akira Nagafuchi). This diversity reflects the school’s culture that promoted individual interest-oriented choices of organisms and strategies.
TSO devoted himself to the study of the flexibility of differentiated states. As a student, Yoshiaki Ito observed a mass of lens cells that developed in a long-term culture of chicken embryonic neural retina. TSO immediately realized that this represented transdifferentiation from the retina into the lens and started an in-depth analysis of this phenomenon (Okada et al., 1975). He and his student Masasuke Araki identified two different mechanisms by which lens can form from neural retina culture (Araki and Okada, 1977). At early stages (around E3.5), before neuronal differentiation, neural retinal cells behaved like stem cells of all ocular tissues (Okada et al., 1979), whereas at later stages, after retinal cell differentiation (around E8), generation of lens appeared to be genuine transdifferentiation – re-fating of differentiated cells. TSO himself performed many experiments involving retinal cultures and immunohistochemistry. His last series of experiments dealt with the mechanism of lens transdifferentiation from the E8 retina. He found that an approximately 10-day period of spreading culture was required for lens transdifferentiation to occur (Okada et al., 1983). Thirty-five years later, it was shown that the spreading culture condition results in reduction of Notch signaling, which otherwise inhibited the intrinsic lens-generating potential of the neural retina (Iida et al., 2017).
TSO also routinely used mouse teratocarcinomas as a model with which to investigate his interest in the concept of flexible differentiation; this then permitted our use of embryonic stem cells (ESCs) shortly after they were first reported by Martin Evans in 1981 (Evans and Kaufman, 1981). Yoshio Hamada and others from the school made full use of ESCs to knock out their favorite genes. Tadao Atsumi established a monolayer culture line from the embryoid body cell line OTT6050, and this facilitated the discovery of E-cadherin by Masatoshi Takeichi. Although induced pluripotent stem cells were only produced many years later by Shinya Yamanaka, the ideas underlying their isolation had already been introduced to developmental biologists in Japan under the prevailing influence of TSO.
When the cloning age arrived in the late 1970s, TSO was eager to introduce molecular biology to the study of developmental biology. He invited Kunio Yasuda and one of us (H.K.) to join his group as assistant professors, asking us ‘do anything challenging, with the condition that it involves the keywords “genes” and “lens” ’. Yoshiro Shimura provided technical supervision during the cloning of crystallin genes. We were given tremendous liberty, but were subject to monitoring by TSO’s extraordinarily sharp eyes, being told ‘Stop it, it’s trivial’, as soon as we developed irrelevant ideas. One successful outcome was the demonstration that the chicken δ-crystallin gene is correctly regulated in a lens-specific manner in mouse cells, indicating the existence of evolutionarily conserved lens-specific gene regulatory mechanisms (Kondoh et al., 1983). This study developed further, leading to the discovery of Sox2 and Pax6 as interacting transcription factors for the initiation of lens development (Kamachi et al., 1995, 2001), and the identification of the Maf family of transcription factors as essential regulators of lens maturation (Ogino and Yasuda, 1998). The electroporation technique for gene manipulation in chicken embryos was also developed along this line (Nakamura, 2009).
Masatoshi Takeichi, who was then an associate professor, set forth to characterize Ca2+-dependent, trypsin-sensitive adhesion molecules and discovered the cadherins (Takeichi, 1986, 1988), while Hajime Fujisawa (who left the group at an early stage of the 10-year period) later discovered neuropilin and plexin (Satoda et al., 1995; Takagi et al., 1995). Although TSO did not participate directly in the molecular characterization of cell-cell interactions, he successfully furnished his laboratory with an environment to encourage such investigations.
The flexibility of the differentiated cell state is perhaps best manifested during tissue regeneration. Thus, modern studies of regeneration using planarians, pioneered by Kiyokazu Agata and Kenji Watanabe (Agata and Watanabe, 1999), can be regarded as a direct reflection of TSO’s interests. In a similar vein, many researchers who joined the TSO school have developed their individual talents and have been successful in various branches of developmental biology.
TSO had planned to keep the laboratory in Kyoto for several more years, but this did not happen. Haruo Kanatani, the Director of the National Institute for Basic Biology (NIBB) in Okazaki, who had also been a friend of TSO at Konan High School, died an untimely death, and TSO was asked to succeed him. He accepted the NIBB Director position and left Kyoto in 1984. He re-formed a tag team with his former colleague Goro Eguchi, who was by then a professor there investigating pigment cell-derived lens development. TSO compiled studies on transdifferentiation and related phenomena in a volume of Current Topics in Developmental Biology (Okada and Kondoh, 1986), and summarized his work in the book Transdifferentation (Okada, 1991).
During his six years in Okazaki, TSO further promoted international collaborations among developmental biologists; he organized many international meetings on different themes in Japan and other Asian countries. These meetings provided hubs for the interaction of developmental biologists on a global scale during the period when international meetings were less frequent than they are today. The small scale of these meetings facilitated trans-generational discussions among participants from different backgrounds. His dedication toward forming global links presumably compensated for his loss of laboratory activities during the period.
For 10 years from 1993, TSO was the Director of the Biohistory Research Hall in Takatsuki, a newly opened private museum owned by Japan Tobacco, which was located midway between Kyoto and Osaka. TSO, together with Vice Director Keiko Nakamura, enjoyed operating this research museum. The research section covered the embryonic and phylogenetic development of various non-mammalian animals, while the museum section aimed to expose a wide audience – ranging from elementary school pupils to nonprofessional biology lovers – to the wonder and beauty of developmental processes. Different types of exhibitions and small concerts were part of the museum’s events and were an amalgamation of his enthusiasm for science and music. This was a joyful period for TSO, allowing him to fully express his esthetics. In 2007, TSO received the Order of Cultural Merit, the most prestigious award in Japan.
TSO had various and serious interests in subjects other than developmental biology. One example was his collection of longicorn beetles. His most profound interest was in Western classic music, and he wrote many critiques on 20th century compositions. His son, Akeo Okada, is a professor of musicology at Kyoto University. In the same way that various elements of his broad scientific interests were elaborated by his colleagues and students, one of TSO’s talents was clearly passed on to his son.
The life of Tokindo S. Okada was rich, influential and joyful. He was an exceptionally attractive and great mentor. We miss him, but he lives vividly in our memories.
References
Agata, K. and Watanabe, K. (1999). Molecular and cellular aspects of planarian regeneration. Semin. Cell Dev. Biol. 10, 377-383.
Araki, M. and Okada, T. S. (1977). Differentiation of lens and pigment cells in cultures of neural retinal cells of early chick embryos. Dev. Biol. 60, 278-286.
Evans, M. J. and Kaufman, M. H. (1981). Establishment in culture of pluripotential cells from mouse embryos. Nature 292, 154-156.
Hadorn, E. (1968). Transdetermination in cells. Sci. Am 219, 110-114.
Iida, H., Ishii, Y. and Kondoh, H. (2017). Intrinsic lens potential of neural retina inhibited by Notch signaling as the cause of lens transdifferentiation. Dev. Biol. 421, 118-125.
Kamachi, Y., Sockanathan, S., Liu, Q., Breitman, M., Lovell-Badge, R. and Kondoh, H. (1995). Involvement of SOX proteins in lens-specific activation of crystallin genes. EMBO J. 14, 3510-3519.
Kamachi, Y., Uchikawa, M., Tanouchi, A., Sekido, R. and Kondoh, H. (2001). Pax6 and SOX2 form a co-DNA-binding partner complex that regulates initiation of lens development. Genes Dev. 15, 1272-1286.
Kondoh, H., Yasuda, K. and Okada, T. S. (1983). Tissue-specific expression of a cloned chick delta-crystallin gene in mouse cells. Nature 301, 440-442.
Le Douarin, N. (1973). A biological cell labeling technique and its use in experimental embryology. Dev. Biol. 30, 217-222.
Nakamura, H. (2009). Electroporation and Sonoporation in the Study of Developmental Biology. Tokyo: Springer Japan.
Ogino, H. and Yasuda, K. (1998). Induction of lens differentiation by activation of a bZIP transcription factor, L-Maf. Science 280, 115-118.
Okada, T. S. (1960). Epithelio-mesenchymal relationships in the regional differentiation of the digestive tract in the amphibian embryo. W. Roux Arch. EntwMech. Org. 152, 1-21.
Okada, T. S. (1965). Immunohistological studies on the reconstitution of nephric tubules from dissociated cells. J. Embryol. Exp. Morphol. 13, 299-307.
Okada, T. S. (1991). Transdifferentiation. Oxford: Clarendon Press.
Okada, T. S. and Kondoh, H. (ed.) (1986). Commitment and Instability in Cell Differentiation: Current Topics in Developmental Biology, Vol. 20. London, UK: Elsevier.
Okada, T. S., Ito, Y., Watanabe, K. and Eguchi, G. (1975). Differentiation of lens in cultures of neural retinal cells of chick embryos. Dev. Biol. 45, 318-329.
Okada, T. S., Yasuda, K., Araki, M. and Eguchi, G. (1979). Possible demonstration of multipotential nature of embryonic neural retina by clonal cell culture. Dev. Biol. 68, 600-617.
Okada, T. S., Nomura, K. and Yasuda, K. (1983). Commitment to transdifferentiation into lens occurs in neural retina cells after brief spreading culture of the dissociated cells. Cell Differ. 12, 85-92.
Sato, Y., Yasuda, K. and Takahashi, Y. (2002). Morphological boundary forms by a novel inductive event mediated by Lunatic fringe and Notch during somitic segmentation. Development 129, 3633-3644.
Satoda, M., Takagi, S., Ohta, K., Hirata, T. and Fujisawa, H. (1995). Differential expression of two cell surface proteins, neuropilin and plexin, in Xenopus olfactory axon subclasses. J. Neurosci. 15, 942-955.
Steinberg, M. S. (1970). Does differential adhesion govern self-assembly processes in histogenesis? Equilibrium configurations and the emergence of a hierarchy among populations of embryonic cells. J. Exp. Zool 173, 395-433.
Takagi, S., Kasuya, Y., Shimizu, M., Matsuura, T., Tsuboi, M., Kawakami, A. and Fujisawa, H. (1995). Expression of a cell adhesion molecule, neuropilin, in the developing chick nervous system. Dev. Biol. 170, 207-222.
Takeichi, M. (1986). Molecular basis for teratocarcinoma cell-cell adhesion. Dev. Biol. 2, 373-388.
Takeichi, M. (1988). The cadherins: cell-cell adhesion molecules controlling animal morphogenesis. Development 102, 639-655
Applications are invited for a postdoctoral research assistant/associate to join the group of Dr Emma Rawlins at the Gurdon Institute, University of Cambridge to work on the regulation of human lung development with the long-term aim of developing strategies for regenerative medicine (http://www.gurdon.cam.ac.uk/research/rawlins). Our recent research has focused on cellular mechanisms of lung development, homeostasis and repair using the mouse as a model system (e.g. Balasooriya et al., Dev Cell 2016; Laresgoiti et al., Development 2016; Watson et al., Cell Reports 2015). We have now established novel in vitro systems for studying human lung development which will be the primary focus of our work over the next few years. We aim to recruit an outstanding individual who is interested in lung developmental mechanisms, their contribution to disease development and therapeutic potential. The research will combine the application of single cell techniques to human embryonic lungs with the in vitro genetic analysis of mechanisms using our organoid system.
Applicants should have a PhD in a relevant subject, or be close to completion of their degree. Expertise in general areas of developmental/stem cell biology including single cell biology, live cell imaging, image analysis and cell signalling mechanisms would be suitable for these positions. Experience of in vitro models and use of experimental animals would be an advantage.
Salary: £29,301-£38,183
Closing date: 15 June 2017
For further information and to submit an application for this vacancy visit
The recent bloom of genomic data from all of life’s kingdoms is revealing a novel perspective of gene loss as a pervasive source of genetic variation with a great potential to generate phenotypic diversity and to shape the evolution of gene networks. How do genes become dispensable and subsequently lost? Are patterns of gene loss stochastic or biased? What is the effect of gene loss on the evolution of gene networks? What is the influence of gene loss on the evolution of mechanisms of development (that is, ‘Evo-Devo,) and the diversification of species?
I am Alfonso Ferrández a PhD student working in the group of Prof. Cristian Cañestro and Prof. Ricard Albalat (@EvoDevoGenomeUB) in the Section of Biomedical, Developmental and Evolutionary Genetics in the Department of Genetics (@GeneticsUB), Microbiology and Statistics, and in the Institute of Biodiversity Research (@IRBioUB), of the University of Barcelona, Spain (Fig. 1). We are trying to address some of the previous questions related to gene loss in the field of Evo-Devo using a curious chordate called Oikopleura dioica, which despite having suffered an extreme process of genome compaction along with massive gene losses, still preserves a typical chordate body plan.
Fig. 1: The 2017 Barcelona Oikopleura team.
Why did we choose O. dioica?
Fig. 2: O. dioica inside the mucous house.
Ecological relevance: O. dioica is a peaceful filter-feeding and free-swimming planktonic organism, about 2-3 mm long. It lives inside a secreted mucous house that it uses as a food trapping device by filtering the water current propelled by its stylish and grooving tail movements (Fig. 2). O. dioica is the only known urochordate species that have separated female and male individuals, which are indistinguishable until maturity. O. dioica has a cosmopolitan world-wide distribution including seas of Europe, Asia and America and it is so abundant in the zooplankton community that plays a key role in marine trophic webs serving as food for fish larvae. Moreover, because of the small size of the pores of their mucous houses, they can also trap the smallest microalgae, thus creating a short circuit that accelerates the transference of organic matter both through the marine trophic web and towards vertical flux of carbon-rich organic material (i.e. marine snow) that sinks to the bottom of the oceans.
Phylogenetic position within our own phylum: O. dioica belongs to the larvacean class inside the urochordate subphylum, the closest sister group to vertebrates. Urochordates diverged prior to the two rounds of whole genome duplication (2R-WGD) that occurred early in the vertebrate lineage (Albalat and Cañestro, 2016; Cañestro, 2012), and consequently, the mutational robustness of most gene networks appears to be much lower in O. dioica than in vertebrates (Fig. 3). The availability of many deeply sequenced genomes of several other chordates –3 species of cephalochordate, 10 urochordate ascidians, and >100 vertebrates– provides a perfect phylogenetic framework for the identification of gene loss events in O. dioica by comparative genomics with other chordates.
Fig. 3. Chordate phylogeny. O. dioica belongs to the Larvacean class inside the Urochordate subphylum, sister group of vertebrates.
Genomic plasticity: O. dioica has the smallest chordate genome known so far with only 65Mb (even smaller than the 175 Mb of Drosophila or the 100 Mb of C. elegans), which results from an extensive process of compaction that has been accompanied by an extraordinary amount of gene losses. One striking example is the loss of all genes of the non-homologous end joining DNA repair system, plausibly one of the reasons that accounts for the elevated propensity for gene loss of this organism. Among key developmental genes, O. dioica has lost more than 30% of the homeobox gene groups, including all central Hox genes, and key genes involved in retinoic acid (RA) signaling (Albalat and Cañestro, 2016; Cañestro et al., 2006; Denoeud et al., 2010; Edvardsen et al., 2005; Martí-Solans et al., 2016; Seo et al., 2004).
A simple and transparent model for Developmental Biology: The embryonic development of O. dioica is very fast, and in less than 20 hours a transparent juvenile already shows a typical chordate body plan with organs that are unequivocally homologous to those in vertebrates, including a notochord anchoring muscle cells throughout a post-anal tail, a dorsal neural tube, brain, thyroid, pituitary, gill slits, pharynx, esophagus, gut and a heart (Fig. 4). In addition, O. dioica shares with other urochordate species, such as ascidians, a very similar embryonic developmental program, both at the morphological and molecular level, with the important difference that O. dioica does not suffer the drastic metamorphosis that ascidians do, and therefore, in contrast to ascidians, maintain all chordate features throughout its life.
Fig. 4. Organ homologies between an Oikopleura and a zebrafish larvae.
A day in the life of an Oikopleura lab
Our laboratory is one of the few laboratories in the world able to culture O. dioica all year round, which means almost unlimited availability of biological material (i.e. mature males and females, eggs, sperm, embryos and larvae) to carry out functional experiments. We maintain them at 19ºC, which results in a generation time of only 5 days (Fig. 5). Our lab has set up a low-cost maintenance regime by reducing as much as possible the amount of water, space and manpower to handle the animals (Martí-Solans et al., 2015). All these characteristics, together with its high fecundity and transparency makes it an attractive model for developmental studies.
Fig. 5. O. dioica facility in the University of Barcelona (Catalonia, Spain). (a) Animals were collected in the coast of Catalonia near Barcelona using a plankton net or directly with a bucket. (b) Seawater is filtrated at 50–20 µm (fSW) in the facility to remove excess of sand particles that could affect O. dioica buoyancy. (c) The production of the four microalgae for O. dioica feeding (Bouquet et al., 2009) was scale down in an adaptable fashion to the weekly needs of the facility (round-bottom glass flasks in upper shelves). Long-term stocks (100 mL Erlenmeyers in lower shelves) were renewed just once per month. (d-g) The use of agar plates provides an alternative method to maintain long-term microalgal stocks: (d) Isochrysis sp., (e) Chaetoceros calcitrans, (f) Rhinomonas reticulata and (g) Synechococcus sp. (h) O. dioica animals were maintained in suspension by the rotation of a paddle driven by a motor mounted on the lid of polycarbonate beakers. (i) Animal lines were maintained in a small room (5 m2) in four shelves (1.5 m2) at 19°C using a standard air conditioner device. (j) Protocol of husbandry. (Martí-Solans et al., 2015).
A typical day in the Oikopleura lab starts looking for ripe animals to mate and start a new generation (this happens early in the morning, since we have synchronized the animals to mature at that time of the day). At day 5 of their cycle, males and females are easily distinguishable. Males have a yellowish gonad full of sperm, whereas females have a refringent and translucent gonad full with 100 to 400 eggs (Fig. 6 K). Since O. dioica has external fertilization, to mate them we only have to put together about 20 females and 10 males, and wait for their spontaneous spawn that will give rise to the next generation. Both males and females naturally die after the spawning, which unfortunately does not allow us to keep the parental generations. Next day (day 1), we are normally happy to see hundreds of juveniles beating their tales inside their already inflated houses. Yes! They have a brisk development! The first division occurs just 20 minutes after fertilization; by 4 hours after fertilization (hpf) they break their chorion, and few minutes after the hatch we can already see them graciously twitching their tails in their first attempts to swim. By 5 hpf the tail movements are rhythmic and harmonious, which allow them to swim up and being suspended in the column of water. By 8 hpf, the heart is vigorously beating, and the ciliary rings are working at full speed creating the water to circulate through the pharynx. By 9 hpf, the animals do the tailshift, characterized by the shift of the tail to an acute angle relative to the trunk, flagging the end of embryonic development, and competence to secrete and inflate their first filter-feeding house (Fig. 6).
From day 1 to day 5, we need to feed them every single day (weekends and bank holidays included, aggh!), and to transfer them to fresh seawater to keep them happy, and not too crowded. Their diet consists of a mix of four different species of algae that we also grow in the lab, at different ratios depending on the size and needs of the animals each day of the culture (Fig. 5 c-g).
Functional approaches of gene knockdown or inhibition are amenable. The syncytial gonad of females is easy to inject with RNAi, morpholinos or even, the recent discovered DNAi (yes, dsDNA rather dsRNA of your target gene…cheap and effective), obtaining a massive generation of knockdown embryos (Omotezako et al., 2015). Moreover, permeability and small size of O. dioica embryos allow us to easily treat them with drugs or specific inhibitors of signaling pathways to modify their developmental programs.
Fig. 6. Embryo development in O. dioica is very fast. (A) Two cell estage embryo 30 minutes post fertilization. (B-D) From 2 to 4 hours post fertilization (hpf) we can identify the tailbud stage in which the embryo resides inside the corion. (E-H) At 4 hpf the embryo leaves the corion and became a swimming larvae during the hatchling stages. (I) The metamorphosis, that only consist in a 180º rotation of the tail, takes place 9 hpf giving rise to the Tailshift embryo. (J) Adult animal of 2 days of life. (K) Adult animals of 5 days of life. The female gonad contains hundreds of eggs, the male gonad is dark and contains the sperm.
Current research lines
To address the fundamental question of how gene loss affects the evolution of the mechanisms of development, as a case study, our research focuses on the analyses of the striking loss in O. dioica of the retinoic acid (RA) signaling pathway, which is conserved and essential for many developmental and physiological roles in all other known chordates. First, we have described a process of gene co-elimination of nearly the entire classic metabolic and signaling pathways. This analysis allowed us also to recognize surviving genes to the dismantling of those pathways, and to recognize processes of neofunctionalization and hidden pleiotropy as the probable causes that preserved the genes of vanishing. Currently, our focus of attention is on O. dioica heart development, since RA plays a fundamental role in all other chordates. Finally, two new lines of applied research are starting to fly in our lab, using O. dioica as an evolutionary knockout model to study the genetic bases of some human cardiomyopathies, as well as a model to better understand the limits of the genetic responses of chordate development towards environmental threats from anthropogenic origin such as heavy metal from industrial wastes or global warming (but these are two long new stories that would need another thread in the Node).
References
Albalat, R., Cañestro, C., 2016. Evolution by gene loss. Nat. Rev. Genet. doi:10.1038/nrg.2016.39
Research assistant position for subsequent appointment as PhD fellow in ‘Epithelial cell renewal’ to join the Sedzinski lab.
The Danish Stem Cell Center (DanStem) at Faculty of Health & Medical Sciences at the University of Copenhagen is looking for a Research assistant subsequent appointed as PhD fellow to join the Sedzinski group starting September 2017 or upon agreement with the chosen candidate.
The position as Research assistant is for 1 year. The position as PhD fellow is for 3 years.
DanStem comprises of two sections: The Novo Nordisk Foundation Section for Basic Stem Cell Biology, where we address basic research questions in stem cell and developmental biology (BasicStem). The second Section for Strategic Translational Stem Cell Research and Therapy (TransStem) is focused on the translation of promising basic research results into new strategies and targets for the development of new therapies for cancer and diabetes. Find more information about the Center at http://danstem.ku.dk
We are seeking a highly motivated and ambitious candidate to join the Sedzinski lab with the following project:
Job description
The Sedzinski lab (http://danstem.ku.dk/research1/sedzinski-laboratory/) is interested in understanding mechanics of epithelial tissue homeostasis and morphogenesis. Particularly, we want to determine both the mechanics and molecular regulation of epithelial cell renewal. For this, we study how forces generated by the actomyosin cytoskeleton and adhesion molecules shape and move epithelial cell progenitors within tissues. We use high-resolution microscopy to image dynamics of progenitor cells, biophysical and theoretical methods to describe the forces, and genetic to perturb the system.
We are seeking highly motivated and ambitious candidates to join our team.
Qualifications
Candidates must hold a master’s degree in biology, biophysics, biochemistry, bioengineering, bio-informatics, or similar, and possess a general understanding of cell and developmental biology.
Previous practical experience in quantitative biology, biophysics, computational biology, microscopy, and image processing is considered of great advantage.
Publications and practical experience are beneficial.
Good English communication skills, both oral and written, are prerequisite for the successful candidate
Terms of salary, work, and employment
The employment is for 4 years, as research assistant is for 1 year and as PhD fellow for the following 3 years, and is scheduled to start on September 2017 or upon agreement with the chosen candidate. The employment as a PhD student is conditioned upon a positive assessment of the candidate´s research performance and enrolment in the Graduate School at the Faculty of Health and Medical Sciences. The PhD study must be completed in accordance with the ministerial orders from the Ministry of Education on the PhD degree and the University´s rules on achieving the degree.
The place of work is at DanStem, University of Copenhagen, Blegdamsvej 3B, Copenhagen. Salary, pension and terms of employment are in accordance with the provisions of the collective agreement between the Danish Government and AC (the Danish Confederation of Professional Associations). In addition to the basic salary a monthly contribution to a pension fund is added (17.1% of the salary).
The application must include
1. Motivation letter
2. Curriculum vitae incl. education, experience, previous employments, language skills and other relevant skills
3. Copy of diplomas/degree certificate(s)
Questions
For further information about the position please contact group leader Jakub Sedzinski, jakub.sedzinski@sund.ku.dk
How to apply
The application, in English, must be submitted electronically by clicking APPLY below.
The University of Copenhagen wishes to reflect the diversity of society and welcomes applications from all qualified candidates regardless of personal background.
Only applications received in time and consisting of the above listed documents will be considered.
Applications and/or any material received after deadline will not be taken into consideration.
The application will be assessed according to the Ministerial Order no. 284 of 25 April 2008 on the Appointment of Academic Staff at Universities.
Assessment procedure
After the expiry of the deadline for applications, the authorized recruitment manager selects applicants for assessment on the advice of the Appointments Committee. All applicants are then immediately notified whether their application has been passed for assessment by an expert assessment committee. Selected applicants are notified of the composition of the committee and each applicant has the opportunity to comment on the part of the assessment that relates to the applicant him/herself. You can read about the recruitment process at http://employment.ku.dk
Application deadline: June 30 2017
Founded in 1479, the University of Copenhagen is the oldest university in Denmark. It is among the largest universities in Scandinavia and is one of the highest ranking in Europe. The University´s eight faculties include Health Sciences, Humanities, Law, Life Sciences, Pharmaceutical Sciences, Science, Social Sciences and Theology.www.ku.dk
Innovations in science are happening all over the nation! Visit the science videos at the 2017 STEM for All Video Showcase, funded by the National Science Foundation. Interact with the community by asking questions, getting answers, and voting for your favorite video! Support STEM education and let your voice be heard from May 15-22 at the STEM for All Video Showcase!
Your vote counts!
Did you see an amazing video at the showcase? Let us know! You can vote for your favorite video through Twitter or the showcase website. The video with the most votes will be given the “Public Choice” award on Tuesday, May 23, and the winning participants will be recognized by the National Science Foundation!
Do you support STEM? Let the world know!
You make the difference. Please share the STEM for All Video Showcase with your friends and colleagues, on social media, and with your local science centers. Help us spread the word about the amazing people working in STEM education and the awesome work they are doing for our communities!
More details on the event:
The Stem for All Video Showcase is funded by the National Science Foundation. This year’s theme is “Research & Design for Impact.” In short 3 minutes videos, leaders of STEM education initiatives describe how their projects are meeting some of today’s most pressing scientific and technological challenges. The STEM for All Showcase is a collaborative effort of the following NSF resource centers: MSPnet, CADRE, CAISE, CIRCL, STELAR, CS For All Teachers. It is funded by the National Science Foundation (#1642187).
The Showcase is powered by the Videohall.com platform developed by TERC, a STEM-focused education research nonprofit based in Cambridge, MA.
I had started to become a little worried when I didn’t see Eric on the opening day of the conference, but it turned out that his plane to Germany had been delayed by the snowstorms blanketing the Eastern seaboard of the US and he made it in the end. Between sessions later on, we found a slightly chilly empty computer lab to sit down in, and over the course of an hour talked about his history in research and views on science in general. My lasting impression was of an enthusiasm and passion for research that was by equal measure refreshing and infectious – it made me want to look down a microscope again!
Eric Wieschaus is a HHMI Investigator, and the Squibb Professor in Molecular Biology at Princeton University and the Lewis-Sigler Institute for Integrative Genomics. In 1995, he was awarded the Nobel Prize jointly with Edward Lewis and Christiane Nüsslein-Volhard for discoveries about the genetic control of Drosophila embryogenesis. We caught up with Eric at the joint meeting of the German and Japanese Societies of Developmental Biologists held in Kiel in March 2017, and discussed his career, his thoughts on the field and the impact the Nobel award had on his life.
Eric in the lab
I understand that after your first experience with Drosophila – as an undergraduate in Harvey Bender’s lab at Notre Dame – you said never wanted to see a fly again! What led you to change your mind?
I suppose the formative undergraduate experience wasn’t washing dirty fly bottles and making fly food, which is what I had done in Harvey Bender’s lab, but an embryology course where we looked at frog and chick embryos, which were the embryos people worked with at the time. I knew then that I wanted to be an embryologist, to study development. But I think I just didn’t know that flies had embryos! Anyway, I ended up at Yale in New Haven, and for various reasons I was assigned to the lab of Donald Poulson. Poulson had, in the 1930s and 1940s, first as a graduate student and through to his professorship, described all of Drosophila embryogenesis. So I knew that I wanted to study embryos, and I had by chance landed in the laboratory of the one person in the world who really knew how Drosophila embryos develop. Poulson was a very kind person: he took me in to his lab when he specifically didn’t want to have a graduate student. It took him a little while to find a young assistant professor who was willing to take me on, and that turned out to be Walter Gehring.
And as a graduate student with Gehring, you were trying to map cell lineage in Drosophila with somatic clones and disc transplantation. What was the drive behind this work?
Walter had developed a tool to culture embryos, based on the strategies people had used to culture imaginal discs. He was able to convince me, a naïve young graduate student entering the lab, that the best experiment in the world would be to isolate a single cell from the blastoderm, grow it in these culture conditions and measure its developmental capacity, to establish whether it had already undergone some determination or programming. And that was my project – I spent my first three years grinding up embryos and trying to get single cells to grow. I have to say that none of that ever worked, but at some point I decided that I needed to have a control – once I had these wonderful cultures going, and these cells displaying their inherent potential, I wanted to be able to compare them to what a blastoderm cell actually did if left in situ. I decided I would produce clones at the blastoderm stage and follow those cells, but I never really looked at them because I still believed my experiments would work. But at the end of four and a half years, when I realised that I wasn’t going to have cultured cells as part of my thesis, I decided to go back and look at the controls. Luckily, it turned out that what cells normally do in development is almost as interesting as what you can get them to do when you artificially manipulate them.
When Walter’s lab moved to Basel, you were brought into contact with Christiane (Janni) Nüsslein-Volhard. How did that work out?
Janni and I both finished our thesis work at around the same time – she was in Tübingen, and having done molecular biology was an attractive person for Walter to bring to his lab. She actually came to Basel specifically not to do molecular biology, but instead to learn how to look at embryos, and as I was the only person in the lab who was working with embryos, we bonded over that in the four months before I left. And later, we both were fortunate to get positions in Heidelberg – these were real group leader positions, but we didn’t know that we had to bargain for space! So we ended up together, sharing a lab, and we already knew kind of what we wanted to do; it was a perfect set of circumstances.
And in Heidelberg, you began work on the screen that would eventually win you the Nobel Prize. Screens obviously require a lot of time and effort: how did you stay focussed and cope with the practical demands at the time?
Well it was a lot of work, but the payoffs were real. We spent a certain amount of time figuring out how to do the screen, and that was maybe more frustrating than actually doing it. But once we started the big screen, scaled up and lasting about two and a half months, we discovered something new every day, and so focus was not at all hard. We would work from nine in the morning to around midnight, pretty much every day of the week, but the payoff was already there – not in understanding as such, but in the realisation that we were seeing stuff that no one had ever seen before, and thinking thoughts that no one had ever thought before. I don’t even remember being frustrated or wanting to slow down at all – my memory of that time is just of the fascination and excitement of doing it.
We were seeing stuff that no one had ever seen before, and thinking thoughts that no one had ever thought before
Was there a key lesson the screen taught you about how development works?
Before we did the screen, we really didn’t know what we would get out of it. The two dangers were that every gene that you mutated would mess up development, and that the phenotypes would be complicated and heterogeneous and impossible to interpret. But what actually happened was that the number of zygotically active genes we found was small – about 120 to 130 – and the mutants had unique phenotypes that identified specific processes, which meant you could immediately group things. This small number contrasted with the characterisation of RNA heterogeneity carried out at the time, which estimated there were thousands of different mRNAs present in the embryo, but most of these turned out to come from the mother. So the conceptual lesson was that the embryo uses transcription of a small number of genes to drive specific events forward. If gene products can be supplied to all cells, they’re provided maternally, but if you want to turn the gene on at a specific place or at a specific time, zygotic transcription does that. That was the power of the screen, to pick up these regulators: the genes controlling developmental decisions. It seems quite obvious now, but did I predict that at the time? Absolutely not.
Following the screen, you moved your lab to Princeton in 1981. What did you hope to achieve in the early days in New Jersey?
Initially, when we returned to Princeton, I helped Trudi (Schüpbach) a bit when she was setting up her maternal effect screen to find the determinants deposited in the egg by the mother. But what I really wanted to understand was morphology, and this very quickly became a cell biological problem, of how cells move and change shape and so on. This obviously required a lot of observation, and I remember at the time the great discovery was that there was something called phalloidin (which labels F-actin) – it was fluorescent, and you could buy it from a supplier! You could get a fluorescence microscope and just look – it really allowed you to see embryogenesis differently. So this period was also a time where, as well as advances in molecular biology, there were great advances in cell biological reagents, which you could begin to apply to embryos to think about these processes from a more cell biological perspective. It all came together – the observation of the real things that cells do, and their manipulation by genetics (a tool that cell biologists did not have at the time).
And how did you approach the cell biological side of development, particularly since many of the factors involved would have been missed by the screen?
The screen picked switches, decision-making mechanisms: most of these are transcription factors and the cell signalling pathways that govern them. Once you go downstream of decisions, it becomes more complicated and more synthetic, and less easily approachable by a genetic screen that looks for survival, for instance. I think that genetic manipulations are still really powerful here, but you can’t take shortcuts like only looking for genes essential for viability. A number of biological processes go into a change in cell shape, for example, and if you remove one of them, the cell shape transition may not occur in the same way, but the animal may be robust enough that a proportion of embryos survive and you’ll miss it. So understanding morphology turns out to be a process where you use genetics, but need other tools too, and much of this concerns imaging. I think that a major transformation in biology has been the development of microscopes of different kinds, whether confocal or light sheet, or whatever. And digital cameras! To those of us who can remember, images used to be things you had on film and you would spend artistic moments in the darkroom trying to bring out what you wanted; now they are just a matrix of pixels. And I think that aspect – doing microscopy on embryos, and doing it in a quantitative way – meant you learned stuff that you didn’t know before. It’s just transformed the field.
The Wieshcaus lab’s focus on the cell biology of development includes work on nucleolus assembly. In Falahati, et al. (Current Biology, 2016), the dynamics of the nucleolar protein Fibrillarin were tracked in a mutant embryo lacking rDNA repeats
To look at embryos with a quantitative eye, your lab has in recent years incorporated physics and mathematical modelling. From your background, to what extent do you personally need to understand the physics and the maths?
Well I’m totally dependent on having collaborators who have the patience to explain things to me three or four times, and also the tolerance to accept when I decide that I’ve got it, even though there might be a deeper level of understanding available that they feel I haven’t reached yet. But it’s also an interesting question to ask what your thresholds are for understanding: I think you set them, and that sets the style of science that you do. The fact that different scientists have different thresholds is one of the reasons why it is so valuable to collaborate, and also why the social nature of science is so important to its productivity. You profit from talking to people who are interested in working with you, but frankly not willing to invest the same amount of energy into your topic as you do. Of course there’s always danger in not understanding everything, but we live in a dangerous world. Scientifically, if you are going to be on the edge of discovery, you just take the best path forward even if you don’t understand everything in a deeply defendable way.
As someone who has worked on Drosophila for 40 years, what do you think this animal still has to teach us about development?
Science is hard, and in biology we are only at the beginning of a truly deep understanding. To understand development, you work on embryos, and you choose a model organism because to do things well, you need the technologies that everybody else has developed; you need to parasitise a field.
Working with Drosophila today, you have two avenues. If you want to strike out into some totally unexplored area, for instance in something behavioural or evolutionary, flies allow you to do this. And then there are the old areas, things that we think of as established and successful problems, like patterning in the early embryo. What we have there is actually a wonderful cartoon understanding of the process, and this is so rare and valuable in biology because it can provide the basis for building a deeper understanding. So if you want to understand transcription, for example, I think you’d be crazy not to work in flies! In flies you have this organism where you can study a real process with biological meaning that has been selected for in evolution, and which is primed by our existing knowledge for you to start. So the things that we know prime us to go deeper, and for the things that we don’t know, we’re in a position where we can profit from the community and the tools that are available.
In biology we are only at the beginning of a truly deep understanding
It’s now 22 years since you won the Nobel Prize along with Christiane Nüsslein-Volhard and Edward Lewis. What did the award mean to you at the time, and does it still have an impact on your academic life?
Of course it was a wonderful thing to happen for me and my family. It wasn’t really anything I’d foreseen, and then I got ripped out of my normal life for a week in Stockholm. But I also felt like it was wonderful for the community as a whole: at least from my perspective, my colleagues were happy because they saw it as recognition of a set of accomplishments that the field had made.
The other thing that it gave me was a certain power to control my work life. I’ve never assumed major administrative roles at the university – it gave me the power to insist that I would continue to work four hours a day at the bench myself, and meant that people were happy to have me around even if it forced some others to assume extra responsibilities. I could set certain standards for my life: I wanted to be a productive scientist, not to just run a lab and tell other people what to do. I wanted to be the person who did interesting things.
You’ve described yourself as a very visually oriented person, both in terms of science and as someone who loved to draw and paint in your youth. Does this dictate the way you do your science?
Since the public is giving their own money to support science, there are limited resources, an obligation to produce and a certain competition among scientists. So as a young scientist you have to ask yourself – what is my competitive advantage? What am I good at? And almost inevitably that means: what do I enjoy? And the answer will go deeper than science itself. So when I look at most things that I’ve done in the lab, they’ve always had a visual component to them. In the Heidelberg screen, we identified mutants based on our abilities to look at things and recognise what was worth following up on. And to look at gastrulation is a matter of reconstructing and visualising form, and form follows force, so an understanding of physics, of flow and force and mechanics, has, to my mind, at least a strong visual component to it. I am the scientist that I am because of the talents and particular proclivities I have, and clearly painting was another expression of that same quality.
And do you still paint?
I’ve got back in to painting, but it’s challenging because I think to be really good, you have to invest more time than I have. I’m still too hung up on the idea of getting a product at the end, getting the picture to work, rather than doing a hundred versions of it to figure out how to get it to work. Every step in becoming better costs time, which of course is not very different from science, where to be really great, you have to invest and risk, and do things a bunch of times. That’s why science is not a nine to five job: so much of what we do doesn’t quite work out!
What would Development readers be surprised to find out about you?
I am an aggressive card player, and I play to win (though I must say don’t win all the time!). Trudi and I have trained all of our children to be aggressive pinochle players, and also to play to win. Pinochle is different from poker: with poker there is an element of bluff, and while I am a theatrical enough person that I could pull that off, I feel so emotionally dishonest! So I tend towards card games like pinochle where you are confronted with a problem and have to figure out how to win with the cards you have. As long as I don’t have to be dishonest about it!
Check out Development’s collection of print interviews here, and our video interview collection here
A postdoctoral BBSRC funded position is available to study role of Par-3 in E-cadherin recycling and signalling in the group led by Dr Natalia Bulgakova http://www.sheffield.ac.uk/bms/research/bulgakova
The focus of the Bulgakova group is on discovering roles and regulation of cell-cell adhesion using Drosophila as a model organism. We have recently discovered that the protein Par-3 maintains the key cell-cell adhesion protein E-cadherin within the recycling route. The successful applicant will study the mechanism of this Par-3 function and its role in the development and physiology of the animal. The project will employ a combination of molecular genetics, live imaging, and protein biochemistry.
The successful applicant will have a PhD or equivalent experience relevant to studying cell-cell adhesion or intracellular trafficking, knowledge of working with genetic model systems and be highly motivated. Experience in using Drosophila, molecular biology, protein biochemistry, and confocal microscopy, and knowledge of cell-cell adhesion, cell polarity and intracellular trafficking will be an advantage. The successful candidate will be expected to have excellent interpersonal and communication skills, be highly independent, and committed to research in a fast-moving and competitive field.
Fixed-term with an immediate start date and an end date of 31 March 2020
For more information and to apply, please see http://www.jobs.ac.uk/job/BBL246/research-associate/
The publication of Marc Tessier-Lavigne’s seminal Cell papers (1, 2) in 1994 describing the identification of netrin1 (from the Sanskrit word, netr, meaning “one who guides”) was a defining moment in my graduate career. My friends and I talked about those papers for weeks, from the incredible technical feat, the biochemical purification of netrin1 from tens of thousands of chicken brains, to the commonality of the neural developmental mechanism, based on the homology of netrin1 with Unc6, a gene previously identified by Ed Hedgecock and Joe Culotti in a C.elegans screen for axon guidance defects (3).
A couple of years later, I interviewed for postdoctoral positions in San Francisco Bay area in neural development laboratories, and had arranged to stay with Marya Postner, a fellow Princeton graduate alumnus. Marya just happened to be married to Tito Serafini, who, with Tim Kennedy, was one of two lead postdocs on the seminal 1994 Cell papers. Tito was still working with Marc at the time. I asked him breathlessly about the status of the netrin1 project – did they have a netrin1 loss of function phenotype yet? Tito looked happy, triumphant even. “Yes!” he said. “We’ve knocked out the mouse homologue. Commissural axons stall before they reach the floor plate.”
This Cell paper came out a couple of months later (4) and cemented the idea of chemotaxis as the prevailing model of axon guidance (5): netrin1 is secreted from floor plate (FP) cells at the ventral midline of the spinal cord, and like a beacon in a harbor guiding ships in the night, orients commissural axons to grow towards it (Fig. 1A). This model fit with insight dating back as far as Cajal (6), including work performed in grasshoppers (7, 8) and then in vertebrates (9, 10). Together, these studies suggested that axons could be guided in a stepwise manner over long distances by chemotropic attractive or repellent cues diffusing from “guidepost” cells that they encountered along the way. The axon guidance field also recognized that axons also grew on local substrates provided by extracellular matrix components, such as laminin. But their contribution was generally considered prosaic, i.e. passive carpets that permitted or prevented axon growth. The more important and interesting contribution to neural circuit formation came from the chemotropic cues, the netrins, semaphorins, the slit/robo pathway and morphogens.
Problems with the chemotropic model for netrin1
I have taught the chemotropic model of axon guidance for years in my undergraduate and graduate lectures, with netrin1 as the centerpiece discovery. It was a beautiful example of scientific daring being rewarded with mechanistic understanding. I was able to trace a path from the first century-old scientific insight to the netrin1 mutant phenotype that strongly supported that hypothesis.
However after starting my own laboratory in 2004, I began to wonder about a reported, but underappreciated, feature of the netrin1 expression pattern. Textbooks often show the distribution of netrin1 in the chicken spinal cord, where netrin1 is expressed at high levels specifically in the FP. However, my first undergraduates – Joe Herrold and Anna Maglunog – found that mouse netrin1 is also expressed in the ventricular zone (VZ), which is the central compartment where the neural progenitor cells reside, oscillating back and forth on radial process as they proliferate. Spinal axons uniformly avoid growing in the VZ, staying rather at the “sides” of the spinal cord in a region that will ultimately segregate into the grey and white matter. Anna found that netrin1 expression extended into the dorsal VZ and appeared to strengthen, rather than diminish, over time (Fig. 1B).
This distribution had been accurately described in Serafini et al (4). But, it remained unresolved why spinal commissural axons first grew around the domain of VZ-derived netrin1 before growing towards FP-derived netrin1 (Fig. 1A). Were commissural axons unresponsive to ventricular netrin1? In that case, how did commissural axons then become responsive to FP-derived netrin1 to grow towards the ventral midline? Was there a molecular switch that controlled this process? Joe also noticed that in netrin1 mutant mice, while the vast majority of commissural axons of the Tag1+ subtype stalled as they entered the ventral spinal cord as published (4), there were always some Tag1+ axons that entered the VZ. Again, this latter phenotype was reported by Serafini et al, but it was not a major focus of the paper. We also wondered why spinal axons usually grew around the VZ. Was there a repellent in the VZ? Could netrin1 be that repellent?
I wrote some of these ideas into an R01 grant application that was eventually funded in 2008, which allowed me to bring a postdoctoral fellow on board to work on these questions. However, the project stalled for two years; the Tag1+ axon mispolarization phenotype was subtle and it remained stubbornly unclear whether these axons originated from within the spinal cord or from the dorsal root ganglia (DRGs) in the adjacent peripheral nervous system. Moreover, my plan to tackle the problem by recapitulating the putative VZ-repellent activity in a tissue co-culture assay proved challenging. No progress was made and alas, the postdoc left my lab. With time running out to fulfill this aim of my grant, I recruited a student, Supraja (Sup) Varadarajan onto the project with the idea that she could complete the characterization of the netrin1 phenotype. It would be a quick paper, I reassured her. One of the first ideas that we had was to characterize the netrin1 mutant using a wider range of axonal markers.
Figure 1. (A) In the canonical chemotaxis model, axons grow towards a diffusible source of floor plate (FP)-derived netrin1. (B, C) In our haptotaxis model, netrin1 is expressed by neural precursor cells in VZ (red domain) and then netrin1 protein is transported to the pial surface via their radial processes to form a growth substrate (green line). Axons extend adjacent to this substrate in a Dcc-dependent manner. (D) Axon growth is stalled, disoriented, and/or defasciculated in the absence of netrin1 (or Dcc). (E, F) Conditionally ablating netrin1 supports the haptotaxis model: VZ-derived netrin1, not FP-derived netrin1, is required to guide spinal commissural axons. Figure adapted from Varadarajan et al (11).
First moment of clarity: many types of spinal axons invade the VZ in netrin1 mutants
Not long afterwards, Sup called me over to her computer very excitedly: “LOOK at the pattern of neurofilament innervation!” she said. Neurofilament (NF) is an intermediate filament present ubiquitously in axons. Sup had made transverse slices of control and netrin1 mutant spinal cords and stained them with antibodies against NF. While control NF+ axons grew their usual orderly way avoiding the VZ, to our amazement we saw that NF+ axons were now profusely growing into the VZ in the netrin1 mutant (Fig. 1D, Fig. 2). Antibodies against another protein, Robo3, which labels all commissural axons, showed a similar phenotype; axons were no longer tightly bundled or fasciculated. Rather, the axons radiated in all directions, including into the VZ (Fig. 2). Thus, the observed stall of Tag1+ commissural axons appeared to be an anomaly: the loss of netrin1 resulted in other spinal axons extending wildly into the VZ. We suddenly had clear evidence that there might be a repellent in the VZ, with netrin1 as a top candidate for that repellent.
Sup then spent considerable time mapping domains of netrin1 expression in the mouse spinal cord at different stages of development. She found two general patterns of behavior: 1) spinal axons rarely grow on netrin1-expressing cells, and 2) spinal commissural axons grow precisely around the boundary of netrin1-expressing cells in the VZ. But were these activities mediated by a long-range activity from the FP or a more local, short-range activity from the VZ? Our genetic manipulations strongly supported the presence of a VZ-derived netrin1 repellent. First, we characterized Gli2 mutants, which have no FP (12) either singly or in combination with a netrin1 mutation. The result was clear-cut – it was not enough to remove the FP; NF+ axons only invaded the VZ in the absence of VZ-derived netrin1. Second, in collaboration with Jennifer Kong and Bennett (Ben) Novitch, we were able to ablate netrin1 expression from a stripe of neural progenitor cells, thereby creating two de novo netrin1(-):netrin1(+) boundaries. To our amazement, axons now detached from their normal trajectories and extended into the VZ to follow along one of the ectopic boundaries of netrin1 expression. Everyone in the Butler/Novitch joint lab meeting applauded when Sup showed these results for the first time.
Figure 2: Transverse section of a control (left of dotted line) and netrin1 mutant (right of dotted line) mouse embryonic spinal cord, showing NF+ (green), Robo3+ (red) and Tag1+ (blue) axonal staining. Figure adapted from Varadarajan et al (11).
Revised model: netrin1 provides a growth boundary for axon extension
Our first model was that netrin1, present in the VZ, was repulsive for axon growth. But our results were now suggesting a more complex activity. While spinal axons did generally avoid growing on netrin1-expressing cells, commissural axons appeared to grow preferentially along a netrin1(-): netrin1(+) boundary. Since there wasn’t an obvious term for this phenomenon, which was unwieldy to constantly explain, Sup came up with the concept of a “hederal” boundary. Ivy (genus, hedera) uses a wall (c.f. netrin1) as a necessary scaffold for growth, but it is unable to penetrate this wall as it grows. We wondered whether this hederal activity of netrin1 was more attractive or more repulsive, and tested this idea by examining mice lacking different classes of netrin1 receptors, sent to us by Artur Kania.
In the canonical model, netrin1 results in attractive or repulsive responses in axons by activating different receptor complexes. Thus, Dcc translates the attractive responses of netrin1 (13), whereas the Unc5 family mediates the repulsive responses (14). I was confident we would find that a member of the Unc5 family decoded the netrin1 growth boundary in axons, thus confirming that we were describing a repellent activity. But I was wrong: Sup found only minor axon guidance phenotypes in the Unc5 mutants, many of which had been reported before, and stemmed from the loss of Unc5 expression in the DRGs. However, the Dcc mutants looked just like the netrin1 mutants: Tag1 axons stalled, as already described (13) and NF and Robo3 axons dramatically extended into the VZ (Fig. 1D). Thus, Dcc appears to be the chief receptor that mediated the ability of spinal axons to avoid the VZ, and grow alongside a netrin1(-):netrin1(+) border. Moreover, these findings suggested that netrin1-Dcc might be working through an attractive, rather than repulsive, mechanism.
Second moment of clarity: netrin1 protein is deposited on the pial surface of the spinal cord where it acts as a haptotactic growth substrate
As we started to write the paper, Sup had a thesis committee meeting with Kelsey Martin, Alvaro Sagasti and Larry Zipursky where she was questioned skeptically about the feasibility of our hederal model. Larry, in particular, wanted to know more about the distribution of netrin1 protein. Larry, working with his postdoc Orkun Akin, had just shown that netrin had an adhesive, rather than chemotropic, role in the fly medulla (15). We grudgingly admitted that we had never looked, because we had no expectation that netrin1 protein would be anywhere other than the VZ. A couple of days later, Sup appeared at my door, looking worried. “The staining doesn’t look right,” she said. “There’s a lot of background staining on the pial surface of the spinal cord, and in axons.” I internally cursed the non-specificity of our antibody. “Try antigen retrieval.” I said, “That’s what Tim Kennedy had to do!” (16). Sup came back a day or two later, looking even more worried, and told me that the antigen retrieval protocol made the staining “worse.” Both the pial and axonal staining looked brighter than ever. “It looks real!” she said glumly. This distribution made no sense in our model, and I complained to Larry about his opening up a can of worms, when I saw him at a seminar that afternoon. “It’s a bad molecule,” Larry joked.
Sup was right; the antibody staining did look real. There were very low levels of netrin1 in the VZ as we had predicted, but there were much higher levels on most of the pial surface around the circumference of the spinal cord, and on axons (Fig. 1B). I showed James Briscoe the distribution pattern when he visited UCLA a couple of weeks later. “Of course it’s real,” he said, “The neural progenitor cells are making netrin1, and then depositing it on the pial surface using their radial endfeet.” We corralled Ben in from his office next door to mine to assess what he thought. He thought the staining was real too. The three of us peered excitedly at my computer screen, the realization of how the pattern of the netrin1 transcript related to the distribution of netrin1 protein slowly sinking in. We then discussed nothing else for the rest of the weekend.
Sup went on to show that indeed James’ supposition was correct. Through a trick of their cellular geometry, the neural precursors that make netrin1 then use their radial processes to deposit it as a growth substrate onto the pial surface (Fig. 1B, C). No netrin1 is made by the dorsal-most neural precursors, and indeed there is no netrin1 on the most dorsal pial surface. Sup pointed out that the resulting sharp on:off netrin1 boundary on the dorsal pial surface really suggested that netrin1, a member of a laminin family, could not be highly diffusible. In other exhilarating moments of understanding, we realized that spinal neurons specifically initiate NF+ axonal growth on this netrin1 pial-substrate. And that netrin1 only accumulates on commissural axons as they grow adjacent to the netrin1 pial-substrate, as if netrin1 can transfer from this substrate to axons. In a further remarkable result, Sup found that the putative axonal-transfer of netrin1 requires Dcc.
We finally understood how to model our results: VZ-derived netrin1 acts locally as a substrate to promote fasciculated axonal growth in an oriented manner towards the ventral midline. Netrin1 and Dcc appear to cooperate within axons as part of the mechanism that orients growth (Fig. 1C). The innervation of the VZ that we had observed in both netrin1 and Dcc mutants might in fact be randomized, or defasciculated, growth as a result of losing this adhesive interaction (Fig. 1D). VZ-derived netrin1 thus appears to act by haptotaxis, i.e. by the establishment of a local adhesive surface, the alternative model to chemotaxis. As a mechanistic side note: it remains unclear how this adhesive surface acts to orient and fasciculate commissural axon growth and whether there is an additional “no go” activity in the VZ. Is this mechanism functioning solely through haptotaxis? Or is there also a “hederal” component? Nonetheless, these activities – the pull of an adhesive substrate that promotes fasciculation, perhaps coupled with the push of a “no go” signal – permit axons to grow along the netrin1(-):netrin1(+) border, i.e. in a circumferential path precisely around the VZ.
Third moment of clarity: FP-derived netrin1 is dispensable for axon guidance
Our model – the ability of neural progenitor cells to deposit a haptotactic substrate of netrin1, which promotes ventrally-directed, fasciculated axon growth – was a notable departure from the textbook view of netrin1. But was this model the only mode of action? Or did netrin1 have long-range or short-range activities depending on the cell type? In his original papers, Marc Tessier-Lavigne argued the case for and against netrin1 acting by chemotaxis or haptotaxis (1), but ultimately settled on the idea, well supported by the data at the time, that netrin1 was presented as a long-range cue from the FP. I discussed this point vigorously and endlessly with Sup, Ben, Artur, Larry and Orkun. We had not doubted this model until now. Could our phenotypes still be explained by a long-range activity from the FP? Were the activities of short-range VZ-derived netrin1 in addition to the long-range activities? Or could FP-derived netrin1 really be dispensable for axon guidance?
In the end, the reviewers decided it. The beauty of the finding that neural progenitor cells lay down a substrate of netrin1 that orients and promotes the axonal trajectories of their own neural progeny was not considered sufficiently novel. It was clear what experiment needed to be done, thus we requested the conditional netrin1 lines from Holger Eltzschig and set about breeding them to Cre recombinase driver lines that would remove netrin1 specifically from either FP cells (ΔFP, Fig. 1E) or the dorsal VZ (ΔVZ, Fig.1F). Sup accomplished her breeding scheme astonishing quickly and the day came when she finally had the answer sitting on a microscope slide. I ran to the confocal room, both anxious and excited, to discover the result. “There’s no effect,” she said. “It doesn’t matter if you remove netrin1 from the FP!” In contrast, removing netrin1 from the VZ had profound effects. Axons grew in all directions, but only locally, specifically in the region where netrin1 had been removed from the VZ. Together with the Gli2-mediated FP ablation data, our studies had found no evidence for long-range chemotaxis in the spinal cord. Who knew? Perhaps Cajal’s model was wrong!
The paper was finally accepted at Neuron (11), and came out at the same time as a paper from Alain Chedotal in Nature (17), with complementary findings in the hindbrain. A few days later, I ran into a colleague on the street onside my lab. “I saw your paper in Neuron!” she laughingly scolded me, “But please don’t tell me I have to change my lecture on netrin1! I liked that lecture!” I liked my lecture on netrin1 too, but now I also have to change it. Netrin1, of course, remains the supreme architect of spinal circuitry, but now acts locally as a directional surface along which axons can extend, akin to the holds used by climbers to pioneer their way to the top of a mountain.
Samantha Butler
Department of Neurobiology,
Eli and Edythe Broad Center of Regenerative Medicine and Stem Cell Research,
Intellectual and Developmental Disabilities Research Center
University of California, Los Angeles.
 (No Ratings Yet)
(No Ratings Yet)
 (2 votes)
(2 votes)


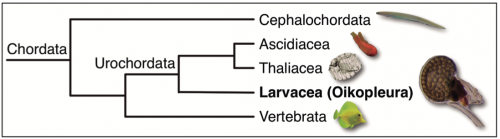
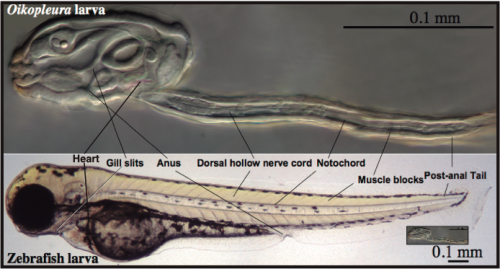

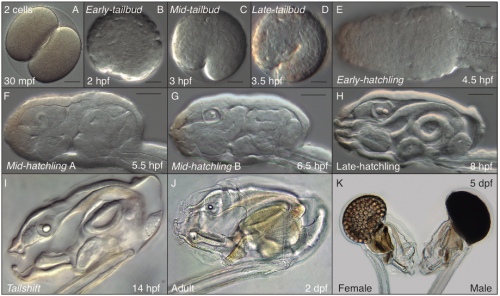






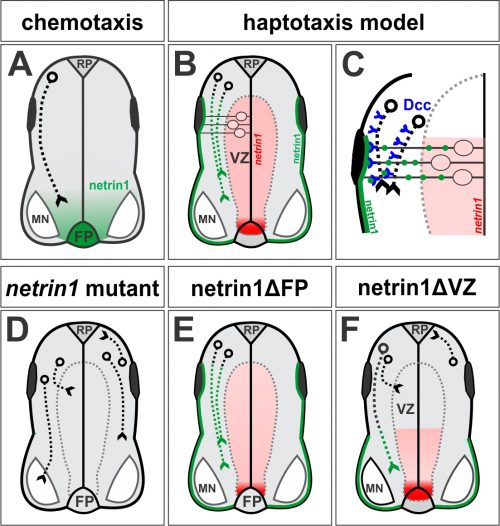
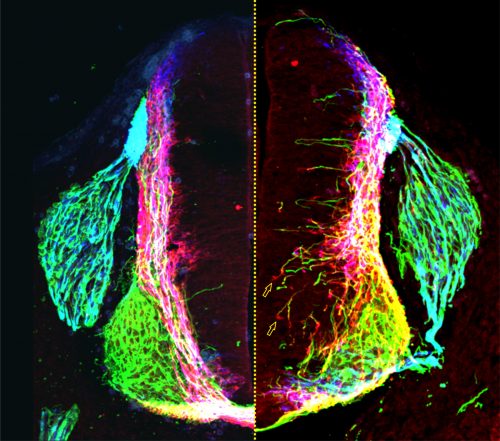
 (16 votes)
(16 votes)