We are looking for an enthusiastic and highly motivated Staff Scientist or experienced Research Assistant/Technician to join the stem cell and regenerative medicine-based laboratory of Dr. April Craft, within the department of Orthopaedic Research at Boston Children’s Hospital and Harvard Medical School. Our lab studies the development of musculoskeletal tissues, primarily articular cartilage, using human pluripotent stem cells as a model system. Projects in the lab range from basic to translational research.
The candidate will be essentially involved in all aspects of the laboratory but will primarily focus on 1-2 prioritized scientific projects, including modeling of cartilage disorders using patient specific iPSCs. The successful candidate will work closely with the PI and post-doctoral fellows in the lab, and be responsible for maintaining human pluripotent stem cell differentiation cultures and analyzing/recording results using various established assays. Individualized research projects will be available based on proven experience and independence. Additional tasks include preparation of reagents and media, and a supporting role in lab supplies inventory/ordering and maintenance of lab equipment. Occasional weekend work is required. This job has physical requirements that include the ability to bend, lift and carry objects weighing up to 50 lbs on occasion, frequently reach and grasp objects above and below shoulder level, and occasional proofreading and checking documents for accuracy.
The applicant must be a US citizen or permanent resident. Applicants must have at least a BS/BA in Biomedical science (or equivalent); Candidates that have a MS or PhD degree are preferred. Applicants must have at least three years of cell culture experience, preferentially human embryonic stem cell culture, and a strong understanding of molecular biology. We will teach the right candidate the necessary skills. Salary will commensurate with experience. Excellent organization skills and communications skills in English are required. A minimum three-year commitment is requested, however this position is potentially long term.
Applicants should email a CV, contact information for three referees and a cover letter describing their background to april.craft@childrens.harvard.edu.
We are looking for an outstanding, highly motivated postdoctoral fellow to join our innovative young department in the stem cell and regenerative medicine-focused laboratory of Dr. April Craft, Assistant Professor of Orthopaedic Surgery at Boston Children’s Hospital and Harvard Medical School. Our lab studies the development of musculoskeletal tissues, primarily cartilage, using human and mouse embryonic (ESCs) and induced pluripotent stem cells (iPSCs) as a model system. Available projects in the lab range from basic developmental biology to translational research. We are interested in applying our knowledge of how to generate cartilage tissues from pluripotent stem cells to further understand how genetic mutations cause skeletal dysplasias in patients, particularly those for which animal models do not exist. We have also recently identified a panel of molecular markers that may help to define progenitor cells that are best suited for cartilage regeneration and repair.
A strong background in one of the following fields is required: pluripotent stem cell biology, basic developmental biology, or musculoskeletal/cartilage development, homeostasis or disease. Candidates with hands on experience using pluripotent stem cells will be given preferential consideration. The successful candidate will have a recently acquired PhD or MD-PhD, a minimum of 3 years laboratory experience including tissue culture, and at least one first author publication. The candidate must be a US citizen or permanent resident, and must possess excellent communication & writing skills in English. Members of the Craft lab participate in seminar series and other events within the Harvard Stem Cell and Regenerative Biology Program, Boston Children’s Hospital Orthopaedic Research Laboratories, Harvard Stem Cell Institute, and Harvard Medical School.
Applicants should email a CV, contact information for three referees and a cover letter describing their background and research interests to april.craft@childrens.harvard.edu.
A Ph.D. position in molecular cell and developmental biology of plants is available in the lab of Prof. Kay Schneitz, Dept. of Plant Developmental Biology, Technical University of Munich in Freising/Germany. The successful candidate will work on the molecular characterization of the signaling pathway mediated by the atypical transmembrane receptor-like kinase STRUBBELIG (SUB) in Arabidopsis (1-4). The SUB pathway functionally links intercellular signaling controlled by receptor kinases and plasmodesmata, gateable channels interconnecting most plant cells, during tissue morphogenesis of for example flowers and ovules. Starting date is negotiable but ideally the position should be filled as soon as possible. The lab is part of the Collaborative Research Centre SFB924 (sfb924.wzw.tum.de) and thus funding comes from the German Research Council (DFG) and is at the usual EG13/2 level. Requirements are e.g. a German masters (with a mark of 2.5 or better), a French DEA (a final average of 13 or more), or a masters thesis. The person needs to have a firm basis in basic molecular techniques. Fluency in English is a must. Freising is located about 35 km to the north of Munich. Munich is a lively, cosmopolitan city close to beautiful lakes and the Alps. For further information please contact Kay Schneitz (kay.schneitz@tum.de) and visit the webpage (plantdev.wzw.tum.de).
(1) Chevalier et.al. 2005 PNAS 103: 9074-9079.
(2) Fulton et.al. 2009 PLoS Genetics 5: e1000355
(3) Vaddepalli et.al. 2011 PLoS One 6: e19730
(4) Vaddepalli et.al. 2014 Development 141: 4139-4148
In an interview, Lewis Cantley, Director of the Sandra and Edward Meyer Cancer Center at Weill Cornell Medicine, talked about discovering the PI3K pathway and the relationship between metabolism and cancer
Chunhui Xu and colleagues presented a human iPSC model for catecholaminergic polymorphic ventricular tachycardia.
Katherine Swenson-Fields and colleagues described a pathway for the differentiation of macrophages in polycistic kidney disease.
We are seeking a creative and exceptionally motivated candidate to fill a post-doctoral position in the field of evolutionary and developmental genetics.
Research in the Kamberov lab is directed at uncovering the genetic basis of human adaptive traits, with a core focus on the evolution of skin appendages, namely sweat glands and hair follicles. In this pursuit, we utilize a highly interdisciplinary approach that combines mouse and human genetics with developmental biology and evolutionary genomics. The culmination of this research program is to not only enhance the understanding of human evolution but also to apply what we have learned in translational efforts that lead to the improvement of human health and alleviation of disease, particularly in the context of skin and skin appendage regeneration.
Projects include:
-Dissection of molecular pathways underlying skin appendage development and evolution.
-High throughput screening for regulatory elements controlling the specification and patterning of hair follicles and sweat glands.
-Dissection of stem cell populations in the skin and the regeneration of human skin appendages in vitro.
-Discovery and functional modeling of evolutionarily significant human variants using comparative genomics and humanized mouse models.
The position provides an exciting opportunity to work at the interface of basic and translational research in a collaborative and stimulating environment, and gain experience in a diverse set of technical approaches at the cutting edge of evolutionary, developmental and regenerative biology.
A doctorate in biology or related field is required. Applicants with a strong background in developmental biology, genetics, genomics or molecular biology are encouraged to apply. Prior experience with mouse genetics and husbandry is preferred.
Interested candidates should provide: 1) a CV 2) a brief letter detailing your interest in the lab and relevant past research experience 3) contact information for three references who can comment on your research. Application materials and any questions regarding the position should be addressed to Yana Kamberov: yana2@mail.med.upenn.edu
We are currently seeking an independent and motivated scientist to join the research group led by Dr Anestis Tsakiridis (https://www.sheffield.ac.uk/cscb/tsakiridis). His work focuses on dissecting the molecular basis of cell fate decisions in the developing embryo and aims to exploit this knowledge for disease-modelling and regenerative medicine applications. The group is particularly interested in the biology of neuromesodermal progenitors (NMPs), the bipotent cell population which drives embryonic axis elongation by generating spinal cord neuroectoderm and paraxial mesoderm.
We have recently described the efficient derivation of NMPs from human pluripotent stem cells (hPSCs) thus establishing a tractable system for studying these progenitors in vitro. The ideal candidate will employ this model in combination with various screens and complementary approaches aiming to define the intrinsic and extrinsic determinants of NMP maintenance and differentiation. Applicants should hold a PhD in a related field and have experience in hPSC culture/differentiation, immunocytochemistry, high content imaging and chromatin immunoprecipitation. A background in Crispr/Cas9-based genome editing and/or bioinformatics would be highly advantageous.
September was notable for the amount of new jobs posted, from Kansas to Copenhagen to King’s College London, and PhD to tenure-track. Check out our jobs page for the latest opportunities. In case you missed them, here are some of our other monthly highlights:
We continued our ‘People behind the papers’ series with Adam Johnston, who talked about his recent work on mouse digit tip regeneration and his new lab in Canada. We also featured an interview with Lewis Cantley from Disease Models and Mechanisms.
Life beyond the bench
A visual report from the Royal Society early career conference by Scriberian
The European Molecular Biology Laboratory is searching for Group Leaders. EMBL offers a highly collaborative, uniquely international culture. It fosters top quality, interdisciplinary research by promoting a vibrant environment consisting of young, independent researchers with access to outstanding graduate students and postdoctoral fellows.
Group Leader Developmental Biology
at EMBL Heidelberg, Germany
The Developmental Biology Unit studies the development of multicellular
organisms. Research in the unit covers all levels, from the cellular to
the whole organism, and is highly interdisciplinary, combining a wide
range of approaches and innovative techniques, with special emphasis
on quantitative and real-time imaging. Research in the unit is firmly
embedded within the overall EMBL research environment, with extensive
in-house collaborations and support from world-class services,
including the gene core, transgenic, metabolomics and mass-spectrometry
core facilities.
We are seeking outstanding candidates using model organisms to
address fundamental principles of multicellular development across the
entire spectrum of developmental biology. Complementary approaches,
including simplified systems, such as organoid and stem cell systems,
are also welcome. Candidates with strong background and research
using theoretical approaches are also encouraged to apply. There is the
possibility to hire two group leaders in the present call.
The successful candidate should have a strong motivation to work in
the multidisciplinary and collaborative environment of EMBL, grasping
the opportunity to interact with many other research groups. In general,
EMBL appoints group leaders early in their career and provides them
with a very supportive environment for their first independent position to
achieve highly ambitious and original research goals.
EMBL is an inclusive, equal opportunity employer offering attractive
conditions and benefits appropriate to an international research
organisation with a very collegial and family friendly working
environment. The remuneration package comprise a competitive
salary, a comprehensive pension scheme, medical, educational and
other social benefits, as well as financial support for relocation and
installation, including your family, and the availability of an excellent
child care facility on campus. EMBL is committed to achieving
gender balance and strongly encourages applications from women.
Appointment will be based on merit alone.
APPLICATION INSTRUCTIONS
Please apply online through www.embl.org/jobs and include a cover letter, CV
and a concise description of research interests and future research plans.
Please also arrange for 3 letters of recommendation to be emailed directly by
your referees to references@embl.de at the latest by 20 October 2016.
Interviews are planned for 19 to 21 December 2016.
For further information about the position can be obtained from
the Head of Unit Anne Ephrussi (anne.ephrussi@embl.de).
Further details on Group Leader appointments can
be found under www.embl.org/gl_faq.
Our monthly trawl for preprints from developmental biology and related research. Previous posts can be found here.
Preprints roll on, and this month’s selection is a reminder of how developmental biology is based on a variety of model organisms. We’ve got two investigations into zebrafish mesoderm from Deborah Yelon’s lab, quite a bit of cell mechanics (in zebrafish, fly and worm), updates on the underlying cell biology of cell division in c. elegans, a transcriptome for the spiny mouse, and much more besides.
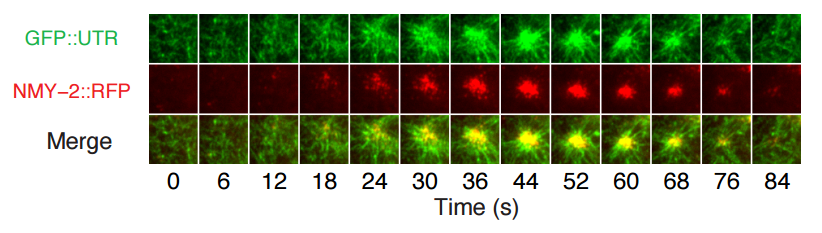
Figure 1C from Robin, Michaux, et al. (Edwin Munro lab), showing dynamics of utrophin and myosin at the one cell stage of the c. elegans embryo
As in the last months, the preprints predominantly came from bioRxiv, but we also found a couple from PeerJ. If we missed anything out, let us know. Happy preprinting!
Landscape of X chromosome inactivation across human tissues. Taru Tukiainen, Alexandra-Chloé Villani, Angela Yen, Manuel A. Rivas, Jamie L. Marshall, Rahul Satija, Matt Aguirre, Laura Gauthier, Mark Fleharty, Andrew Kirby, Beryl B. Cummings, Stephane E. Castel, Konrad J. Karczewski, François Aguet, Andrea Byrnes, GTEx Consortium, Tuuli Lappalainen, Aviv Regev, Kristin G. Ardlie, Nir Hacohen, Daniel G. MacArthur
Nuclear pore-like structures in a compartmentalized bacterium. Evgeny Sagulenko, Amanda Nouwens, Richard I Webb, Kathryn Green, Benjamin Yee, Gary Morgan, Andrew Leis, Kuo-Chang Lee, Margaret K Butler, Nicholas Chia, Uyen Thi Phuong Pham, Stinus Lindgreen, Ryan Catchpole, Anthony M Poole, John A. Fuerst
Contrasting genome dynamics between domesticated and wild yeasts. Jia-Xing Yue, Jing Li, Louise Aigrain, Johan Hallin, Karl Persson, Karen Oliver, Anders Bergström, Paul Coupland, Jonas Warringer, View ORCID ProfileMarco Cosentino Lagomarsino, Gilles Fischer, Richard Durbin, Gianni Liti
Nanopore DNA Sequencing and Genome Assembly on the International Space Station. Sarah L Castro-Wallace, Charles Y Chiu, Kristen K John, Sarah E Stahl, Kathleen H Rubins, Alexa B. R. McIntyre, Jason P Dworkin, Mark L Lupisella, David J Smith, Douglas J Botkin, Timothy A Stephenson, Sissel Juul, Daniel J Turner, Fernando Izquierdo, Scot Federman, Doug Stryke, Sneha Somasekar, Noah Alexander, Guixia Yu, Christopher Mason, Aaron S Burton
This post first appeared on Annotations, the DMDD blog (blog.dmdd.org.uk).
Around a third of targeted gene knockouts in mice are embryonic-lethal. But not all deaths occur during gestation – a significant number of gene knockouts result in death at or shortly after the time of birth. Mice from these knockout lines provide a valuable animal model of human neonatal death and are the focus of a new systematic screen by the DMDD programme.
The study, to be carried out by consortium members at the Wellcome Trust Sanger Institute, will offer new insights into the genetic basis of death in neonates, complementing the efforts of large-scale human gene studies such as the DDD and UK10K.
UNDERSTANDING LETHALITY IN NEONATES
The DMDD programme studies embryonic-lethal knockout mouse lines, where lethal means that no pups are observed 14 days after birth (known as postnatal day P14). Detailed phenotyping of these lines can provide important clues about the genetic basis of human developmental disorders.
Our phenotyping efforts so far have focussed on in-utero development at embryonic days E9.5, E14.5 and E18.5 since, for the majority of embryonic-lethal lines, the embryos die well before birth.
In 5-10% of cases, however, the embryos die at the time of birth or shortly afterwards, and can provide an animal model of neonatal death due to genetic mutation. The pilot neonatal screen will study 20 of these lines during the period E18.5 to P14, gathering systematic phenotype data.
Nesting wild-type pups on their day of birth.
Common causes of lethality for neonates can include problems with partuition, breathing, suckling and achieving homeostasis. To understand the likely cause of death in each case, the neonatal screen will assess the pups’ gross dysmorphology, breathing, milk spot morphology (a test that reveals whether they have been able to feed), weight, measurements, locomotive skills, righting reflex (ability to correct their own body position) and blood glucose levels.
The first data is expected at the end of 2016. For more information about the DMDD programme, visit dmdd.org.uk.
 (No Ratings Yet)
(No Ratings Yet)




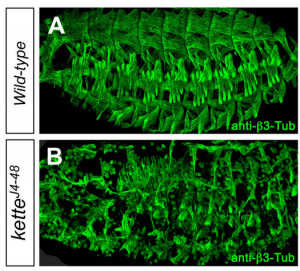

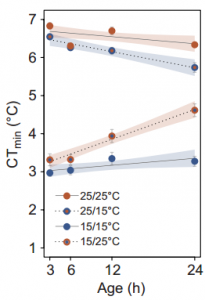
 (3 votes)
(3 votes)