Calling all developmental biology postdocs and graduate students!
Interested in attending an exciting meeting? Want to expand your dev bio network? Want to present your work to a supportive audience?
Come to the 2023 Developmental Biology Gordon Research Conference and Seminar
Date: June 25 – 30
Venue: Mount Holyoke College, Mass, USA
Abstract deadline May 21, 2023
The organisers have raised significant funds to support registration fees and travel grants for postdocs and graduate students. If you need financial assistance please contact the organisers.
“You can boil them, freeze them, throw them into outer space – and they will survive. So how can we use these incredible adaptations to solve some of our own problems?”
Dr Sally Le Plage
In the latest episode of the Genetics Unzipped podcast, we’re going microscopic, exploring what tiny tardigrades can teach us about DNA damage, vaccine distribution and even astronaut health in space.
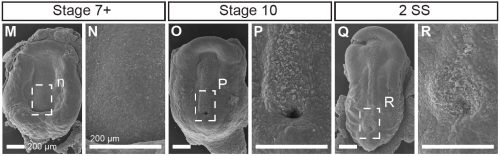
This project began like a lot of very cool studies do, completely accidentally. We were wrapping up a project describing neural crest cell patterning in veiled chameleons started by a former PhD student Dr. Raul Diaz, and Dr. Natasha Shylo created a staging series of veiled chameleon embryo development to go with the study (Diaz et al., 2019). Based on her prior work in left-right patterning, Dr. Shylo then recognized the potential of veiled chameleons to understand how left-right patterning occurs in non-avian reptiles. When we started this work in 2019, nothing was known about left-right patterning in non-avian reptiles. And it was going to be easy (famous last words), since we already had some of the key reagents – Shh and Fgf8 RNA in situ probes from our previous work in chameleon limb patterning (Diaz and Trainor, 2015), and a Nodal probe for a gastrulation study (Stower et al., 2015). Shh was the first molecular marker ever published with asymmetric molecular expression in chicken, so it was a logical starting point. Also given the presence of cilia and their known roles in left-right organizer function, we also searched for cilia using SEM. But Shh was not asymmetrically expressed in veiled chameleon embryos, and we found nothing that resembled motile cilia in the blastopore region of the embryo.
Figure 1. SEM of chameleon embryos across the stages of left-right patterning. Modified from figure 4 in Shylo et al., 2023. No evidence for cilia, exhibiting motile-like identity was found at any of the stages.
What was known about left-right patterning in non-avian reptiles before your work?
Left-right patterning in non-avian reptiles was a black box when we started this project. However, we were not the only ones curious to tackle this question. As we were trying to figure out the steps of the Nodal signaling cascade and what happens in an organism that doesn’t use cilia in its left-right organizer, Dr. Hiroshi Hamada’s group published a paper revealing that turtles and geckos also do not use motile cilia to establish left-right patterning (Kajikawa et al., 2020). We were able to build on those observations and realized we were looking at the “wrong” Nodal gene. Nodal1, as we now refer to it, was not supposed to exist in chameleons. It is the only Nodal paralog present in mammalian genomes, but has been lost from the genomes of chickens, geckos, and turtles, which retained Nodal2 instead. It turned out that the veiled chameleon genome contains two Nodal genes – both retained from a duplication event in jawed vertebrates.
What made you choose the veiled chameleon as your model organism?
Although we have a good understanding of early developmental events in many vertebrate model organisms, few studies if any involved the reptile clade. This is because at the time lizards and snakes lay their eggs, their embryos are already at limb bud stages of development, which is far too late to study left-right patterning, gastrulation and neurulation. We became interested in veiled chameleons because their embryos are pre-gastrula stage at the time of oviposition and because of their sexually dimorphic casque, which we hypothesized was neural crest cell derived. Fortunately, veiled chameleons are a popular pet with established husbandry. They breed year-round and lay large clutches of eggs. Dr. Shylo came to the lab with expertise in left-right patterning, and veiled chameleons proved to be a perfect research organism to study the mechanisms establishing left-right asymmetry in non-avian reptiles.
How is it like working with veiled chameleons?
One must be patient when working with veiled chameleons because their development is really slow. It can take up to 70 days to accomplish gastrulation, and a week to establish left-right patterning, compared to about 6 hours in mice, which means veiled chameleons allow for a much finer temporal resolution of key developmental processes. We stagger chameleon mating, and consequently always have an abundance of embryos at various stages of development throughout the year. Veiled chameleon eggs are soft shelled, which means we can’t manipulate the embryos in ovo like chicken embryos, but they are amendable to ex ovo culture, lineage tracing, time lapse imaging and most molecular techniques.
Can you summarise your key findings?
We used veiled chameleons to study the evolution of developmental mechanisms and obtain a deeper understanding of how left-right patterning is established in non-avian reptiles. We found that like chickens, veiled chameleons do not use motile cilia in their left-right organizer to establish left-right asymmetry. Instead, through live imaging, we observed asymmetric morphological changes in the embryo, which precede, and likely initiate molecular asymmetry. We further discovered that veiled chameleons have retained two Nodal genes – Nodal1 and Nodal2 – from a duplication event in jawed vertebrates. In comparison, mammals have only Nodal1, whereas avians have retained Nodal2. This work has laid the foundation for our future studies, aimed at understanding in greater detail the mechanisms and processes that establish left-right asymmetry in diverse groups of amniotes.
Were you surprised to find that the veiled chameleons do not have motile cilia in their left-right organizer?
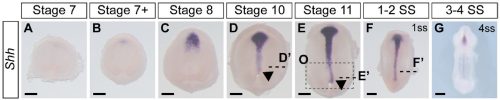
Not entirely. Going into this project, the outcome was always going to be binary – they will have motile cilia or not. It has been known for a long time that avian embryos do not have motile cilia in their left-right organizer, so it was reasonable to expect that all reptiles might be the same. What was surprising was the pattern of Shh expression, because Shh is expressed asymmetrically around the node in chickens and pigs (pigs also lack motile cilia in their left-right organizer). In veiled chameleon embryos, Shh is expressed symmetrically in the floor plate, but major morphological changes in the embryo push the Shh expressing tissues to the left. In another surprise and again distinct from chickens and pigs, veiled chameleons don’t use a primitive streak for gastrulation, and appear to lack a morphological node structure. We presume that the blastopore slit in chameleons carries out the function of the left-right organizer, but this remains to be determined.
Did you have any particular result or eureka moment that has stuck with you?
We knew that imaging development live would be key to our understanding of left-right patterning in chameleons, and we collected multiple movies at various stages of development, but no matter how we squinted, tilted our heads, or what type of analysis we used, we could not find consistent robust asymmetric cellular movements in the embryos we imaged. However, in a fortuitous meeting with collaborator and co-author Dr. Sarah E. Smith, viewing the movies in the transverse optical plane revealed an unmistakable asymmetric tilt that occurred reproducibly in all of our movies, which pushes the Shh expression tissue to the left at the same precise stage of development! We still think that the gross morphological rearrangement that we reported is driven by finer cellular movements, and we will continue our work with chameleons to figure out where, when, and how it all starts.
And the flipside: were there any moments of frustration or despair?
As mentioned earlier, Shh was the first molecular marker that we characterized, and it appeared perfectly symmetrical at every stage that we initially examined. This observation was consistent with a report in turtles and geckos from Dr. Hiroshi Hamada’s lab. Apparently, we had a model system in which the left-right organizer lacked motile cilia, and Shh was symmetrically expressed, and this contradicted everything we knew from mice (motile cilia) and chickens (asymmetric Shh). However, it made some sense, since chameleons don’t have a conventional node that rotates. As we started to prepare the manuscript, Dr. Shylo realized a broader time course of Shh expression was required, and there it was, unmistakable asymmetrically displaced Shh expressing tissue! Only these developmental stages were much earlier than when asymmetric Nodal expression is observed! Thus, an initial moment of frustration turned into weeks of excitement. We had simply been looking too late in development. These data revealed that morphological changes occur in a veiled chameleon embryo well before we can detect asymmetric Nodal expression, opening up new models and mechanisms to explore.
Figure 2. Expression of Shh in veiled chameleons across the stages of left-right patterning. Modified from figure 6 in Shylo et al., 2023. Shh highlights an asymmetry in the midline near the blastopore (Arrowheads).
Natasha – What brought to you join Paul’s lab? And what is next for you after this paper?
My Ph.D. focused on the ciliary gene Tmem107 in a mouse mutant which exhibited left-right patterning defects (Shylo et al., 2020). Work with a collaborator revealed that these mice also had a neural crest defect, which made me curious about roles for cilia in neural crest cell specification and migration (Cela et al., 2018). So I came to Paul’s lab with a plan to study neural crest cells in mice, but I will continue to study the evolution of developmental mechanisms, particularly with respect to left-right patterning and gastrulation.
I plan to seek out a faculty position in the next year or two, and establish a laboratory to study gastrulation, left-right patterning, and other early developmental processes, using veiled chameleons as a model for early amniotic development. Although my focus has changed dramatically through a series of serendipitous events, there is no other place I could have done this work as efficiently as in Paul’s lab at Stowers Institute. To my knowledge, we maintain the largest colony of veiled chameleons used specifically for research. We have recently sequenced the veiled chameleon genome and once the annotation is finalized, we will have all the genetic, molecular and cellular tools we need to functionally study the evolution of developmental mechanisms with an emphasis on early development in these reptiles. My message to other postdocs based on my experience is to be brave, and if an amazing project summons your attention, it is OK to completely switch directions and your scientific focus. Go for it.
Paul – Where will this story take the lab?
Veiled chameleons exhibit a number of interesting morphological features including a cranial casque, forelimb and hindlimb clefting, and wrists and ankles that function like balls and sockets instead of hinges. Chameleons also have a projectile tongue, a prehensile tail, and they can color change. All of these features make chameleons very well suited for arboreal environments. With annotation of our newly sequenced genome nearly complete, we will soon be able to tackle functional genetic questions in ecological-evolutionary-developmental biology using veiled chameleons in concert with traditional model organisms.
References
CELA, P., HAMPL, M., SHYLO, N. A., CHRISTOPHER, K. J., KAVKOVA, M., LANDOVA, M., ZIKMUND, T., WEATHERBEE, S. D., KAISER, J. & BUCHTOVA, M. 2018. Ciliopathy Protein Tmem107 Plays Multiple Roles in Craniofacial Development. J Dent Res, 97, 108-117.
DIAZ, R. E., JR., SHYLO, N. A., ROELLIG, D., BRONNER, M. & TRAINOR, P. A. 2019. Filling in the phylogenetic gaps: Induction, migration, and differentiation of neural crest cells in a squamate reptile, the veiled chameleon (Chamaeleo calyptratus). Dev Dyn, 248, 709-727.
DIAZ, R. E., JR. & TRAINOR, P. A. 2015. Hand/foot splitting and the ‘re-evolution’ of mesopodial skeletal elements during the evolution and radiation of chameleons. BMC Evol Biol, 15, 184.
KAJIKAWA, E., HORO, U., IDE, T., MIZUNO, K., MINEGISHI, K., HARA, Y., IKAWA, Y., NISHIMURA, H., UCHIKAWA, M., KIYONARI, H., KURAKU, S. & HAMADA, H. 2020. Nodal paralogues underlie distinct mechanisms for visceral left-right asymmetry in reptiles and mammals. Nat Ecol Evol, 4, 261-269.
SHYLO, N. A., EMMANOUIL, E., RAMRATTAN, D. & WEATHERBEE, S. D. 2020. Loss of ciliary transition zone protein TMEM107 leads to heterotaxy in mice. Dev Biol, 460, 187-199.
SHYLO, N. A., SMITH, S. E., PRICE, A. J., GUO, F., MCCLAIN, M. & TRAINOR, P. A. 2023. Morphological changes and two Nodal paralogs drive left-right asymmetry in the squamate veiled chameleon (C. calyptratus). Frontiers in Cell and Developmental Biology, 11.
STOWER, M. J., DIAZ, R. E., FERNANDEZ, L. C., CROTHER, M. W., CROTHER, B., MARCO, A., TRAINOR, P. A., SRINIVAS, S. & BERTOCCHINI, F. 2015. Bi-modal strategy of gastrulation in reptiles. Dev Dyn, 244, 1144-1157.
The fourth episode of the Wellcome-funded Human Developmental Biology Initiative‘s new podcast, Made the Same Way, features scientist Emily Calderbank and rapper Olivia (aka FREEQUENCY3) discussing human embryonic haematopoiesis.
At the end of the episode, the pair collaborate on an original piece of music which reflects their conversation.
Right now, every second, your blood is making 3 million cells!
Emily Calderbank
About the participants
Emily Calderbank is a postdoctoral researcher at the Stem Cell Institute, part of the University of Cambridge. Her research focuses on the production and maintenance of human blood throughout life, from embryo to adult, and the role that inflammatory signals may play in this process.
In her spare time, she enjoys listening to podcasts and baking.
Olivia, AKA FREEQUENCY3 is an Alternative HipHop rapper, DJ and host. FREEQUENCY3 explores emotion, values & the ripples of life through music, both written and freestyle. Over the past year FREEQUENCY3 has supported Dizraeli on his UK Tour and rapped in the ‘CalibySnoop’ Cypher, which brought together some of Manchester’s best rappers to launch Snoop Dogg’s CalibySnoop wine in the UK.
In her spare time, she enjoys being around friends and family, listening to music, watching tv and relaxing.
Please subscribe and listen to Made the Same Way on Apple podcasts, Spotify, or wherever you get your podcasts. If you enjoy the podcast, please rate and review us on Apple podcasts to help others find us!
One of the crowd-favourite giveaways here at the Node is our collection of postcards. With our supplies dwindling, we are planning to reprint some of the postcards, and take this opportunity to add some more #devbio favourites to our collection.
Thank you to all the researchers who have submitted their images to the Node postcard competition back in April. We have now narrowed down to the final 8, and would like to see which ones the community like to feature on our postcards. The top 4 will be printed on our postcards, and the winner will be also be featured on the cover of a ‘Development’ issue in 2023.
Thank you and good luck to the following researchers for their contributions:
Daniel Rios, Elio Escamilla, Elisabeth Kugler, Jessica Marin, Margot Smit, Nick Gatford, Özge Özgüç, Valerie Tornini
And a big thank you to everyone who submitted their images to the competition. There were many good quality submissions that it was very difficult to narrow down the selection!
Please vote for your favourite image at the bottom of the page. The voting will close on Sunday 4 June 11:59pm.
“Geographical Maps“
Depicted is a digital section of the developing eye in a zebrafish. Specifically, these are support cells of the eye called glia cells, which have undergone 3D data processing to extract the surface of these cells. Different colours represent different depths in the studied eye. Technique: The image was acquired with the Zeiss LSM 900 AiryScan microscope, using a 40x water-immersion LD C-Apochromat (NA 1.1) objective. Processing was conducted using Fiji. Following image rotation in 3D, data were segmented and the surface extracted. Using depth-coding and application of different colour-palettes achieved the different colorations seen.
Drosophila larvae
This shows heat-fixed Drosophila larvae expressing an infrared fluorescent protein (IFP) in the tracheal system using the Gal4/UAS system. Images were acquired in a confocal microscope (Nikon A1R+) with a 10x objective and using the mosaic modality. Stitching was done using the microscope’s software (Nikon NIS-Elements). The images were Z-projected and pseudocolored in Fiji and further processed using Inkscape.
Hatching embryo
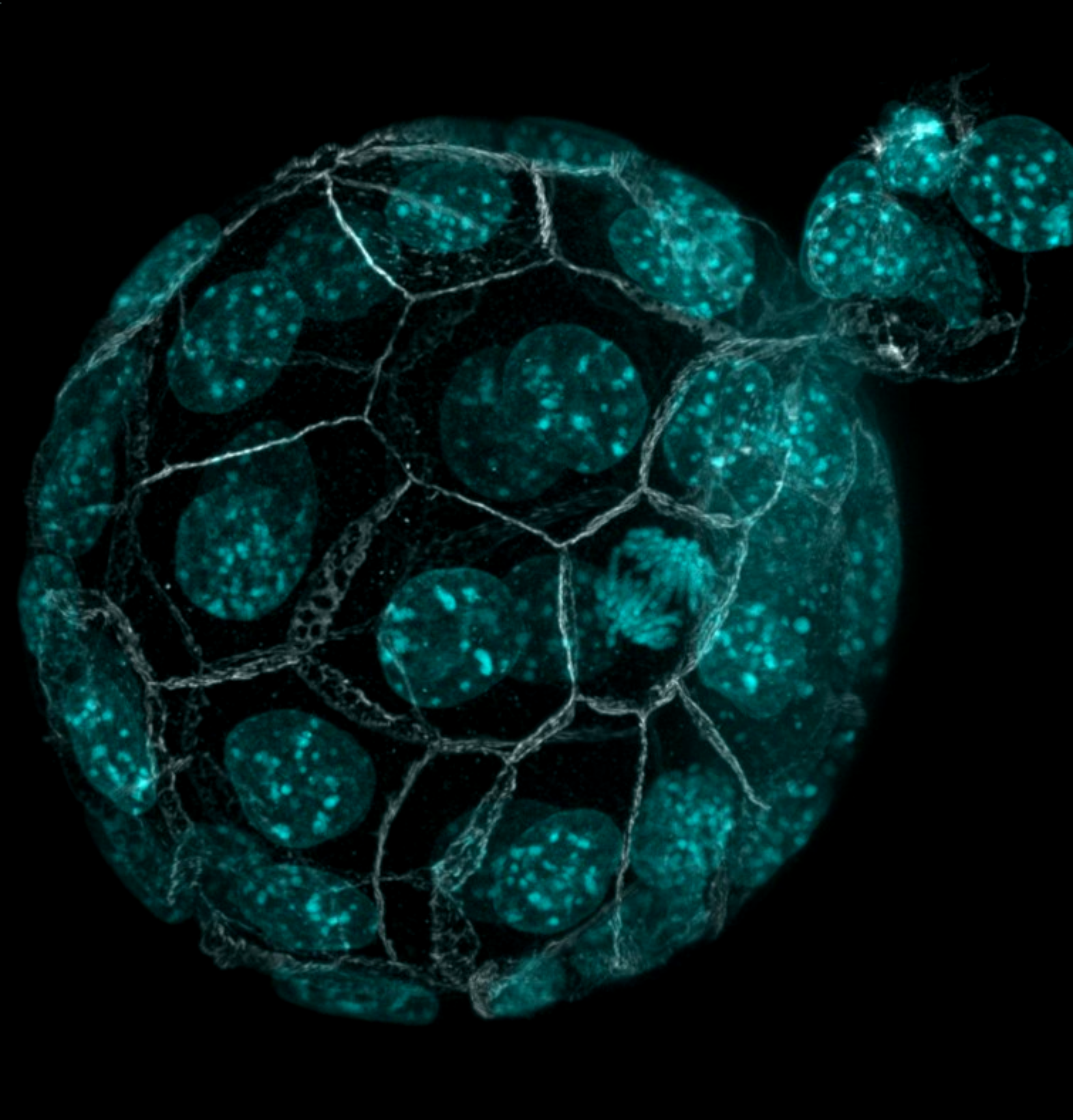
Immunostaining of a hatching mouse blastocyst imaged with confocal microscopy
Tarsal Claw
This scanning electron microscope (SEM) image of the tarsal claw of the horsefly (Tabanus sulcifrons) juxtaposes the complexity and simplicity of “nature’s Velcro.” The menacing sturdiness of the tarsal claws contrasts with the delicate nature of the tarsal pad, with fine, hooked hairs that allow the fly to hold on to animal fur.
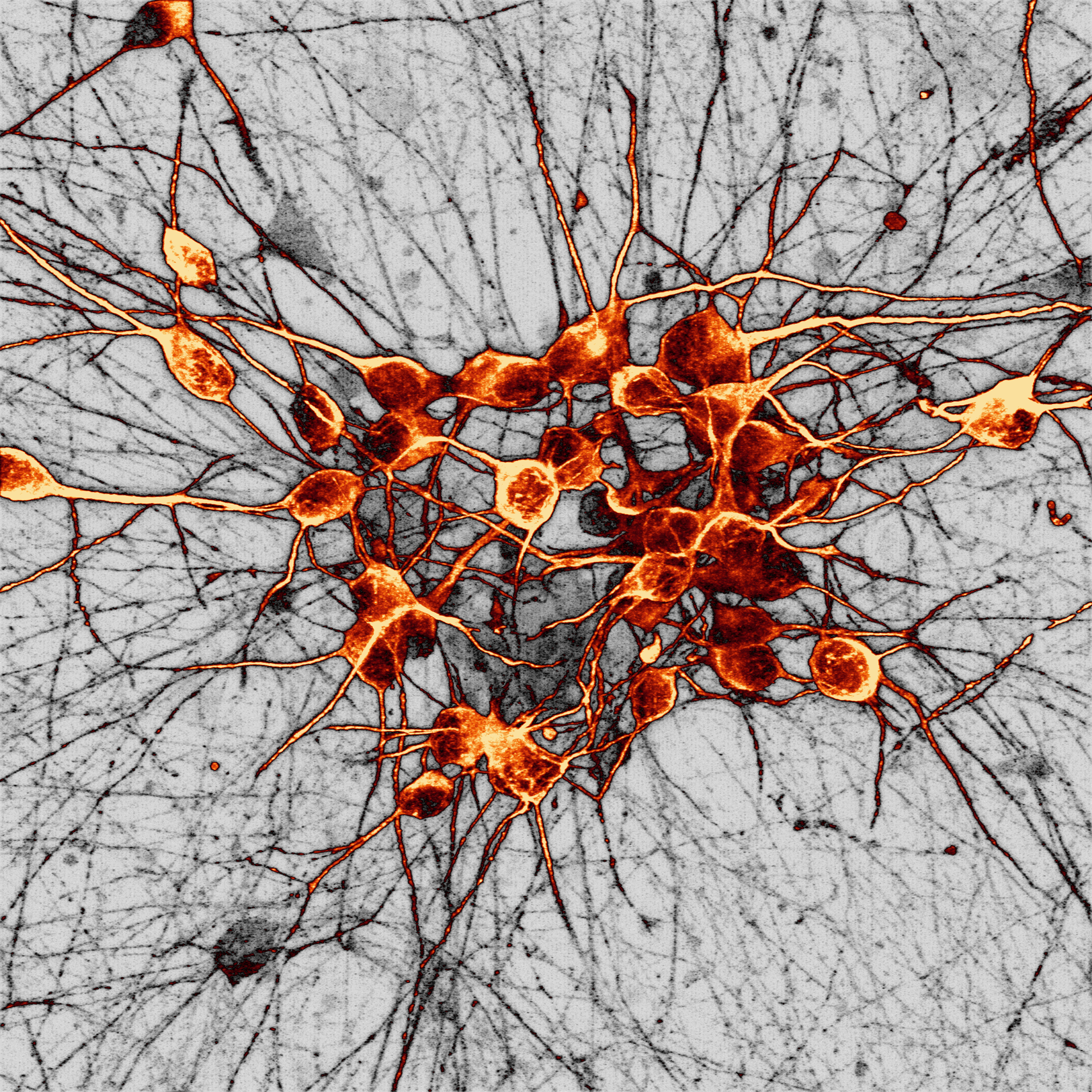
Human dopaminergic neurons
This image shows a culture of human dopaminergic neurons generated from human stem cells acquired via super-resolution Airyscan confocal microscopy at the University of Oxford Micron facility. Dopaminergic neurons are the main cell type that deteriorate in Parkinson’s disease, partly due to toxic build-up of a protein called alpha-synuclein.
Arabidopsis leaf
Cells on the epidermis of a 3 day old Arabidopsis leaf. This is an adaptation of a linocut print created based on a microscopy image.
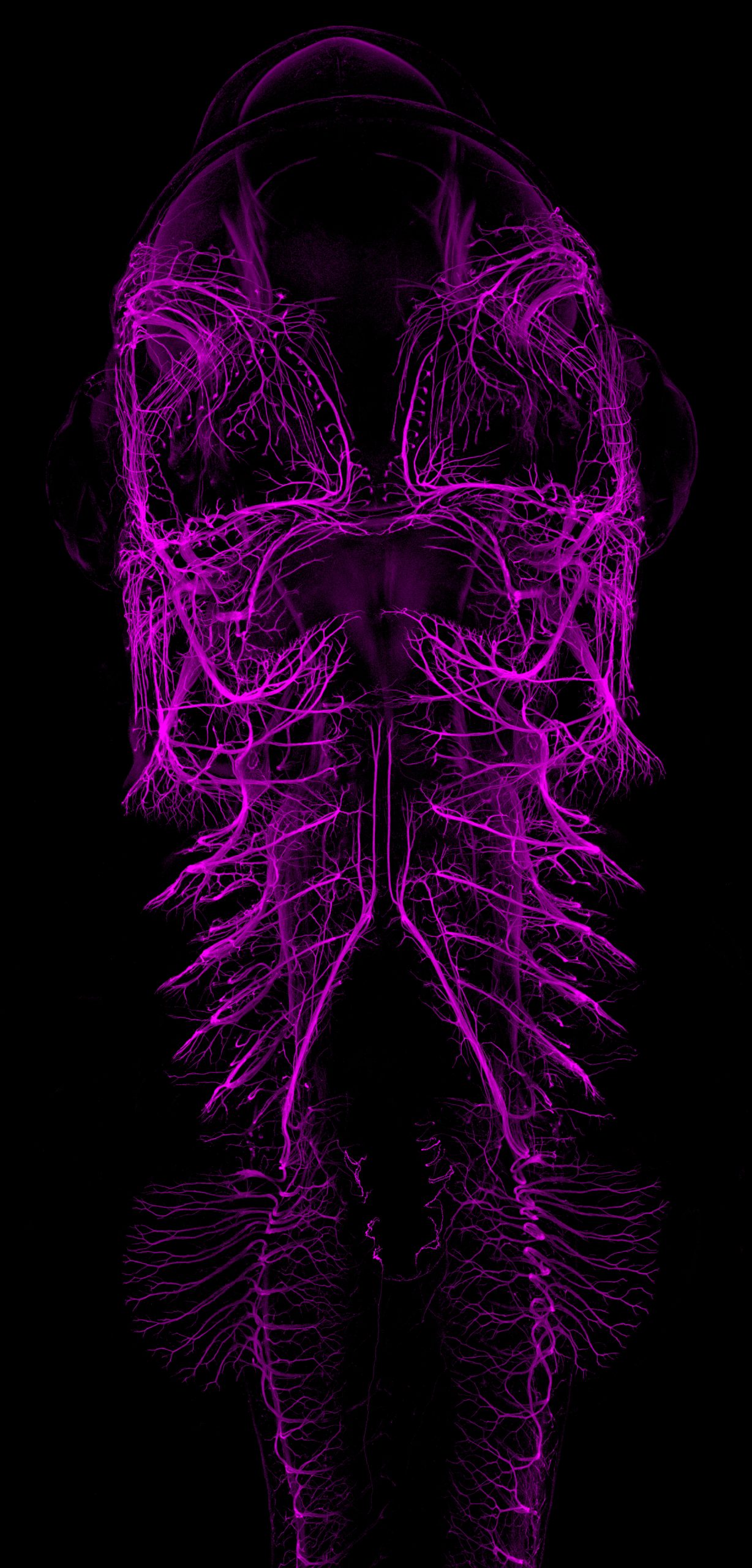
Catshark embryo
Ventral view maximum intensity projection from an immunofluorescence staining labeling the developing nervous system (primarily nerves and ganglia) of a stage 30 small-spotted catshark embryo (Scyliorhinus canicula). The image was acquired using a ZEISS LSM980 with Airyscan2 confocal microscope, stitched and processed using ZEN software from the same microscope.
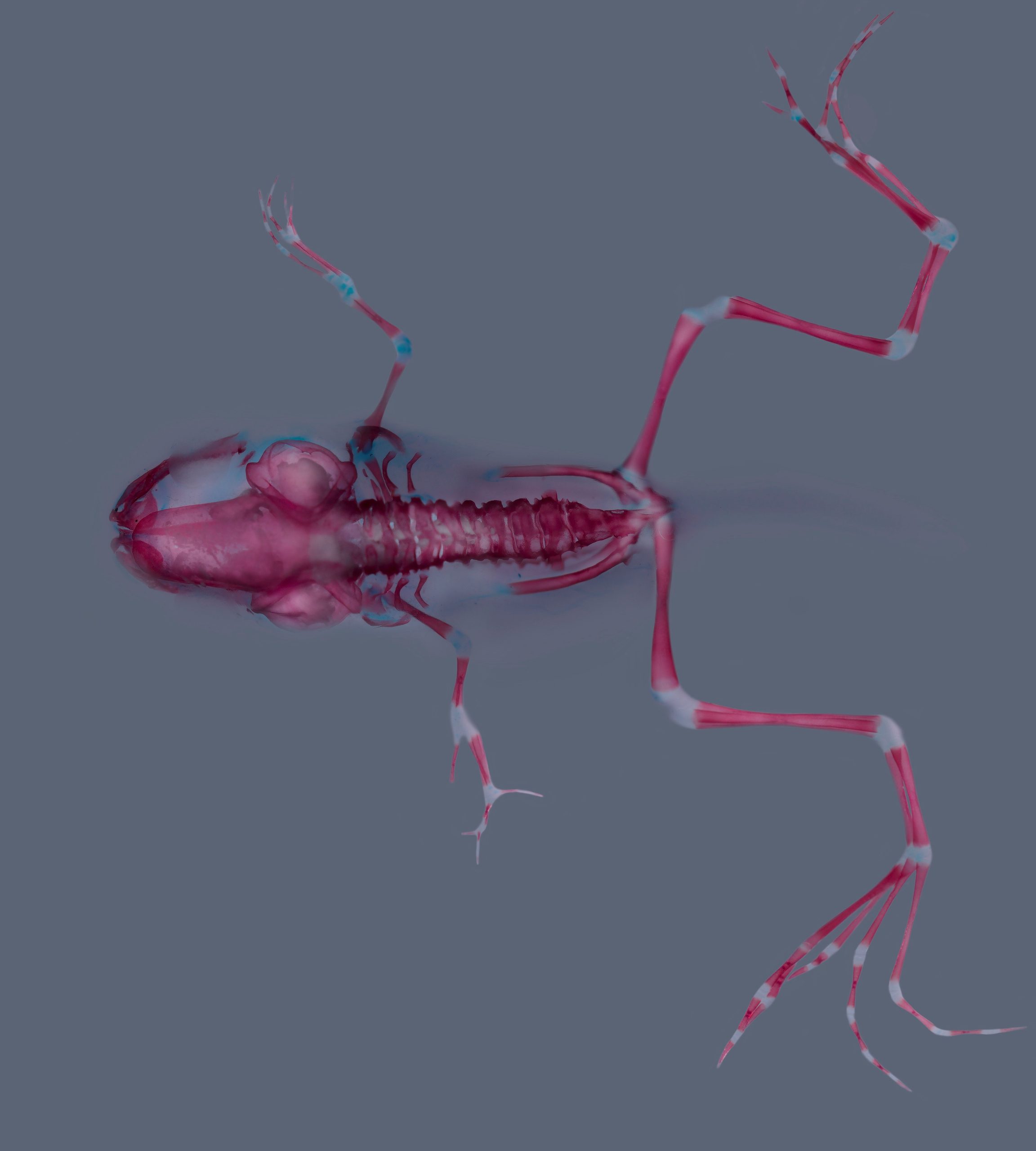
Xenopus laevis skeleton
Skeletal staining (alizarin red and alcian blue) of a Xenopus laevis at stage 62. Stage 51 larva was treated with a Cyp26a inhibitor during forelimb regeneration. Notice proximo-distal duplication in the left forelimb.
Austin Smith: We study pluripotent stem cells in vitro and their relationship to transient pluripotent cells in mammalian embryos. To execute their potential, naive pluripotent cells must gain lineage competence, a process termed formative transition. We want to understand:
how potency and competence are encoded by a dynamic regulatory network of signals, transcription factors and chromatin
how cells transition between states of competence and how fate decisions are made
how the trajectory and regulatory machinery of pluripotency is adapted in different mammals
The Smith lab
Lab roll call
Tao Huang, post-doc, establishing naïve pluripotent stem cells from non-human primates
Zhili Ren, post-doc, investigating self-renewal and formative transition in human naïve stem cells
Arthur Radley, post-doc, investigating cell identities and trajectories by computational analysis of transcriptome data
Francesca Carlisle, experimental officer, supporting stem cell culture and next generation sequencing, plus lab manager for the group
Zhi (Klein) Zhang, PhD student, studying transition from trophectoderm to amnion competence
Jing Yen Yong, PhD student, studying capacitation of naïve cells for definitive endoderm formation
Favourite technique, and why?
Austin: Stem cell culture, because it is life in our hands – a path to understanding and control.
Apart from your own research, what are you most excited about in developmental and stem cell biology
Austin: The regenerative plasticity uncovered in adult stem cell tissues such as intestine and lung is very interesting. The dedifferentiation seen after injury, or during cancer formation, is a striking contrast to how we generally consider developmental trajectories and more complex than a simple reversal process.
How do you approach managing your group and all the different tasks required in your job?
Austin: My role as Institute Director takes a lot of my time so the people in my group have to be quite independent. Joint lab meetings with the group of Ge Guo who also work on pluripotency are the main forum where group members present and get feedback on their research. I often follow up with 1 to 1 discussion. In general, I like to let people get on with their own experiments and ideas. I get more heavily involved as results emerge and require discussion. I usually take the lead in paper planning and writing.
What is the best thing about where you work?
Austin: The Living Systems Institute combines different research interests. The open space design of our building is ideal for research and social interaction. Labs, facilities, and technical support are all good. We have good community space including an outdoor terrace. The atmosphere is friendly and supportive.
What’s there to do outside of the lab?
Austin: Great outdoors along the river, the Jurassic coast, and Dartmoor, with endless opportunities for recreation, relaxation, and exploration
Browse through other ‘Lab meeting’ posts featuring developmental and stem cell biology labs around the world.
Anchel de Jaime Soguero (COS, University of Heidelberg) ‘Cell signaling control of genome stability during early lineage specification and neurogenesis’
Elena Camacho Aguilar (Rice University) ‘Combinatorial integration of BMP and WNT allows BMP to act as a morphogen in time but not in concentration’
Tyler Huycke (UCSF) ‘Patterning and folding of intestinal villi by active mesenchymal dewetting’
In the third episode of the Human Developmental Biology Initiative‘s new podcast, scientist Magomet ‘Mag’ Aushev, a postdoctoral researcher in Mary Herbert‘s lab at Newcastle University, meets Zara, a songwriter and performer from Manchester. The two discuss the very first cell divisions after an egg is fertilised and why a better understanding of very early development may impact fertility treatment and regenerative medicine of the future.
At the end of the episode, the pair write and record an original piece of music inspired by their conversation, exploring the science of human development in a brand new way.
“It really is like a universe on its own, a single cell.”
– Mag Aushev
Please subscribe and listen to Made the Same Way on Apple podcasts, Spotify, or wherever you get your podcasts. If you enjoy the podcast, please rate and review us on Apple podcasts to help others find us!
About the participants
Prior to joining HDBI, Magomet was applying genome editing for the treatment of genetic diseases and is now using it to generate reporter embryos to study early development. In his spare time Magomet works on machine learning pet projects and plays the guitar.
Zara is a song writer and performer – specialist mic controller (rapper)! She considers herself a philosopher, which comes across in her music, often covering about existential topics – love, happiness , grief, identity, self belief, overcoming trauma.
“By and large, animals have the same genes that you and I have. They’re just using them in a different way that makes it less likely for them to get a disease.”
Dr Linda Goodman, Fauna Bio
In the latest episode of Genetics Unzipped, we’re becoming chromosomal criminals and learning about how researchers are stealing genes from the animal kingdom and using them to improve human health. From 13-lined ground squirrels teaching us how to recover from heart attacks, to bowhead whales showing us how to avoid cancer, there’s a lot geneticists can learn from Mother Nature.
 (No Ratings Yet)
(No Ratings Yet)


 (35 votes)
(35 votes)

