Do you have an idea for a post on the Node that you have been postponing writing for a while? Or, perhaps, you have a draft that is waiting to receive your final touches before posting? Good news! This is your opportunity to publish your piece at last for a chance to win £200. For the upcoming month, we are hosting an exciting writing challenge to help battle procrastination and motivate you to share your piece with the Node’s community of developmental and stem cell biologists.
To take part in the challenge, simply post on the Node. The post can be related to current series and themes we have on the Node (or, perhaps, introducing some new exciting topic). All posts* between now and the deadline will automatically enter the pool. Register or log in to share your blog post with the community and enter. If you want to feel inspired, take a look at the amazing entries from our 2025 writing challenge.
The deadline for the challenge is 4 September, after which one winner will be randomly selected for a prize of £200 and an interview with the Node.
Cristian Cañestro’s lab offers a fully funded 4-year FPI PhD contract associated with our newly granted project PID2025-170547NB-I00, “Gene loss impact: evolution of cardioparaxial-neuromesodermal development and genome scrambling in Oikopleura dioica as a case study” (OikoLoss).
The PhD will be carried out within the Genetics Doctoral Programme at the University of Barcelona, in the Section of Genetics and the Biodiversity Research Institute (IRBio).
THE CANDIDATES:
We welcome candidates interested in evolutionary developmental biology, genomics, bioinformatics and evolutionary and population genetics.
Applicants must hold bachelor’s and master’s degrees in a field related to Biological Sciences, such as Biology, Genetics or Bioinformatics
THE PROJECT:
OikoLoss investigates how gene loss can drive evolutionary innovation. Using the chordate Oikopleura dioica as a natural “evolutionary knockout” model, the project integrates EvoDevo, functional genomics, single-cell transcriptomics, population genomics and ecology through two complementary aims:
Aim 1. To determine how massive gene loss has reshaped heart, axial muscle and neuromesodermal development in chordates, using single-cell omics, gene-expression analyses and functional experiments.
Aim 2. To uncover how genome scrambling and chromosomal breakpoints generate gene-loss alleles, and to assess their potential adaptive impact to ocean environmental conditions.
THE TECHNIQUES:
The PhD will involve embryo culture and manipulation, microinjection, CRISPR-knockouts, interference-knockdowns, HCR whole-mount in situ hybridization, confocal microscopy, single-cell RNA sequencing, PacBio HiFi long-read genomic sequencing, comparative and population genomics, and bioinformatics.
TRAINING AND COLLABORATIONS:
The successful candidate will join a multidisciplinary research environment and will enjoy opportunities to interact with international partners and undertake research stays, particularly with collaborators at OIST in Okinawa, IGFL in Lyon, SARS in Bergen and CRG in Barcelona.
FUNDING:
The position consists of a fully funded 4-year FPI predoctoral contract associated with the granted PID2025 project. Salary, doctoral fees, mobility support and social-security conditions will follow the official programme and University of Barcelona regulations.
DEADLINES:
Expressions of interest and pre-selection ARE NOW OPEN. Early contact is strongly recommended. The formal application procedure and incorporation date will follow the official FPI and UB timetable.
– The application process is expected to be open in September 2026
– Incorporation is expected at the end of 2026 or 2027.
CONTACT and SELECTION:
Interested candidates should email Cristian Cañestro at canestro@ub.edu, attaching a single PDF containing:
– A motivation letter explaining why they wish to join the laboratory and which project aims interest them most.
– A brief CV, including Bachelor’s and master’s academic transcripts, including average grades.
Shortlisted candidates will be invited to an online interview. Selection will follow the UB’s responsible recruitment principles and will be based on academic record, relevant experience and motivation, without discrimination and with attention to gender balance.
Further information, related publications and updates:
One limitation of comparative genomics and transcriptomics in understanding the origin of evolutionary novelties is that these approaches typically provide long (or short) lists of genes that differ among the species being compared, but they cannot distinguish causal mechanisms from subsequent adaptive outcomes. The former are most likely involved in the rewiring of gene regulatory networks (GRNs)—a process that does not necessarily require the evolution of new genes, but rather changes in regulatory interactions, including modifications of network connectivity, regulatory inputs, microRNA activity, epigenetic marks, or chromatin remodeling. By contrast, many of the latter changes, although biologically important, may simply reflect adaptation to new lifestyles, ecological conditions, or developmental modes that arose after the novelty itself had originated.
Because we have no direct access to the earliest stages in the evolution of new lineages, it is difficult to determine whether the gain or loss of particular genes played a causal role in the emergence of a novelty. Most extant animal groups diverged from their common ancestors hundreds of millions of years ago and have since experienced long and complex evolutionary histories. During these intervals, lineages presumably adapted to a succession of environments, many of which no longer exist. Such adaptive histories may have favored the expansion of particular gene families, thereby increasing organismal fitness under changing ecological conditions. Consequently, many genomic differences observed among living species may reflect long-term adaptive evolution rather than the mechanisms that originally generated evolutionary innovations. This highlights an obvious but often overlooked point: we have no direct record of the evolutionary trajectories that produced extant lineages. As a result, it is challenging to identify which genomic changes were instrumental in generating a novelty and which merely contributed to its subsequent refinement and ecological success. Ultimately, only regulatory changes can be considered causal in the origin of developmental novelties, since morphological innovation necessarily arises through modifications in the regulation of development. Investigating regulatory genes, their interactions within GRNs, and the molecular mechanisms that modulate them therefore offers the best opportunity to uncover the causal processes underlying evolutionary innovation.
Once these primary regulatory changes are established, subsequent evolutionary refinements can occur through selection acting on downstream targets, including structural, metabolic, physiological, and additional regulatory genes. These later modifications fine-tune the phenotype and improve its performance in particular ecological contexts, but they should not be conflated with the original causal events that produced the novelty.
The central argument I wish to make, therefore, is that research on evolutionary novelties should place greater emphasis on identifying causal mechanisms by reconstructing how GRNs have been modified over evolutionary time. This requires refocusing our attention on the primary regulatory drivers rather than on the accumulation of downstream genomic differences. Moreover, integrating comparative genomics with evidence from the fossil record and with reconstructions of the ecological histories experienced by different lineages would provide a much richer framework for interpreting differences in gene complements. Such an approach would move comparative studies beyond sequence comparisons alone toward a functional understanding of regulatory genes and their evolutionary roles across the animal tree of life. Although this perspective is necessarily nuanced and difficult to implement, it offers a more promising route toward understanding how evolutionary novelties arise through causal developmental mechanisms.
Though the above doesn’t represent a particularly new idea, it is, in my view, a useful reminder of what we need to focus on in order to understand novelty.
Disclosure: ChatGPT was used solely to improve the language and style of the original text.
The adventure of a brave ENGAGE-Bio post-bac scholar at the MBL who set up a new sea cucumber species in the Perillo lab.
Who are we?
Hi, we are a group of scientists who believe establishing new research systems is key to discover new biological features. We work at the Marine Biological Laboratory (MBL) in Woods Hole (MA, USA) in the Perillo lab. Talia (now a graduate student at the University of Virginia) was an ENGAGE BIO post-bac scholar who spent a year establishing the sea cucumber Leptosynapta tenuis as a new lab system.
What are sea cucumbers?
Sea cucumbers are marine invertebrates with amazing abilities: these animals can undergo whole body regeneration, spit out their internal organs if threatened -and completely regenerate them! – change skin strength and elastic stiffness within a timescale of seconds and are rich in novel active bioactive compounds -just to cite a few.
Almost all species of adult sea cucumbers have dark, thick skin and studying their organs in vivo is a challenge. Moreover, the most common species can only be studied close to their collection site, as these animals do not ship well. However, thanks to the knowledge of the Marine Resource Center at the MBL we have access to a local sea cucumber species that has transparent skin and that we found can be easily kept in any lab. These new species is the sea cucumber L. tenuis, a small and clear sea cucumber from the North Atlantic Ocean (Fig. 1A). They feed in the sand and use their tentacles to quickly dig and hide. Not much is known about the organism in terms of its characteristics and behaviors and we are excited to discover new aspects of their biology every day. How do they navigate in space inside the sand? How is the nervous system controlling their behaviors? How do they reproduce? How does a hermaphrodite gonad work? How do their larvae develop? These are some of the questions we are investigating in the lab.
Talia worked on some of these questions and had a chance to present her work at the developmental Biology of the Sea Urchin and Other Marine Invertebrates (DBMUMI) conference (Fig. 2A). In 2026 Aly Rodger, new Research Assistant in the lab, took over the sea cucumber project.
Figure 1:A) Leptosynapta tenuis digging in the sand in our lab setup. B) Live imaging of L. tenuis skin stained with a DNA dye.
Talia’s experience working with sea cucumbers
Due to the niche of this animal, I was interested in Margherita’s project to characterize its structures. When we received a collection for the month, we would dissect and image to understand different structures. We can conduct living imaging of their skin because it is transparent (Fig. 1B). Through this, we were able to observe structures on the skin that we could have seen if they had pigment, as most sea cucumbers do. I spent time staining phalloidin and tubulin to understand the morphology of the skin, intestine, tentacles, and the gonads. While imaging new structures, I was amazed by the amount of muscle and neurons that these small sea cucumbers had (Fig. 2B, Talia working). L. tenuis is a hermaphrodite species, how do they develop both testis and ovaries in the same tube? To understand their oocyte growth, I would image them monthly and measure their size to figure out their seasonal patterns and compare it to testis maturation. Being the first to see these structures through fluorescence microscopy only made me question their development more.
A challenging part of working with these animals is that little is known about their development or their behaviors. Margherita and I spent a lot of time troubleshooting how to house them at the MBL and within our lab. We came up with a water bubbling system to cycle the sand for oxygen flow within their habitat and a sand collection system to ensure their environment has nutrients. Even though trying to understand how to maintain them is difficult, it is exciting to be able to lay the groundwork for a new model system in developmental biology.
Why do we need this for developmental biology?
Sometimes in research, we cannot answer a question because we don’t have the tools or the right model. But the sea cucumber has the possibility to be used in different fields; it is worth investigating and conducting groundwork research for future research to answer larger questions with this organism. Similarly, groundbreaking science came from taking a chance and investing resources in what are now considered traditional research organisms, such as mice, flies, bacteria, and zebrafish. I believe that many research questions, such as neurodevelopment, axis elongation, and body plan development, could be answered by using L. tenuis.
Figure 2: A) Talia presents her work on sea cucumbers at the international conference for the Developmental Biology of the Sea Urchin and Other Marine Invertebrates in 2025. B) Talia taking pictures of sea cucumber gonads at the confocal microscope. (1 votes) Loading...
Mackem lab is recruiting for NIH-funded post-doc position in developmental morphogenesis to study the roles of Shh in vertebrate limb patterning (different digit types, numbers, adaptations) using molecular-genetic and genomic approaches. Our long-term goal is understanding how regulatory networks instruct the formation of complex structures with distinct shapes, such as the varying bony segments and joint numbers in different digits, arising from the same tissues and not based in cell fate changes per se.
Current work focuses on genome-wide approaches to identify key Shh relay signals and their relation to late signaling centers, comparative evolutionary analyses to reveal regulatory mechanisms underlying morphology-based adaptations and combined genetic/genomic strategies to explore the dynamic, robust nature of Shh-producing cells. We recently showed that Shh acts as a trigger, not morphogen, to initiate digit patterning, but the critical targets remain unknown. We have discovered that the Shh-producing ZPA domain in the limb is not a static population but arises from progenitors at the limb border that contribute to ongoing ZPA renewal and plan to characterize progenitor pool regulation. We have also found that the widespread phenomenon of autocrine non-responsiveness in Shh-producing cells is an intrinsic feature of these cells, not due to negative-feedback, and are examining its mechanistic basis.
Applicants should have a strong background in developmental and/or evo-devo biology. Prior experience with cutting-edge genomic approaches is desirable. For more information, please contact Susan Mackem (mackems@mail.nih.gov) directly.
Recent lab publications:
Huang et al. (2026) Nature Communications. A pivotal Wnt antagonist role promoting digit joint specification by constraining Wnt activity. In press May 2026. doi.org/10.1038/s41467-026-73549-4
Patel and Mackem (2025) PNAS. Dual Bmp-negative feedback loops modulate function of both AER and ZPA to buffer and constrain postaxial digit number. 122: e2427249122. doi.org/10.1073/pnas.2427249122
Zhu et al. (2022) Dev Cell. Sonic hedgehog is not a limb morphogen but acts as a trigger to specify all digits in mice. 57: 2048-62.
Trofkaet al. (2021) Current Biology. Genetic basis for an evolutionary shift from ancestral preaxial to postaxial limb polarity in non-urodele vertebrates. 31: 4923-34.
Reviews/perspectives:
Huang et al. (2022) Dev. Dynamics. Rethinking positional information and digit identity: The role of late interdigit signaling. 251:1414–1422.
Zhu et al. (2017) Dev. Biology. John Saunders’ ZPA, Sonic hedgehog and digit identity – How does it really all work? 429:391-400.
Last month, the Node ran a cover competition with images taken by advanced graduate students and postdoctoral fellows from across the Americas who gathered in Chile to participate in the International Course on Developmental Biology, an EMBO Practical Course held at the Marine Biology Station of Quintay (CIMARQ).
We held a vote over a two-week period. Nine impressive submissions were received from the 2025 student cohort, with images ranging from amphipods to Xenopus laevis and the winners of the popular vote were chosen. Among the great selection of images, the image taken by Natália Heloísa de Oliveira, Fernanda Salce, and Federica Salatiello, Amphipod, received the most votes. Congratulations! Their image will be published on the front cover of Development later in the year.
First, we hear from one of the winners, Fernanda Salce.
Amphipod. Amphipod collected in Quintay Bay by students of the 2025 course, and stained with phalloidin and DAPI.
Fernanda, can you describe your research career so far?
Currently, I am a PhD student in Brazil at the Federal University of Rio de Janeiro, Macaé, RJ. Specifically, I work at the Institute of Biodiversity and Sustainability, affiliated with the Morphological Sciences PhD program.
Can you tell us about your current research?
I am focusing on Developmental Biology as I study the regenerative process of locomotor limbs in the beetle Tribolium castaneum. I am trying to analyse how transcriptional perturbations retain enough plasticity to ensure a given biological process is completed. Thus, I have been studying the limits of leg regeneration in Tribolium larvae under different zld dsRNA concentrations. In previous studies, our group showed that zld is a pioneer transcriptional factor whose activity is pivotal to the success of leg regeneration in the Tribolium larvae model, with zld dsRNA treatment fully inhibiting this process. Here, I am investigating at which point this system might recapitulate function even in the partial absence of zld mRNA. So, more specifically, I am trying to observe whether the regenerative process can be characterized as a bimodal switch-like system and, furthermore, which genes are integral and which are dispensable for leg regeneration in a zld-dependent manner. In parallel, I am also observing changes in morphospace exploration during leg regeneration under these different zld dsRNA concentrations.
What is your favourite imaging technique/microscope?
I frequently use the stereomicroscope for imaging. However, after the course in Chile, I have to say that Light-Sheet microscopy has become my new favorite technique. Not for its accessibility, mind you! But for the amount of morphological data you can extract from your samples. It is simply astonishing!
What is the most impactful thing you learned in the course?
That people are more accessible and approachable than they might seem at first. I would never have thought in my life that Roberto Mayor and I would be sharing drinks and grilling steaks together, but there we were, haha! But on a serious note, the course truly highlights how important social interactions and interpersonal skills are for a scientist.
If possible, describe a bit more about the image that made the cover, including what seems to be capturing wild-caught animals!
This is a crustacean in the genus Parhyale. We caught this bad girl with Nipam Patel’s help. He told us to put a few carrot pieces inside a plastic water bottle with some stones to attract a few of these animals overnight. Keep in mind, this was done inside an aquarium-like system with thousands of red sea urchins. The next day, we actually caught some and thus began our fishing minigame, using Pasteur pipettes and clove oil as anaesthetic. This specimen was particularly interesting because it was a female carrying eggs, so we separated the mother for the Light-Sheet microscope and the eggs for downstream analysis on the confocal microscope. The latter was also brought to Chile with Nipam’s aid.
As a side note, the confocal microscope had an extraordinary logistical undertaking behind it, from what we were told. Apparently, each piece of equipment had to be labelled and registered upon entry into the country, so this particular confocal microscope was designed to be “easily” assembled and disassembled. Truly mesmerizing stuff.
Lastly, for staining, we left our specimen in a phalloidin solution for a week, then stained it with DAPI before our first use of the Light-Sheet microscope during the course. Retroactively, I don’t think that we stained for enough time because you’ll notice that there are myofibers clearly visible in the extremities, but on the animal’s central axis, there was barely any staining – the few observations we caught were restricted to the head and portions of the tail. So, for future participants, my advice would be to begin staining as early as possible and to visualize your possible findings during the last week of the course for better results 😊.
The other two winners, Federica Salatiello, who was funded by the BEOM Department, Stazione Zoologica Anton Dohrn, Italy, and Natália Heloísa de Oliveira, a PhD student at the Federal University of São Paulo, Brazil, under the supervision of Prof. Dr. Marimelia Porcionatto, will be interviewed and featured on the Node later in the year.
Thank you to all the students who participated in the course: Victoria Fernandez, Ana Maria Soria, Candelaria Diaz, David Arancibia, Emilia Failache, Erika Venancio, Federica Salatiello, Fernanda Salce, Fernanda Dalami, Gonzalo Spelzini, Ignacio Casanova, Ingrid Pinto, Jaime Aguayo, Juan Pablo Venegas, Luis Eduardo Sánchez , Marco Mundaca, Mariana Tovar, Natália Heloísa de Oliveira, Nicolás Zúñiga and Pablo Fernández.
Did these images inspire you to take the course? The next cohort of the International Course on Developmental Biology is currently accepting applications until 30 July 2026.
A scientists we tend to move around a lot during our training period. Adjusting to new scientific and social environments can be challenging and intimidating but also exciting. Andrew Ramirez writes about navigating this process with the right mindset and some useful strategies.
We are delighted to announce the winners of the Young Embryologist Network Image Competition held as part of the YEN 2026 Conference, which took place on the 15th June at the Francis Crick Institute. A huge thank you to all 30 participants who submitted their beautiful images.
The winning image, Lamination in the Zebrafish Retina by Jack Nicholls, will be featured in the YEN 2027 promotional materials. Congratulations to all of our shortlisted entrants!
Winning image: Jack Nicholls – Lamination in the Zebrafish Retina
Runner-up: Ipek Gassaloglu Guler – The First Contact: Interaction Between an Embryo Model and Maternal Tissue
Runner-up: David Arancibia-Altamirano – Aristotle’s Egg Through a New Lens
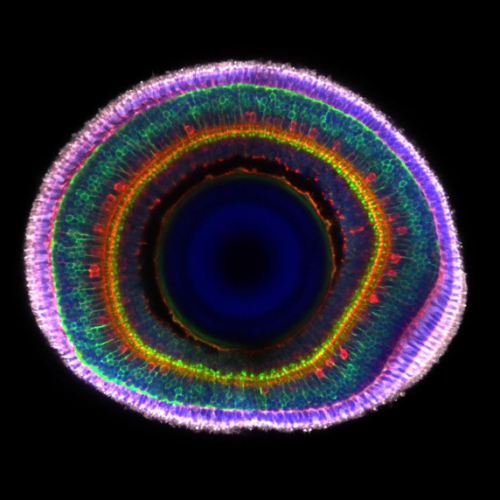
1. Lamination In The Zebrafish Retina
Jack Nicholls, City St George’s, University of London
Immunohistochemistry of an 8-day post-fertilisation larval zebrafish retina stained with anti-ChAT in red (star burst amacrine cells), anti-PKCα in green (ON bipolar cells) and grey (photoreceptors), with a DAPI counterstain. The anti-PKCα signal was manually separated into the bipolar and photoreceptor cell regions and coloured in green and grey to highlight the boundary between the two layers. Imaged on a LSM800 confocal microscope.
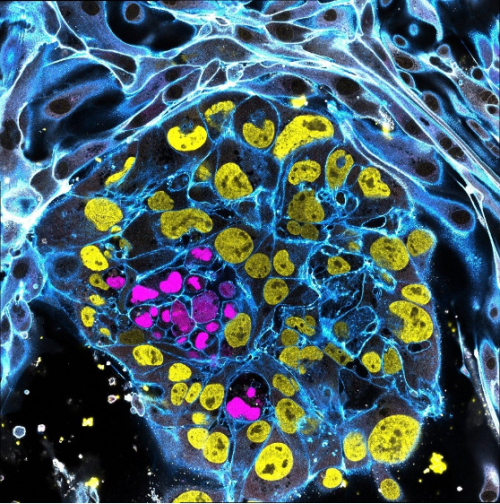
2. The First Contact: Interaction Between an Embryo Model and Maternal Tissue
Ipek Gassaloglu Guler, Yale University
This image portrays a stem cell-derived human embryo model (blastoid) cultured on endometrial epithelial cells. This experiment aimed to visualize the initial contact between the blastoid and endometrial epithelium, to capture one of the earliest stages of embryo-maternal interaction.
Within the blastoid, the epiblast is shown in magenta (SOX2) and the trophectoderm in yellow (GATA3). To highlight the interaction between the blastoid and maternal cells, integrin B1 (ITGB1) is shown in white. F-ACTIN, which outlines cellular architecture is shown in blue. The image was acquired using a Leica STELLARIS 5 confocal microscope in tile-scan mode with a 63x objective.
3. Aristotle’s Egg Through a New Lens
David Arancibia-Altamirano, University College London, Universidad Mayor
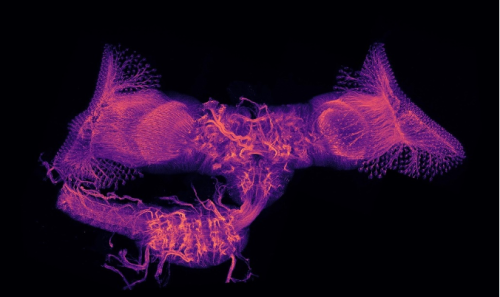
Since Aristotle’s studies in embryology, the chicken embryo has captivated the imagination of scientists and spurred exploration of epigenesis and preformation. Echoing classic experiments with India ink, fluorescent ink injection combined with tissue clearing and light-sheet microscopy now allows us to study the beauty and complexity of vascular networks in 3D and with high resolution. Sample displayed by depth colour-coding across 3.1mm; prepared by me and my friend, Jesus Juarez.
4. Dancing Ghosts
Ornella Clara, Aix-Marseille University
At first glance, the image resembles a ghostly choreography suspended in a cosmic landscape. In reality, it captures the collective cell migration during the early formation of a gut-like tissue structure. These gastruloids spread on a laminin-coated substrate, self-organizing into patterns reminiscent of embryonic development. The transcription factor CDX2 (light blue) marks intestinal identity, while phalloidin staining (yellow-orange) highlights the actin cytoskeleton that drives cell movement, and nuclei are shown in blue-violet.
This image was acquired using confocal microscopy. It illustrates how coordinated cellular behaviour gives rise to complex tissue architecture.
Experimental work by Dalia El Arawi; staining and confocal imaging by Ornella Clara.
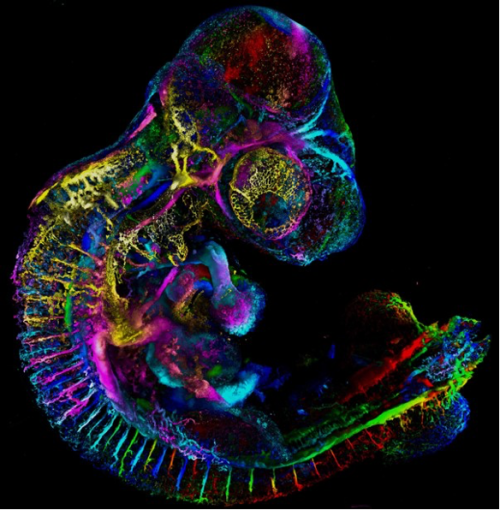
5. Growing Into Form
Matyas Bubna-Litic, University College London
A one-day old zebrafish embryo already has recognisable features such as the early forms of the eye and ear as well as the segmented backbone, which will go on to form vertebrae and muscle. Cell nuclei and filamentous actin are visualised in this stained fixed sample. Imaged using a Zeiss LSM980 with an Airyscan2 detector in multiplex mode.
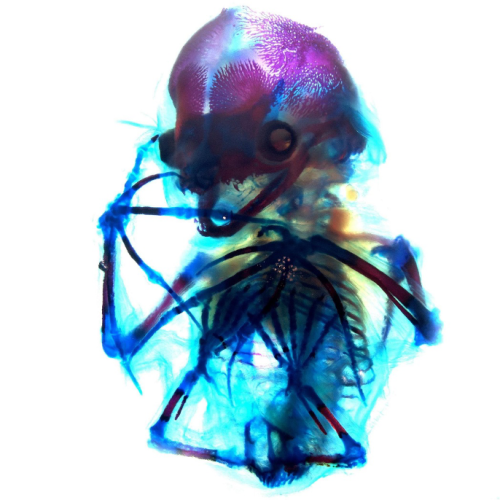
6. Down to the Bone
Alexandra Lion, Brigham and Women’s Hospital and Harvard Medical School
A beautiful network of bones and cartilage, which in life would provide support, protection and facilitate movement of the body. The image shows a short-tailed fruit bat (Carollia perspicillata) embryo at embryonic stage 22 which has been cleared and stained with alcian blue for cartilage and alizarin red for bone. This staining allows for visualization of the still-ossifying bones of the bat autopod, and most strikingly of the skull, which appears to smile with clearly visible canine teeth. Imaged during the 2024 Embryology Course at the Marine Biological Laboratory using transmitted light on a Zeiss Axio Zoom.V16 microscope, then further processed using FIJI.
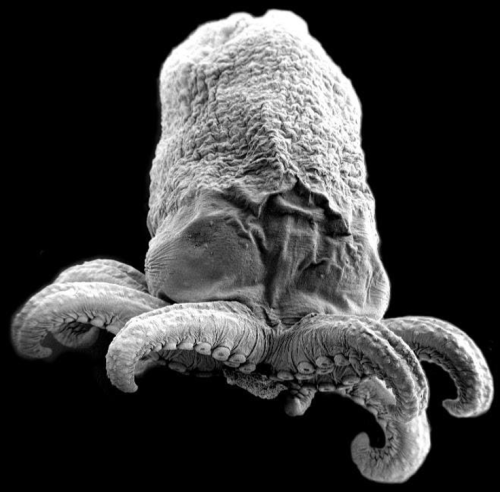
7. Octopus Embryo
Ailen Cervino Len, Baylor College of Medicine
Octopus embryo taken with Scanning Electron Microscopy (SEM).
8. Wiring Diagram Ryan Cheng, Centre for Developmental Neurobiology, King’s College London
Maximum intensity projection image of the central nervous system of Drosophila melanogaster during metamorphosis. This sample was collected at around 24 hours after puparium formation and stained with an anti-Neuroglian antibody to visualize the neurite tracks. Multiple z-stacks were imaged on a Zeiss LSM800 and reconstructed in FIJI.
9. The Phases of Gastrulation Hoang Anh Le, University College London
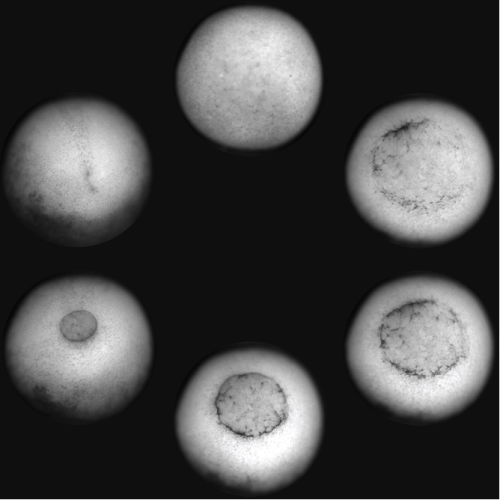
A Xenopus laevis embryo was imaged from the ventral side showing the different phases of gastrulation, from the formation of the blastopore lip to its closure and the beginning of neurulation. The embryo was imaged with an upright brightfield microscope.
10. beCOWming
Noemi Monferini, Developmental Biology Institute of Marseille
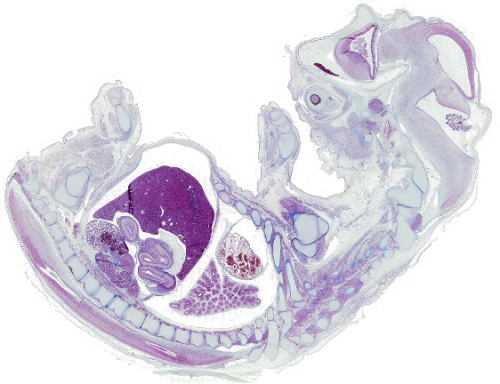
A bovine foetus in histological section stained with Azan trichrome, revealing the delicate architecture of foetal tissues.
We would also like to highlight the winner of the public vote, which has received 128 out of the 773 votes cast:
Weaving a Nervous System
Lamiya Dohadwala, Tata Institute of Fundamental Research
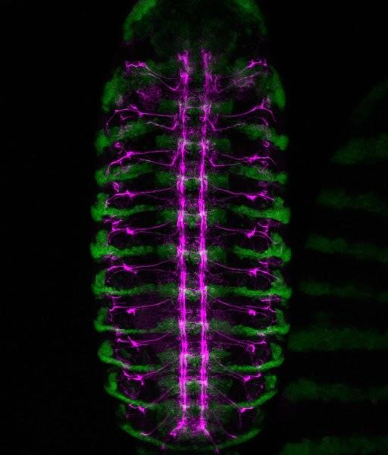
A confocal view of the developing central nervous system in a Drosophila embryo. Green marks engrailed-expressing segmental compartments, while magenta highlights Fasciclin II-positive nerve fibres, tracing the intricate network of axon pathways that form the embryonic nerve cord.
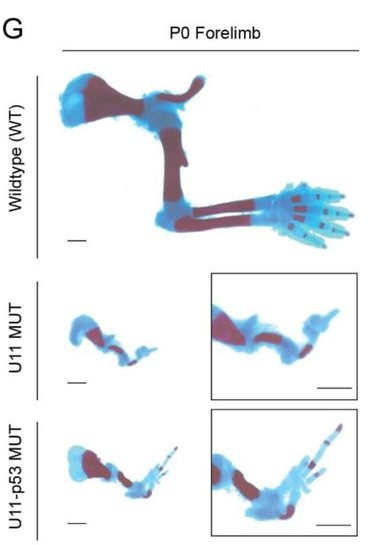
A Perturb-seq screen guided by species divergence uncovers pathways for collateral artery formation Xiaochen Fan, Ronghao Zhou, Brian C. Raftrey, Pamela E. Rios Coronado, Emily Trimm, Erin Clancy, Xinhong Chen, Jamie Bozeman, Maggie S. Chen, Shoxruxxon Alimukhamedov, Juan Alcocer, Idalina Bonham, Stuti Agarwal, Alina Isakova, Vinicio A. de Jesus Perez, Chong Y. Park, Timothy F. Shay, Viviana Gradinaru, Thomas Quertermous, Jesse M. Engreitz, Kristy Red-Horse
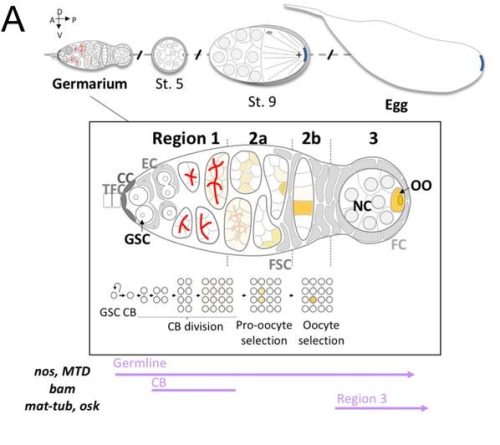
Developmental genetic response of the zooplanktonic tunicate Oikopleura dioica to marine noise pollution Eva R. Quintana, Nuria P. Torres-Águila, Ignasi Nou-Plana, Sissel Norland, Valentina Caorsi, Giorgio Blumer, Matteo Bozzo, Elettra Panarari, Giacomo Sabaddin, Simona Candiani, Irene Guarneri, Lucia Manni, Roberta Pennati, Filomena Ristoratore, Giovanni Zambon, Marios Chatzigeorgiou, Rosa Maria Alsina-Pagès, Cristian Cañestro
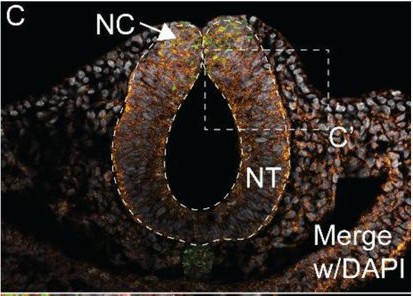
Modeling human B cell development with pluripotent stem cells Xiaoning Sun, Jamie J. Kwan, Krishna Kothari, Alexandra F. Nazzari, Astrid Kosters, Colin A. Fields, Bao Q. Thai, Deepta Bhattacharya, Michael Atkins, Kelvin Chan Tung, Xinyuan Zhao, Vladimir T. Manchev, Marion Kennedy, Eliver Ghosn, Gordon Keller
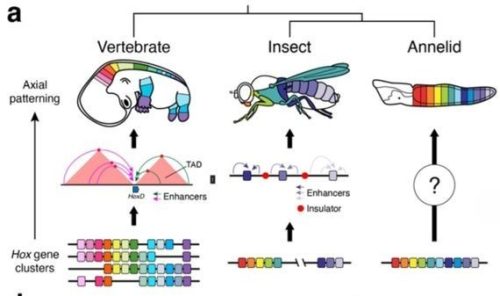
Convergent evolution of cluster-wide Hox gene regulation in Bilateria Billie E. Davies, Francisco M. Martín-Zamora, Tom Frankish, Elise Parey, Nancy Ellis, Noura Maziak, Kero Guynes, Grygoriy Zolotarov, Yi-Jyun Luo, Ferdinand Marletaz, Juan M. Vaquerizas, Arnau Sebé-Pedrós, Nicolae Radu Zabet, Paul J. Hurd, José M. Martín-Durán
The SPARK complex forms the molecular basis of vertebrate fertilization Victoria E. Deneke, Johannes P. Suwita, Haoting Wang, Shingo Tonai, Yonggang Lu, Karin Panser, Alexander Schleiffer, Jeremy A. Hollis, Maria Novatchkova, Gerhard Dürnberger, Karel Stejskal, Gabriela Krssakova, Andreas Blaha, Aleq Adrianne R. Andresan, Muriel Mirus, Hana Marvanova, Hsin-Yi Chang, Taichi Noda, Alejandro Burga, Elisabeth Roitinger, Masahito Ikawa, An-drea Pauli
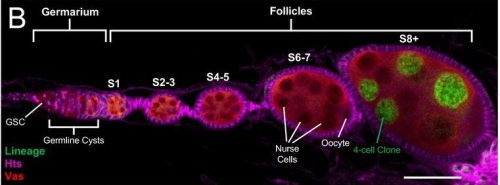
Nucleolar Dynamics During Oogenesis Ruoyu Li, Grace McKown, Dai Tsuchiya, Mark Mattingly, Anna Galligos, Michay Diez, Jui Feng Lu, Mary C McKinney, Sean McKinney, Boris Rubinstein, Timothy J Corbin, Melainia McClain, Carrie Carmichael, Victoria A Hassebroek, Stephanie H Nowotarski, Jennifer L Gerton, Kamena K Kostova
In vitro sexual dimorphism establishment in schistosomes Rémi Pichon, Magda E Lotkowska, Jude L. D. Bulathsinghalage, Madeleine McMath, Mary Evans, Benjamin J. Hulme, Kirsty Ambridge, Geetha Sankaranarayanan, Simon Kershenbaum, Sarah D. Davey, Josephine E. Forde-Thomas, Karl F. Hoffmann, Matthew Berriman, Gabriel Rinaldi
Comprehensive Lineage Tracing Maps the Landscape of Cell Fate Decisions in Mouse Embryogenesis William N. Colgan, Luke W. Koblan, JoAnne Villagrana, Tien-Chi Jason Hou, Minming Wang, Gokul Gowri, Whitney Chandler, Leonardo A. Sepúlveda, Didar Ciftci, Karina Smolyar, Alicia Young, Lars Wittler, Styliani Markoulaki, Kyle Loh, Xiaowei Zhuang, Nir Yosef, Zachary D. Smith, Jonathan S. Weissman

 (No Ratings Yet)
(No Ratings Yet)
 (1 votes)
(1 votes)