“The notion that you can use blood tests that look at circulating DNA to understand whether people do or don’t have cancer and how they’re likely to do will totally transform cancer care. I’m quite confident of that”
Sir Harpal Kumar, President of GRAIL Europe
In the latest episode of the Genetics Unzipped podcast, we’re taking a closer look at the red stuff, finding out what a few millilitres of blood can reveal about the development, progression and treatment of cancer within the body. Rather than painful surgical biopsies, expensive scans or complicated screening tests, what if we could simply take a small tube of blood and discover a wealth of information, such as whether or not you have cancer in your body, where it started, how to treat it, and whether that treatment is actually working?
Presenter Dr Kat Arney finds out what circulating tumour DNA is from Dr Susan Galbraith, how it can be used to monitor the progression of a cancer from Professor Charles Swanton, and what this will mean for future cancer patients from Sir Harpal Kumar.
If you enjoy the show, please do rate and review on Apple podcasts and help to spread the word on social media. And you can always send feedback and suggestions for future episodes and guests to podcast@geneticsunzipped.com Follow us on Twitter – @geneticsunzip
We are currently asking for feedback on our Development presents… webinar series and invite you to complete a short survey using the button below. The survey will close on Wednesday 6 July.
During the pandemic, Development were thinking about ways in which we could support the developmental and stem cell biology communities during this period. What could the journal could offer to substitute the lack of in-person interactions with digital opportunities? We experimented by bringing some virtual elements to the eBSDB/GenSoc 2020 meeting and by the organisation of the Node 10th birthday networking event to bring scientists together in a fun and flexible way. An initial survey of the community determined that the vast majority of respondents (largely based in Europe and the Americas) would be interested in a webinar series featuring developmental biology scientific talks from a mix of early career researchers (ECRs) and principal investigators (PIs) (Fig. 1).
Fig. 1. Initial survey. The bar chart above shows the percentage of respondents for each of the following questions: 1) Would you be interested in a scientific webinar series with talks from developmental biologists? Yes (111/113); No (2/113). 2) Who would you prefer for us to feature? ECRs (12/111); PIs (8/111); A mix of the two (91/111). 3) In which time zone are you based? Asia/Australasia (7/113); Europe/Africa (61/113); Americas (45/113).
Aims
Highlight recent published or soon-to-be published Development articles and interesting preprints. We want to promote the authors of great Development papers, as well as to recognise the growing preprint literature and signal our support. We run the webinars monthly with a different Development Editor chairing each month. The webinars are made up of three short (12-15 minute) talks: two of the speakers are selected from Research Articles handled by the chair and the third speaker is chosen from an interesting preprint in the Editor’s field.
Promote ECRs. Along with various other initiatives in Development, we want to champion ECRs, both by promoting their science and by providing opportunities for career development. We encourage the first authors of the selected papers to present at the webinar, if possible, hoping that reflects greater diversity in career stage than only inviting the corresponding author. We hope that the webinar series provides a forum for ECRs to discuss both of these topics.
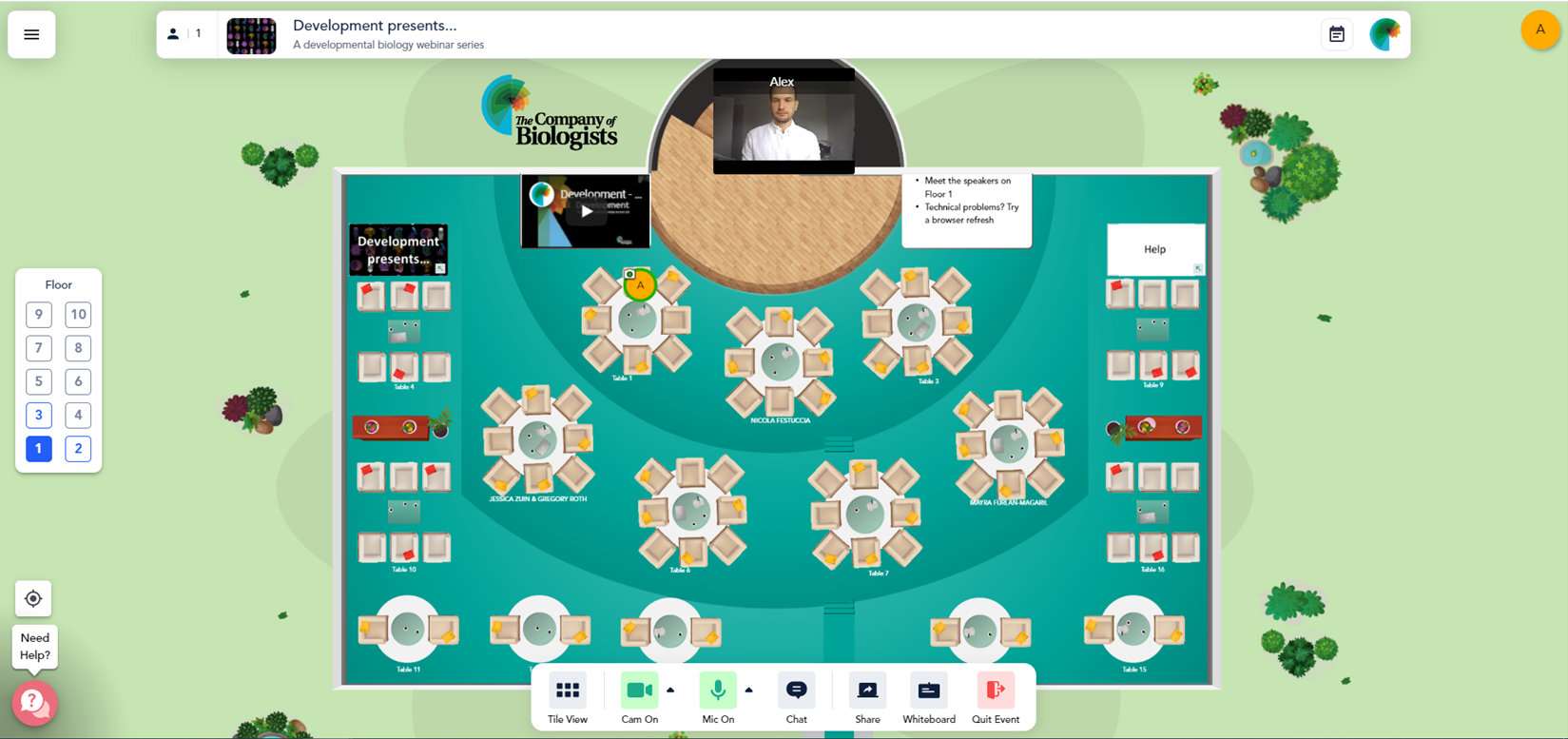
Provide a platform for informal discussions and networking in small, flexible groups. Finally, with the lack of in-person meetings at the time, we wanted to recapitulate some of the ‘chance’ in-person meetings and conversations that occur at seminars and conferences. To achieve this, all participants are invited to join an informal ‘discussion session’. We use the virtual conference platform Remo to host the webinars because, in addition to ‘meeting-wide’ presentations (similar to Zoom), Remo allows dynamic virtual interactions in small groups by allowing participants to quickly move between ‘tables’ (Fig. 2).
Fig. 2. The Remo virtual conference centre for discussion sessions. While seated at one of the virtual tables an attendees speaker and microphone are only shared with others sitting at the same table. It is also quick and easy to move between tables by double-clicking on where you want to move to and you can hover over other participants’ avatars to see who is sitting where.
To keep the conversation going, we also use a dedicated Twitter hashtag (#DevPres) to allow discussion beyond the webinar platform and we deposit the recorded talks on the Node after the webinar so that people unable to attend the live session do not miss out. We host the talks on our YouTube channel and link these videos to the speakers’ preprint manuscripts on bioRxiv, if desired. Occasionally, we also share some talks on our WeChat channel to reach a wider China-based audience where YouTube can be inaccessible.
Review
Since our first webinar in October 2020, we’ve hosted 42 speakers over 14 individual sessions. Most of the webinars have been chaired by Development’s editors with the occasional ‘special’ webinar hosted by guest chairs, such as those celebrating the anniversary of preLights and Development’s zebrafish issue, as well as Development’s special issues. As we now reflect on future directions for ‘Development presents…’ it’s a good opportunity to look at the stats so far.
Speakers
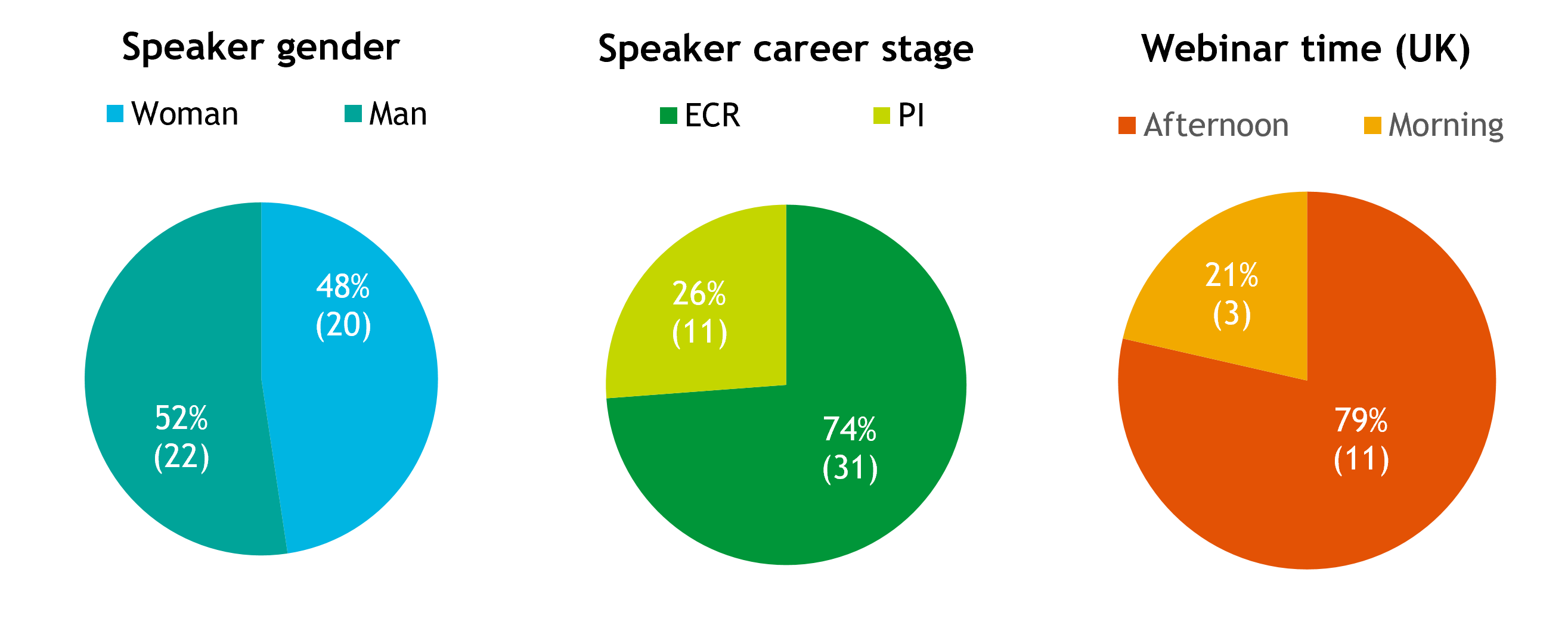
In terms of speakers, we have had a relatively equal split between men and women and almost three-quarters (74%) of the speakers have been first authors, aligning with our aim to promote ECRs (Fig. 3).
Fig. 3. Webinar demographics. Pie charts showing the percentage of the 42 speakers that have been women (48%) or men (52%) and ECRs (74%) or PIs (11). Percentage of the 14 webinars scheduled for the morning UK time (i.e. Europe-Asia; 21%) or afternoon UK time (i.e. Europe-Americas; 29%).
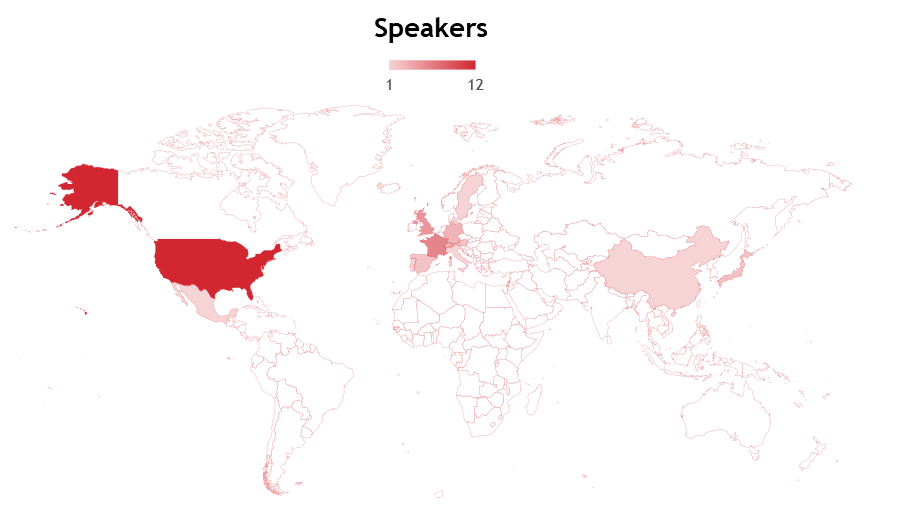
As a UK-based journal, we organise the webinars to begin in the morning (09:00-10:00) or afternoon (i.e. 13:00-19:00) UK time (GMT/BST) so that audiences in Europe-Asia or Europe-Americas, respectively, can attend live. We are also careful to make sure that all the invited speakers (as well as the chair) are in the same or a complementary time zone. The majority (79%) of the webinars have taken place in the afternoon UK time (Europe-America accessible; Fig. 3), which partly reflects the audience from our initial survey (>90% of respondents were in Europe or the Americas; Fig. 1). The majority of speakers have been from Europe-based institutions (this is perhaps unsurprising because European speakers could attend both morning and afternoon sessions). The USA has made up the most number of speakers from a single country (Fig. 4). Moving forward, we could certainly think about how to improve the geographical diversity of our speakers to better reflect the community and our attendees.
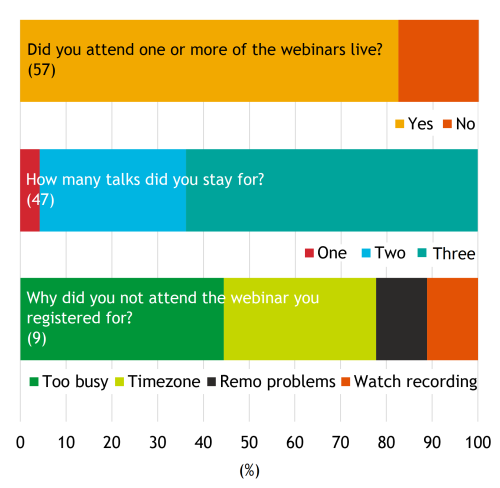
Usually, around 100 people or more (mean 137±50) register for the webinar but this has declined towards the end of 2021. Registration was initially lower for the morning-based webinars, although there hasn’t been much difference between morning vs afternoon registration and attendance in recent months. Overall, almost two-thirds of registrants have signed in to watch the talks live (mean 61%; range 44-98%) and the results from our 2021 survey show that the majority (>60%) of attendees stay to watch all three talks (Fig. 5). Webinar scheduling and timing seemed to be the biggest factor for those that registered but did not attend live (Fig. 5) but the number of people in this category (9) is low. On average, 20% of attendees stay logged on after the talks, presumably to participate in the discussion. While this represents only a relatively small number of people, we hope that those who did stay found the opportunity to talk to the speakers and network more generally to be useful.
Fig. 5. Results from Development presents… most recent survey. Questions regarding attendance: 1) Did you attend one or more of the webinars live? Yes (47/57); No (10/57). 2) How many talks did you stay for? One (1/47); Two (16/47); Three (30/47). 3) Why did you not attend the webinar you registered for? Too busy (4/9); Timezone (3/9); Remo problems (1/9); Decided to watch the recorder (1/9).
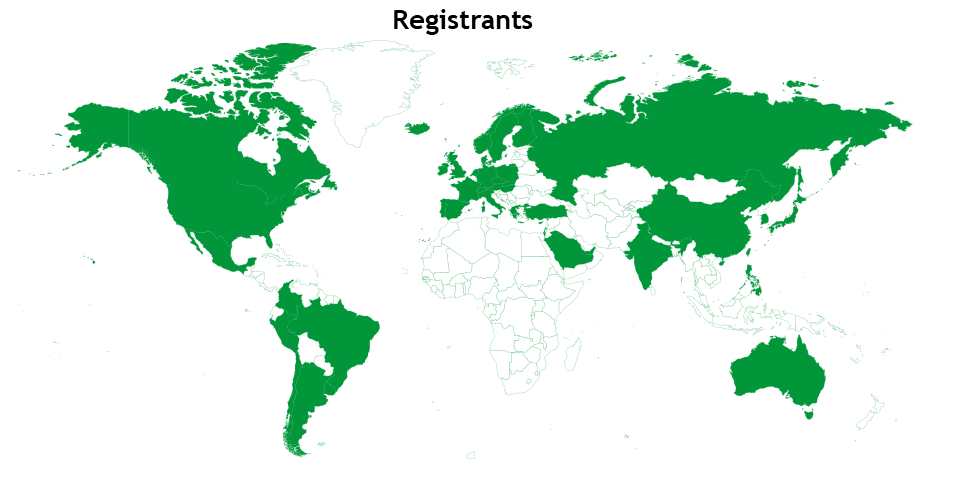
Although we do not collect data from registrants, an email address is required to sign up for the webinars via Remo. When a country-level domain is present in the email, we can estimate where some of the ‘Development presents…’ audience is attending from (Fig. 6). Encouragingly, we have a broad representation of registrants from around the globe, which suggests that the webinars are able to reach many countries and audiences.
What is next for ‘Development presents…’? First, we would like to understand the current thoughts and requirements for these types of virtual sessions so we can best adapt as the needs of our audience change. With this in mind, we’d like to invite you to have your say by completing the survey by clicking the link below. It shouldn’t take more than 6 minutes to complete!
A postdoctoral research fellow position is available in the Stathopoulos Lab at Caltech in sunny Los Angeles County to study how cell migration contributes to organ formation during Drosophila embryogenesis. In particular, we are interested in understanding recently identified signaling checkpoints that ensure that cells migrating to wrong positions are eliminated through a specialized programmed cell death “Anoikis“. Our aim is to build this Drosophila cell migration model to identify the molecular mechanisms that control metastasis. This project will build on our recent progress in this area (Macabenta, Sun, & Stathopoulos Developmental Cell 2022) by using cutting-edge live imaging, single cell sequencing, and molecular approaches to analyze both homo- and heterotypic cell-cell interactions in vivo. The successful applicant will join Angelike Stathopoulos’s lab at Caltech to work on this NIH funded project. For further details of research within the Stathopoulos lab, see: http://stathlab.caltech.edu. You should hold or be working towards a PhD in a biological science, be highly motivated and have an interest in studying cell-cell interactions. The position is open immediately and available for 3-5 years, with the possibility of extension based on good progress.
Candidates should send CV, include list of publications, a brief statement of research interests, and names and contact details for threes references to angelike@caltech.edu.
We are an equal opportunity employer and all qualified applicants will receive consideration for employment without regard to age, race, color, religion, sex, sexual orientation, gender identity, or national origin, disability status, protected veteran status, or any other characteristic protected by law.
Fin ray branching is defined by TRAP+ osteolytic tubules João Cardeira-da-Silva, Anabela Bensimon-Brito, Marco Tarasco, Ana S. Brandão, Joana Rosa, Paulo J. Almeida, António Jacinto, M. Leonor Cancela, Paulo J. Gavaia, Didier Y. R. Stainier, Vincent Laizé
A quantitative biophysical principle to explain the 3D cellular connectivity in curved epithelia Pedro Gómez-Gálvez, Pablo Vicente-Munuera, Samira Anbari, Antonio Tagua, Carmen Gordillo-Vázquez, Jesús A. Andrés-San Román, Daniel Franco-Barranco, Ana M. Palacios, Antonio Velasco, Carlos Capitán-Agudo, Clara Grima, Valentina Annese, Ignacio Arganda-Carreras, Rafael Robles, Alberto Márquez, Javier Buceta, Luis M. Escudero
Zygotic genome activation by the totipotency pioneer factor Nr5a2 Johanna Gassler, Wataru Kobayashi, Imre Gáspár, Siwat Ruangroengkulrith, Maximilian Kümmecke, Pavel Kravchenko, Maciej Zaczek, Antoine Vallot, Laura Gomez Hernandez, Laura Cuenca Rico, Sabrina Ladstätter, Kikuë Tachibana
PKN2 deficiency leads both to prenatal ‘congenital’ cardiomyopathy and defective angiotensin II stress responses Jacqueline J T Marshall, Joshua J Cull, Hajed O Alharbi, May Zaw Thin, Susanna TE Cooper, Christopher Barrington, Hannah Vanyai, Thomas Snoeks, Bernard Siow, Alejandro Suáarez-Bonnet, Eleanor Herbert, Daniel J Stuckey, Angus Cameron, Fabrice Prin, Andrew C. Cook, Simon L Priestnall, Sonia Chotani, Owen J L Rackham, Daniel N Meijles, Tim Mohun, Angela Clerk, Peter J Parker
The autism-associated gene SYNGAP1 regulates human cortical neurogenesis Marcella Birtele, Ashley Del Dosso, Tiantian Xu, Tuan Nguyen, Brent Wilkinson, Jean-Paul Urenda, Gavin Knight, Roger Moore, Ritin Sharma, Patrick Pirrotte, Randolph S. Ashton, Eric J. Huang, Marcelo P. Coba, Giorgia Quadrato
Machine-learning dissection of Human Accelerated Regions in primate neurodevelopment Sean Whalen, Fumitaka Inoue, Hane Ryu, Tyler Fair, Eirene Markenscoff-Papadimitriou, Kathleen Keough, Martin Kircher, Beth Martin, Beatriz Alvarado, Orry Elor, Dianne Laboy Cintron, Alex Williams, Md. Abul Hassan Samee, Sean Thomas, Robert Krencik, Erik M. Ullian, Arnold Kriegstein, Jay Shendure, Alex A. Pollen, Nadav Ahituv, Katherine S. Pollard
Morphodynamic Atlas for Drosophila Development Noah P Mitchell, Matthew F Lefebvre, Vishank Jain-Sharma, Nikolas Claussen, Marion K Raich, Hannah J Gustafson, Andreas R Bausch, Sebastian J Streichan
Reconstruction and deconstruction of human somitogenesis in vitro Yuchuan Miao, Yannis Djeffal, Alessandro De Simone, Kongju Zhu, Andrew Silberfeld, Jong Gwan Lee, Jyoti Rao, Oscar A. Tarazona, Alessandro Mongera, Pietro Rigoni, Margarete Diaz-Cuadros, Laura Min Sook Song, Stefano Di Talia, Olivier Pourquié
Parallel clonal and molecular profiling of hematopoietic stem cells using RNA barcoding Edyta E. Wojtowicz, Jayna Mistry, Vladimir Uzun, Anita Scoones, Desmond W. Chin, Laura Kettyle, Francesca Grasso, Allegra M. Lord, Graham Etherington, Charlotte Hellmich, Petter S. Woll, Mirjam E. Belderbos, Kristian M. Bowles, Claus Nerlov, Wilfried Haerty, Leonid V. Bystrykh, Sten Eirik W. Jacobsen, Stuart A. Rushworth, Iain C. Macaulay
The Network Zoo: a multilingual package for the inference and analysis of biological networks Marouen Ben Guebila, Tian Wang, Camila M. Lopes-Ramos, Viola Fanfani, Deborah Weighill, Rebekka Burkholz, Daniel Schlauch, Joseph N. Paulson, Michael Altenbuchinger, Abhijeet Sonanwane, James Lim, Genis Calderer, David van Ijzendoorn, Daniel Morgan, Alessandro Marin, Cho-Yi Chen, Alex Song, Kate Shutta, Dawn DeMeo, Megha Padi, John Platig, Marieke L. Kuijjer, Kimberly Glass, John Quackenbush
The Beddington Medal is awarded by the BSDB for the best PhD thesis in developmental biology, defended in the year prior to the award. The medal is named in memory of Rosa Beddington, who made major contributions to both the field of developmental biology and the BSDB. The artwork on the medal is from Rosa’s own drawings.
In 2022, the medal was awarded to Guillermo Serrano Nájera. Guillermo completed his PhD in Cornelis Weijer’s lab in Dundee and he presented his work at the recent BSCB/BSDB annual meeting in Warwick. I caught up with Guillermo over Teams to find out more about his career so far.
Guillermo receiving his medal from Val Wilson
Where are you originally from?
I’m from Spain, from Madrid. I studied biochemistry at the university there. After completing my undergraduate degree, I decided I wanted to do something more interdisciplinary, so I stayed on to do a master’s degree in biophysics. At this point, I moved to Newcastle in the UK to do a second master’s degree in computational biology, but my main focus for this move was to improve my English. I could read very well but wanted to improve my speaking skills to be able to communicate more easily in science.
When did you first become interested in science?
I’ve been very interested in science for a long time, since childhood really. But I was also interested in just about everything! I liked archaeology, anthropology, literature, even philosophy. Then little by little, I became more convinced about focussing on science. Before university, I was choosing between physics and biology. I like the methods of physics, but ultimately decided to go along the biology route because I’m more interested in asking questions about living things. That’s why ultimately, I wanted to study science, and biology in particular.
How did you come to do a PhD in Cornelis (Kees) Weijer’s lab?
I was completely convinced that I wanted to study developmental biology for my PhD. I had wanted to study developmental biology for a long time, but for one reason or another, I couldn’t join a suitable lab. The second factor in choosing my thesis lab was that I wanted to do something interdisciplinary, using computing, modelling, as well as biology. Of course, there are many labs that are interdisciplinary, but I found that most of them were looking for biologists and physicists to collaborate in the lab, rather than truly interdisciplinary researchers. But I told Kees, my supervisor, that I wanted to work on all aspects of our research. He told me, ‘of course, there’s no other way!’ So, we were a match, and ended up working together.
Tell us about your PhD project: what were the main questions you were trying to answer?
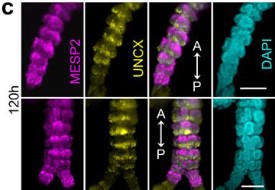
The main question that I wanted to answer was, how are cell behaviours coordinated during animal development to generate shapes. We use chick embryo gastrulation as our model, and in this continuous epithelial layer, the cells can only have four different behaviours. This makes things easy, right! The cells can rearrange or interchange neighbours, they can change shape, they can divide, or they can ingress and abandon the layer. All these four cell behaviours coordinate in space and time over thousands and thousands of cells to generate the primitive streak during avian gastrulation. My question was, how do all these cells know what to do in each moment during this process. I used chemical perturbations to disrupt different cell behaviours as this was an easy method to combine with imaging. And the surprise was that by disrupting these behaviours, we generated structures that are very similar to gastrulation in other organisms. After these initial observations, we began to focus on our interpretation of these results in our next experiments. But the original question was, how are cell behaviours coordinated during development.
Figure from Chuai and Serrano Nájera, et al. (2021)
You have two bioRxiv preprints arising from this work, where you use both experimental data and modelling to address the differences in gastrulation in vertebrates, can you tell us about this collaboration and your results?
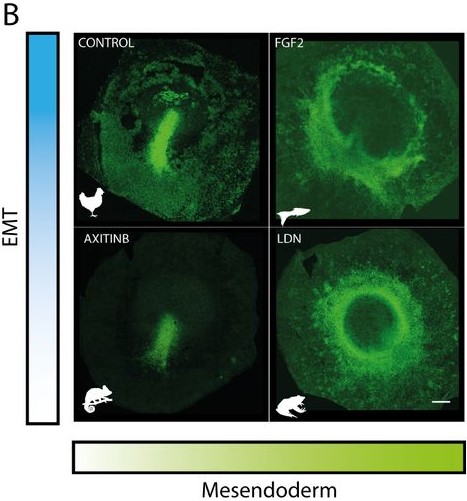
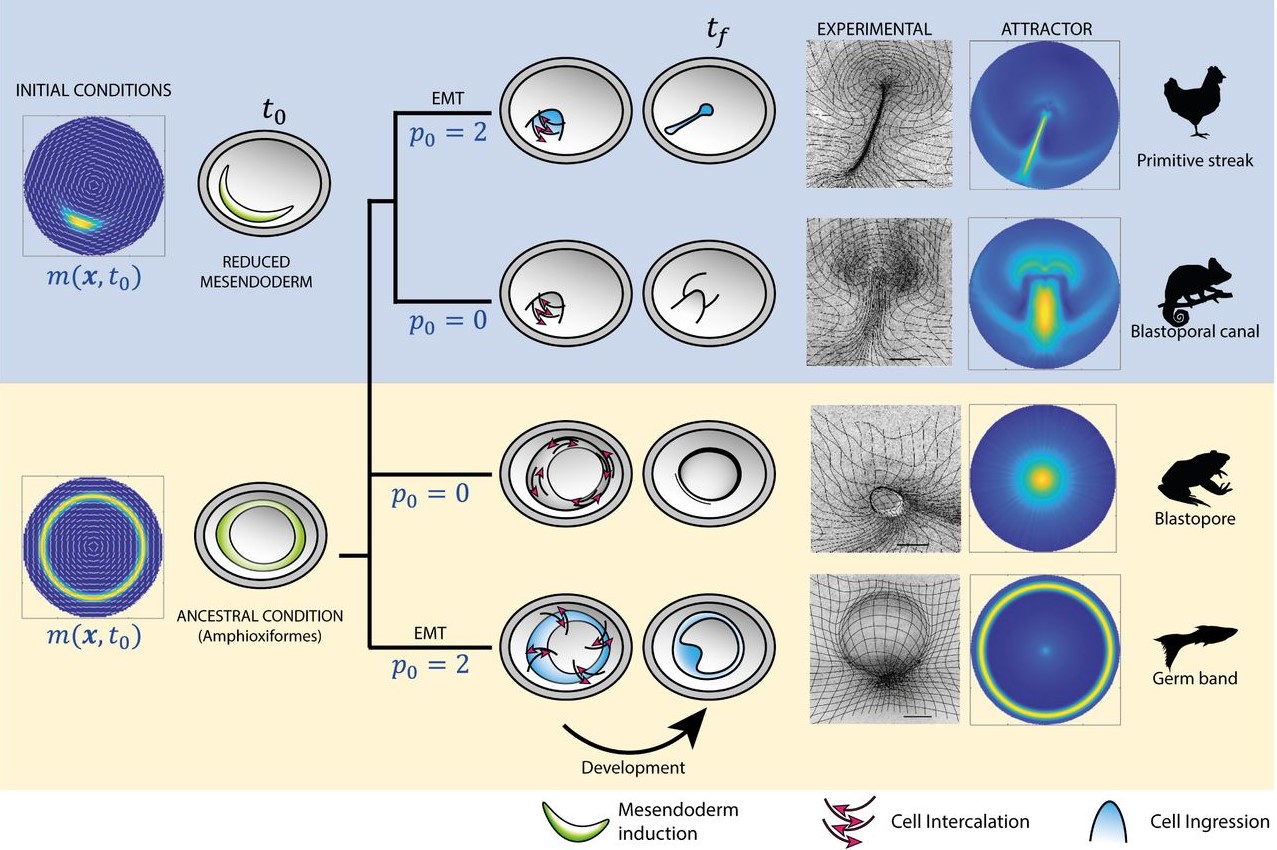
The biology paper is mostly my own work from my PhD, where we used chemical perturbations to change two parameters during gastrulation in the chick embryo; one was the amount of mesoderm, and the other was whether the cells in the mesoderm territory can undergo cell ingress. We found that by changing just these two parameters, you can generate different morphologies in the embryo, which look very similar to gastrulation in other organisms. At the same time, we observed myosin cables in the mesoderm territories, and we think these cables are producing the force to move the cells. For a long time, Kees had thought that these cables could be mechanosensitive, and that tension helps with their assembly. This could explain the movement of the cells and the self-organisation of the tissue flows. He was looking for collaborators to try to explore this in a more formal way using mathematical models. Kees was meeting with Mattia Serra from the Mahadevan lab about another project, and he asked if they would be interested in collaborating on this research. They began working on the model and how to implement it. I joined them because I had a way to test the model experimentally. We introduced the same perturbations into the model and were able to reproduce the experimental results. So, it was a confirmation that this principle of myosin self-organisation, and the resulting tissue flows could be underlying the structures that are formed during gastrulation.
Figure from Serra, Serrano Nájera, et al. (2021)
It must have been exciting when you saw that you could phenocopy the different types of gastrulation from other organisms.
In the beginning, we were not thinking about that at all. Actually, the first treatment that I performed resulted in an embryo that looked similar to the gastrulation in reptiles, with a small invagination in front of the primitive streak. We had that result for a long time before we thought about it from that perspective. Then, after reading papers about gastrulation in chameleons, I told Kees that I thought that our result looked similar to gastrulation in reptiles. We knew about the paper from Alev Cantas, Guojun Sheng and colleagues, where they create a circular primitive streak in chick embryos using FGF. They compare this to the circular primitive streak in reptiles, and the evo-devo perspective that they had in their paper was super inspiring for me. They called this circular primitive streak a blastopore, and in the beginning that is what we were calling it. But later, one of our treatments caused a complete invagination of the epiblast and we realised that our first result was not the blastopore! It was very, very exciting, especially when this last invagination happened and we realised that we could phenocopy all the gastrulation modes, and we began to interpret all our results in a new light.
If you took one abiding memory with you from your PhD, what would it be?
I had a lot of fun during my PhD, so choosing one moment is hard! I think I will have to choose an everyday moment that I always enjoyed. So, I have seen the movement of cells and the formation of the primitive streak thousands and thousands of times, but each time, after doing my experiments, I loved looking at the movies! I find the process of gastrulation very mysterious and beautiful, and that hasn’t changed since my first experiments. And I think that kept me going a lot – just to feel that mystery.
You been involved in a number of different projects including publishing a paper on ‘Trendygene’, a pipeline for detecting targets for drug discovery, how did you come to work on this project?
In general, I fall in love with everything, that’s my problem! At the beginning of my PhD, I was very interested in artificial intelligence. It’s a fascinating topic, machine learning; I find it amazing. In my PhD programme, the BBSRC EASTBIO Doctoral Training Partnership, you have to complete an internship. I decided that I wanted to do an internship where I could learn some applications of artificial intelligence. I had already started studying by myself with a few online courses. And for my internship, I went to a pharmaceutical company called Exscientia. They wanted to be able to detect emerging areas in the biomedical literature to focus their efforts in producing new drugs. I came up with this idea, which we called Trendygene. I can briefly explain the background: we trained a deep neural network, to predict the number of publications about each gene in the human genome in a particular year, for example in 2020, but only using the data from 1980 to 2013. As well as publications, we incorporated data from recent reviews, citations, clinical trials, everything. And it works very well! But we were more interested in looking at when it fails, because in some cases, it fails very badly. These were unusual, because, in general, most of the trajectories of publications are very, very predictable – that’s why the machine learning approach works, but it makes huge mistakes for a small subset of genes. The interpretation for these genes is that something happened in the last few years, which are not included in the time series, that radically change the publication dynamics in unexpected ways. I think that our work is original because instead of using artificial intelligence to predict something, we’re interested in when it fails. The cases when it fails tend to be due to a recent discovery. We selected a couple of genes to talk about in the paper, and to say why we think they are trendy. One of them is PIEZO1, the mechanosensor which was recognised in the Nobel Prize in Physiology or Medicine awarded in 2021; we could detect it in July, which was very nice.
Did the network ever fail in the opposite direction?
We did find that we could have anti-trendy genes. In general, the network fails more often in the other direction; the anti-trendy genes tend to have smaller errors. In the first versions of the manuscript, we included some analysis of why some things are anti-trendy, but they were more difficult to analyse. Some examples of anti-trendy genes were genes that have become obsolete in diagnostics due to a new alternative. So, we could detect these genes, but they were not as interesting so we decided just to focus on what is trendy!
You also had a poster at the BSCB/BSDB meeting on self-domestication in naked mole rats – how did you start working on this project?
So, just before the pandemic, I became obsessed with domestication. I think it is a very interesting problem. Domestication is probably one of the longest running biology experiments that we have ever done! Which means we have all these results out there just ready for us to look at. One of the first scientists to look was, of course, Darwin. He wrote a massive book on the topic called ‘The Variation of Animals and Plants Under Domestication’. I went to the library in Dundee to try to borrow the book, and they told me that it would be a couple of days before I could collect it. When I arrived, I found that it was the first edition of the book! It had been in the basement of the library for 97 years before I got it. It is a beautiful book with original binding, it is incredible! I spent the lockdown with that book in my home library because the main library was closed.
So, you were allowed to take the book out of the library?!
Yes, I was. I actually wrote to them because, although the book was in the catalogue, I wasn’t sure that they realised what they had. They are now hoping to showcase the book at the library in the future. Anyway, as I said I became obsessed with domestication. One of the things that was most interesting to me is something called domestication syndrome, which is a series of traits, both morphological and behavioural that are characteristic of domestic animals but are not very common in wild species. The most famous example is drooping ears; only elephants have drooping ears in the wild, and they use them to control their body temperature. But all the other animals that you can think of with drooping ears are domesticated animals, dogs, pigs, cows. So, why do domesticated animals have drooping ears and while wild animals do not? Another famous example attributed to domestication syndrome are the white patches on the skin, like you see on cows. This kind of coloration is not present in the wild or is at least very uncommon. There are many other characteristic traits, for example a small snout and a reduction in the number and size of teeth. Interestingly, all these tissues, the teeth, the snout, the melanocytes responsible for the pigmentation, are derived from neural crest cells. The neural crest can generate very different tissues during development including the peripheral nervous system and the adrenal glands. That is really important because domestication syndrome theory says that domesticated species have a reduced fear response, they don’t have a fight or flight response. This is why you can pet a cow and it will not attacked you or run away. So, if you have problems in your neural crest cells, and they cannot migrate well or proliferate well or differentiate well, then they you will have a reduced peripheral nervous system, which transmits the fear response from the brain to the body and you will have lower adrenaline levels because of the effect on adrenal gland and so on. And as a side effect, these defective in the behaviour of the neural crest cells, will generate problems in other tissues such as the cartilage of your ear, or with your teeth etc. It’s just a theory, but I think is really beautiful. It fits with many animals, although it’s probably not the only way to domesticate. And then another one of these domestication syndrome traits is to be fully naked. For example, there are naked dogs, naked cats, naked horses (which looks extremely bizarre), pigs, and of course the naked mole rat. I was really interested in the naked mole rat because they are eusocial animals, they they behave like bees, with a queen and then other workers. There are rat miners, and rat nurses, I’m oversimplifying of course! But basically, they have this kind of social organisation. So, it occurred to me that, and it has also proposed in the literature, that animals could self-domesticate to live in groups, to increase their social tolerance. This led me to think that maybe the naked mole rat is a self-domesticated animal that self-domesticated to improve its social tolerance and that’s how they managed to evolve this eusocial behaviour. They live in very large groups of up to 300 individuals, which is unusual.
I began to look at this project with a friend, Koryu Kin, during the pandemic. We met in the street as I was wheeling my lab chair down the street to my home and Koryu was on the way to the lab to collect his things at the beginning of the lockdown. We had discussed the naked mole rat project before and when me met, Koryu told me, maybe this is the moment to work on it. He was right! We are now trying to publish our paper, where we defend our hypothesis that the naked mole rat presents self-domestication syndrome traits, and that this could improve the social tolerance and be related to the evolution of eusociality in these animals. We also present a hypothesis that the eusocial behaviour is also linked with the properties of the soil.
Sticking with side-projects that you have done during your PhD, you were selected for the Roche Continents programme which brings together artists and scientists, can you tell us more about how you became involved in the programme?
During my PhD, I have been involved in mixing science with art. For example, I am very interested in computational art. In the past, I created a couple of bots online, for example Twitter bots, which detect haiku, Japanese poems with a very particular set of syllables. The bot I created detects when something written in Spanish can be cut in such a way to reproduce a haiku. Basically, the bot retweets you to say ‘hey, you wrote a haiku, though you probably didn’t know it!’ While in Dundee, I’ve also been a tutor for a programme where students in art are encouraged to link up with scientists, to find inspiration from science, and to work on a project together. Because of my interest in art and science, I decided to apply to Roche Continents, which is a programme that Roche organises where they bring together art students and science students for one week every summer in Salzburg. We did workshops together, and we also went to the opera and the theatre. It was very, very interesting and a great experience. I would recommend it to everybody!
What are you working on now, as a postdoc with Ben Steventon in Cambridge, and how does Cambridge compare with Dundee?
I only moved here two months ago so I have not really begun working on my project yet. But I was attracted to Ben’s lab because of the different model organisms they are working on. They work on chick, fish, and mouse gastruloids. I’m exploring all of these models with different people in the lab and collaborating on different projects. However, my intention is probably to move more towards the mouse gastruloids. And in terms of Cambridge, when I first moved here, I found it very busy, noisy and with cars everywhere! I was so use to Dundee, which is a quiet city with a fantastic river. I thought that it was a very good place to be during lockdown because nature is very close. I really enjoyed my life there. But now here in Cambridge, I really like the massive community of scientists associated with the university, there are physicists, engineers, biologists; you have world experts in every aspect of science. That is fantastic.
Longer term, do you know if you plan to stay in science?
I don’t really have long term plans. For the moment, I would like to continue in science, and in academia in particular. I think that the kind of questions I want to answer are a better fit for academic science. That’s at least my medium-term plan!
Where do you think developmental biology will be in ten years?
I would like to see us have a very good understanding of developmental principles so that we can begin to manipulate them. Now, for example, we have these in vitro systems like gastruloids and other types of organoids; we understand the some of the developmental principles, and we can guide the development, but in the future, we could begin to engineer development. We could manipulate these programmes to generate things that are not in nature, or to find new purposes for the in vitro models. I think this engineering will happen, but I don’t know if it will be within 10 years or later.
When you’re not in the lab, what do you do for fun?
As I mentioned, I like computational art, I do that a lot. I also like walking in the forest, cooking with my girlfriend and spending time with friends. I think I have a very normal life outside of the lab!
Doing great science depends on teamwork, whether this is within the lab or in collaboration with other labs. However, sometimes the resources that support our work can be overlooked. Our ‘Featured resource’ series aims to shine a light on these unsung heroes of the science world. In our latest article, we hear from Steve Lisgo and Janet Kerwin, who describe the work of the Human Developmental Biology Resource (HDBR) Atlas.
When was the HDBR Atlas established and what are its aims?
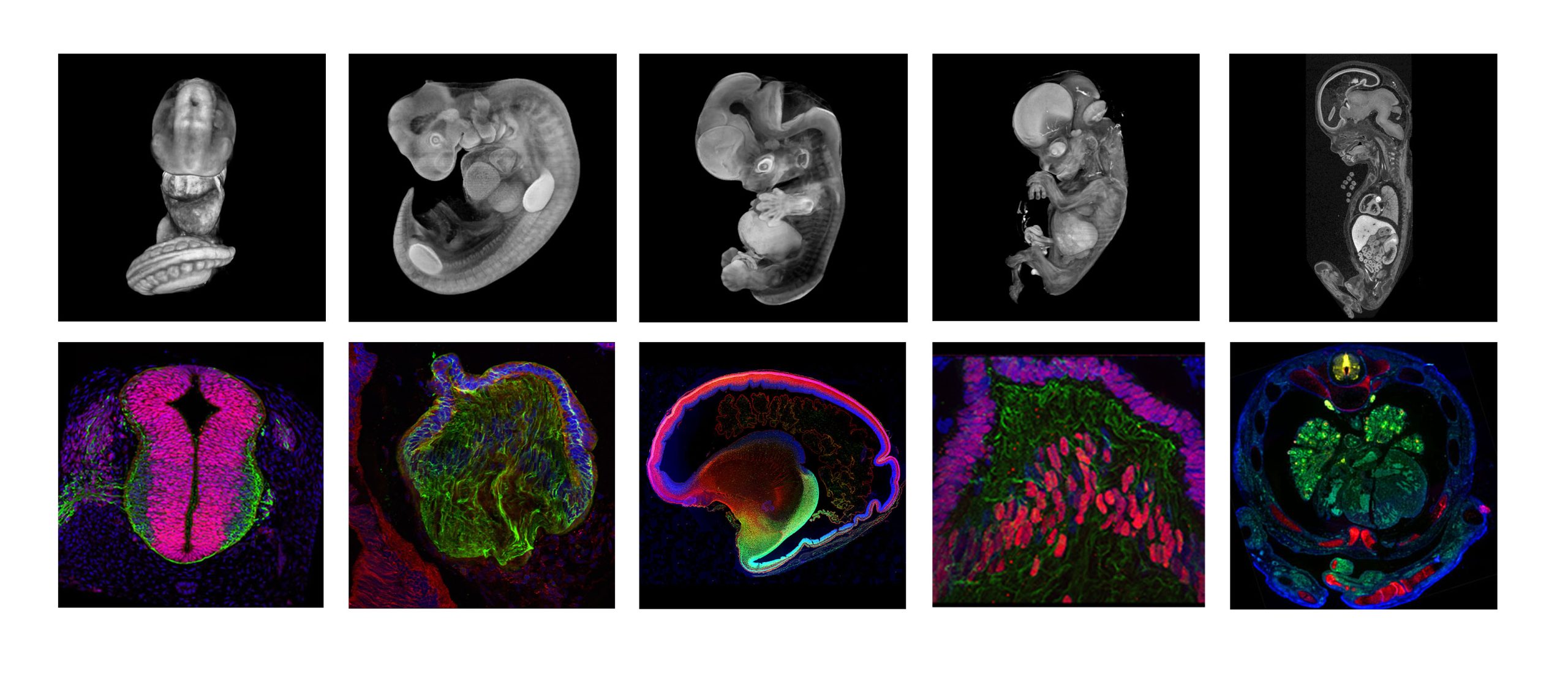
The HDBR Atlas was established in 2020 and was developed from the Human Developmental Studies Network (HuDSeN) and Electronic Atlas of the Developing Human Brain (EADHB) projects, which began in 2002. The HDBR Atlas brings together 3D models, high resolution histology sections and gene expression data, and aims to facilitate the learning and understanding of human embryonic and fetal development.
Images courtesy of the HDBR Atlas
Who runs the HDBR Atlas?
The Human Developmental Biology Resource (HDBR; www.hdbr.org) is an MRC/Wellcome funded human embryonic and fetal tissue bank, managed across two centres: Newcastle University and UCL. The samples collected by the resource are used in registered research projects in the UK and overseas. Data generated both by HDBR users and produced in house at London and Newcastle HDBR centres are disseminated through the HDBR Atlas. The Newcastle HDBR group lead on curating and developing the website.
What can researchers find in the HDBR Atlas?
The HDBR Atlas is a freely available website comprising four sections: 1) 3D models of normal embryos (Carnegie stages (CS)12 to CS23) and fetuses (8-11-post conception weeks), plus a series of karyotypically abnormal embryos. 2) Annotated histology sections of whole CS12-23 embryos at low and high resolution. 3) Organ systems, including gastrointestinal, nervous, urogenital and visual systems, and with a new feature on the developing cardiovascular system. The latter includes movies and interactive 3D models (CS12-16), annotated H&E sections (CS14-22), and fetal heart dissections (8-17 post conception weeks (pcw)). 4) Gene expression data for 129 genes from CS11 to 19 pcw. All the HDBR Atlas resources are available to download on request.
How can the community contribute?
We encourage users of HDBR material to return data images (or physical slides, if this is easier) so that the data can be made publicly available via the HDBR Atlas. Anyone who has human embryonic or fetal protein or gene expression data generated from microscope slides can submit data to us using the submission form available on our website (https://hdbratlas.org/data-submission.html). Expertise in different organ systems is always welcome to define anatomy in the 3D models and annotate 2D histology sections.
Any hidden gems, features that are new, or that researchers might be less aware of?
Previously, we have shown 2D images and movies of 3D models on the Atlas. Recently, we have added interactive 3D reconstructions of early heart development (https://hdbratlas.org/interactive/heart.html) which can be manipulated and zoomed in/out by the user.
If money was no object, what would you like to add to the HDBR Atlas?
Generate 3D models for all stages of development from 3 to 20pcw; create more detailed/comprehensive labelling of the histology sections; further develop the high resolution eHistology viewer to include transverse, sagittal and frontal section planes at each Carnegie Stage, and expand this into fetal stages; produce a fully searchable gene expression database with sets of marker genes for key structures e.g. cardiac markers and distinct brain regions; and create a section on comparisons between human and mouse development for different organ systems.
Feedback from users of the Atlas
‘Many thanks for a wonderful resource!’
‘I would like to take this opportunity to reiterate how useful the HDBR is. It is an amazing resource, unequal in its ability to support developmental biology research. My sincere gratitude to all the dedicated staff involved.’
‘I have used the 3D images in lectures (thanks!) and would really like to see these accompany any further additions/stages. 2D histology sections are also extremely useful to help lab members understand what they should be looking for in our own experiments.’
‘This is a great project! We linked to certain images for our graduate embryology workshops last semester. The students really liked it!’
‘Again, thank you for putting together this incredible resource.’
Thanks to everyone who has contributed to our ‘Featured resource’ series so far. You can read the whole series here. Let us know which resource(s) is essential for your research and we’ll add them to our list. Get in touch at thenode@biologists.com.
For our latest SciArt profile, we continue our adventure into 3D artwork and showcase the the neuro-woodwork sculptures from Louis-Jan Pilaz. LJ is an Assistant Professor at the University of South Dakota, Sanford School of Medicine, where his lab works on early cortical development.
Where are you originally from and what do you work on now?
I was born in France. This is where I got my PhD, related to neural stem cell biology. During my master’s, I worked in Colette Dehay’s lab and I completely fell in love with radial glia. I think they are the most amazing cells in the universe, so full of mysteries. Flash-forward 13 years, after a couple postdocs in Nick Gaiano’s lab and Debby Silver’s, I am now working on them in my own lab. We are interested in many aspects of early cortical development and our daily bread is working with microscopes.
A 2-foot radial glial cell sculpted in ashwood on a walnut podium.
Were you always going to be a scientist?
Science has always been a huge part of my life. Going to college, I wanted to be an ethologist. Then I ran across a pamphlet on cognitive sciences and majored in that. Afterwards, I wanted to go back to heavy-duty biology, so I started working in the Dehay lab and so on.
Microglia cut out in a walnut slab.
And what about art – have you always enjoyed it?
While I was born in France, my mom is Dutch. This meant that every summer we would travel to the Netherlands, where we would spend a lot of time in museums. I am a huge fan of Vermeer and Rembrandt. I can also get really moved by contemporary art. Also, my grandfather was a painter and my great-grandfather was a carpenter. Those sets of genes probably gave me this irresistible need to tinker, to try artsy things.
Oligodendrocyte sculpted in sapele wood.
What or who are your most important artistic influences?
I don’t feel I am consciously influenced by any artist in particular. All the pieces of art I was exposed to as a kid are probably doing their job subconsciously. There are so many talented neuroscientists turned artists! I love the works of Greg Dunn, Needles and neurons, Laura Bundesen, Maria Zanfir…
This human brain was cut out of a slab of walnut.
How do you make your art?
I have to admit that I use a lot of power tools. And I keep learning how to use new ones to shape the wood the way I want to. The most critical tool in my workshop is the scroll saw, it allows me to give the wood the intricate shape of a neural cell, and then I have a whole arsenal for the final touches. Importantly, most of my creations start on my computer.
Epoxy-filled astrocytes in oak.
Does your art influence your science at all, or are they separate worlds?
They are quite separate for now, but I get to interact with a lot of amazing scientists who are interested in my work, that I may not have met if it weren’t to talk about my creations. While I love creating cells in wood, my priorities are my lab and my family.
Astrocyte carved in walnut wood. This piece is now in Australia.
What are you thinking of working on next?
I want to scale up. I want to make 6-foot neurons, astrocytes, microglia and of course radial glia. I also want to combine several cells together to create big installations.
Thanks to LJ and all the other SciArtists we have featured so far. You can find the full list here. We’re always on the lookout for new people to feature in this series – whatever kind of art you do, from sculpture to embroidery to music to drawing, if you want to share it with the community just email thenode@biologists.com (nominations are also welcome!)
We offer one fully funded postdoctoral position up to five years in the Laboratory of Genome Integrity located at the National Institutes of Health (NIH, Bethesda, MD). NIH is the largest biomedical research agency in the world, fosters world-renowned researchers and provides access to state-of-the art innovative technologies and scientific resources.
Our laboratory uses human and mouse embryonic stem cells (ESCs) as well as mouse embryos to understand the molecular mechanisms underlying cell fate decisions. The applicant should have or about to have a PhD in Developmental Biology, Genetics, Molecular Biology or similar, and must have demonstrated expertise on molecular biology/mammalian cell culture (preferably in embryonic stem cells). Knowledge in mouse embryology, single-cell RNAseq, chromatin architecture and/or next generation sequencing technologies will be considered as an advantage.
The applicant will be involved in a variety of exciting projects ranging from studying the relation between cell plasticity/totipotency and chromatin architecture to exploring the underlying mechanisms of new regulators of Zygotic Genome Activation. We seek a highly motivated, creative individual, eager to learn and develop new technologies and complex cell systems based on live cell/embryo imaging, single-cell technologies and CRISPR-based editing interested in understanding how a single cell can develop into a complex multicellular organism in vitro and in vivo.
Applications will be accepted until the position is filled.
Recent publications: • Vega-Sendino, et al (2021) The ETS Transcription Factor ERF controls the exit from the naïve pluripotent state. Sciences Advances, 7(40): eabg8306. • Olbrich, T., et al (2021) CTCF is a barrier for 2C-like reprogramming. Nature Communications 12, article number: 4856. • Markiewicz-Potoczny, M., et al (2020) TRF2-independent protection of telomeres in pluripotent stem cells. Nature, 589: 103-109.
Debate on the status of the field of developmental biology continue to rumble on, including an open letter to Claudio Stern from Peter Lawrence published on the Node. You can read the letter here, as well as a reply from Alfonso Martinez Arias. Remember, the Node is your site and if you would like to share your hope and fears for the future of developmental biology all you need to do is register and then you are free to post. Please get it contact if you need any support doing this, or if you would like to collaborate with others on a post.
If you would like to write for the Node, check out our recent list of writing ideas. If you would like to contribute to our ‘Developing news’ blog, please get in touch at thenode@biologists.com
At the end of 2021 and beginning of 2022, Journal of Cell Science (JCS) put out a call for essays on ‘Equity, diversity and inclusion in cell biology’. JCS introduced the series to amplify voices that are not always heard in this space. After receiving some fantastic essays on a range of different topics, they selected three winners and have now published these essays, as well as those from the runners-up.
If you have a story about equity, diversity and inclusion that you’d like to share, we’d love for you to post it on the Node. Get in contact (thenode@biologists.com) if you have any questions or would like help posting your article.
 (No Ratings Yet)
(No Ratings Yet)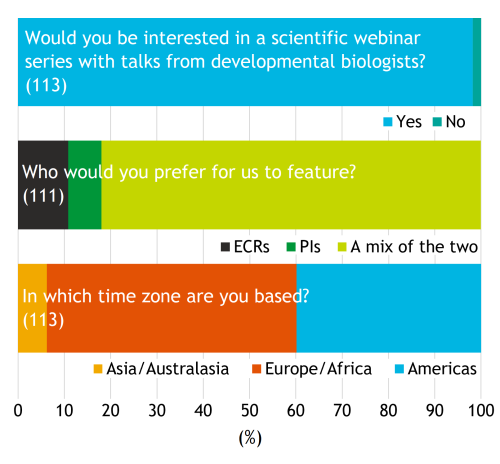





 (1 votes)
(1 votes)