Are you an early career researcher interested in the cell or molecular mechanisms underlying disease? Do you have an outstanding record and an innovative research plan?
The Sir William Dunn School of Pathology at the University of Oxford is looking for outstanding early career researchers seeking a stimulating and supportive environment in which to establish their research group as externally-funded fellows. We are specifically looking for researchers seeking mentoring and sponsorship to apply for career development fellowships (e.g. Wellcome Trust Career Development Award, MRC Career Development Award, CRUK Career Development Fellowship, UKRI Future Leaders, etc). Researchers who succeed in securing a fellowship will then be invited to establish their independent group in the department, benefiting from a generous support package, comprehensive mentorship, career development training and opportunities to recruit Oxford undergraduate and postgraduate students.
Successful candidates will have an outstanding track record in any area of biomedical research, with a particular focus on the fundamental cell and molecular biology underlying disease. The Department celebrates diversity and we welcome applicants from diverse backgrounds that are currently underrepresented at the University of Oxford.
The Dunn School is a dynamic and collaborative department, with 30 research groups and roughly 300 research staff investigating the biology underlying disease, using a wide range of basic and translational approaches. Our interests span many disciplines including cell and molecular biology, microbiology, development, immunology and cancer biology. Our researchers have access to excellent scientific facilities and support services, and the stimulating environment of the South Parks Road science area in particular, and the wider University of Oxford in general.
In the latest episode of Genetics Unzipped we’re taking a trip back in a virtual time machine, soaking in the primordial soup to discover the origins of DNA, find out where genes come from and how some species have stolen theirs from viruses, and explore what’s next for the genetic code.
Experts think that the first step towards life was simply a molecule that was capable of self-replicating. As a geneticist, your mind might jump straight away to the most famous self-replicating molecule of them all, DNA. As we discover, that’s probably the least likely scenario, but what actually happened is still a hot topic of debate among researchers searching for the origins of life.
Moving from DNA to genes, as far as we can tell, all of life on earth evolved from one common ancestor, LUCA, which must have had one set of genes, whatever they looked like. But that leaves the question of how this simple set of genes diversified to encompass the incredible diversity of genes that now exist in trillions of extant and extinct species on earth. We look at where genes come from, and how we’ve managed to steal some from our mortal enemies, viruses.
Finally, you may think you know your A, C, T and G when it comes to DNA, but what about B, P, S and Z? We discover how the genetic code is expanding, thanks to Hachimoji DNA.
If you enjoy the show, please do rate and review on Apple podcasts and help to spread the word on social media. And you can always send feedback and suggestions for future episodes and guests to podcast@geneticsunzipped.com Follow us on Twitter – @geneticsunzip
We built an anatomy ontology. You should too – here’s why.
We have an information scale problem. I’m hardly the first to note the exponentiality and rapidity of information growth, as it was a keenly felt sentiment even at the dawn of the Industrial Age: “Knowledge begets knowledge as money bears interest (Conan Doyle, 1885).” Today there are a staggering 32,948,436 papers in the PubMed database (Aug 17th, 20:07 PM CT). Consider this sobering perspective: if a single person wanted to read each and every one of these papers, and, optimistically(!), read 5 papers per day, it would take 6,589,687 days. Or, 18,053 years. Which is the average lifetime of 229 people. And that’s just the papers written about all of the data that was collected.
The information problem in the life sciences in particular is further compounded by the varied types of data in current publications: supplemental figures, spreadsheets, stand-alone databases. With technological advances and increased storage capacity, collecting big data is no longer the bottleneck. New information is cheap. From single cell sequencing to large scale volumetric microscopy imaging, we have more data than we can wrap our heads around. How then do we effectively mine data to generate testable hypotheses with the potential to transmogrify information into new knowledge? Part of the answer lies in creating unified analysis schemes across platforms that are both human and machine readable. One of the ways we are doing this is by using ontologies as frameworks for organizing data.
What is an ontology?
Ontology might be a new word to you, but more than likely you’ve already been using them. Ontologies organize and link data for social media sites and big retailers. Have you ever saved an item to a Pinterest board (Gonçalves et al., 2019)? Used a filter to shop for a specific color, brand, and size of clothing from an online retailer? Run a GO (Gene Ontology) enrichment on a differentially expressed gene set? Used FlyBase or WormBase to browse gene pages? If so, you’ve interacted with an ontology. And you are going to interact with more.
An ontology by definition (Oxford Languages) is:
(1) the branch of metaphysics dealing with the nature of being (not this one!)
(2) a set of concepts and categories in a subject area or domain that shows their properties and the relations between them. (this one!)
If you’re familiar with libraries and the Dewey decimal system, this will all start to sound very familiar. To explain, let’s jump into an example:
“On the Origin of Species” IS A book.
That statement is an ontological axiom. An ontological axiom is a simple sentence that follows a pattern: concept / relationship / concept. In our example, both “On the Origin of Species” and “book” are concepts; IS A is the relationship. Now, let’s take it one step further: the idea of the concept in your head likely has some specific attributes. In ontological terms, those specifics are known as properties.
A set of properties for “On the Origin of Species” could be: Author: Charles Darwin Publication Date: November 24, 1859 ISBNs: 9780521867092, 9780857088475, 9788423918164…
Now we have a concept with properties and the categorical relationships between them. But we don’t have to stop there! We can define other relationships that exist for “On the Origin of Species” and string them together, like this: “On the Origin of Species” IS A scientific non-fiction book; a scientific non-fiction book IS A non-fiction book; and a non-fiction book IS A book. Here’s the super power of ontologies: by adding properties via relationships, we create a clear structure that can be used to run searches of either the properties (return all books where Author = Charles Darwin) or on the relationship (return all non-fiction books), and get resulting sets that include “On the Origin of Species.”
When concepts are visualized with their relations, ontologies are a web of information. Using common rules make ontologies interoperable. This interoperability allows information from different knowledge domains to be connected.
How do we use ontologies in biological sciences?
From how individual genes and what anatomical structures contribute to an organism, to a chemical library of compounds and molecules, to scientific evidence arising from laboratory experiments, ontologies are instrumental for data organization in the biological sciences (Chibucos et al., 2014; Degtyarenko et al., 2008; Haendel et al., 2009; The Gene Ontology Consortium, 2019). Arguably, the Gene Ontology (GO) (http://geneontology.org/), is the most familiar ontology in biology. GO describes how individual genes contribute to the biology of organisms at the organismal, cellular, and molecular levels. Another widely used ontology is the Uber anatomy ontology (Uberon, http://uberon.github.io/) (Haendel et al., 2009), a GO-integrated framework that describes body parts, organs, and tissues across animal species. Uberon unites anatomy ontologies for a growing variety of traditional and emerging research organisms, facilitating comparative evolutionary and developmental studies.
Why build an anatomy ontology?
Everything we study in biology comes down to a process that is happening in a place, in an organism. That single cell data? It came from stem cells sorted from the intestine of a mouse. That volumetric electron microscopy data? It came from mouse intestinal crypts. That in situ data that shows Lgr5 expression in mouse intestinal crypt stem cells… that crypt cell remodeling phenotype… all these disparate data, have the context of anatomy in common. Thus, anatomy is at the root of organizing seemingly disparate datasets and is a de facto way to aggregate data.
Does my research organism have an anatomy ontology?
If you work in an established research organism, great! You likely already have an anatomy ontology to hook your data up to. Check to see if your organism of choice has one at the Ontology Lookup Service. Almost any organism with a “base” (Flybase, WormBase…) already has an ontology and uses it to organize data within the base and as a framework for other tools, like Virtual Fly Brain (Osumi-Sutherland et al., 2014). If you work on an emerging research organism, and you are poised to generate a lot of data, there’s good news here, too. Many research communities are generating anatomy ontologies, notably Ciona and recently, Planarians (Hotta et al., 2020; Nowotarski et al., 2021).
What if my research organism doesn’t have an anatomy ontology?
If your research organism does not have an anatomy ontology, consider starting one! Assemble a squad with an expert(s) on the anatomy of your organism and at least one person who has some coding experience, and you can build an ontology for your data. The tools in the field are easy to use (Web Protégé, Git Hub and Google Sheets) and are becoming increasingly accessible with the ontology-development-kit (Matentzoglu, 2021), ROBOT (Jackson et al., 2019), and COGs (https://github.com/ontodev/cogs).
When’s the best time to build an anatomy ontology?
It is never too soon to put frameworks in place to organize and connect big data. For example, a growing number of labs use the planarian flatworm Schmidtea mediterranea as a research organism to model regeneration and stem cell biology, but there are still far fewer when compared to labs using Drosophila or C. elegans. Searching Pubmed for “Drosophila”, “C.elegans”, and “Planaria” yields 113,316; 35,030; and 1,884 papers, respectively. Going back to our original 5 paper a day example, it would take one person 62 years to read all of the Drosophila papers, 19 years to read all of the C. elegans papers, and just over a single year to read all of the Planaria papers. For the planarian field, this meant we were at a point where our data and information base was manageably small for a team of curators to capture all the anatomical terms needed for an ontology. As a general rule of thumb, it is a good time to build an ontology for data organization when the published record is still small enough for humans to read and process. That way, we ensure we can capture old data, as well as promote and ensure that future data can be integrated into a unified framework.
Why we need to use ontologies to organize big data:
If you’ll allow a somewhat geeky paraphrase, with big data, comes great responsibility. How do we handle big data responsibly? Efficiently? And in a way that is accessible and reusable? Luckily, we already have a framework in the form of FAIR. FAIR data practices insist that data be Findable, Accessible, Interoperable, and Reproducible (Wilkinson et al., 2016). When data is acquired and handled according to FAIR practices, everyone wins. Anatomy ontologies are Findable and Accessible when available through the Ontology Lookup Service (Jupp et al., 2015), are interoperable when using relationships found in the Relationship Ontology, and are Reproducible when reported in adherence to the Minimum Information for Reporting an Ontology (MIRO) practices (Matentzoglu et al., 2018). Adhering to FAIR practices while annotating anatomical data using an ontology ensures that all folks can access research and data more easily, source data has an opportunity to gather more citations, and importantly we all get more accessible science for our money.
Anatomy ontologies are the difference between hoarding data in piles versus curating and organizing biological data into a searchable library. Building an anatomy ontology for a research organism may seem like a big undertaking, but it is a necessary investment in the community, a tool everyone can benefit from. Consider our own experience: if two biologists and someone who scripts could build an anatomy ontology with help from the great community at the Open Biological and Biomedical Ontologies (OBO)foundry, so can you.
References
Chibucos, M. C., Mungall, C. J., Balakrishnan, R., Christie, K. R., Huntley, R. P., White, O., Blake, J. A., Lewis, S. E. and Giglio, M. (2014). Standardized description of scientific evidence using the Evidence Ontology (ECO). Database 2014,.
Conan Doyle, S. A. (1885). The Great Kleinplatz Experiment. The New York Times.
Degtyarenko, K., de Matos, P., Ennis, M., Hastings, J., Zbinden, M., McNaught, A., Alcántara, R., Darsow, M., Guedj, M. and Ashburner, M. (2008). ChEBI: a database and ontology for chemical entities of biological interest. Nucleic Acids Res. 36, D344–50.
Gonçalves, R. S., Horridge, M., Li, R., Liu, Y., Musen, M. A., Nyulas, C. I., Obamos, E., Shrouty, D. and Temple, D. (2019). Use of OWL and Semantic Web Technologies at Pinterest. arXiv [cs.CL].
Haendel, M., Gkoutos, G., Lewis, S. and Mungall, C. (2009). Uberon: towards a comprehensive multi-species anatomy ontology. Nature Precedings 1–1.
Hotta, K., Dauga, D. and Manni, L. (2020). The ontology of the anatomy and development of the solitary ascidian Ciona: the swimming larva and its metamorphosis. Sci. Rep.10, 17916.
Jackson, R. C., Balhoff, J. P., Douglass, E., Harris, N. L., Mungall, C. J. and Overton, J. A. (2019). ROBOT: A Tool for Automating Ontology Workflows. BMC Bioinformatics20, 407.
Jupp, S., Burdett, T., Leroy, C. and Parkinson, H. E. (2015). A new Ontology Lookup Service at EMBL-EBI. SWAT4LS2, 118–119.
Matentzoglu, N. (2021). INCATools/ontology-development-kit: June 2020 release.
Matentzoglu, N., Malone, J., Mungall, C. and Stevens, R. (2018). MIRO: guidelines for minimum information for the reporting of an ontology. J. Biomed. Semantics9, 6.
Nowotarski, S. H., Davies, E. L., Robb, S. M. C., Ross, E. J., Matentzoglu, N., Doddihal, V., Mir, M., McClain, M. and Sánchez Alvarado, A. (2021). Planarian Anatomy Ontology: a resource to connect data within and across experimental platforms. Development148,.
Osumi-Sutherland, D., Costa, M., Court, R. and O’Kane, C. J. (2014). Virtual Fly Brain – Using OWL to support the mapping and genetic dissection of the Drosophila brain. CEUR Workshop Proc.1265, 85–96.
The Gene Ontology Consortium (2019). The Gene Ontology Resource: 20 years and still GOing strong. Nucleic Acids Res.47, D330–D338.
Wilkinson, M. D., Dumontier, M., Aalbersberg, I. J. J., Appleton, G., Axton, M., Baak, A., Blomberg, N., Boiten, J.-W., da Silva Santos, L. B., Bourne, P. E., et al. (2016). The FAIR Guiding Principles for scientific data management and stewardship. Sci Data3, 160018.
The Human Cell Atlas (HCA) Developmental and Pediatric Cell Atlas meeting will be held August 25-27, 2021 online. Free registration up to and during the meeting is available at https://devped2021.humancellatlas.org/
This meeting will bring together diverse communities of scientists to plan how to map human development from conception to adolescence, and how to apply this knowledge to address important scientific and clinical challenges. We are particularly looking to engage developmental biologists and pediatric community members, as well as computational biologists, single-cell and imaging genomics experts, clinicians and ethicists who are interested in forging new collaborations to support this effort.
We have an amazing presenter line up, including: Hans Clevers, Kat Hadjantonakis, Muzz Haniffa, Aviv Regev, Sarah Teichmann, Sten Linnarsson, Deanne Taylor, Amos Tanay, and Barbara Treutlein. The meeting also will feature lightning talks, poster sessions and an ethics panel led by Bartha Knoppers and Jonah Cool, with panelists Helen Firth, Dimitri Patrinos, Vasiliki Rahimzadeh and Deanne Taylor.
To help participants connect around key research areas, a large part of the meeting will involve scientific discussion at breakout sessions covering the following topics:
Regulatory mechanisms in development (led by James Briscoe and Jesse Gillis)
Single cell genetics to highlight genes, pathways, cell types and tissues (led by Gray Camp and Xiao Chen)
Understanding cellular decision-making during development (led by Anne Grapin-Botton and Cantas Alev)
Imaging and spatial omics technologies and applications (led by Ali Erturk and Hiroki Ueda)
Lineage tracing, recording, clonal evolution, tagging and its applications (led by Samantha Morris and Nozomu Yachie)
Abnormal development in humans (led by Heather Etchevers and Stéphane Zaffran)
Clinical genetics – use of development and pediatric single-cell atlas data to identify disease causing variants in patients (led by Sarah Henrickson and Helen Firth)
Regenerative medicine (led by Guoji Guo and Jason Rock)
Developmental origins of health outcomes over a lifespan/Challenges of studying an organ from development to aging at the single cell level (led by Kricket Seidman and Arnold Kriegstein)
The ethics of working with human developmental and pediatric samples (led by Bartha Knoppers and Jonah Cool)
This meeting is generously supported by University of Toronto’s Medicine by Design program, the Children’s Hospital of Philadelphia, Cincinnati Children’s hospital, the Hospital for Sick Children, Toronto, Genome Canada, the Development journal, and the McLaughlin Centre at the University of Toronto. HCA gratefully acknowledges the Chan Zuckerberg Initiative and the Klarman Family Foundation for additional organizational support.
Postdoctoral fellow, Centre de Recherche en myologie, France
This Workshop was really on point for me, as it fitted very well with my research interests about intercellular communication and aging. First planned to take place at Wiston House, UK, it was rescheduled as a virtual meeting for safety reasons. I am so glad that the organizers and The Company of Biologists decided to go through with it in a virtual format. I do not think a virtual meeting is equivalent to an in-person event but the way it was set up on the Remo platform truly permitted great exchanges and even informal discussions. It also allowed for fantastic scientific discussions with great participants and we learned a lot from speakers from different fields.
The interdisciplinarity nature of the Workshop was highlighted by talks that discussed the relationship between inflammation, cellular senescence and regeneration in different tissues such as lung, cardiac or skeletal muscles and through different approaches from organoids to multi-omics. It was really fascinating to see how immune modulators or senescent cells can have such an impact on the orchestration of effective but also ineffective regeneration, depending on their timing and course of action. The relevance of the senescence-associated secretory phenotype in regeneration and aging was pointed out all along and concomitantly the importance of research concerning the drugs associated with it.
The Workshop was concluded on a satisfactory note emphasizing the fruitfulness of the interdisciplinarity of this field of study. As such, it also showed how we could gain much more by establishing collaborations between researchers working in these different disciplines.
In the end, it is true that we did not have crumpets with melted butter nor croquet on the lawn, as mentioned in the introductory talk, but we had wonderful scientific talks and the opportunity to engage in great discussions. I am grateful for the opportunity to have been part of it.
Laura Muraine
PhD student, Centre de Recherche en myologie, France
In the context of a pandemic, it is essential, more than ever, to maintain links and scientific discussions. This was achieved by the recent Workshop, organized by Helen Blau and Nadia Rosenthal, which focused on the question of pain and partnership between inflammaging and regeneration, and which brought together a number of great talks from junior and senior scientists from across the world.
As a PhD student focusing on fibrosis in skeletal muscle, I learned a lot about the inflammation part of the process and the remodeling of the immune system associated with aging. The presentation of a lot of ongoing unpublished work as well as the convergence of different fields, all part of the problem at hands, was highly informative. I really enjoyed the discussions on cellular senescence as an evolvable phenomenon and a physiological response, as well as the point made on the key secretory phenotypes of those cells.
The virtual setting did not take anything away from the relevance and quality of the talks and The Company of Biologists made a great effort of running the 3 days event smoothly. Even without the scones, tea and perfect scenery, the Remo platform made it easy to virtually go from table to table to further discuss the talks as well as share ideas and opinions with other attendees from the comfort of your own home. It was the best alternative to in-person meetings I have experienced so far and all the attendees were more than keen to discuss their research, leaving me feeling part of a community.
The opportunity to share and discuss my work without getting out of my sofa made this a real “feel at home” Workshop. To participate in this meeting was very rewarding and, as an early career researcher, I left feeling quite inspired and with a lot of new questions and approaches to explore in the context of my own research project.
Smilodon californicus and Canis dirus fight over a Mammuthus columbi carcass in the La Brea Tar Pits. Public Domain, via Wikipedia
In the latest episode of Genetics Unzipped, Dr Kat Arney delves back into the ancient past, winding the clock back thousands of years to discover the stories of Denisovans and direwolves that researchers are now able to read in the fragments of DNA left in bones or even dirt.
One of the people who’s digging into the past through the use of ancient DNA to understand why a species might have vanished is Dr Kieren Mitchell from the University of Adelaide. While many may think that his species of choice – the direwolf – is fictional, they were definitely real, but the reasons they went extinct may come down to being too choosy about their meals and their mates.
Kat also speaks with Dr Benjamin Vernot – a researcher at the Max Planck Institute for Evolutionary Anthropology in Leipzig, Germany. Rather than studying bones, he’s been digging for DNA in more unlikely places in order to unearth the stories from our ancient ancestors.
If you enjoy the show, please do rate and review on Apple podcasts and help to spread the word on social media. And you can always send feedback and suggestions for future episodes and guests to podcast@geneticsunzipped.com Follow us on Twitter – @geneticsunzip
Welcome to our gallop through the preprints in developmental biology (and related subjects) published in July.
The preprints this month are hosted on bioRxiv, arXiv and preprints.org – use the links below to get to the section you want. Stand-out section for this month may be the Tools & Resources with some exciting new tools and methodologies.
PPARdelta signaling activation improves metabolic and contractile maturation of human pluripotent stem cell-derived cardiomyocytes Nadeera M. Wickramasinghe, David Sachs, Bhavana Shewale, David M. Gonzalez, Priyanka Dhanan-Krishnan, Denis Torre, Elizabeth LaMarca, Serena Raimo, Rafael Dariolli, Madhavika N. Serasinghe, Joshua Mayourian, Robert Sebra, Kristin Beaumont, Ravi Iyengar, Deborah L. French, Arne Hansen, Thomas Eschenhagen, Jerry E. Chipuk, Eric A. Sobie, Adam Jacobs, Schahram Akbarian, Harry Ischiropoulos, Avi Ma’ayan, Sander M. Houten, Kevin Costa, Nicole C. Dubois
The blood vasculature instructs lymphatics patterning in a SOX7 dependent manner Ivy Kim-Ni Chiang, Winnie Luu, Key Jiang, Nils Kirschnick, Mehdi Moustaqil, Tara Davidson, Emmanuelle Lesieur, Renae Skoczylas, Valerie Kouskoff, Jan Kazenwadel, Luis Arriola-Martinez, Emma Sierecki, Yann Gambin, Kari Alitalo, Friedmann Kiefer, Natasha L. Harvey, Mathias Francois
The Imprinted Igf2-Igf2r Axis is Critical for Matching Placental Microvasculature Expansion to Fetal Growth Ionel Sandovici, Aikaterini Georgopoulou, Vicente Pérez-García, Antonia Hufnagel, Jorge López-Tello, Brian Y.H. Lam, Samira N. Schiefer, Chelsea Gaudreau, Fátima Santos, Katharina Hoelle, Giles S.H. Yeo, Keith Burling, Moritz Reiterer, Abigail L. Fowden, Graham J. Burton, Cristina M. Branco, Amanda N. Sferruzzi-Perri, Miguel Constância
Retinal ganglion cell-specific genetic regulation in primary open angle glaucoma Maciej S. Daniszewski, Anne Senabouth, Helena H. Liang, Xikun Han, Grace E. Lidgerwood, Damián Hernández, Priyadharshini Sivakumaran, Jordan E. Clarke, Shiang Y. Lim, Jarmon G. Lees, Louise Rooney, Lerna Gulluyan, Emmanuelle Souzeau, Stuart L. Graham, Chia-Ling Chan, Uyen Nguyen, Nona Farbehi, Vikkitharan Gnanasambandapillai, Rachael A. McCloy, Linda Clarke, Lisa Kearns, David A Mackey, Jamie E. Craig, Stuart MacGregor, Joseph E. Powell, Alice Pébay, Alex W. Hewitt
Liang, et al. use scRNAseq to identify the major cell types in the mouse liver
Downregulation of WT1 transcription factor gene expression is required to promote myocardial fate Ines J. Marques, Alexander Ernst, Prateek Arora, Andrej Vianin, Tanja Hetke, Andrés Sanz-Morejón, Uta Naumann, Adolfo Odriozola, Xavier Langa, Laura Andrés-Delgado, David Haberthür, Benoît Zuber, Carlos Torroja, Ruslan Hlushchuk, Marco Osterwalder, Filipa Simões, Christoph Englert, Nadia Mercader
Differential impact of a dyskeratosis congenita mutation in TPP1 on mouse hematopoiesis and germline Jacqueline V. Graniel, Kamlesh Bisht, Ann Friedman, James White, Eric Perkey, Ashley Vanderbeck, Alina Moroz, Léolène J. Carrington, Joshua D. Brandstadter, Frederick Allen, Adrienne Niederriter Shami, Peedikayil Thomas, Aniela Crayton, Mariel Manzor, Anna Mychalowych, Jennifer Chase, Saher S. Hammoud, Catherine E. Keegan, Ivan Maillard, Jayakrishnan Nandakumar
Evolution of a chordate-specific mechanism for myoblast fusion Haifeng Zhang, Renjie Shang, Kwantae Kim, Wei Zheng, Christopher J. Johnson, Lei Sun, Xiang Niu, Liang Liu, Theodore A. Uyeno, Jingqi Zhou, Lingshu Liu, Jimin Pei, Skye D. Fissette, Stephen A. Green, Sukhada P. Samudra, Junfei Wen, Jianli Zhang, Jonathan Eggenschwiler, Doug Menke, Marianne E. Bronner, Nick V. Grishin, Weiming Li, Kaixiong Ye, Yang Zhang, Alberto Stolfi, Pengpeng Bi
Comparing mice and zebrafish tissues from Matsubara, et al.
An open-access volume electron microscopy atlas of whole cells and tissues C. Shan Xu, Song Pang, Gleb Shtengel, Andreas Müller, Alex T. Ritter, Huxley K. Hoffman, Shin-ya Takemura, Zhiyuan Lu, H. Amalia Pasolli, Nirmala Iyer, Jeeyun Chung, Davis Bennett, Aubrey V. Weigel, Melanie Freeman, Schuyler B. van Engelenburg, Tobias C. Walther, Robert V. Farese Jr., Jennifer Lippincott-Schwartz, Ira Mellman, Michele Solimena, Harald F. Hess
A Comprehensive Overview of the Physical Health of the Adolescent Brain Cognitive Development Study (ABCD) Cohort at Baseline Clare E. Palmer, Chandni Sheth, Andrew T. Marshall, Shana Adise, Fiona C. Baker, Linda Chang, Duncan B. Clark, Rada K. Dagher, Gayathri J. Dowling, Marybel R. Gonzalez, Frank Haist, Megan M. Herting, Rebekah S. Huber, Terry L. Jernigan, Kimberly LeBlanc, Karen Lee, Krista M. Lisdahl, Gretchen Neigh, Megan W. Patterson, Perry Renshaw, Kyung E. Rhee, Susan Tapert, Wesley K. Thompson, Kristina Uban, Elizabeth R. Sowell, Deborah Yurgelun-Todd
Spatial mapping of protein composition and tissue organization: a primer for multiplexed antibody-based imaging John W. Hickey, Elizabeth K. Neumann, Andrea J. Radtke, Jeannie M. Camarillo, Rebecca T. Beuschel, Alexandre Albanese, Elizabeth McDonough, Julia Hatler, Anne E. Wiblin, Jeremy Fisher, Josh Croteau, Eliza C. Small, Anup Sood, Richard M. Caprioli, R. Michael Angelo, Garry P. Nolan, Kwanghun Chung, Stephen M. Hewitt, Ronald N. Germain, Jeffrey M. Spraggins, Emma Lundberg, Michael P. Snyder, Neil L. Kelleher, Sinem K. Saka
My name is Helen Zenner and I am delighted to introduce myself as the new community manager of the Node. I started my scientific life as a cell biologist, specialising in membrane trafficking, but over the years I have found myself being drawn more and more towards developmental biology. Of course starting a postdoc in Daniel St Johnston’s lab really accelerated this progression, and although I spent most of the first two years claiming not to be a fly person, I have now fully embraced both the fly and developmental biology community!
As community manager at the Node I am excited to build on the excellent work of Aidan Maartens and the Development team. Fortunately Aidan has left me instructions on how to run the Node, and although it doesn’t have ‘Do Not Panic’ written in reassuring large letters, it looks to be an excellent guidebook and hopefully the change will be seamless. The transition has also been supported by Helen and Esperanza who run our sister sites, preLights and FocalPlane, so thanks to them as well.
Going forward, I hope to bring you some of the new features that you asked for in our recent community survey, as well as some additional ideas that we hope you will enjoy. But first and foremost, I would like to remind you that anyone within the community is welcome to post on the site. You can either post directly on the site once registered, or if you have any questions or would like support I would love to hear from you at thenode@biologists.com or helen.zenner@biologists.com
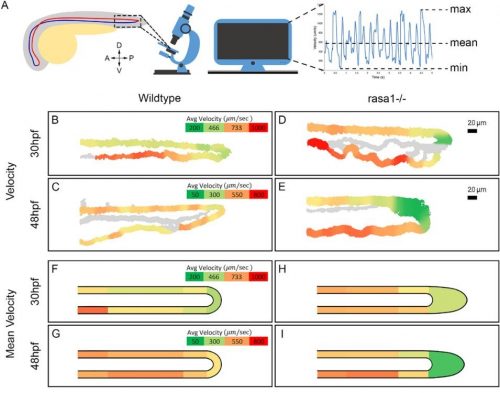
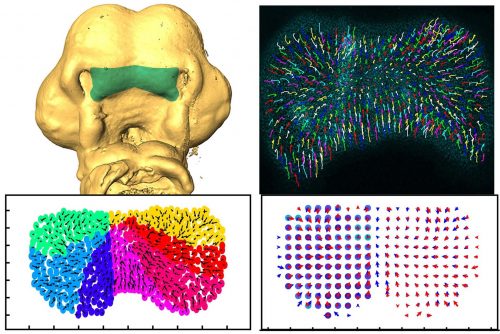
During vertebrate face development, bilateral streams of neural crest cells migrate from the neural tube to give rise to the facial prominences. A new study in Development combines high-resolution live imaging of chick facial development with a mathematical examination of cell behaviour to understand the dynamics of facial symmetry. We caught up with Adrian Danescu, Lisanne Rens and corresponding author Joy Richman (Professor and Director of the Pediatric Dentistry Graduate Program in the University of British Columbia in Vancouver, Canada) to find out more about the work.
Adrian, Lisanne and Joy (L to R)
Joy, can you give us your scientific biography and the questions your lab is trying to answer?
JR: I was originally trained as a dentist and then specialized in paediatric dentistry. It was while I was doing my specialty training that I first encountered the field of developmental biology. The tooth development project with Ed Kollar was so enjoyable that I went on to do a PhD in craniofacial development with Cheryl Tickle at UCL. She was an outstanding mentor and although your audience will be very familiar with her pioneering work on limb development, she also had three students that worked on the face and I was one of them. After that experience in London, the chicken was my main model organism until the last 15 years when I started also working on non-avian reptiles (lizards, snakes, turtles). My group has made discoveries concerning the molecular mechanisms of facial morphogenesis; however, all our previous work was done with static analysis. This is our first foray into time-lapse imaging to describe cellular behaviours in real time. We certainly did not anticipate the striking choreography of cell movements in the face way back when we started this project.
Adrian, how did you come to join Joy’s lab and what drives your research today?
AD: Like Joy, I also trained as a dentist, in Romania, and then came to Canada to take an MSc degree. I was interested in embryology and I decided to enrol in a PhD project in a lab that focused on craniofacial development – Joy’s lab at UBC was a perfect fit for me. My project involved facial birth defects, and their proper study required a suitable model. The lab was well versed in avian techniques and for me it was essential to learn quickly all the technical aspects necessary for the ongoing project. My entire PhD was a dynamic journey, with lots of opportunities for exciting projects and networking.
My main accomplishment in the lab was to develop a system to observe the movement of mesenchymal cells within the face during the early stages of development, prior to lip fusion, with high-resolution microscopy. This took several years of optimization and painstaking attention to subtle things such as finding a way to label individual cells. At the beginning of the project, I collaborated with an expert in lipid nanoparticles so I could deliver plasmids to the chicken face without electroporation. I now recognize the practical applications of these in vivo transfection methods after the implementation of the same technology to make COVID-19 mRNA vaccines. After gathering detailed tracking data, we started to collaborate with Leah Edelstein-Keshet and her postdoc Lisanne Rens from the department of Mathematics at UBC. They came at the data from a different angle and thanks to their insights the paper reached a deeper significance.
Lisanne, what is your research history and how did you come to be involved in this project?
LR: I am from the Netherlands, which is also where I obtained my PhD in mathematical and computational biology (from Leiden University). I typically use mathematical modelling techniques to describe processes in development, such as cell migration, angiogenesis, branching morphogenesis. At the start of this project, I was a postdoc at the mathematics department at UBC, working with my advisor Leah Edelstein-Keshet. I became involved in this project because Joy and her group were looking to quantify their data and understand it better. They got in contact with Leah, and I was very excited to be involved because I was already familiar with suitable methods, which I previously used to quantify simulated data. Just to be able to work with experimental biologists is a great opportunity. After our first meeting, Adrian sent me their data, and then it all started.
How has your research been affected by the COVID-19 pandemic?
AD & JR: I think we were fortunate enough that most of our experiments and analyses were completed before COVID-19 changed our way of working. There were only a few experiments left to finalize the paper and we planned them right after the restrictions were lifted. Communication via online platforms was convenient enough, especially for being able to bring other people from various places to participate in our discussions. However, the lab closure delayed the time we initially planned to finish the paper.
LR: For me, as a computational biologist, working at home is not as big of a deal compared to wet-lab biologists. However, like with any person, working from home comes with challenges. I find it lonelier, and it’s harder to keep being motivated. Also, with school closures, the kids are around.
What is the theory of developmental instability, and how does it relate to craniofacial abnormalities?
AD, LR & JR: Developmental instability refers to the range of fluctuations during development that can be usually compensated by various mechanisms to maintain normal development. There are both genetic and environmental factors at play, but our focus was to find evidence of instability at the cellular level. The theory is that all embryos will have some degree of developmental instability and, in the majority of cases, normal morphogenesis occurs. However, a slight increase in instability may be enough to lead to congenital malformations such as cleft lip. This instability is not only particular to the craniofacial area, but it may also concern all organs. One of the ways to measure developmental instability in other systems is to look at symmetry. Since we had dissected the midline of the face, we were able to compare the left versus right side. Through fruitful discussions with Lisanne and Leah Edelstein-Keshet, the idea to map the data back onto a grid was developed. In that way, we could compare directly each grid reference point in the equivalent anatomical location.
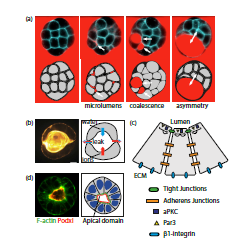
Composite figure of the key experimental data in the paper; for more details, see the full manuscript
Can you give us the key results of the paper in a paragraph?
AD, LR & JR: In this study, we wanted to understand the striking shape changes in early facial development prior to lip fusion. We turned to high-resolution live imaging in order to globally track hundreds of individual mesenchymal cells across the frontonasal mass. First, we discovered that mesenchymal cells moved; second, the movements alternated between states of order and disorder; third, clustering algorithms revealed that the movements are coordinated over large distances; and fourth, by interpolating the data we found symmetry that also fluctuates over time. We then used this interpolated data to map patterns of divergence and convergence that are again cyclical. We showed that all these cell behaviours are dependent on the actomyosin network. One of the most interesting observations made through mathematical modelling was the correlation between the switches in direction of movement with transitions from states of order to disorder.
What did the mathematical analysis of cell behaviours reveal that simple observation could not?
AD, LR & JR: First, the analysis confirmed some of the patterns we thought we were seeing. For instance, after quantification, the symmetry we noticed by eye became very clear, and the loss of symmetry was tremendous in the knockout tissue. We also noticed changes in the direction of the cells, but the pattern was not clear. This is where a divergence analysis helped. It revealed bands of convergence and divergence, and how they changed over time. Our velocity correlation analysis revealed the spatial distance across which cells are seemingly able to communicate. By clustering algorithms, we identified the spatial regions of coordinated motion, which was not possible to do by hand.
Furthermore, the order/disorder and the K-means clustering analyses revealed fluctuations of cell behaviour at a smaller scale, indicating developmental instability during normal midface development. Modelling also helped us discover the rapid switches of cell direction between divergence and convergence that happens within 20 minutes. The overall symmetry and periodicity during midface development were identified by the mathematical modelling as well.
The cell movements you observe are often symmetrical – what might explain this coordination of behaviour over such a long range?
AD, LR & JR: Several pathways, such as WNTs, BMPs, SHH, FGFs and so on, are at play during midface morphogenesis, and they may have a role in regional or more global coordination of cell movement. Furthermore, the forces generated through the extracellular matrix may contribute to these movements as well, due to cells being connected as a network. We identified one potential candidate to be WNT5A signal, with its expression overlapping the band of divergence and convergence that we identified. Our observations will pave the way for future investigations into the molecules that play an essential role in either buffering against instability or promoting the fluctuations. Ultimately, gene pathways associated with increased risk of clefting will be tested in this system.
When doing the research, did you have any particular result or eureka moment that has stuck with you?
AD: The first moment that stuck with me was observing how dissected faces grew and developed normally in a culture environment. I realized the advantage of this system, being suitable for direct observation under the microscope, in contrast with the side positioning that can be accessed in a developing embryo inside the egg. However, this was just the beginning of a long and laborious process to set up a series of methods to pursue my project.
LR: I was most surprised that we were able to link changes in divergence/convergence and coordinated/uncoordinated motion of the cells. Both of these quantities varied in time in a similar periodic fashion. This bears the question which one of those precedes the other, or how they are linked.
And what about the flipside: any moments of frustration or despair?
AD: There were countless moments of frustration, as always happens in research. Along the years, I learned to manage and embrace them as a way to grow. Learning the dissection techniques, optimizing the imaging setup for better clarity and stability of the culture during imaging, finding the right people to contribute to the project were all challenging, but what matters in the end is never, ever give up.
LR: For me, luckily not as many frustrations as with other projects I’ve done! I think the biggest challenge was how to interpret what the quantification of the data was telling us: what are the possible underlying mechanisms?
What next for you two after this paper?
AD: I graduated from both the PhD and the clinical program in Orthodontics last year. I have started to look for opportunities to find my way back into research. I intend to find the optimal way to work as a clinician and dedicate time for research. I expect this process to take longer as we are in the third wave of COVID-19 now in Canada, and there is still a lot of concern and uncertainty for the near future.
LR: I recently started a tenure track position at the TU Delft in my home country. I work in the mathematical physics group, continuing my line of research into mathematical modelling of cell and tissue biology.
Where will this story take the Richman lab?
JR: I would love to go further into the environmental influences that most affect developmental instability. I will expand the work to test specific pathways as mentioned above. In the current paper, we employed a global block on small GTPase signalling, but in the future we will be more specific.
I would love to go further into the environmental influences that most affect developmental instability.
Finally, let’s move outside the lab – what do you like to do in your spare time?
AD: I recently moved to Ontario after many great years in beautiful Vancouver. I generally try to allocate time for cooking, reading, travelling, doing outside activities, and spending time with my wife. I am learning that Ontario is fantastic for outdoor explorations with numerous lakes and parks. The summer is coming, and hopefully we will be able to enjoy a more normal summer after a tough year with so many lockdowns.
LR: Classical ballet is a big passion of mine, so during my time in Vancouver, I was practicing ballet downtown for two hours per week. Now in the Netherlands, I am back at my old ballet school. I also have two young kids who I spend most of my spare time with.
JR: I enjoy walking along the beach or in one of the many lovely urban forests near the UBC campus with my dog. I also am a passionate Masters swimmer and hope to return to club swimming soon. Right now, the pool is shut because of COVID-19. I also have really enjoyed spending lots of quality time with my two children in their twenties who came home to shelter during the pandemic. Oh yes, I have also kept my sour dough starter alive for a year now. Being a scientist helps!
Kat explores the world of transmissible tumours, looking at the history of contagious cancers in Tasmanian devils, dogs, clams and cannibal hamsters. Plus, the story of the man who caught cancer from his tapeworm.
If you enjoy the show, please do rate and review on Apple podcasts and help to spread the word on social media. And you can always send feedback and suggestions for future episodes and guests to podcast@geneticsunzipped.com Follow us on Twitter – @geneticsunzip

 (No Ratings Yet)
(No Ratings Yet)





{kind=link}
.jpg){kind=link}