I am excited to announce the release of our event carbon calculator, our latest resource to support the creation of sustainable events. Estimate your event’s carbon footprint in five minute or less. Our event carbon calculator provides insights into the main drivers of your meeting’s greenhouse gas emissions, offers reduction scenarios to test and provides resources to help you build a sustainable strategy for your event.
Our calculator is still in its beta stage, so please explore our tool and send us feedback on how to make it better suited for your needs. We hope you will enjoy using our event carbon calculator and that it will help to inspire more sustainable actions in your academic meetings.
At the end of each month, I pick the same month from a random year from the past 15 years of the Node, and take a look at what people were talking about back then.
Previously, I’ve been busy travelling back to February 2011, March 2013, April 2014 and May 2016 to have a look around the Node. It’s been really fun peeking around the archives. This June, let’s jump back to 2013…
We used to partner with the Woods Hole embryology course to run an image competition, with the winning image being featured on Development’s cover. We’re hoping to revive this tradition – watch this space!
The Fly Board is excited to announce the second cycle of FlyCROSS (2025–2026), following a successful launch in 2024. The goal of FlyCROSS is to empower early career Drosophila researchers by connecting them with mentors who can help them fly in their careers – whether in academia or beyond.
Through FlyCROSS, early career scientists in the Drosophila community will be paired with an experienced mentor who will help them navigate the world of fly research with ease. Designed to prioritize the needs of mentees by matching them with mentors of their preferences, the program offers seasoned researchers an opportunity to share their knowledge and pay it forward by supporting scientists who are seeking guidance and fresh perspectives. The program consists of three parts: the mentor survey, the mentee survey, and matching.
We are currently seeking mentors. Scientists with experience in Drosophila research, includingpostdoctoral researchers, faculty members, and those in equivalent roles across diverse scientific career paths are eligible to serve as mentors and invited to complete the Mentor survey by July 14. The mentor survey will gather important details about your areas of expertise and personal career journey. We’re looking for mentors from all career paths—academia, industry, biotech, science communication, policy, and more—who have experience with Drosophila research and are eager to support the next generation of fly scientists. (If you have earned a PhD and are currently involved in any scientific career path and have experience working with fruit flies as a model system at any stage of your career, we encourage you to volunteer to support the next generation scientists)
The prospective mentee survey will be open July 21 through September 1 and will collect in-depth information about the type of mentorship each mentee is seeking. Early-career scientists including graduate students, postdoctoral researchers working in fly labs or equivalent settings, and pre-tenure faculty leading Drosophila research groups are eligible to participate as mentees.
Mentees will review the list of available mentors and their responses, with the option to indicate their preferred mentors. After receiving mentee submissions, the FlyCROSS Committee will carefully review the preferences and finalize pairings, ensuring an optimal match between mentors and mentees. Matches will be announced by October. Incase of any queries reach out to dmelcross@gmail.com. FlyCROSS was adapted from a similar mentoring program running in the worm community, and we sincerely appreciate their support of our endeavor.
Thankyou Shefali FlyCROSS Co-chair Grad student rep, Fly Board PhD candidate, Tennessen Lab IU Bloomington Bluesky : @iamshef.bsky.social X : @iam_shef
A fever dream of science and spirit wrapped in the forests of Bohemia. Here’s an escalating list — starting with the standard (though nothing was standard about this) — and spiraling toward the legendary. This was what made Woodstock.Bio² & Night Science the most unforgettable conference of its kind (there is no such kind).
What We Had:
· Scientists across all career stages — from undergrads to full professors (yes, there was one undergrad! He’s ahead of his time).
The people. Each and every participant of this meeting.
In retrospect, being forced to choose a walk-up song might have been one of the most brilliant conference design decisions. It pushed us out of default mode, pipetted creativity and humor into the air, and made each session feel more like a show than a slot.
And because everyone was “performing” just a little — rethinking how to present their science in a more human, fun, or offbeat way — we found ourselves better able to stay tuned to ALL talks, even when they spanned wildly different disciplines. There was something about the levity and variety that made us listen more deeply.
Even the hashtag had a story. We used #TCTeAC on BlueSky – a compact string chosen as shorthand for #TheConferenceToEndAllConferences. It’s an acronym of the full phrase, but we also loved how it resembled a DNA sequence or binding motif. In a way, it was a motif — one you had to chant silently each time you typed it out.
TCTeAC… The Conference To end All Conferences…
TCTeAC… The Conference To end All Conferences…
But maybe it wasn’t just the events. It was the environment. Once a space was created where it felt safe — and common — to laugh, to question, to try weird things, to fail publicly, to be human — everything else followed. For all of us, as a collective.
People stepped out of their academic armor. Speakers forgot their next line, slides froze, and the crowd cheered. Honest dialogue blossomed. Ideas were born not just during talks, but on trains, under canopies, beside the fire.
“This project? It started with a chat on the train to the woods during Woodstock.Bio².”
Already thinking about the next one? The only spoiler I would give is that less than 24 hours after the ending of the meeting, the instigators and bohemian fixers were already conspiring on ideas for the next meeting. Needless to say the ideas are beyond imagination and bigger than life.
My talk in the first session was accompanied by a local bass guitarist:
* Thank you Patric! You rock! (and roll…)
We had a huge amount of exciting talk across a broad range of scientific fields:
The final location was a YMCA campsite at Southeast Central Bohemia:
But most importantly, we were strict with time, if a speaker went overtime and would not leave the stage – we took care of that, to keep the tight schedule of the meeting:
* This act was coordinated with the speaker ahead of time (in other words… just a joke).
This is part of the ‘Lab meeting’ series featuring developmental and stem cell biology labs around the world.
Lab introduction
Mariana S. Silveira: I am the head of LINDes, my current laboratory established in 2023. Prior to that, I shared another research space, the Neurogenesis Lab, with colleagues and my former advisor, Dr. Rafael Linden. I held the position of Associate Professor at the Institute of Biophysics Carlos Chagas Filho, part of the Center for Health Sciences at the Federal University of Rio de Janeiro, Brazil. Notably, this institution is the first university founded in our country, previously known as the University of Brazil, and our Institute is now approaching its 80th anniversary. In Brazil, this represents an esteemed and traditional institution, as our country is relatively young. Our Institute’s Program of Biological Sciences–Biophysics, one of the first doctoral programs in Brazil, has a history spanning 62 years.
The lab is in Rio de Janeiro and our city boasts unparalleled natural beauty and a vibrant cultural scene.
Research summary
Our current research focuses on leveraging advancements in retinal development and cell reprogramming to explore potential therapeutic strategies for vision loss. Specifically, we focus on retinal ganglion cells, the projection neurons of the retina, working in collaboration with both Brazilian and international research groups. Additionally, we are investigating the retinal microglia in a collaborative project aiming to design innovative tools employing machine learning for morphological categorization.
Lab roll call
José Nilson dos Santos – He’s been the go-to technician ready to help anyone in the lab ever since the lab head was a PhD student in the Neurogenesis Lab. Always there to tackle any kind of problem, it’s awesome to have him around!
Mariana S. Silveira – Associate professor and lab head. Honestly, leading this fresh and young team is both a challenge and a joy. My goal is to put all my energy into making sure they have a productive and happy experience in the lab.
Viviane Oliveira Valença – Postdoctoral researcher, Viviane serves as my right hand. Having recently completed her PhD; she is currently assisting in training the group and moving forward some new collaborations while finalizing manuscripts. Her presence is truly invaluable to the lab.
Daianne Torres – A combination of lab manager and technician, Daianne is consistently available to address both administrative and technical challenges.
Isabel Guedes Ferreira – A highly skilled Master’s student currently focusing on the study of retinal microglial morphology and the development of computational tools to enhance and optimize these analyses.
Bernardo Benincá – He has recently started his Master’s program, where his research project focuses on stimulating the reprogramming of Müller glial cells in vitro through the overexpression of transcription factors.
Kauã Mourão – An undergraduate student in Biological Sciences who joined the lab as a trainee in 2024. He is currently exploring various techniques to evaluate the potential for reopening the window of retinal ganglion cell generation and the synaptic integration of newly formed neurons.
Victoria Mattos – An undergraduate student in Biomedical Sciences who joined the lab in 2024. She is currently acquiring the essential foundational techniques needed to initiate her own project. Presently, she assists other students with their work and participates in routine laboratory activities.
Camila Barbosa – An undergraduate student in Microbiology and Immunology who joined the lab in 2024 and is currently enhancing her technical skills.
Loreena Klein – A PhD candidate who has recently joined the laboratory after completing her Master’s in the field of pain regulation, is currently investigating the regenerative potential of Müller glial cells for the generation of retinal ganglion cells in vivo.
Roberto Matias – An undergraduate student specializing in Biophysics, Roberto contributes to Isabel’s research by assisting in the morphological analysis of microglial cells across various functional states and investigating their correlation with neurodegenerative processes.
Talia Pontes – An undergraduate student in Biomedical Sciences, has recently joined the laboratory with the intention of pursuing a graduate program in the future.
Top row, from left to right: Jose Nilson, Daianne, Bernardo, Isabel, Kauã, and Roberto. Bottom row, from left to right: Loreena, Viviane, Mariana, Camila, Victoria and Talia.
Favourite technique, and why?
Mariana: In my opinion, microscopy remains an invaluable tool, particularly with advancements in resolution and the ability to detect multiple antigens simultaneously. This approach provides precise in situ information when combined with cell morphology, defining in a remarkably accurate way, cell identity within tissue. Nevertheless, employing a combination of various methodologies is always the optimal strategy. Recently, I have also become fascinated by scRNA-seq as a very relevant tool.
Mariana, apart from your own research, what are you most excited about in developmental and stem cell biology?
Mariana: The emerging field of organoids and assembloids, particularly for investigating early stages of brain development and the underlying mechanisms of diseases, is truly captivating.
Mariana, how do you approach managing your group and all the different tasks required in your job?
Mariana: I must admit it’s not an easy task. My approach involves holding regular meetings with the team, where we not only discuss relevant literature related to our projects and related fields but also hold individual and group follow-ups to review the goals set for each member. Despite time being limited, I always keep my office door open for them.
What is the best thing about where you work?
Mariana S. Silveira – Although science funding in Brazil remains quite limited, which makes competing for international grants an essential challenge, I truly appreciate working at such a prestigious institution. Here, we are often supported and encouraged to strive for quality and excellence, especially in training the next generation of researchers. While the number of young individuals pursuing this demanding career is gradually declining, it’s rewarding to discover talented individuals and witness their scientific growth and development. Celebrating small achievements serves as a motivation to keep moving forward.
Viviane Oliveira Valença– We are at one of Brazil’s top universities, which is definitely the highlight for me, as numerous scientific contributions are made here. Even though we lack good infrastructure, safety, and other resources, being inside the university and surrounded by students from different fields fosters interaction and knowledge exchange.
Daianne Torres – The best thing about our workplace is the people who make up our lab. Even when we’re stressed about failed experiments, having supportive and caring teammates always makes the effort worthwhile.
Isabel Guedes – The best thing about our lab is the camaraderie and sense of community. We genuinely look out for each other, and that support makes even the hardest days a lot easier to get through.
Bernardo Benincá – It is the people. The scientific environment is challenging and can often be frustrating, but we take great pride in the quality of the work we produce despite numerous hardships and limited funding. Another important factor is how we always support each other, creating a welcoming and inclusive space. It’s an honor to be part of our lab.
Kauã Mourão – The people. They are not only my lab group with whom I talk about research and papers, but also my friends whom I know I can count on in this crazy work routine. It is really great to share my workspace with them because I learn more every day. They are truly skilled at what they do.
Victoria Mattos – It’s all about the connections we build with people and what we learn. Joining the lab helped me grow, both as a student and as a person. Being part of this environment and contributing to our research is very rewarding.
Camila Barbosa – The great thing about working in this lab is the constant exchange with the whole team, which always helps me learn something new, whether during experiments or in our meetings. Outside the lab, we have the privilege of being close to renowned professionals, and we get to attend various lectures and conferences, which also helps broaden our scientific perspectives.
Loreena Klein – The best thing about where I work is the strong sense of community. Everyone is very supportive and willing to help, which has made a big difference for me as a new PhD student adjusting to the environment.
Roberto Matias – I can say for sure that the best thing about where I work is the patient, dedicated and fun people who guide me in everything I need to learn, whether they are colleagues or teachers. The Institute is a peaceful and friendly place capable of comforting anyone in difficult times.
Talia Pontes – Honestly, the best thing about working here is getting to learn hands-on lab techniques from experienced researchers right at my own university. Plus, it feels great to know I’m helping push the boundaries of retinal studies.
What’s there to do outside of the lab?
Mariana S. Silveira – Besides enjoying the quiet pleasure of reading a captivating romance, I’m a big fan of Brazilian music. I enjoy attending live shows and participating in groups that use samba school instruments to explore the diverse rhythms of our rich musical culture.
Viviane Oliveira Valença – Here in Rio de Janeiro, there are plenty of things to do outside the lab, such as hiking in various places where you can enjoy breathtaking views of the city. Besides that, you can go to the beach, soak up the sun, and spend quality time with friends.
Daianne Torres – During my free time, I enjoy curling up on the couch with a good TV series to unwind or diving into the pages of a new book to escape into a world of fantasy.
Isabel Guedes Ferreira – Outside of the lab, I like to unwind by reading, playing games, and spending time with my friends. It helps me recharge and keep a healthy balance.
Bernardo Benincá – Sometimes it is important to take a break from the routine and do things to relax. Hobbies like watching movies, reading, playing video games, going out with friends, or simply going to the garden and looking at the trees.
Kauã Mourão – We are talking about Rio, so I must mention the beaches! They are amazing! There is nothing better than heading to the sea and unwinding from everything.
Victoria Mattos – Outside of the lab, I enjoy spending my time with my family and friends, it’s always a lot of fun to be surrounded by good people in a nice place. I like to go to the beach, enjoy samba and visit new restaurants.
Camila Barbosa – In my free time, I enjoy spending time with friends, going hiking, connecting with nature, watching series, and reading.
Loreena Klein – Outside of the lab, I love spending time with my friends, whether we’re going out or just hanging out together. I also enjoy staying home and watching some TV shows, which help me relax and recharge.
Roberto Matias – Rio de Janeiro offers many interesting activities, such as modern museums, restaurants with unique and exotic cuisine, and adrenaline-pumping amusement parks. With such a beautiful city full of activities and people, it is impossible to decide what to do.
Talia Pontes – Outside of the lab, I enjoy watching movies, reading, and spending quality time with my family. I also dedicate time to studying extracurricular subjects and learning new languages—currently, I am studying Spanish.
The recent Special Issue from Journal of Cell Science focussed on the cell biology of mitochondria. To showcase some of the research in this issue, FocalPlane and MITOtalks have teamed up to host a webinar on Thursday 3 July at 16:00 BST (17:00 CEST, 11:00 EDT, 08:00 PDT). MITOtalks organisers Nuno Raimundo and Sjoerd Wanrooij have invited Yuli Buckley, Mireia Nager and Dikaia Tsagkari to present their research.
Prof. Caren Norden (@nordenlab.bsky.social) appointed Darwin Professor of Animal Embryology in the Department of Physiology, Development, and Neuroscience at the University of Cambridge.
Dr. Júlia Peloggia de Castro (@jupeloggia.bsky.social) awarded the Behrensen – Guzmán Palma Award for the most outstanding research paper by a predoctoral student at Stowers Institute for Medical Research.
Special thanks to Maddie Ryan, Charli Corcoran & Michaela Noskova Fairley for putting this digest together! If you would like to thank the Zebrafish Rock! team for their time & effort, you can buy us a strong cuppa at the link below. Every little bit keeps us caffeinated and motivated! We appreciate your support 🙂
Do you have news, jobs or research that you want to add to the next digest but don’t have a social media account? Use the submission form at our website below:https://linktr.ee/zebrafishrock
Our July webinar features three early-career researchers working on early embryogenesis and will be chaired by Development’s Executive Editor, Alex Eve.
André Dias (Universitat Pompeu Fabra) ‘Opposing Nodal and Wnt signalling activities govern the emergence of the mammalian body plan’
Nikhil Mishra (Institute of Science and Technology Austria) ‘Geometry-driven asymmetric cell divisions pattern cell cycles and zygotic genome activation in the zebrafish embryo’
At the speakers’ discretion, the webinar will be recorded for viewing on demand. To see the other webinars scheduled in our series, and to catch up on previous talks, please visit: thenode.biologists.com/devpres
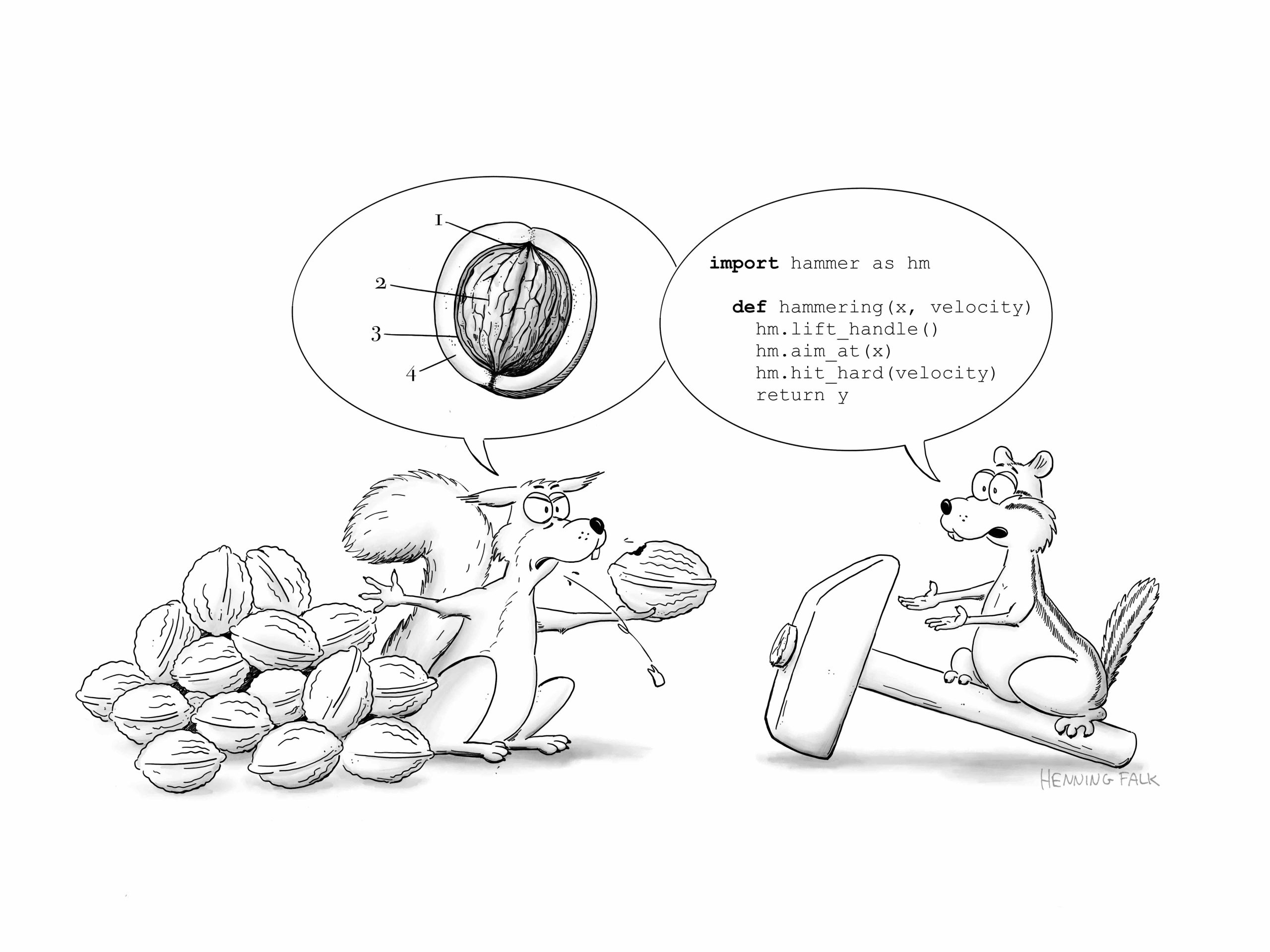
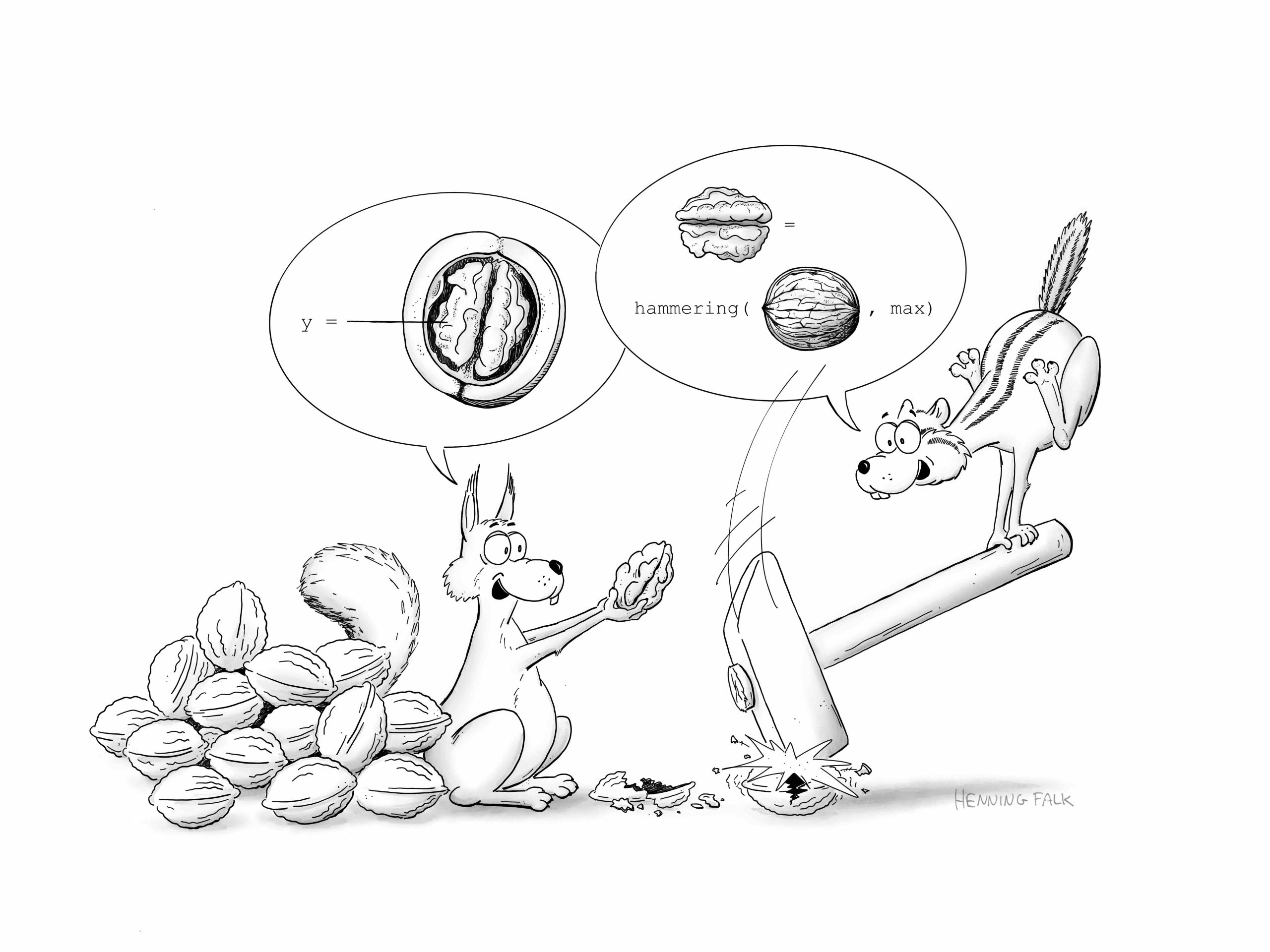
In this SciArt profile, we find out more about Henning Falk, who did a PhD in developmental biology, and now enjoys creating science-related cartoons as a freelancer.
[Click on the images to view the full-size versions of the cartoons]
Can you tell us about your background and what you work on now?
I studied molecular biotechnology and completed my PhD at EMBL Heidelberg, where I focused on somitogenesis in mouse embryos. Microscopy has always been my favorite technology, and during my PhD project, I developed a method to live-image mouse embryo development for up to three days. After finishing my PhD, I left research and transitioned from microscope development to CNN-based image analysis, and eventually to intelligent document processing using large language models. I currently work as a project manager for a company that builds document processing solutions. Creating cartoons for science communication serves as my connection back to my former researcher self.
The cartoon was done for German BioImaging (GerBI-GMB) to illustrate the enabling power of research data management (RDM)
Were you always going to be a scientist?
No, my early career ideas went all over the place, though they did include both biologist and artist – sometimes leaning more toward one, other times toward the other. Now I’m a customer AI project manager – life certainly takes surprising turns sometimes.
Yes, absolutely. In kindergarten, I gave myself the self-imposed task of drawing one picture every day to bring home. During primary school, I was a regular participant in local painting competitions. Later, I focused more and more on cartoon drawing – I’m not really sure why, but somehow it stuck with me.
This cartoon is part of a series of post cards raising awareness for the work of the NFDI4BIOIMAGE consortium. Published under a CC BY 4.0 licence.
What or who are your most important artistic influences?
My biggest influences are other cartoonists who manage to convey their message – their joke – in just one simple drawing and a sentence or two. For me, that’s the real essence of good cartoons: condensing a story so drastically that one image is enough to give you the background story, introduce the characters, and deliver a punchline. It’s pretty remarkable. Ralph Ruthe and Joscha Sauer (maker of “Nichtlustig”) are two German cartoonists I find quite inspiring and funny.
Does your science influence your art at all, or are they separate worlds?
I started creating scientific cartoons for friends and colleagues when I was still working on my PhD, and I also used drawings in my own presentations to illustrate my results and theories. I received a lot of positive feedback. A few years ago, I began doing scientific cartoons and illustrations as a freelancer. So today, it’s not my science influencing my art, but rather the science of my “clients.” I find it tremendously fun to work together with researchers to figure out exactly what message they want to convey and how to transform that abstract concept or complicated experimental result into a simple drawing. It’s a tug of war between completeness and simplicity, and sometimes we go a long way together – from a page-long list of detailed sub-results to nailing down the 2-3 main takeaway points that fit into one cartoon. This is where my background in science, my love for cartoon drawing, and my project management skills come together nicely. Voilà!
Figure for a publication depicting the challenges bio imaging scientists are faced with. Proper handling of large-scale, complex data is a challenge for researchers, data providers and users. https://f1000research.com/articles/11-638
I just finished a project for a psychologist, and before that, there were several projects in biological data management. As for what comes next, I don’t know yet.
This figure was created for a talk by Christina Derksen as basis to talk about the best practices for the development of clinical decision support systems (CDSS) in medicine by highlighting the most common pitfalls.
How/where can people find more about you?
I have a small website advertising my services and showcasing some of my recent projects. It’s falk-illustrations.de.
As mammals, we like to think of ourselves as dominant animals, and to some extent, that is true. Humans, specifically, have altered nearly every part of our planet and reshaped the climate. We’ve domesticated cattle and chickens for food and bred cats and dogs for pets. We exterminate countless other species across the tree of life. Yet, we and our mammalian relatives were not always in control, and throughout life’s history, our influence has ebbed and flowed.
Dr. Caleb Gordon, a paleontologist in the Earth and Planetary Sciences Department at Yale University, spent several years unraveling a dynamic tale that began 300 million years ago. At that time, “the biggest animals on land were our ancestors—the sprawling, sharp-toothed cousins of mammals, and the biggest animals in the oceans were giant armored or fleshy-finned fishes,” Caleb speaks enthusiastically. “The earliest reptiles appeared around this time too,” he introduces them with a smile. “Reptiles are a big, quirky family with lizards, crocodiles, turtles, dinosaurs, and birds. They include many other species, both living and extinct, that descended from an ancient common ancestor. Reptiles were diminutive little creatures in a world dominated by bigger, scarier animals who probably ate them whenever they found them.” On land, reptiles lived in the shadows of large mammal ancestors, eating insects and other small prey. They were not thriving in the oceans either, where various big fish were the top predators.
Then, some 50 million years later, a sudden surge of volcanic activity and environmental upheaval transformed the landscape and the plot. “The Great Dying, we call it, triggered the worst mass extinction in the history of animal life, around 252 million years ago,” Caleb explains in a more serious tone. “Up to 95% of species were wiped out, and almost anything bigger than a modern-day deer – gone.” But reptiles were lucky. In the aftermath, they thrived, both in aquatic and terrestrial arenas. In their “glow-up era – the Age of the Reptiles,” they became the new top predators on land and in the oceans.
Back in the present day, I sit in a local New Haven coffee shop with Caleb as he draws an evolutionary tree in his notebook. He shows me when reptiles and mammals first evolved and how they have competed throughout Earth’s history. “It’s hard for many of us, at least here in the northeast U.S., to imagine encountering big reptiles today. If we hike through a national park or drive through our local neighborhood, most of the big animals we’ll see are mammals — deer, cows, bears, dogs, wolves, coyotes, and especially humans. And if we swim in the cold coastal waters, most of the big animals we’ll see are sharks, bony fish, or whales,” he says. For his dissertation, Caleb explored a different landscape. He wanted to understand how, in the aftermath of the Great Dying, reptiles managed to win both on land and in sea as the Age of Reptiles began.
“There is an underappreciated role for the evolutionary innovations that helped reptiles win,” he explains. By evolutionary innovations Caleb means “complex features they evolved that allowed them to do new things and invade new environments after the Great Dying.” Such traits appear randomly, and on rare occasions, favoured their survival in certain circumstances. Little by little, through small changes over many years and generations, these beneficial traits became long-lasting innovations in reptiles.
To transition to water and survive the rough seas, reptiles needed special evolutionary innovations. “Marine reptiles were, over the course of millions of years, able to change the dimensions of their arms to create their own flippers. They did this by making their hands much longer while shortening their lower arm bones.” Caleb demonstrates by shrugging his shoulders close to his ear, squeezing his arms by his body, while extending and flapping his hands out. “Reptiles were able to do this quickly and easily, a bunch of times, in a bunch of different groups. But in mammals and their ancestors, flippers didn’t appear for another 200 million years.” At this broad time scale, mammals were worse at making flippers, and it rarely happened. While flippers evolved some eight to ten times in reptiles, they only evolved four times in mammals. In part because of their flippers, some aquatic reptiles evolved to become massive creatures that could now prey upon the biggest sharks.
To thrive on land, one group of reptiles managed to shape their skulls to catch big prey,” Caleb says with a smile, “imagine, a T. rex, the biggest meat-eating animal that ever walked the Earth, or its relative, the modern-day saltwater crocodile, which has one of the strongest bites among all living animals today.” To become terrifying carnivores, terrestrial reptiles needed to open their jaws wide, snap it shut quickly, and bite down hard, destroying their prey before it escaped. Caleb demonstrates this by cupping his hands open and shut like a clamshell. But nature gives and takes, and powerful jaws tend not to open as wide or bite down as fast, while nimble and lightweight jaws open wide, but usually fail to bite down as hard. Reptiles managed to overcome these limitations, Caleb explains, by “changing the architecture of their jaw muscles.” Shortly before the Great Dying, some transformed their tiny deep jaw muscles into massive ones that could open wider while maintaining a powerful bite. These reptiles were the ancestors of dinosaurs. They ate bigger prey, became fearsome land predators, and reached a new level of carnivory.
Reconstructing such details of a distant past is not easy. “Studying the evolution of ancient reptiles is tricky. We rely on scarce fossil records they left behind, and the vast majority of them left no trace at all.” To make these discoveries, Caleb does some of the classic paleontology work you may envision from the movies. He sifts through ancient bones at the Yale Peabody Museum and precisely records lengths, widths, heights, and depths. With these, he built the largest dataset of its kind: around 16,000 measurements from over 800 museum specimens, representing both living and extinct species, across hundreds of millions of years of evolution.
Caleb Gordon collecting data at the Yale Peabody Museum. Here, he measures an arm bone of an ancient marine reptile that lived during the Jurassic Period, over 140 million years ago.
“But these classic methods can only get you so far. To understand all of these fossils, and all this information, we need to use modern techniques and technologies.” He feeds his masses of data into a machine-learning algorithm to compare competing stories of whether extinct species occupied land, water, or both, and predict which scenario was most likely. Given the nature of the data, Caleb had to introduce a statistical method first devised by the US Navy, and almost never used to study fossils. “We also adapt 3D computer animation programs, the same software used by Pixar and Marvel Studios, to ‘repair’ and ‘un-crush’ fossils so that they look more like they would have when the animal was alive,” he explains. These approaches let us recraft ancient structures that are unseen in modern life. With these methods, Caleb can tell which past aquatic and terrestrial reptile stories are likely true.
3D skull segmentation in action.Caleb segments a skull bone of a Euparkeria, a close relative of crocodilians and dinosaurs.
Caleb’s work recreates scenes from prehistoric times of life forms fighting for their existence. His PhD advisor, Dr. Bhart-Anjan Bhullar, emphasizes, “Caleb’s research represents a series of fundamental advances in our understanding of the lives of ancient organisms.” These ancient organisms, the diverse family of reptiles, flew, crawled, swam, and roamed. Through luck and remarkable evolutionary innovations, they managed to conquer two vastly distinct environments and replace our ancestors at the top of the global food chain…at least for a while.
This work was supported by training from Carl Zimmer, and feedback from Zili Shen at the Yale University Graduate Writing Lab.
 (No Ratings Yet)
(No Ratings Yet)









 (5 votes)
(5 votes)