The Tata lab in the Department of Cell Biology, Duke University School of Medicine has two openings for postdoctoral researchers to study cellular plasticity mechanisms in lung injury repair and tumorigenesis. We seek to understand the genetic and epigenetic basis of organ regeneration and tumorigenesis. We study the properties of stem/progenitor cells in diverse epithelial tissues (with a primary focus on lung) and their relationships with neighboring tissues in pathophysiological conditions. We utilize in vivo mouse genetics, live imaging, 3D organoids, genome-wide Cas9/Crispr based functional genetic screening, and next generation sequencing technologies to study the behavior of tissues at single cell level. We offer an inspiring intellectual, collaborative and multidisciplinary research environment to support your career goals and provide access to state-of-the-art facilities. Candidates with background knowledge and hands-on experience in transcriptional regulation, 3D-organoids and bioinformatics skills are particularly welcome.
Requirements
– A PhD or MD/PhD (or equivalent) in biological sciences (cell & developmental biology or a related field).
– Strong research background in transcriptional regulation, cell biology, molecular biology, mouse models of cancer, and/or biochemistry. Prior experience in stem cells, Cas9/Crispr gene editing, 3D-organoid cultures, chromatin biology and bioinformatics is advantageous.
– Evidence of successful completion of a research project (publications)
– Ability to work independently; interpret, present and discuss experimental data
– Excellent communications skills
My mentor, Bruce Appel, emphasizes the importance of communicating science clearly and precisely. Consequently, I have watched my peers and myself deliver ever-improving talks, posters, and manuscripts during our time in the lab. I think that many people in science appreciate that clear communication is essential for others to be able to interpret findings and effectively build upon what has been done. A corollary lesson that I wasn’t expecting to learn from our latest project, published recently in Nature Neuroscience (Hughes and Appel, 2020), is how imprecise language can muddle and confound understanding, obstructing progress.
Our lab studies oligodendrocyte lineage cells using zebrafish as a model system. In the central nervous system, oligodendrocytes wrap neuronal axons with myelin, a lipid-rich membrane that increases conduction velocity. Zebrafish are both genetically and optically accessible, allowing us to image cellular interactions during myelination in live larvae. A live-imaging experiment can catch oligodendrocytes extending numerous, arborizing processes that search for and begin to wrap axons with myelin membrane. Some of these nascent wraps continue to grow and mature, whereas other wraps appear to be eliminated. How are myelin sheaths eliminated?
Many structures that also are studied by live-imaging, like neuronal neurites, undergo similar deformations during development. Neurons elaborate and occasionally withdraw neurites, and this latter process is termed “retraction”. Oligodendrocytes generate processes that branch similarly to neurites. Like neurites, these processes withdraw, and these events have been described as “retraction”. But oligodendrocyte processes also do something very different than neurites: upon contacting a target axon, an oligodendrocyte process can deposit a reservoir of myelin lipids that spreads like a liquid droplet as the process begins wrapping the axon (Nawaz et al., 2015). Can fluid-like myelin sheaths, like the simple processes that gave rise to them, also be withdrawn? Live confocal microscopy doesn’t show us sheaths unraveling or processes reeled in by the cell body. Instead, sheaths merely reduce in size and disappear. By using the word “retraction” to describe all oligodendrocyte process disappearances, an untested mechanism was invoked to account for all myelin sheath elimination.
If myelin sheaths aren’t retracted, what alternative mechanisms could remove sheaths? Many structures in the developing nervous system, including synapses, neuronal precursors, and neurons, are overproduced and can be pruned by microglia, the resident immune cell type of the CNS. We had previously found a number of similarities between the formation of myelin sheaths and neuronal postsynapses (Hughes and Appel, 2019), raising the possibility that these structures might also be eliminated similarly.
Admittedly, I spent a lot of time thinking about microglia before they constituted a reasonable suspect in sheath elimination. Early in grad school, I had read a paper that found that microglia regularly survey the zebrafish spinal cord and quickly swoop in to clear laser-ablated neurons (Morsch et al., 2015) and I was really curious to see if these cells interact with oligodendrocytes during normal development. Something I particularly like about working with zebrafish is how easy it is to casually pursue these types of side-curiosities. I lost no time to generating the construct to label microglia, because I did it in parallel with other cloning I needed to do; I injected the construct to generate a microglia reporter transgenic line after I had performed my priority injections for experiments that week. A few months later, I had a microglia reporter line and was ready to find out if microglia and oligodendrocytes interact during myelination.
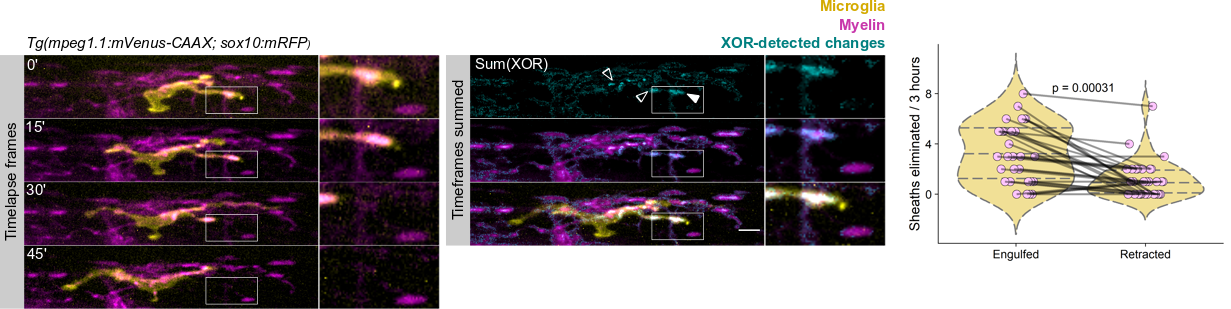
A microglia (yellow) engulfing nascent myelin sheaths (magenta) in the spinal cord of a zebrafish. Scale bar, 10 um.
The first time I timelapse imaged microglia interacting with myelin, to my surprise and delight I found microglia engulfing nascent myelin sheaths. I presented a video at lab meeting and was encouraged by the enthusiasm and questions raised by my labmates. How many microglia are there and how many sheaths are they eating? Do oligodendrocytes die when their sheaths are eaten? What regulates sheath phagocytosis? These tractable questions started to crystallize the phenomenon into a project.
At this stage, our recognition that myelin sheath development shares numerous similarities with synapse development was pivotal. Work by Dorothy Schafer, Beth Stevens, and Marie-Ève Tremblay had previously shown that microglia contact and phagocytose synapses in a neuronal activity-regulated manner (Schafer et al., 2012; Tremblay et al., 2010). Inspired by this work, the hypothesis and predictions that would form the foundation of the project emerged by analogy. Like synapses, do microglia phagocytose myelin in an activity-regulated manner? Will preventing pruning cause extra myelin to persist? With a path laid out, experiments moved swiftly. We found that microglia do survey and phagocytose myelin sheaths, and neuronal activity spurs microglia to trade-off between interacting with neuronal somas and phagocytosing myelin from myelinated axons. We further found that this program is robust enough that blocking it (via microglial ablation) caused excessive and ectopic myelin to develop.
We wrote up and submitted a much shorter version of our paper, concurrent with deposition to the preprint server BioRxiv, in summer 2019. In review, it became clear that microglia-mediated myelin pruning was incompatible with our field’s understanding that sheaths are solely eliminated by retraction. At first, I saw this as a purely semantic problem: myelin sheaths disappear, and the word “retract” has been used to describe this observation but oversteps into a mechanism that hasn’t been tested. I was resistant to confronting retraction but was persuaded by a reviewer’s argument that it would be informative to know what fraction of sheath elimination is contributed by microglia. Figuring out how to quantify sheath loss in an unbiased way took longer than any other experiment in the paper and pushed my image analysis forward in new ways. These data were the very last addition to the paper, but they appear inconspicuously in the middle of Figure 2! Ultimately, we found that sheaths are both phagocytosed by microglia and disappear independently of microglial contact, with phagocytosis accounting for the majority of sheath loss. This latter, microglia-independent category of lost sheaths might be called “retraction”, as we do in the paper.
How many sheaths are eliminated by microglia? Method to detect changes in myelin over time (cyan) allowed us to determine how many disappearing sheaths are engulfed vs disappear without microglia contact (“retract”). Arrowhead marks an engulfed sheath; open arrowheads mark engulfed myelin shuffling within microglia. Scale bar, 10 um.
I still have some reservations about “retraction”. Prior to labeling microglia, we accepted that sheaths were solely eliminated by retraction: when oligodendrocytes were the only cell type visible, it was easy to grant cell autonomy to all visible changes and to forget that other cell types are lurking in the dark. Similarly, it’s possible that additional unlabeled cell types might engulf the microglia-independent subset of disappearing sheaths.
References
Hughes, A. N. and Appel, B. (2019). Oligodendrocytes express synaptic proteins that modulate myelin sheath formation. Nat. Commun.10, 4125.
Hughes, A. N. and Appel, B. (2020). Microglia phagocytose myelin sheaths to modify developmental myelination. Nat. Neurosci. in press.
Morsch, M., Radford, R., Lee, A., Don, E. K., Badrock, A. P., Hall, T. E., Cole, N. J. and Chung, R. (2015). In vivo characterization of microglial engulfment of dying neurons in the zebrafish spinal cord. Front. Cell. Neurosci.9, 321.
Nawaz, S., Sánchez, P., Schmitt, S., Snaidero, N., Mitkovski, M., Velte, C., Brückner, B. R., Alexopoulos, I., Czopka, T., Jung, S. Y., et al. (2015). Actin Filament Turnover Drives Leading Edge Growth during Myelin Sheath Formation in the Central Nervous System. Dev. Cell34, 139–151.
Schafer, D. P., Lehrman, E. K., Kautzman, A. G., Koyama, R., Mardinly, A. R., Yamasaki, R., Ransohoff, R. M., Greenberg, M. E., Barres, B. A. and Stevens, B. (2012). Microglia Sculpt Postnatal Neural Circuits in an Activity and Complement-Dependent Manner. Neuron74, 691–705.
Tremblay, M.-È., Lowery, R. L. and Majewska, A. K. (2010). Microglial Interactions with Synapses Are Modulated by Visual Experience. PLoS Biol.8, e1000527.
The University of Lyon is a worldwide academic site of excellence. Labelled IDEX in 2017, it is located in the heart of the Auvergne-Rhône-Alpes region, in the Lyon Saint-Étienne basin. Structured around 12 members, the University of Lyon has three major ambitions:
· To design a large, attractive, responsible university with a reputation for excellence and innovation and a strong international reputation;
· Propose a training and research program of excellence, in line with the expectations and changes in society;
· Develop and enhance the dynamics of the Lyon Saint-Étienne site, in conjunction with all the players in the area: citizens, associations, businesses, local authorities (Lyon and Saint-Étienne metropolitan areas, Auvergne-Rhône-Alpes Region, other local authorities).
The University of Lyon is looking for a research engineer (IR) in bioinformatics for the LabEx CORTEX. The LabEx CORTEX brings together 22 research teams in Neuroscience who conduct multiscale research, from molecular and cellular bases of neuronal physiology to cognitive functions in normal and pathological contexts. It is an exciting international multidisciplinary research environment. The LabEx activities are divided according to three axes: research, education and dissemination.
JOB DESCRIPTION
A full-time position is available for a period of 2 years (renewable) to establish a bioinformatics platform within the LabEx CORTEX of the University of Lyon.
We are seeking highly motivated candidates with a PhD in biological sciences (ideally in Neurosciences) and strong experience in Bioinformatics, in particular with next generation single cell RNA-Sequencing analysis. The candidate will have a published track record within this field and strong knowledge of up to date bioinformatics tools. She/he will have good communication skills in French and English, strong capacity to develop networks (e.g. a previous experience in managing a platform or a common facility). Finally, the candidate will have the task of developing/establishing new protocols and teaching/sharing them to the CORTEX community.
The selected candidate will work closely with local genomic facilities in order to lay the foundation of a bioinformatics platform to assist LabEx CORTEX teams in performing single cell RNA-Sequencing experiments. Hence, she/he will be involved in:
Facilitating the planning of new single cell RNA-Sequencing experiments.
Assisting teams in the analysis of currently available datasets, either directly or by the training/supervision of students/staffs from the participating teams.
Establishing new techniques or procedures of general interest, e.g. establishment and comparison of adult CNS tissue dissociation protocols… gradually evolving to integrate complementary datasets (e.g. Chip-Seq, Slice-Seq)
PROFILE
Competences:
– PhD in biology/neurosciences
– Expertise in bioinformatics analysis
– Advanced expertise in data collection, processing and analysis
– Good knowledge of programming languages (R, Python…)
– Technical English
– Teaching and knowledge transfer competences
– Build and manage databases
Soft skills:
– Excellent communication skills
– Proactive, autonomous and versatile
– Strong ability to work in a team
APPLICATION
Send resume, reference (name, email, address, phone) and application letter to jennifer.beneyton@inserm.fr
A postdoc position is available within the Conduit lab at the Institut Jacques Monod in Paris. See job advert attached and get in touch if you’re interested in studying microtubule nucleation in Drosophila (paul.conduit@ijm.fr).
In this episode, Kat Arney takes a look at the world of epigenetics to find out if more than DNA passes on to the next generation, whether Darwin was wrong and Lamarck was right, and how to pimp your genome. Plus, we meet the Mickey Mouse Mice – a strange (but cute!) example of transgenerational epigenetic inheritance.
If you enjoy the show, please do rate and review on Apple podcasts and help to spread the word on social media. And you can always send feedback and suggestions for future episodes and guests to podcast@geneticsunzipped.com Follow us on Twitter – @geneticsunzip
Happy to share the latest manuscript by André Castro @castro_neuro from the @HCuntz with @GTavosanis labs! We show how a temporal arrangement of stochastic developmental processes achieves efficient dendritic trees both in terms of wire and function. #Drosophila#comp_neuro
One PhD student and one postdoc position will be available in the @GTavosanis lab starting from September 2020.
The two planned projects will focus on:
The cellular mechanisms that support the differentiation of neuronal dendrites (see Stuerner et al., Development 2019). This project will combine the generation of molecular tools for acute manipulation of protein activity with high-resolution in vivo microscopy. We will closely cooperate with neuronal morphology modelling groups.
The circuit mechanisms that support the consolidation of memories in the adult fly brain. Combining detailed anatomical insight and refined genetic tools to manipulate identified neurons, you will investigate how experience and learning modify a circuit’s output. This project is embedded within the frame of the DFG-funded Research Unit FOR 2705 that includes highly interactive groups with complementary expertise (https://www.uni-goettingen.de/en/601524.html ).
All infos and application: https://jobs.dzne.de/en/jobs/50580/phd-student-fmd-and-postdoc-fmd-182520206-182620206
Alistair McGregor’s group at Oxford Brookes University uses Drosophila and the common house spider Parasteatoda tepidariorum to understand how different shapes and sizes of animals evolve. Back in 2016, two PhD students in Alistair’s lab shared a day in the life of a spider lab.
Four years and one pandemic later, we caught up with Alistair to find out how his lab has responded to the ongoing Covid-19 crisis.
‘Normal’ spider care
The lab’s spider culture was originally founded from spiders collected from a basement in Göttingen, Germany. The spiders are fed twice a week, Mondays and Fridays, and mated females get an extra feed on a Wednesday. Females get crickets while males and juveniles are given flies.
In normal circumstances, lab members collect cocoons from females, which are produced every four days. There are usually hundreds of embryos per cocoon. Developing embryos can be kept in halocarbon oil so that the team can identify the exact stage of embryogenesis. This is key for the different approaches the lab uses to study development. It takes about 10 days for the embryos to develop into translucent, hairless, immotile hatchling spiders. Once the juveniles have progressed through several moults and eaten some flies, and siblings, they are separated into individual vials. It takes several weeks after hatching for the spiders to reach reproductive maturity.
The return of the Spider Lab
Over Zoom, we spoke with Alistair to find out about his lab’s lockdown and the return to the bench.
We have made a safe plan.
“People are going in on Mondays to Fridays in shifts. There are three shifts a day and we have divided the lab up into different zones. In the first phase, there will be one person in each zone. As we move forwards I hope we can put more people in the labs while maintaining social distancing. The teaching labs are available over the summer, so that has given us a bit more space to use!”
We had reached the stage where people were keen to return.
“The rest of the UK is opening up to some extent, but we want people to feel comfortable and safe. Therefore the university has permitted researchers to voluntarily return to the research labs to carry out work that cannot be done at home. To avoid using public transport we have allowed people to drive in and use the car park even if they do not have a permit.
The university allowed essential maintenance during lockdown, so some people could go in and cook fly food or flip the stocks. There were three major Drosophila flips during lockdown, so that kept us going. As for the spiders, they are not dangerous or genetically modified, and so one of my postdocs took them home! They are happily back in the lab, but she was feeding them at home for a few weeks.”
We had two new PhD students do their rotations during lockdown.
“They were expecting lab work but they got bioinformatics projects instead! They were both successful though and have resulted in two submitted papers. We have an annual monitoring process for PhD students where they have to submit a report every summer, so my students also worked on that.”
We are trying to prioritise access for final year PhD students.
“There might be a second lockdown, of course we have no idea. That is why we have started to go back with a conservative plan. We are trying to prioritise access for final year PhD students so that they can finish vital experiments.”
Morale has stayed high!
“The lab is a positive bunch. We still had meetings and journal clubs, and a virtual pub quiz every week. At the start I think we were wondering what we could do at home, but then it became overwhelming because there was so much that could be done!”
We’d love to hear how any of our ‘A day in the life’ labs are doing and how you’re returning to the bench, so please do get in touch if you have a story to share!
My university has reopened and being reunited with the lab bench after months have reminded me again how much I love what I do for a living. Of course I enjoy the entire process of research from bench to publication, troubleshooting experiments and brainstorming ideas to presenting research stories. However, hands-on working is something I relish the most. It has a tangible and stimulatory effect on my experimental thought process and in general how I perceive and approach scientific questions.
Staying home for more than three months has been the largest break in my life since kindergarten and has affected my mental wellbeing. I had moved to Boston as a postdoc to study how forces shape tissue patterning and morphogenesis in zebrafish, after finishing PhD in single-molecule plant development in Tübingen, Germany. However, within two months of my arrival in Boston, Harvard stopped all the non-essential research due to soaring coronavirus pandemic.
Living pretty much alone in house arrest in a new city away from my partner and family has been extremely stressful and emotionally difficult. The new lab has been very supportive and connected throughout but the isolation – paired with the rejection from the postdoctoral fellowship I applied for – has given me an imposter syndrome. I felt the sadness growing, my moods shifting, confidence losing, and fear building in me. For example, during scientific zoom meetings with the lab and the department, I was not able to communicate my thoughts and ideas properly because being out of the lab and staying in isolation was toying with my state of mind. Of course, afterwards I then dealt with the guilt of it. The murder of George Floyd further added frustration and anger about the world we live in and how we treat each other. I could feel and acknowledge my worsening mental health. Therefore, I constantly talked about my feelings with my partner and my best friend who have been incredibly supportive irrespective of our separation by the continents. Although communication helps, I felt I still need tangible positivity to pull me out of the mental black hole. So, I have been taking time to reflect on positive things. For example, I have been excitedly rethinking about the scientific problems I am interested in and ways to approach them differently. And besides science, reading literary and historical fiction and baking cakes and pastries. I am sure I am not alone in experiencing such mental instability in this time. Each one of us is experiencing stress and anxiety in our own ways. What is most important is to acknowledge the symptoms and seek the help and support we need.
When I heard the university was reopening and we could go back to the lab, I was thrilled. This is exactly the positivity I think I have been longing for. The flipside to this positivity is that I was nervous about going back. I joined a zebrafish developmental and systems biology lab and had learned how to do crossings and microinjections in zebrafish right before the shutdown. Obviously, I have been anxious that I had forgotten the newly learned methods and whether I would be able to find reagents and consumables in the lab.
Now that I am in the lab again, of course, it is all the same as I knew it, physically. Except that the labs, corridors, hallways, bathrooms, meeting points, and common areas are labelled with safety distance reminders and are awfully silent and empty. There are no social hours or physical meetings. It feels like working late over Christmas. Even fish in the lab probably miss the hustle-bustle of colleagues. I, on the other hand, am delighted to be back in the lab and to see a few familiar masked faces around. Foremost, it is getting me back into the workday routine and breaking the monotony of alone and ‘relaxed’ working from home. More importantly, I am thrilled to be working on the bench again. I think there is an eternal joy that we wet-lab scientists find in bench work. To get back into the familiar working spirit, I resumed by molecular cloning experiments, which I am most comfortable with. Seeing bacterial colonies on the agar plate the day after bacterial transformation have already started to diminish my feelings of being an imposter. Turns out, I can still cross zebrafish and pull needles for microinjections. I am starting to find purpose again, after all work is the reason I moved to the US.
Happy to be back at the bench although atmosphere in the lab and institute is not the same as pre-CoViD era.
Although we are working in spatially and temporally coordinated lab shifts, the motivation to be in the lab and conduct experiments is more than enough for me to be feeling mentally healthy again. I understand it is not the ideal situation for any of us. But neither is the uncertainty of normality. I think by embracing little steps towards normality, we can overcome the mental hurdles posed by the pandemic. I realise my mental clock has started to shift towards normal: being back at the bench is definitely the sweeter side of our bitter-sweet labcoming experience.
DanStem addresses fundamental research questions in stem cell and developmental biology and has activities focused on the translation of promising basic research results into new therapeutic strategies for cancer and chronic diseases. DanStem is a vibrant, internationally diverse and ambitious research center with state-of-the-art facilities. The setting is ideally suited for seamless collaboration and exchange with other centers and departments of the Faculty and Copenhagen science community.
CPR promotes basic and applied research on human proteins of medical relevance. CPR spans broad areas of protein research, ranging from structural and mechanistic understanding of proteins and their cellular functions in chromatin biology and genome maintenance, to development of clinical applications of proteomics and systems biology. The vision of CPR is to combine the power of integrative protein technologies and their application to accelerate understanding of the biological processes underlying health and disease.
The genomics platform was established in 2017 by DanStem and CPR and currently employs a sequencing specialist and a bioinformatics specialist. We are seeking a highly motivated and ambitious candidate to join the platform and replace our current sequencing specialist.
Job description
The role of the next generation sequencing (NGS) specialist is to ensure daily management of the Genomics Platform. The tasks include: training and assistance on library preparation, running the sequencer, training the users on platform instruments (e.g. Fragment Analyzer, liquid handling robot), providing advice on genomics tools and troubleshooting. It also includes ordering, stocking and supplying consumables. In addition, the applicant will be expected to push forward new genomics applications in collaboration with users.
Your profile
A degree in biotechnology, molecular biology, or appropriate biological or biomedical science with either PhD or MsC.
Theoretical and practical knowledge of NGS sample preparation workflows and applications.
Previous experience with NGS bioinformatics is a plus.
Good English communication skills, both oral and written, are required.
We offer
stimulating, challenging and multifaceted research environment
possibility for continued education and training
attractive employment conditions
central located workplace.
Employment conditions
The employment is a permanent position and is scheduled to start in September 2020 or upon agreement with the chosen candidate. The place of work is at DanStem, University of Copenhagen, Blegdamsvej 3B, Copenhagen. Salary, pension and terms of employment are in accordance with the provisions of the collective agreement between the Danish Government and AC (the Danish Confederation of Professional Associations).
In addition to the basic salary, a monthly contribution to a pension fund is added (17.1% of the salary).
The employment will be as Research consultant (Specialkonsulent) depending on the selected candidate’s experiences and qualifications and the working time are 37 hours per week on average
Application
Your application must be submitted in English by clicking “Apply now” below and must include:
Motivation letter
Curriculum vitae incl. education, experience, previous employments, language skills and other relevant skills
Copy of diplomas/degree certificate(s).
Only applications received in time and consisting of the above listed documents will be considered. Applications and/or any material received after the deadline will not be taken into consideration.
Deadline for applications: Sunday 9 August 2020, 23:59 p.m. CET.
Questions
For further information about the position, please contact Professor Joshua Brickman by e-mail: joshua.brickman@sund.ku.dk.
The Faculty of Health and Medical Sciences comprises approximately 7,850 students, 1,700 PhD students and 4,800 employees. The Faculty advances the field of health sciences through its core activities: research, teaching, knowledge sharing and communication. With basic research fields ranging from molecular studies to studies of society, the Faculty contributes to a healthy future through its graduates, research findings and inventions benefitting patients and the community. The University of Copenhagen wishes to reflect the surrounding community and invites all regardless of personal background to apply for the position.
 (No Ratings Yet)
(No Ratings Yet)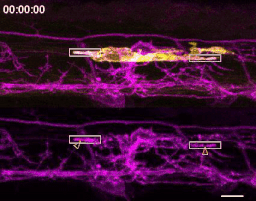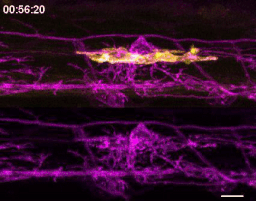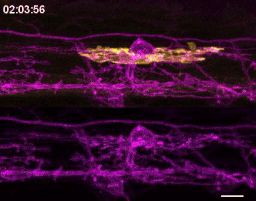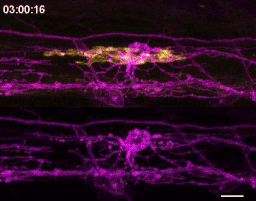
 (4 votes)
(4 votes) In this episode, Kat Arney takes a look at the world of epigenetics to find out if more than DNA passes on to the next generation, whether Darwin was wrong and Lamarck was right, and how to pimp your genome. Plus, we meet the Mickey Mouse Mice – a strange (but cute!) example of transgenerational epigenetic inheritance.
In this episode, Kat Arney takes a look at the world of epigenetics to find out if more than DNA passes on to the next generation, whether Darwin was wrong and Lamarck was right, and how to pimp your genome. Plus, we meet the Mickey Mouse Mice – a strange (but cute!) example of transgenerational epigenetic inheritance.

 (11 votes)
(11 votes)