Research assistant position for subsequent appointment as PhD fellow in stem cell biology and epigenomics
at DanStem, Faculty of Health and Medical Sciences
University of Copenhagen
The Danish Stem Cell Center (DanStem) at Faculty of Health & Medical Sciences at the University of Copenhagen is looking for a Research assistant subsequent appointed as PhD fellow to join the Żylicz group starting June 2020 or upon agreement with the chosen candidate.
The position as Research assistant is for 1 year. The position as PhD fellow is for 3 years.
This position is available in the group of Jan Żylicz at DanStem. The team studies fundamental mechanisms of early mouse development and stem cell biology. The Żylicz group wants to understand how metabolic and epigenetic mechanisms cooperate to regulate transcription during early development. In particular the team is interested in how metabolism regulates histone modifiers, and how these in turn affect lineage choice and embryo growth at around the time of implantation. To achieve this, the group utilizes both in vivo mouse models as well as in vitro stem cell culture systems and state-of the-art ultrasensitive transcriptomic, epigenomic and metabolomic techniques. This research project will employ a multi-disciplinary approach to understand how first lineage choices are influenced by metabolic and chromatin states. Starting date
for this position is the 1 July 2020, or upon agreement with the chosen candidate.
Job description:
We are seeking a highly motivated and ambitious pre-doctoral candidate with a strong educational background in either stem cell and developmental biology and/or metabolomic research. The candidate will investigate the molecular mechanisms controlling first lineage choice of early development using both in vivo and in vitro stem cell models. By employing genomic engineering methods the candidate will go beyond description of epigenetic and metabolic states and uncover novel functional regulatory mechanisms at the interplay of epigenetics and metabolism.
About DanStem
The Novo Nordisk Foundation Center for Stem Cell Biology – DanStem – addresses basic research questions in stem cell and developmental biology and translates results from basic research into new strategies and targets for the development of new therapies for diabetes and cancer. DanStem was established as a result of a series of international recruitments coupled with internationally recognized research groups focused on insulin producing beta cells and cancer research already located at the University of Copenhagen.
Qualifications:
Candidates must hold a master’s degree in biotechnology, bio-informatics, biology or similar, and possess a general understanding of developmental and stem cell biology and/or metabolomics.
Previous practical experience in bioinformatics, analysis of epigenomic data and/or software programming is considered of great advantage.
Practical experience in working with stem cells or mouse models is considered of great advantage.
Practical project experience in basic lab techniques is considered beneficial.
Good English communication skills, both oral and written, are prerequisite for the successful candidate
Terms of salary, work, and employment :
The employment is for 4 years, as research assistant is for 1 year and as PhD fellow for the following 3 years, and is scheduled to start in July or upon agreement with the chosen candidate. The employment as a PhD student is conditioned upon a positive assessment of the candidate ́s research performance and enrolment in the Graduate School at the Faculty of Health and Medical Sciences.
The PhD study must be completed in accordance with the ministerial orders from the Ministry of Education on the PhD degree and the University ́s rules on achieving the degree.
The place of work is at DanStem, University of Copenhagen, Blegdamsvej 3B, Copenhagen. Salary, pension and terms of employment are in accordance with the provisions of the collective agreement between the Danish Government and AC (the Danish Confederation of Professional Associations). In addition to the basic salary a monthly contribution to a pension fund is added (17.1% of the salary).
The application must include:
Motivation letter
Curriculum vitae incl. education, experience, previous employments, language skills and other relevant skills
List of two references (full address, incl. email and phone number)
Copy of diplomas/degree certificate(s)
Questions:
For further information contact Associate Professor Jan Żylicz, jan.zylicz@sund.ku.dk.
How to apply
The application, in English, must be submitted electronically by clicking APPLY below.
The University of Copenhagen wishes to reflect the diversity of society and welcomes applications from all qualified candidates regardless of personal background.
Only applications received in time and consisting of the above listed documents will be considered.
Applications and/or any material received after deadline will not be taken into consideration.
The application will be assessed according to the Ministerial Order no. 284 of 25 April 2008 on the Appointment of Academic Staff at Universities.
Assessment procedure
After the expiry of the deadline for applications, the authorized recruitment manager selects applicants for assessment on the advice of the Appointments Committee. All applicants are then immediately notified whether their application has been passed for assessment by an expert assessment committee. Selected applicants are notified of the composition of the committee and each applicant has the opportunity to comment on the part of the assessment that relates to the applicant him/herself. You can read about the recruitment process at http://employment.ku.dk
Application deadline: 14 April 2020
Founded in 1479, the University of Copenhagen is the oldest university in Denmark. It is among the largest universities in Scandinavia and is one of the highest ranking in Europe. The University´s eight faculties include Health Sciences, Humanities, Law, Life Sciences, Pharmaceutical Sciences, Science, Social Sciences and Theology. www.ku.dk
Part of the International Alliance of Research Universities (IARU), and among Europe’s top-ranking universities, the University of Copenhagen promotes research and teaching of the highest international standard. Rich in tradition and modern in outlook, the University gives students and staff the opportunity to cultivate their talent in an ambitious and informal environment. An effective organisation – with good working conditions and a collaborative work culture – creates the ideal framework for a successful academic career.
An interdiciplenary collaboration of scientists from the Novo Nordisk Foundation Center for Stem Cell Biology and animators in Denmark is aiming to facilitate a dialogue between the two diciplines, and ultimately disseminate stem cell research to different segments of the society.
The primary objectives are:
To train scientists to pitch scientific stories to non-scientists, and in particular to visual story-tellers.
To introduce visual story-tellers to stem cell research and to how science is conducted.
To establish a common language between these professionals.
In May 2019, a few scientists from DanStem went to a match-making event in Viborg, and in August the painter and animator Cosimo Miorelli visited DanStem to meet with scientists and platform specialists.
Following these interactions, Cosimo conducted the animation ‘Cells of Knowledge’; a short introduction to stem cells research and to the techniques used in research.
More specilized animations are currently in the pipeline.
The Kim Lab at Molecular, Cell, and Developmental Biology Department, the University of California, Santa Cruz is seeking highly motivated and talented postdoctoral research fellows with a Ph.D. degree to join our new and innovative research group.
The University of California, Santa Cruz is one of 10 universities within the prestigious University of California system. MCD department has top-tier neuroscience and molecular biology labs and UCSC with its genomics institute is an undisputed leader in genomics and bioinformatics. Santa Cruz is a wonderful small progressive town on the central coast of sunny California, nestled into mountains that teem with giant redwood trees, approximately 35 minutes to San Jose/Silicon Valley or 1 hour and half to San Francisco Bay.
The Kim Laboratory aims to investigate connectivity, development, genetic identity, and function of neural circuits using mouse cerebral cortex as a model system. Our ultimate goal is to understand the fundamental principle of neural connectivity and its functions in animal’s perception and behavior. We address our questions using novel neural circuit tracing systems with next-generation trans-synaptic viral tracers, mouse genetics, single-cell genome-wide sequencing, and in vivo imaging. My lab is determined to offer excellent research opportunities to advance your scientific career, strong academic interactions and collaborations across the neuroscience and other biology laboratories at UCSC and more. For additional information, please refer to the following webpage: http://www.ejkimlab.com/
We prefer, but not limited to, candidates with expertise in the following areas: (neuro)developmental biology with genomics experiences, mouse surgery and handling related to neural circuit tracing and manipulations, and molecular and cellular neuroscience. Above all, outstanding applicants with strong quantitative skills are strongly encouraged to apply. Interested individuals should submit an application with a curriculum vitae, a brief cover letter including research interests, and the contact information of three individuals who will provide letters of reference to:
Euiseok Kim, Ph.D.
Assistant Professor
Department of Molecular, Cell, and Developmental Biology University of California, Santa Cruz ekim62@ucsc.edu
The University of California is an Equal Opportunity/Affirmative Action Employer. All qualified applicants will receive consideration for employment without regard to race, color, religion, sex, sexual orientation, gender identity, national origin, age, disability, protected veteran status, or any other characteristic protected by law.
Uri Manor’s primary focus is the integration and application of optical and charged particle detection technologies to study problems of critical biological significance. His current research focuses on developing deep learning-based computational approaches to increase the resolution, sensitivity and speed of the next generation of microscopes, as well as designing nanoprobes for high spatiotemporal resolution imaging of subcellular dynamics.
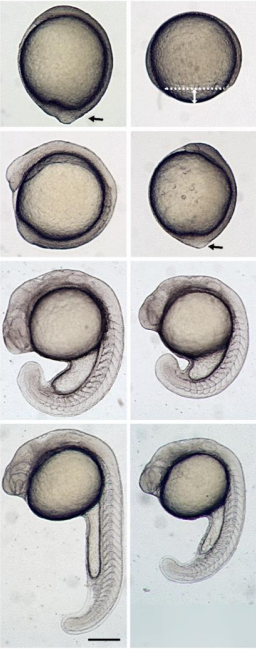
We seek a candidate for a Wellcome Trust-funded Research Associate position working on an interdisciplinary project investigating the role of cell shape in controlling asymmetric cell divisions, fate decisions and tissue morphogenesis. This project is based in the laboratory of Dr Shane Herbert (University of Manchester, UK) and builds upon our recent work revealing that asymmetric divisions fundamentally coordinate tissue morphogenesis during blood vessel formation (Costa. G., et. al. 2016. Nat. Cell Biol. and unpublished work). In particular, we aim to determine how and why acquisition of specific interphase cell geometries modifies the biomechanics of cell division to trigger asymmetric divisions, both during blood vessel formation and development of other tissues. Using in vitro micropatterning to modulate cell shape, omics techniques and live cell imaging approaches, both in vitro and in vivo during zebrafish embryonic development, we aim to define how cell shape globally impacts cellular transcription, translation, actomyosin dynamics, chromosome segregation and DNA damage to drive asymmetric division in diverse tissue contexts in vivo.
The successful applicant will already hold (or have nearly completed) a PhD in a relevant discipline and have experience of either live cell imaging, the mechanisms of cell division, micropatterning tools and/or the zebrafish model system.
The position is available from 1st April 2020 and is tenable for up to 60 months full time (later start dates are negotiable). For more details of the laboratory, please see www.herbertlab.com.
We seek a candidate for a Wellcome Trust-funded Research Associate position working on an interdisciplinary project investigating the roles of subcellular targeting of mRNAs in the spatial control of local cell shape remodelling and tissue morphogenesis. This project is based in the laboratory of Dr Shane Herbert (University of Manchester, UK) and builds upon our recent work revealing that the precise subcellular targeting of mRNAs and resulting local protein translation acts as a molecular compass that drives local cell shape changes, orients motile cell polarity and spatially directs tissue movement during blood vessel formation in vivo (Costa. G., et. al. 2019. bioRxiv). In particular, this project will exploit novel optogenetic tools to achieve light-induced dynamic modulation of mRNA subcellular localisation, both in vitro and in vivo during zebrafish embryonic development, with an aim to define the fundamental mechanisms and function of mRNA targeting in the control of broad aspects of in vivo tissue behaviour.
The successful applicant will already hold (or have nearly completed) a PhD in a relevant discipline and have experience of either live cell imaging, optogenetics, CRISPR gene-editing and/or the zebrafish model system.
The position is available from 1st April 2020 and is tenable for up to 60 months full time (Later start dates are negotiable). For more details of the laboratory, please see www.herbertlab.com.
Informal enquiries can be made to Dr. S. Herbert.
Tel: 0161 275 1140
Email shane.herbert@manchester.ac.uk
Application forms and further particulars can be obtained at http://www.manchester.ac.uk/aboutus/jobs/
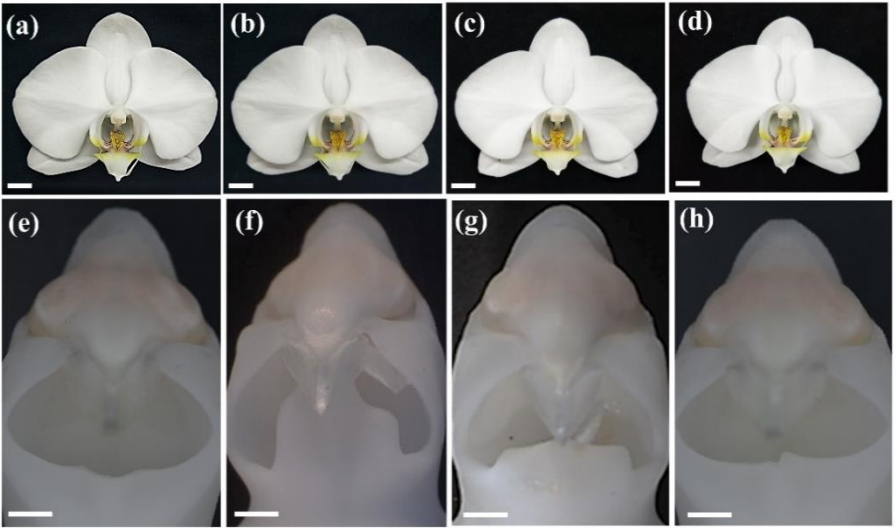
Welcome to our monthly trawl for developmental biology (and related) preprints.
February’s haul features blooming orchids, regenerating lungfish and human intestines, and also lots of interesting stuff in the ‘Research practice and education section’, from preprint impacts to the dominance of the English language in science.
They were hosted on bioRxivandarXiv. Let us know if we missed anything. Use these links to get to the section you want:
LSD1 represses a neonatal/reparative gene program in adult intestinal epithelium
Rosalie T. Zwiggelaar, Håvard T. Lindholm, Madeleine Fosslie, Marianne T. Pedersen, Yuki Ohta, Alberto Díez-Sánchez, Mara Martín-Alonso, Jenny Ostrop, Mami Matano, Naveen Parmar, Emilie Kvaløy, Roos R. Spanjers, Kamran Nazmi, Morten Rye, Finn Drabløs, Cheryl Arrowsmith, John Arne Dahl, Kim B. Jensen, Toshiro Sato, Menno J. Oudhoff
Lgr5+ telocytes are a signaling hub at the intestinal villus tip
Keren Bahar Halpern, Hassan Massalha, Rachel K. Zwick, Andreas E. Moor, David Castillo-Azofeifa, Milena Rozenberg, Lydia Farack, Adi Egozi, Dan R. Miller, Inna Averbukh, Yotam Harnik, Noa Weinberg-Corem, Frederic J. de Sauvage, Ido Amit, Ophir D. Klein, Michal Shoshkes-Carmel, Shalev Itzkovitz
Urethral luminal epithelia are castration-insensitive progenitors of the proximal prostate
Diya B Joseph, Gervaise H Henry, Alicia Malewska, Nida Iqbal, Hannah M Ruetten, Anne E Turco, Lisa L Abler, Simran K Sandhu, Mark T Cadena, Venkat S Malladi, Jeffrey C Reese, Ryan J Mauck, Jeffrey C Gahan, Ryan C Hutchinson, Claus G Roehrborn, Linda A Baker, Chad M Vezina, Douglas W Strand
“Enteric glia as a source of neural progenitors in adult zebrafish”
Sarah McCallum, Yuuki Obata, Evangelia Fourli, Stefan Boeing, Christopher J Peddie, Qiling Xu, Stuart Horswell, Robert Kelsh, Lucy Collinson, David Wilkinson, Carmen Pin, Vassilis Pachnis, Tiffany Heanue
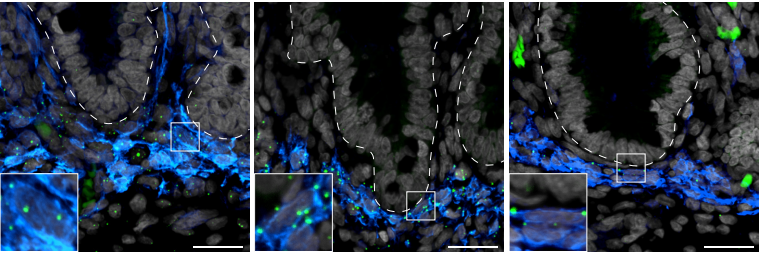
Par3A and Par3B orchestrate podocyte architecture by regulating RhoA levels
Sybille Koehler, Johanna Odenthal, David Unnersjö Jess, Martin Höhne, Christian Jüngst, Ferdi Grawe, Martin Helmstädter, H. Henning Hagmann, Gerd Walz, Wilhelm Bloch, Carien Niessen, Bernhard Schermer, Andreas Wodarz, Barry Denholm, Thomas Benzing, Sandra Iden, Paul Thomas Brinkkoetter
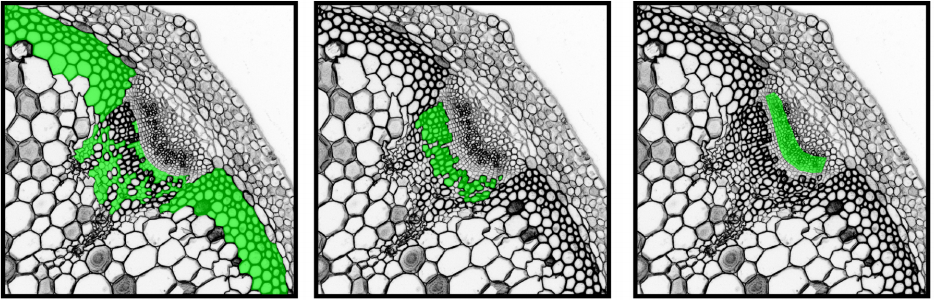
Jeremy Lotto, Sibyl Drissler, Rebecca Cullum, Wei Wei, Manu Setty, Erin M. Bell, Stéphane C. Boutet, Sonja Nowotschin, Ying-Yi Kuo, Vidur Garg, Dana Pe’er, Deanna M. Church, Anna-Katerina Hadjantonakis, Pamela A. Hoodless
Platynereis gene expression/EM overlay from Vergara et al.
Whole-body integration of gene expression and single-cell morphology
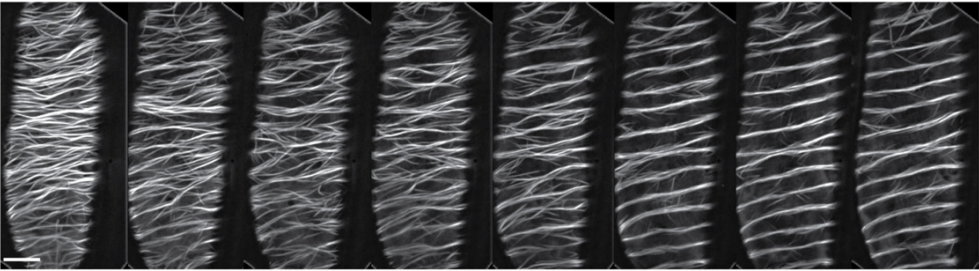
Hernando M. Vergara, Constantin Pape, Kimberly I. Meechan, Valentyna Zinchenko, Christel Genoud, Adrian A. Wanner, Benjamin Titze, Rachel M. Templin, Paola Y. Bertucci, Oleg Simakov, Pedro Machado, Emily L. Savage, Yannick Schwab, Rainer W. Friedrich, Anna Kreshuk, Christian Tischer, Detlev Arendt
Hnrnpul1 loss of function affects skeletal and limb development
Danielle L Blackwell, Sherri D Fraser, Oana Caluseriu, Claudia Vivori, Paul MK Gordon, Amanda V Tyndall, Ryan E Lamont, Jillian S Parboosingh, A Micheil Innes, François P Bernier, Sarah J Childs
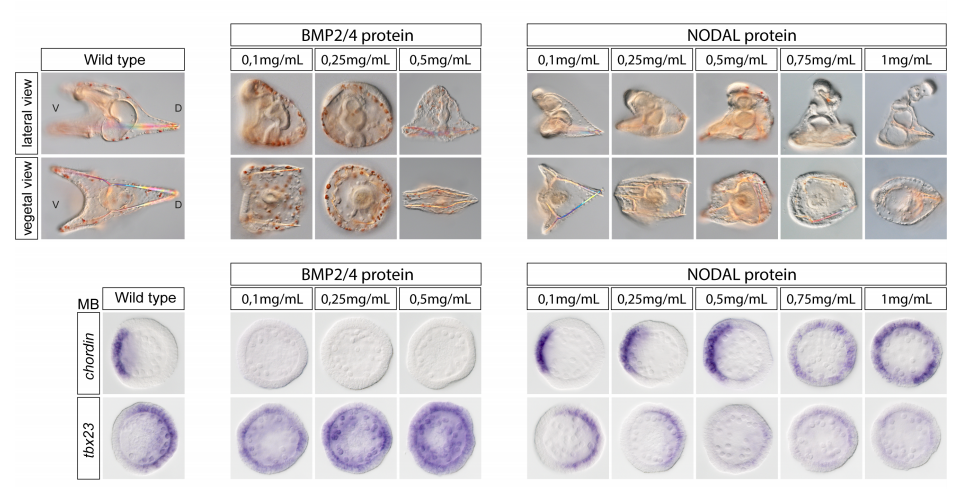
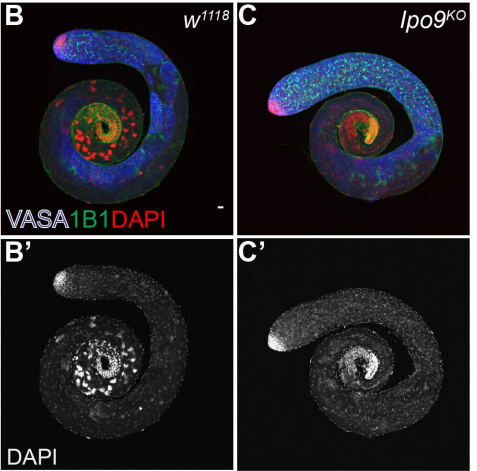
DUX4 regulates oocyte to embryo transition in human
Sanna Vuoristo, Christel Hydén-Granskog, Masahito Yoshihara, Shruti Bhagat, Lisa Gawriyski, Eeva-Mari Jouhilahti, Anastassius Damdimopoulos, Vipin Ranga, Mahlet Tamirat, Mikko Huhtala, Kosuke Hashimoto, Kaarel Krjutškov, Gaëlle Recher, Sini Ezer, Priit Paluoja, Pauliina Paloviita, Yujiro Takegami, Ai Kanemaru, Karolina Lundin, Tomi Airenne, Timo Otonkoski, Juha S. Tapanainen, Hideya Kawaji, Yasuhiro Murakawa, Thomas R. Bürglin, Markku Varjosalo, Mark S. Johnson, Timo Tuuri, Shintaro Katayama, Juha Kere
Smchd1 is a maternal effect gene required for autosomal imprinting
Iromi Wanigasuriya, Quentin Gouil, Sarah A. Kinkel, Andrés Tapia del Fierro, Tamara Beck, Ellise E.A. Roper, Kelsey Breslin, Jessica Stringer, Karla Hutt, Heather J. Lee, Andrew Keniry, Matthew E. Ritchie, Marnie E. Blewitt
PEA15 loss of function and defective cerebral development in the domestic cat
Emily C. Graff, J. Nicholas Cochran, Christopher B. Kaelin, Kenneth Day, Heather L. Gray-Edwards, Rie Watanabe, Jey W. Koehler, Rebecca A. Falgoust, Jeremy W. Prokop, Richard M. Myers, Nancy R. Cox, Gregory S. Barsh, Douglas R. Martin, 99 Lives Consortium
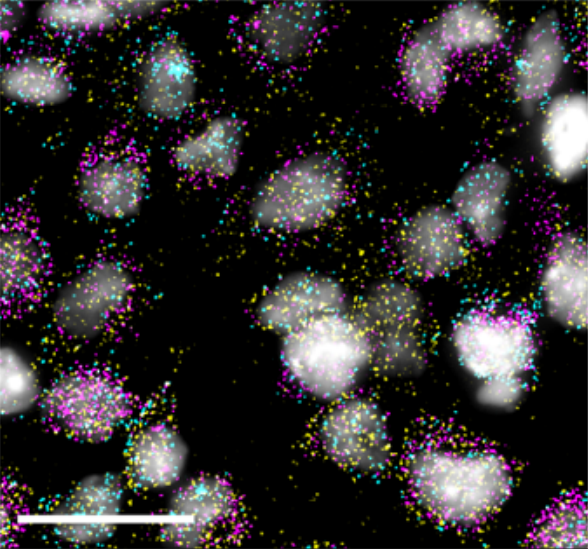
Generation of twenty four induced pluripotent stem cell lines from twenty four members of the Lothian Birth Cohort 1936
Jamie Toombs, Lindsay Panther, Loren Ornelas, Chunyan Liu, Emilda Gomez, Raquel Martín-Ibáñez, Simon R. Cox, Stuart J. Ritchie, Sarah E. Harris, Adele Taylor, Paul Redmond, Tom C. Russ, Lee Murphy, James D. Cooper, Karen Burr, Bhuvaneish T. Selvaraj, Cathy Browne, Clive N. Svendsen, Sally A. Cowley, Ian J. Deary, Siddharthan Chandran, Tara Spires-Jones, Dhruv Sareen
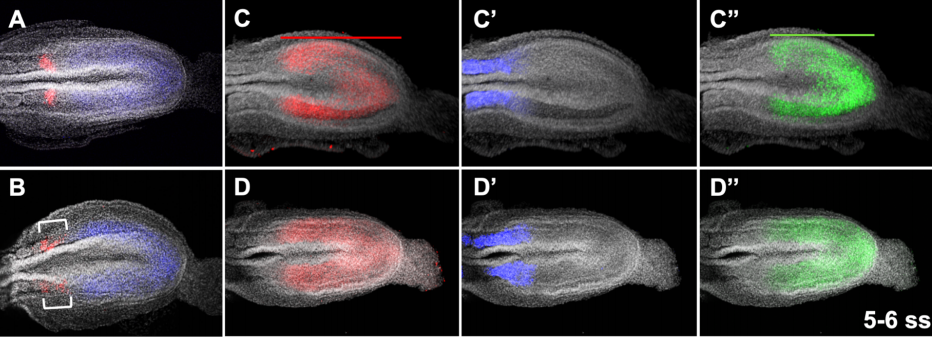
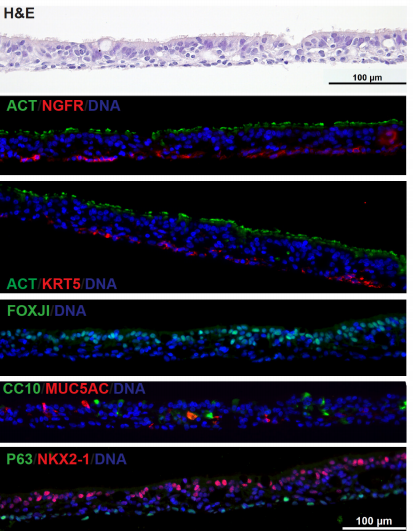
Airway epithelial cells from Hawkins, et al.
Derivation of Airway Basal Stem Cells from Human Pluripotent Stem Cells
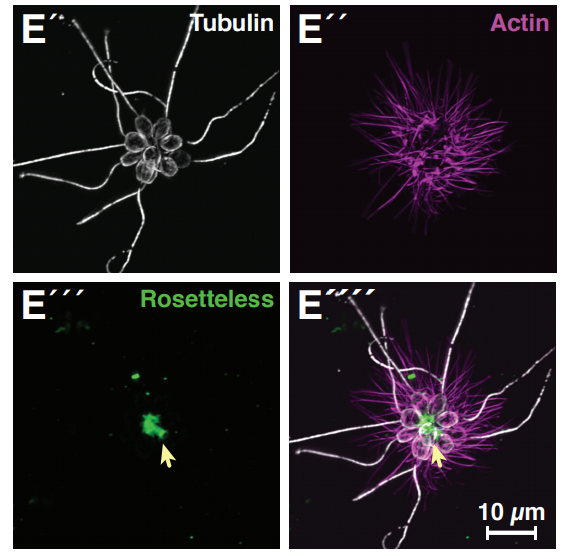
Finn J. Hawkins, Shingo Suzuki, Mary Lou Beermann, Cristina Barillà, Ruobing Wang, Carlos Villacorta-Martin, Andrew Berical, J.C. Jean, Jake Le Suer, Chantelle Simone-Roach, Yang Tang, Thorsten M. Schlaeger, Ana M. Crane, Sarah X. L. Huang, Scott H. Randell, Andras Rab, Eric J. Sorscher, Amjad Horani, Steven L. Brody, Brian R. Davis, Darrell N. Kotton
Neural G0: a quiescent-like state found in neuroepithelial-derived cells and glioma
Heather M. Feldman, Chad M. Toledo, Sonali Arora, Pia Hoellerbauer, Philip Corrin, Lucas Carter, Megan Kufeld, Hamid Bolouri, Ryan Basom, Jeffrey Delrow, José L. McFaline-Figueroa, Cole Trapnell, Steven M. Pollard, Anoop Patel, Christopher L. Plaisier, Patrick J. Paddison
Class III peroxidases PRX01, PRX44, and PRX73 potentially target extensins during root hair growth in Arabidopsis thaliana
Eliana Marzol, Cecilia Borassi, Philippe Ranocha, Ariel. A. Aptekman, Mauro Bringas, Janice Pennington, Julio Paez-Valencia, Javier Martínez Pacheco, Diana Rosa Rodríguez Garcia, Yossmayer del Carmen Rondón Guerrero, Mariana Carignani, Silvina Mangano, Margaret Fleming, John W. Mishler-Elmore, Francisca Blanco-Herrera, Patricia Bedinger, Christophe Dunand, Luciana Capece, Alejandro D. Nadra, Michael Held, Marisa Otegui, José M. Estevez
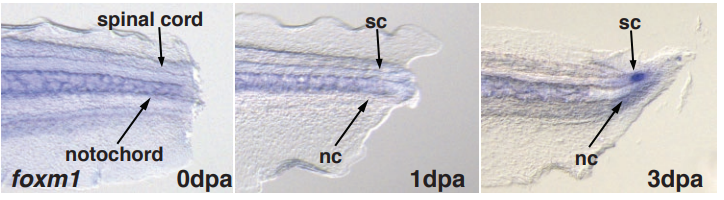
The West African lungfish provides insights into the evolution of tetrapod tail regeneration
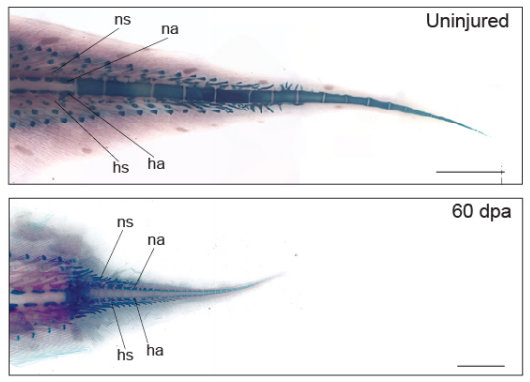
Kellen Matos Verissimo, Louise Neiva Perez, Aline Cutrim Dragalzew, Gayani Senevirathne, Sylvain Darnet, Wainna Renata Barroso Mendes, Ciro Ariel dos Santos Neves, Erika Monteiro dos Santos, Cassia Nazare de Sousa Moraes, Neil Shubin, Nadia Belinda Frobisch, Josane de Freitas Sousa, Igor Schneider
CRISPR-Cas12a-assisted PCR tagging of mammalian genes
Julia Fueller, Konrad Herbst, Matthias Meurer, Krisztina Gubicza, Bahtiyar Kurtulmus, Julia D. Knopf, Daniel Kirrmaier, Benjamin Buchmuller, Gislene Pereira, Marius K. Lemberg, Michael Knop
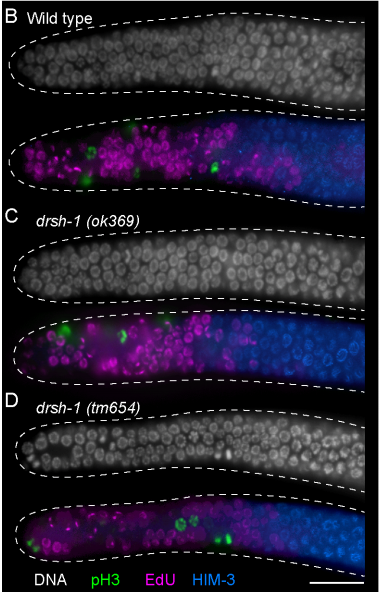
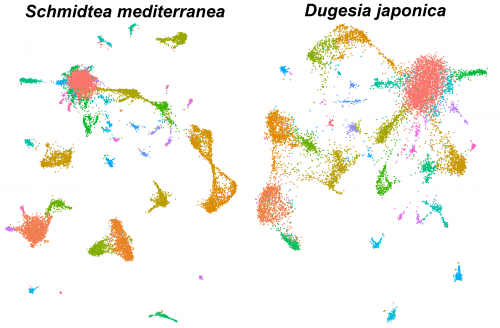
Single-cell analysis of planarian stem cell differentiation and regeneration
The Solana lab seeks to recruit two postdocs (an experimental and a computational researcher) to develop projects on single-cell analysis of planarian stem cell differentiation. These positions are funded by the Leverhulme Trust.
Single-cell sequencing methods are revolutionising the study of stem cells. The flatworm Schmidtea mediterranea is a very promising model for in vivo stem cell biology. They can regenerate thanks to stem cells that continuously differentiate to all adult cell types. We have previously used single-cell transcriptomics to study stem cell differentiation in planarians. Combining clustering algorithms with graph mathematics and RNA metabolism information we were able to reconstruct the differentiation lineage tree of planarian stem cells. We are now using SPLIT-seq to analyse tens of thousands of single cell transcriptomes under different conditions.
The successful applicants will have:
a: experimental) a PhD or equivalent experience in biology, with experience in cell biology and molecular biology, and familiarity with RNA transcriptomics and/or genomic approaches
b: computational) a PhD or equivalent experience in genomic or transcriptomic analysis and must have good computational skills, including experience in R and Unix.
Additionally, the applicants must have organisational and time management skills, and ability to work independently.
You would join an international and growing group of researchers. We are located at Oxford Brookes University. Currently there are two PhD students, one experimental postdoc, two computational postdocs and one technician. There are opportunities to collaborate with other researchers in the UK, Spain, Germany and Italy. The positions are fully funded but the successful applicants will be encouraged to apply for their own funding.
To apply to either the experimental or computational positions, please use the Oxford Brookes University online system following the links. The deadline is March 29th.
The group of Chen Luxenburg at the Faculty of Medicine, Tel Aviv University, invites applications for a Ph.D. student position.
Our laboratory is looking for excellent and highly motivated Ph.D. students to study the role of the actin cytoskeleton in skin development. Our goal is to understand how cytoskeleton derived signals regulate stem cell ability to create the skin epidermis during development, maintain it in the adult, and repair it upon wounding. On top of molecular biology, tissue culture, advanced microscopy, and mouse work, we utilize state of the art technology that allows us to rapidly manipulate the function of any gene of interest in epidermal stem cells in utero. Several exciting projects are available for successful candidates.
We offer state of the art laboratory with a dynamic and international atmosphere and full financial support (tuition and stipend)
Candidates should hold a Master’s degree in Biology/Life-sciences or related fields.
Interested candidates should email their CV and a brief paragraph describing their research experience and career plans to Chen Luxenburg (lux@tauex.tau.ac.il)
The four letters of life – A, C, T and G – are ingrained into the scientific lexicon and burned into the brain of anyone who’s ever worked with or even just learned about genes, genomes and DNA. It’s a code that’s as inseparable from genetics as the double helix itself.
It might therefore be surprising to learn that scientists knew that DNA was made up of these four iconic chemicals – adenine, cytosine, thymine and guanine – long before its double helical structure was figured out in the 1950s.
So when were they discovered? And how did they get their unforgettable names? To find out, we need to go back to the bird poop boom of the 1840s.
If you enjoy the show, please do rate and review on Apple podcasts and help to spread the word on social media. And you can always send feedback and suggestions for future episodes and guests to podcast@geneticsunzipped.com Follow us on Twitter – @geneticsunzip
 (No Ratings Yet)
(No Ratings Yet) (3 votes)
(3 votes)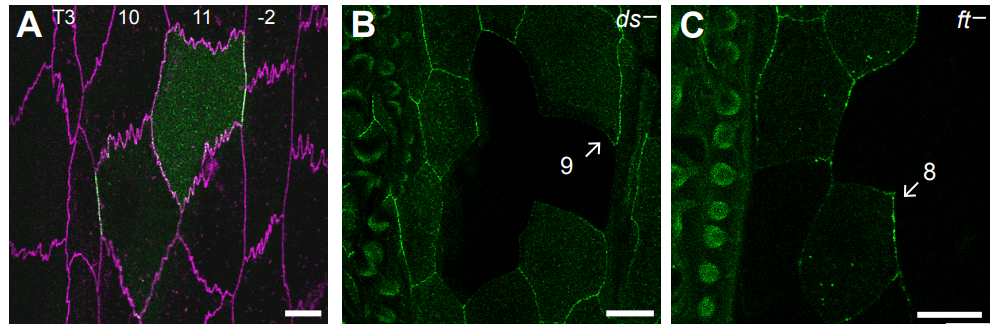


 The four letters of life – A, C, T and G – are ingrained into the scientific lexicon and burned into the brain of anyone who’s ever worked with or even just learned about genes, genomes and DNA. It’s a code that’s as inseparable from genetics as the double helix itself.
The four letters of life – A, C, T and G – are ingrained into the scientific lexicon and burned into the brain of anyone who’s ever worked with or even just learned about genes, genomes and DNA. It’s a code that’s as inseparable from genetics as the double helix itself.