The Company of Biologists (Development’s not-for-profit publisher) is currently seeking proposals for Workshops to be held during 2021.
The Workshops provide leading experts and early-career researchers from a diverse range of scientific backgrounds with a stimulating environment for the cross-fertilisation of interdisciplinary ideas. The programmes are carefully developed and are intended to champion the novel techniques and innovations that will underpin important scientific advances.
Workshop proposals should take into account the following points:
Proposals should focus on cutting-edge scientific research in topic areas that are new, novel and not covered by traditional conferences.
Proposals that concentrate on emerging or cross-disciplinary themes are particularly encouraged.
Organisers should be experts in their field, with sufficient standing to attract world-class speakers and attendees.
Proposals should indicate how the Workshop will contribute to establishing new collaborations or research directions.
Each Workshop will consist of 30 delegates to include 10 early-career researcher places to be applied for and proposers should ensure diversity in the proposed attendee list. You will need to propose 20 speakers (but they don’t have to be confirmed).
As the scientific organiser your involvement will be focused on the science. We will undertake all of the logistical arrangements, liaise with the venue, organise speaker travel, help with arranging the programme and fund the meeting.
The first human genome sequence cost billions of dollars. Today, we’ve broken through the thousand dollar barrier and are heading towards $100. In this episode of Genetics Unzipped we talk to pioneering geneticist George Church about his plans for the ‘Zero Dollar Genome’ and his thoughts on the future of genome reading, writing and editing.
Plus, we find out what happened when one scientist’s interest in personal genomics got a little too close to home, when Manuel Corpas invited his whole family to share their personal genomes with the world.
If you enjoy the show, please do rate and review and spread the word. And you can always send feedback and suggestions for future episodes and guests to podcast@geneticsunzipped.com
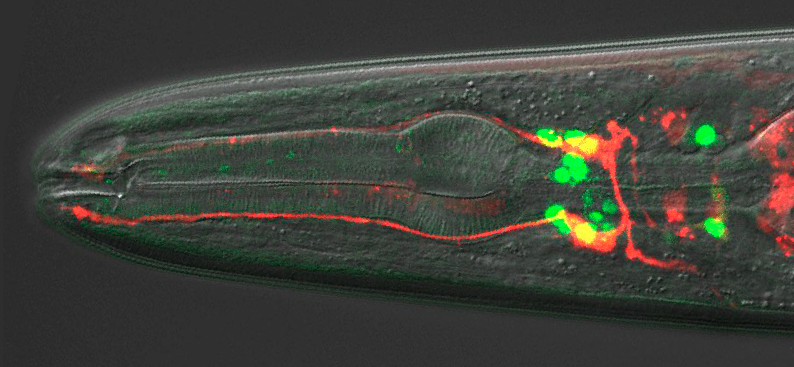
A fundamental aim in developmental biology is to understand how the various cell types of the body are specified by differential gene regulation. Caenorhabditiselegans nervous system development provides a powerful system for studying this, as exemplified by a new Development paper reporting on how the BAG neurons that help the worm sense oxygen and carbon dioxide are specified. We caught up with first authors Julia Brandt and Mary Rossillo and their supervisor Niels Ringstad (Associate Professor at the Skirball Institute of Biomolecular Medicine and Department of Cell Biology at New York University) to find out more about the story.
From left to right: Niels, Julia and Mary.
Niels, can you give us your scientific biography and the questions your lab is trying to answer?
NR I’ve had a longstanding interest in molecular neurobiology. This started during my graduate studies with Pietro De Camilli at Yale University, where I studied molecules required for the synaptic vesicle cycle. During my postdoctoral training with Bob Horvitz at the Massachusetts Institute of Technology I was introduced to behavioural genetics of C. elegans as a method to identify molecular mechanisms that are required for the function of specific neuron-types or for specific neurotransmitter signals. My own group continues to study the genetic basis of C. elegans behaviours to advance understanding of neuromodulation and sensation.
Julia and Mary: how did you each come to join the Ringstad lab, and what drives your research?
JB I’ve always been interested in how specific cells acquire the characteristics that distinguish them from other cells. The nervous system is where this question really stands out. There are hundreds of kinds of neuron, and each kind has a unique morphology and distinctive physiology.
MR I was drawn to the lab by the possibility of using C. elegans to understand how neurons sense respiratory gases. The worm gas-sensing neurons are specified by a transcription factor called ETS-5, which has a vertebrate homologue that is required to specify neurons in the mammalian brainstem that sense respiratory gases in arterial blood. I thought that this similarity offered the opportunity to leverage techniques in developmental genetics to interrogate mechanisms that might be involved in human pathology, specifically fatal respiratory disorders like Sudden Infant Death Syndrome. Because I am training to be a physician-scientist, I wanted my graduate research to impact clinical medicine.
Why are the BAG neurons an interesting developmental model, and what was known about their specification before your work?
NR We have colleagues studying chemosensation or neurodevelopment who previously identified and characterized mutants that are defective in BAG development. Elissa Hallem at the University of California, Los Angeles, had identified the ETS-5 transcription factor through an independent study. Roger Pocock at Monash University in Australia collaborated with us in our study of ETS-5, and his group independently identified two other transcription factors that are important for BAG development, EGL-13 and EGL-46. From this work, we had the sense that the genetics of BAG neuron development would be complicated and rich. Indeed, our own genetic screens yielded mutants that did not come up in Elissa’s or Roger’s studies. Our screens also yielded a mutation that causes ectopic expression of a BAG fate – a phenotype that is the mirror-image of the more common absence-of-BAG fate phenotype. This mutation was the starting point for the work that Julia and Mary did.
JB The BAG neurons exhibit some remarkable physiological characteristics, and they have an interesting and easy-to-measure function. BAGs are able to sense the respiratory gas carbon dioxide as if it were an odour. The carbon dioxide that they sense is coming from environmental microbes, which the worms eat. We think that worms prefer to eat dead microbes, or microbes that are growing slowly, and BAGs are used to sense whether microbes are metabolically active. I think that the development of these neurons is interesting because understanding how the BAGs are put together can help us understand how they do what they do.
MR I agree with Julia. The BAG neurons are an interesting developmental model because they have remarkable physiology that is imparted by a developmental programme. And that programme requires a transcription factor, which is conserved between worms and humans. I like the idea that, by studying the developmental mechanisms that make a BAG neuron a BAG neuron, we can identify molecules required for its chemosensory function.
A wild-type adult hermaphrodite expressing an integrated ETS-5::GFP translational reporter and a Pflp-17::dsRed transcriptional reporter.
NR I agree with Julia and Mary. Understanding the developmental programme that generates a BAG neuron is going to teach us how these neurons execute a pretty cool function. And we now have the opportunity to study how that developmental programme is initiated and organized, which is new territory for us.
Can you give us the key results of the paper in a paragraph?
JB, MR & NR Chemosensory BAG neurons of C. elegansrequire the transcription factor ETS-5 to express a host of genes that are required for their specialized function. How expression of ETS-5 is regulated during development was unknown. Julia and Mary have found that ETS-5 expression is regulated by another transcription factor – VAB-3 – which is the nematode homologue of Pax6/Eyeless. Remarkably, the vab-3 locus encodes VAB-3 isoforms that have opposing functions with respect to regulating expression of a BAG fate. One isoform represses expression of ETS-5 in cells that are not supposed to become BAGs. Another isoform, however, is expressed in BAGs and is required for them to fully differentiate. Julia and Mary concluded that the decision to express one VAB-3 isoform or another is crucial for determining which cells become BAG neurons.
Have you got any ideas what controls which vab-3 isoform is expressed in a given cell?
JB, MR & NR That’s the key question! We had a short-lived feeling that we had learned something when we realized that ETS-5 expression was regulated by VAB-3. But now we feel like we have simply re-framed the question as one that concerns promoter-selection in the vab-3 locus. But we’re not completely in the dark, and we now have some ideas about what is going on at the vab-3 locus. Mary has been pushing hard to establish methods for purifying and sequencing chromatin associated with sparsely expressed transcription factors, and some of her studies are identifying factors that bind to vab-3 promoters. Stay tuned.
Why might the system of broad repression/selected activation of cell fates have evolved in animal development?
JB, MR & NR Our first thought is that biological systems aren’t designed or engineered, so we shouldn’t be surprised when we discover that they are rife with needless complexity and inefficiency. During evolution, a set of transcriptional repressors and activators might have been randomly permuted until they generated a pattern of ETS-5 expression that worked. We might be looking at the mess that was left behind in that workshop. But it is also possible that we are looking at a legacy of an ancestral and now-obsolete developmental programme. Cells with the characteristics of BAG cells might have been more widespread in the ur-nematode, and over time more efficient body plans that used those cells for other purposes became the norm. A final thought we have entertained is that there is some drive to carefully regulate expression of a BAG fate. A good control system will use both positive and negative regulators of its output, which allows rapid but controlled responses to inputs.
When doing the research, did you have any particular result or eureka moment that has stuck with you?
JB: Finding a mutant is always a eureka moment. I had found a large number of mutants that lacked expression of a BAG marker. It was pretty surprising to find a mutant with extra cells expressing that marker. And it was exciting to discover that the phenotype was caused by ectopic expression of ETS-5.
MR Julia had found that vab-3 mutated to cause ectopic expression of ETS-5, and she had tested a number of vab-3 alleles. We had the idea that one isoform was repressive and the other not, but we were confused by reports that vab-3 is highly expressed in BAG neurons. It was satisfying to design isoform-specific reporters and see that the long repressor isoform was not expressed by BAGs. This really crystallized our model of a promoter-selection event that patterns expression of a BAG fate.
Finding a mutant is always a eureka moment
And what about the flipside: any moments of frustration or despair?
JB & MR: Some of the mutants we used are difficult to work with because they are barely viable or the mutations cause lethality. ceh-32 was awful. It is an essential gene, but we also found that ceh-32 reporter transgenes could cause lethality. We had to figure out how much DNA to use to make the ceh-32::GFP trangenics that we needed for the project.
So, what is next for you two after this paper?
JB I’m wrapping up a post-doc and deciding on the next steps for my career.
MR I am halfway through my dissertation research. Now I’m focusing on using mRNAseq and ChIPseq to generate a genome-wide map of ETS-5 targets. I’m hoping this will lead to novel genes that function in carbon dioxide sensing.
Where will this work take the Ringstad lab?
NR We’re still trying to link the developmental programmes that specify a BAG cell to genes required for gas-sensing. Julia and Mary have put another transcription factor on the board – the isoform of VAB-3 that promotes a BAG fate. I think we want to know how that factor functions together with ETS-5 and what genes it regulates.
Finally, let’s move outside the lab – what do you like to do in your spare time in New York?
JB I enjoy life in Brooklyn, spending time with my family and reading.
MR I enjoy distance running, yoga and spending time with my cat, Ernie.
NR Family life, home repairs, and I’m trying to catch up on all the maths and physics I didn’t do at university because I was too preoccupied with biology.
Normal Human Chromosomes. John Campbell. Public Domain
Everyone knows that humans have 23 pairs of chromosomes, right? But back in the 1930s, the correct answer would have been 24. So where did that extra pair go?
In this episode from our series exploring 100 ideas in genetics, we explore the discovery of chromosomes – the strands of genetic material within every living cell – take a look at Lyonisation, and solve the case of the missing chromosomes.
If you enjoy the show, please do rate and review and spread the word. And you can always send feedback and suggestions for future episodes and guests to podcast@geneticsunzipped.com
In this episode of Genetics Unzipped we ask, what would have happened if Darwin had read Mendel? And what if they’d been on Twitter? Professor Greg Radick, a historian of genetics at the University of Leeds, has a few ideas. Plus, something else that Darwin would have loved – an ambitious project to sequence the DNA of everything across the tree of life from the team at the Wellcome Sanger Institute.
If you enjoy the show, please do rate and review and spread the word. And you can always send feedback and suggestions for future episodes and guests to podcast@geneticsunzipped.com
We are inviting applications for a 3-year PhD position in the general area of Cell Biology and Cell Biophysics, to commence in October 2019. The student will be physically based within the Department of Physiology, Development and Neuroscience (PDN, https://www.pdn.cam.ac.uk) at the University of Cambridge, UK. The Department has significant cell biology, biophysics and imaging infrastructure.
The proposed research project aims to explore how cells control their mechanical properties during cell shape changes. Cellular shape changes are driven by mechanical forces generated primarily by intracellular cytoskeletal networks. The project will focus on a key cytoskeletal network, the actin cortex, and investigate how cortex organization and mechanics are controlled at the molecular level. The project is interdisciplinary and will involve the combined use of live cell imaging, super-resolution microscopy, advanced electron microscopy, as well as molecular cell biology and cell biophysics techniques.
The cell cortex is a thin network of actin filaments, myosin and associated proteins that underlies the plasma membrane. The physical properties of the cortex, and primarily its contractile tension, determine the shape of animal cells. Precisely controlled changes in cortical contractility are at the basis of most cell deformations, and deregulation of cortical tension is implicated in a number of cancers.
The overall aim of the project is to understand how contractile tension is generated from the microscopic organisation of the cortical network. We will develop an interdisciplinary research plan to study:
– the spatial organization of cortical actin filaments, using cryo-electron microscopy and super-resolution optical microscopy,
– the biophysical basis of cortical contractility.
We will investigate these questions in the context of cell division, where the cortex reorganizes to create a tension gradient that drives cell cleavage during cytokinesis.
Candidates should have, or expect to obtain, a first class or upper second class honours degree in subjects such as Molecular Cell Biology or Biophysics. Students with a Physics degree who wish to work at the interface of physics and biology are also strongly encouraged to apply.
Funding is available to UK and EEA candidates. This project is a part of a collaboration with a partner at the University of Zurich (Switzerland) and will necessitate some international travel.
Suitably qualified and enthusiastic candidates interested in performing research in a dynamic multidisciplinary scientific environment should apply online via the University’s Applicant Portal for a PhD in Physiology, Development and Neuroscience (BLPN22).
List the name of your prospective supervisor Professor Ewa Paluch and the project title in the Research section of the form. You will also be required to submit a research proposal (1-2 pages of A4). An application is only complete when all supporting documents, including the 2 academic references, are submitted. It is the applicants responsibility to ensure their referees submit their references before the closing date Monday 27 May 2019.
Informal enquiries can be addressed to Prof. Ewa Paluch (ekp25@cam.ac.uk).
Our laboratory of synthetic development at IMBA (Vienna) is looking for a postdoctoral colleague to decode the self-organising behaviours of early embryos. This position is part of a HFSP with Shantanu Singh (Broad Institute) and Kyogo Kawaguchi (Riken Institute), and requires regular and frequent exchanges with these labs.
Requirements. We are looking for a colleague with:
a strong interest and experience in quantitative high-content imaging including significant experience with microscopy, biological assay development, and some experience using image processing software on biological assays
an ability to clearly communicate with biology collaborators about project aims and results, and with software engineers about image processing and software needs
a personality that is helpful, professional, highly organized, and goal-oriented
useful but not required: stem cell culture experience, CellProfiler expertise, programming experience (e.g., Python), and experience with Linux environments, cloud computing clusters, and bioinformatics/statistics tools (e.g., R).
About us. Our laboratory gathers stem cell biologists, geneticists, and engineers who study organizing principles in the early embryo. We created the blastoid system, a currently unique in vitro model of the pre-implantation conceptus generated solely from stem cells [Nature 2018]. Upon transfer in utero, blastoids implant and initiate a pregnancy. We also develop microsystems, genetic and computational tools to modulate and analyse self-organisation. We are conscientious, enjoy a professional, no-fuss environment, and are passionate about what we do.
About IMBA. The IMBA is an institute of the Austrian Academy of Science and of the Vienna Biocenter. It hosts 15 research groups that foster strong creativity and world-class science in biology, including stem cells, organoids and development. IMBA and the Vienna Biocenter provide the utmost technical support to researchers, including in microscopy (e.g., lattice, mesoscale and macroscale LSFMs handled by technical experts), so that you can focus on the science. See https://www.imba.oeaw.ac.at/scientific-facilities/
We are seeking to recruit a new member to our team at the University of Cambridge that contributes to two Drosophila databases: Virtual Fly Brain (VFB; http://virtualflybrain.org) and FlyBase (http://flybase.org/). Virtual Fly Brain is a resource that helps Drosophila neuroscientists map and visualize neuronal circuits, and find the experimental reagents to manipulate the circuits to reveal their role in perception, learning, memory and complex behaviours. The successful applicant will aid this process in three ways: 1) by annotating the expression of Gal4 drivers within the adult brain, using data from large scale datasets and the primary research literature; 2) assisting in the integration of data from single-cell RNA sequencing datasets; and 3) leading the effort to assess the functionality of improvements to the website, by recruiting members of the local neuroscience community and working with them and other VFB members to perform usability testing of the website. Much of the curated data will also be integrated into FlyBase, the primary community resource for Drosophila genetic, genomic and functional data. You will interact day to day with the rest of the expert scientist-curators in the Virtual Fly Brain and FlyBase team, housed in the Department of Physiology, Development and Neuroscience, and VFB colleagues in the Cambridge area and Edinburgh. Members of VFB and FlyBase will provide appropriate training.
The Mokalled lab in the Department of Developmental Biology at Washington University School of Medicine is seeking highly motivated scientists to join our vibrant and collaborative research team (http://www.mokalledlab.com/). We use zebrafish as a discovery platform to study spinal cord regeneration across species. Candidates with expertise in zebrafish regeneration models or cell reprogramming models are encouraged to forward a cover letter, CV, and list of 2-3 references to mmokalled@wustl.edu.
The Young Tunisian Researchers in Biology Network:
Tunisia is suffering brain drain, especially post Tunisian-revolution (2011). Recent non-official reports say that over 90.000 senior quitted Tunisia since 2012, most of them are doctors, pharmacists and engineers. Aware of considerable role that may Tunisian scientist diaspora could play in the development of Tunisia in general and science and education in particular, the YTRB project was born (Young Tunisian Researchers in Biology). The project is all the more credible as all the founders were graduated from Tunisian universities and spent at least one post.doc outside Tunisia. The idea came out in early 2015 during a biology conference in Tunisia. Young Tunisians post.doc working in France, Tunisia, Sweden and Germany discussed the opportunity to organise a scientific event in Tunisia dedicated to young principal investigator (PI) as speakers essentially. The first YTRB committee was created with a fixed target: May 2016 will be the date of the 1st International Symposium of Young Researchers in Biology in Tunisia. We had an excellent speakers selection from Tunisia, Brazil, Europe and Asia. We had also a plenty of short oral talks and posters presented by local and international participants and definitely putting the event as the major scientific symposium in biology ever organised in Tunisia. Encouraged by this big success, we decided to continue building the Network. The major aim of YTRB is to create a new and young collaboration network and to help young Tunisian colleagues to connect to their peers all over the world, in order to provide knowledge, experience and technology exchange. The main activities of the YTRB Network are first conducting biannually an international biology symposium for young researchers. In collaboration with Tunisians universities and faculties, the organisation of several workshops in different thematic, Skype conferences and webinars, technology advances days wish are a mini symposium with focus on single thematic. Moreover, we created an annual science vulgarisation prize to help young Tunisian scientists to communicate their findings to a large public. In summary, we believe that YTRB is creating a new science climate in Tunisia marked by increasing the meetings, symposia, workshops and even chair in universities with the tag “young scientist” since the first symposium May 2016. This definitely encourages us to continue our work and investment to contribute modestly renewing the scientific research and build together the future science in Tunisia with the motto: Be Young, be Brilliant, Be YTRB!
Audience during the first international symposium of the YTRB Network, Monastir, Tunisia, May 2016
Organisers and volunteers during the second international symposium of the YTRB Network, Hammamet, Tunisia, May 2018
Our Thanks To
None of the work by the YTRB Network would be possible without the amazing devotion of Chamseddine Kifagi, Myriam Fezai, Fakher Frikha, Imen Chamkha, Walid Elfalleh, Radhouène Doggui, Ons Mamai, Chaker Slaymi, Radhouène Doggui, Nourhène Fendri, Abdelaziz Tlili, Alif Chebbi, Azza Sellami, Hana Trigui, Dhouha Daassi and many others. In addition, the YTRB activities could not be performed without the generous help from various organisations and scientists. I would like to thank the German, Italian, Spanish and French cultural institutes in Tunisia. Moreover, I’m grateful to the MNS” (Mediterranean Neuroscience Society, https://mnsociety.live/) and DrosAfrica (http://drosafrica.org/). We encourage every reader of this article to visit our website (https://ytrbiology.weebly.com/) and help the Network from their own work and time with any advertisement and financial support.

 (No Ratings Yet)
(No Ratings Yet) The first human genome sequence cost billions of dollars. Today, we’ve broken through the thousand dollar barrier and are heading towards $100.
The first human genome sequence cost billions of dollars. Today, we’ve broken through the thousand dollar barrier and are heading towards $100. 




 (7 votes)
(7 votes)