The Department of Genetics at Yale University School of Medicine invites applications for junior or senior tenure-track faculty positions. The search is open to investigators from all areas of biological and biomedical research. We are particularly interested in applicants working in one of the following areas Developmental Biology, Imaging, Quantitative Biology, Computational Biology, Genomics, Systems Biology, and Genetics. Applications from investigators working at the interface of these areas will be strongly considered. The rank of the appointment will be commensurate with experience and the positions come with a substantial start-up package.
The Department of Genetics comprises an exceptional group of 31 primary basic science faculty with research interests including fundamental aspects of Developmental Biology, Genetics, Genomics and Epigenetics, using different model systems including flies, worms, fish and mouse, and humans (https://medicine.yale.edu/genetics/). The Department is closely associated with science initiatives at Yale including The Cancer Center, The Center for Neuroscience, The Stem Cell Center and the Yale Center for Genome Analysis
Candidate must hold a Ph.D., M.D., or equivalent degree. Applicants should upload a cover letter, a curriculum vitae, a description of previous research (1 page), a concise statement of research plans (up to 2 pages), reprints of 2 publications, and the names of 3 references to the Interfolio website at: https://apply.interfolio.com/52614). Specific inquiries about the position may be sent to the attention of Dr. Antonio Giraldez, Chair of the Department of Genetics, at genetics.admin@yale.edu. Applications will begin to be evaluated on November 1, 2018.
Interviews will take place as part of a multidisciplinary symposium including candidates for different searches. Please reserve the dates of January 15, January 22 and February 11 (snow date) as potential dates for the symposium in case you are selected for an interview.
Yale University is an Affirmative Action/Equal Opportunity employer. Yale values diversity among its students, staff, and faculty and strongly welcomes applications from women, persons with disabilities, protected veterans, and underrepresented minorities.
We are delighted to invite you to attend the international conference: Assisted Reproduction Technologies – long term perspectives, that will take place in Auditorium Maximum of the Jagiellonian University in Cracow, September 7-8, 2018.
The program of the conference will include invited lectures, presentations of the latest research, many constructive discussions to share and explore new horizons in the field of Reproductive Medicine and Epigenetics between leading specialists/scientists from around the world.
The registration is now open. Participation in the conference is free of charge. We also encourage you to send abstracts and to active participation in the poster session.
About a decade ago I came to the National Institutes of Health (NIH) to work with my mentor, Jeff Baron, to study childhood growth and to tackle one of the unsolved mysteries in biology – mechanisms for body size determination (1). Years have passed, and we probably still don’t understand what makes an elephant an elephant. What we do know is that body growth happens for much longer period of time in an elephant than in a mouse, which contributes to it being bigger. Body growth slows and eventually stops in all mammals, and we think that what regulates the timing of “growth deceleration” holds the key to understanding the mechanism for body size control (2).
Growth does not only slow with age in soft tissues. The bones that constitute our skeletal system – more than 200 of them – also grows quickly in young children but gradually slow with age. It has been well-established that longitudinal bone growth is driven primarily by the growth plate – a small piece of cartilage tissue found near the ends of our long bone. As you grow taller, your growth plate also gradually becomes thinner and thinner. Eventually, all the growth potential in your growth plate became exhausted, and that is when growth plate fuses at around 20 years of age for most people and when linear growth stops. We call this age-related deterioration of growth plate function that limits bone growth “growth plate senescence” (senescence is another word for aging).
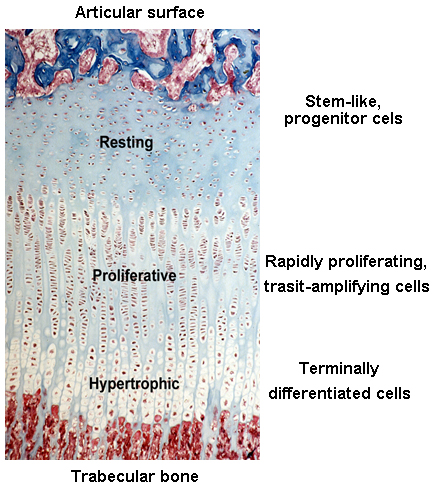
Before and right around the time I joined the group, Jeff and other colleagues were studying the expression of different paracrine signaling pathways in the growth plate (3) using a new (at the time) technique called growth plate microdissection. The growth plate has an interesting and elegant spatial arrangement. From one side of the growth plate (closer to the joint surface) to the other (closer to bone marrow), cells in the growth plate called chondrocytes gradually transition from progenitor cells (in the resting zone) to transit-amplifying cells (in the proliferative zone) and finally becomes terminally differentiated cells (in the hypertrophic zone). This cellular transition contributes to the expansion of cartilage tissue and subsequent bone growth. Chondrocytes at each of these zones have dramatically different gene expression profiles, which is fundamental to their corresponding cellular functions.
Before then, most of the studies elucidating the changes of gene expression across the growth plate were done by performing in situ hybridization or immunohistochemistry, interrogating expression of one gene at a time. These techniques are however semi-quantitative at best and could be prone to false positive results. Technically, the only way you can be sure the immunohistochemistry signals are real is when you do it in a knockout mouse and the signal disappears.
Ola Nilsson, a fellow in the lab at the time and a good friend of mine and now professor at Karolinska Institute, was developing a technique called growth plate microdissection. He would make frozen sections of the rat growth plate and cut it with a razor blade under an inverted microscope into different zones, one slide at a time. It was a game-changer because that allowed us to extract RNA from different growth plate zones to study gene expression using qPCR or microarray. This technique has its downside of course, as it requires steady hand to make precise cut on tiny tissues, extremely long day (you need to combine 40 dissections for each sample), and occasional inhalation of xylene vapors (because you need to keep the tissue on the slide in xylene when you cut). Nevertheless, over the years, this technique led us to many interesting discoveries, like the regulation of bone growth by growth plate senescence (4, 5).
In addition to body size control mechanisms, Jeff and I were also wondering how different bones in our body grew to different lengths to achieve skeletal proportions. How does the left and right leg always grow to (almost) the same length (6), and what makes the finger bones so much shorter than the leg bones? We have a hypothesis that maybe senescence in the growth plate are happening at different rates in the longer and the shorter bones. However, to test this hypothesis, we want to compare the gene expression between growth plates from the leg bones and the finger bones, which are way too small to dissect using our method. And even if we were able to dissect the finger growth plates, it would take many animals to get enough RNA to study at the time.
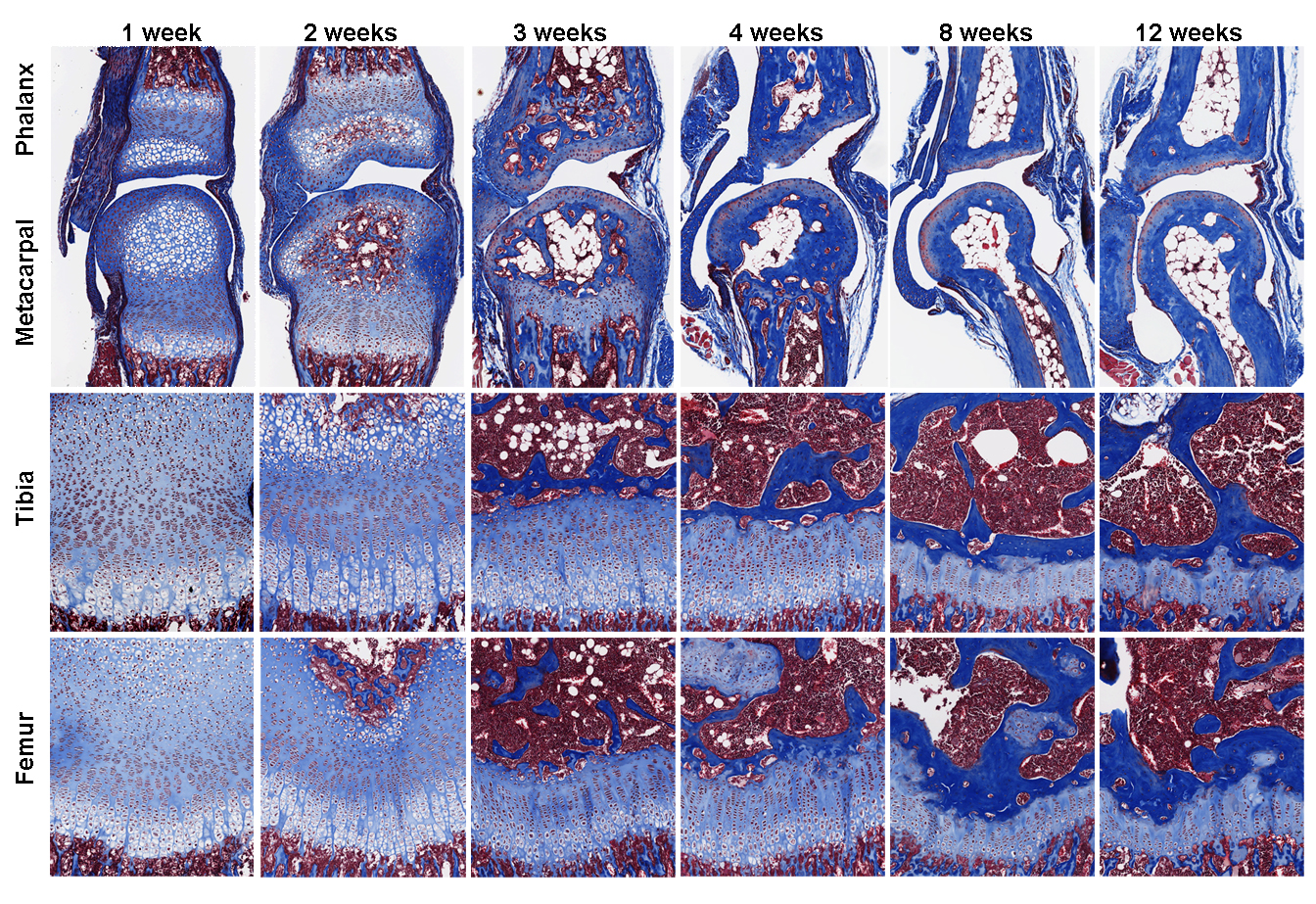
Luckily, remarkable advancement in sequencing technology in recent years significantly reduced the amount of starting material needed for whole transcriptome interrogation. More importantly, a few years ago I met two gentlemen from National Cancer Institute, Jaime Rodriguez (now at MD Anderson) and Jeffrey Hanson, who taught me all about laser capture microdissection (LCM). LCM is a much-improved version of our original microdissection method, because now it is relying on the machine-controlled laser, rather than my shaky hands, to make the cut. After years of optimization, we can now isolate cells from much smaller tissues, such as the growth plate zones from mouse proximal phalanx, for RNA extraction. We have achieved what was (to us) seemingly impossible few years ago, using RNA-Seq to compare zonal expression profile between the leg bones and the finger bones.
Our findings, which was published recently in PLOS Biology (7), showed that growth plate senescence is more advanced in shorter bones than in longer bones, and this differential aging contributes to the disparities in bone growth. Our RNA-Seq data showed that the molecular mechanisms that underlie this differential aging between bones involve differential expression of signaling pathways between longer and shorter bones, such as IGF1, BMP, and WNT signaling, which regulate cell proliferation/differentiation in the growth plate. We are currently trying to understand whether differences in bone lengths between different species are also attributable to similar difference in growth plate senescence.
The neural crest is a progenitor population with the capacity to contribute to all vertebrate germ layers. The transcription factor and signalling pathway activity underlying this remarkable pluripotency have been well studied, but the role of the epigenetic state is less well understood. A new paper in Development examines the role of histone acetylation in regulating the neural crest’s remarkable pluripotency. We caught up with authors Anjali Rao and Carole LaBonne, Erastus O Haven Professor of Life Sciences at Northwestern University, to find out more about the story.
Anjali and Carole
Carole, can you give us your scientific biography and the questions your lab is trying to answer?
CLB I have always been in love with the beauty of embryonic development, and the elegance of developmental decision-making. I did my PhD work at Harvard studying germ layer formation; specifically the role of FGF signalling in mesendoderm formation. During my graduate work I learned about the neural crest and I knew that was what I wanted to work on as a post-doc. It is an absolutely fascinating cell type, and this year is the 150th year of its discovery by Wilhelm His. Acquisition of the neural crest drove the evolution of vertebrates because these stem cells contribute a myriad of novel structures to the basic chordate body plan. Another interesting feature of these cells is that they are migratory and invasive; they need to actively disperse throughout the embryo to the places where they will form those structures. The mechanisms that they use to do this have been co-opted by cancer cells to mediate metastasis, so we can actually learn important things about tumor progression by studying the neural crest.
Two of Carole’s early papers (in Development!), from her PhDand postdoc
After my post-doc at Caltech I moved to Northwestern, where my lab has continued to study various aspects of neural crest development, including the signalling pathways and GRN components that control the stem cell attributes, behaviour and lineage decisions of these cells. More recently we have also been studying the pluripotent cells of the early blastula embryo. We do most of this work in Xenopus, which is a fantastic system for asking questions about early vertebrate development. Xenopus is closely related to mammals but the embryos develop rapidly and externally in simple saline solution, and the large (~1.2mm) size of the embryos facilitates isolation of explants or single cells. These embryos also provide copious materials for genomic and proteomic studies, which we are doing a lot of these days. Xenopus embryos are ideal for rapid gain/loss of function studies, including CRISPR-mediated genome editing, and the fate of every early embryonic cell has been mapped. We are very fortunate to have a fantastic resource center for this model, the National Xenopus Resource (NXR), that is located at the MBL in Woods Hole MA. The NXR generates and distributes transgenic lines and CRISPR mutants to the community, and also holds training workshops.
Anjali, how did you come to join the LaBonne lab, and what drives your research?
AR I have always been interested in understanding the regulation of stem cell maintenance. Before starting my PhD, I was working in a lab that studied the regulation of kidney stem cells during kidney fibrosis. Often diseases are caused by the misregulation of developmental pathways, and I hence got interested in studying the regulation of stem cells during embryonic development. The LaBonne lab was the perfect fit. Early during my PhD, our lab proposed a new model for the genesis of the neural crest that suggested that these cells arise due to a retention of stem cell potential, and we became really interested in identifying the mechanism for this. In particular, I was tasked with identifying chromatin remodelers required for neural crest stem cell maintenance, and characterizing the epigenetic changes that take place during the process of neural crest formation.
What led you to become interested in the epigenetic regulation of neural crest and blastocyst pluripotency?
CLB A few years ago we realized that neural crest cells and pluripotent blastula cells share a remarkable number of gene regulatory components. In particular, many of the transcription factors that we had long studied in the neural crest are first expressed in, and play essential roles in, blastula stem cells. That led us to hypothesize that neural crest may have arisen (and thus vertebrates evolved) via retention of features of these early pluripotent cells, even as neighbouring cells became lineage restricted. We began actively studying the shared features of these two cell types at the level of signalling pathways and transcription factors. For example some of our most recent work has focused on FGF-mediated Map Kinase signalling, which is essential for maintaining the developmental potential of both cells types. This was fun for me because it brought me full circle back to my graduate studies.
There is growing literature on the roles of epigenetic readers, writers, and erasers in controlling stem cell attributes and lineage decisions in both cultured ES cells and embryos. Thus, it was clear that we would need to examine conserved and divergent roles for these factors, and epigenetic mechanisms more generally, in neural crest and pluripotent blastula cells. We are finding interesting roles for a number of epigenetic regulatory factors, and so Anjali’s HDAC paper is just the tip of that iceberg.
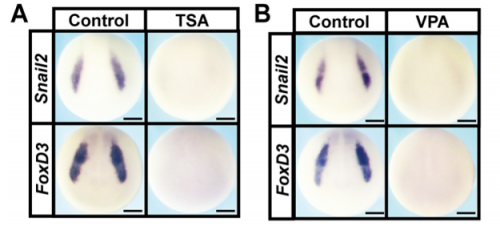
HDAC inhibition leads to loss of expression of neural crest factors, from Fig. 1 in the paper
Can you give us the key results of the paper in a paragraph?
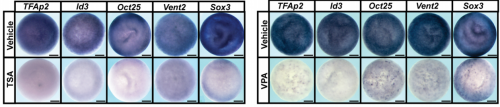
AR & CLB We found that Histone Deacetylase (HDAC) activity is essential for neural crest formation, both in whole embryos and explants of pluripotent cells. HDAC activity is also required for the pluripotency of blastula stem cells. Importantly, both cell types are characterized by low levels of histone acetylation, highlighting another shared feature of these cell populations, and HDACs function to maintain this state. Interestingly, we found that blocking HDAC activity in blastula-derived cells results in aberrant expression of markers of multiple lineages, and prevents the cells from committing to any lineage. Finally, we found that increasing HDAC activity can enhance the reprogramming of cells to a neural crest state. Our data suggests that HDACs play an important role in retaining the cells that will become neural crest cells in a stem cell state through gastrulation and neurulation, even as other cells in the early embryo become lineage restricted.
I was intrigued by your finding that HDAC inhibited cells co-expressed several lineage markers at once. What do you think explains these cells’ confused identities?
AR That is a great question. It seems that HDAC activity must be critical for keeping the acetylation state of genes controlling a number of different lineage states low in pluripotent stem cells, so that these genes are not expressed before the cells have made a specific lineage decision. Inhibition of HDAC activity releases this control, and therefore causes low-level expression of markers of multiple lineages simultaneously. An apparent consequence of co-expressing genes that promote multiple different lineage states is an inability to commit to any one lineage. During normal development, signals instructive of a specific lineage state likely release HDAC control of gene expression essential to that lineage, while ensuring that other lineage control factors remain unexpressed.
Late blastula embryos showing reduction in pluripotency factors following HDAC inhibition, from Fig. 2 in the paper
What do you think might be downstream of HDAC in the neural crest, and do you expect these targets to be the same as in the naïve blastula?
AR & CLB We know that global levels of H3K9 and H3K27 acetylation are similar in blastula and neural crest stem cells, and we think that the key targets of HDAC activity are likely to be similar in both of these cell types. We have used RNA-Seq to characterize the changes to the transcriptome that occur in response to HDAC inhibition, and that gives us important clues to what the key targets might be. We are currently examining the deposition of specific acetylation marks genome wide using ChIP-Seq to investigate this further.
When doing the research, did you have any particular result or eureka moment that has stuck with you?
AR I was really excited when I found that increasing HDAC activity enhanced the ability of blastula cells to be reprogrammed to a neural crest state. I think it definitely was my eureka moment, because it gave us the fundamental insight that HDACs actively promote the retention of pluripotency that leads to formation of the neural crest.
Animal pole explants in various experimental conditions, from Fig. 3 in the paper.
And what about the flipside: any moments of frustration or despair?
AR My most frustrating moments during this project were actually during the revisions for the paper. The reviewers asked us to confirm the phenotypes we observed following chemical inhibition of HDAC activity by using an independent means of down-regulating this activity. This was very challenging because HDAC1 is very highly expressed in the early embryo. However, with Carole’s guidance, I was able to target a morpholino that blocks HDAC1 translation to the neural crest, and show that this phenocopied the effects of the chemical inhibitors.
What next for you after this paper?
AR I am currently diving deeper into understanding the epigenetic regulation of the neural crest using ChIP-Seq and proteomics, and I hope to gain further insights into the role of specific epigenetic marks in stem cell maintenance.
And where will this work take the LaBonne lab?
CLB We are continuing to drill down on the epigenetic control of pluripotency in both naïve blastula cells and neural crest cells, and in developmental decision making more generally. Ultimately we want to understand these processes at single cell resolution using both quantitative imaging and single cell genomics. We are fortunate to be a part of a new NSF-Simons Center for Quantitative Biology focused on understanding emergent properties in developmental biology. Our project is focused on understanding dynamical transitions in cell states, and we have amazing math and modelling collaborators in the Center who are going to help us to take these studies to whole new levels of analysis.
Finally, let’s move outside the lab – what do you like to do in your spare time in Illinois?
AR In my spare time, I enjoy running on the beautiful trail along Lake Michigan and I am currently training to run the New York City marathon in November. I also love reading, particularly historical fiction, and trying out new restaurants around Chicago.
CL I am the mother of two and the Chair of my department, so spare time is a bit of a foreign concept for me! In the time I do have I love to cook, travel and take long walks along the lake with my two standard poodles.
Studying mechanisms of genetic renal disease in Drosophila and mouse
The Renal Division of the University Medical Center Freiburg is offering a PhD position (DFG-funded). The position is available for three years and should be filled as soon as possible (starting in fall 2018).
Project:
The lab is interested in studying mechanisms of genetic renal disease with a focus on monogenic causes of nephrotic syndrome.
To this end we are utilizing the Drosophila and mouse animal models in conjunction with approaches in vitro.
The podocyte-like Drosophila nephrocytes will be employed to characterize disease genes functionally and for whole-animal drug screening to develop novel therapeutic strategies.
The candidate will work in diverse scientific environment with intensive supervision and support.
Requirements:
We are looking for a highly motivated and ambitious candidate with a strong interest in basic research with clinical relevance.
The applicant should hold a master’s degree in Biology, Molecular Medicine or an equivalent discipline.
Experience in cell biology, mice and/or Drosophila is desirable but not mandatory.
Application: Interested candidates should send an application including a CV, a brief motivation letter and two references to
Dr. Tobias Hermle (tobias.hermle[at]uniklinik-freiburg[dot]de).
Selected References:
1. Hermle, T, Schneider, R, Schapiro, D, Braun, DA, van der Ven, AT, Warejko, JK, et al.: GAPVD1 and ANKFY1 Mutations Implicate RAB5 Regulation in Nephrotic Syndrome. J Am Soc Nephrol, 2018.
2. Helmstadter, M, Huber, TB, Hermle, T: Using the Drosophila Nephrocyte to Model Podocyte Function and Disease. Front Pediatr, 5: 262, 2017.
3. Hermle, T, Braun, DA, Helmstadter, M, Huber, TB, Hildebrandt, F: Modeling Monogenic Human Nephrotic Syndrome in the Drosophila Garland Cell Nephrocyte. J Am Soc Nephrol,28: 1521-1533, 2017.
A postdoc position is available in the Heemskerk lab, Department of Cell and Developmental Biology at the University of Michigan Medical School.
We are looking for a postdoc that shares our excitement about using stem cells to discover the developmental mechanisms underlying embryogenesis and organogenesis. The lab is highly interdisciplinary and combines experimental and theoretical methods from biology, physics, and engineering, focusing on quantitative live-cell measurements to study spatial organization and cell fate determination in human pluripotent stem cells.
The broader institutional environment of the University of Michigan – the public university with the highest research spending in the United States – provides exceptional resources for professional development and trailblazing scientific exploration. The university is located in Ann Arbor, which is consistently named among cities with the highest quality of life in the US.
The successful candidate is a rigorous thinker with either a relevant biology background and strong interest in quantitative methods, or a quantitative background (e.g. physics, engineering, math) and strong interest in wet lab biology.
For more information see http://idseheemskerk.com.
To apply, please send a CV with publication list and three references, as well as a cover letter stating your motivation to iheemske@umich.edu.
Welcome to our monthly selection of developmental biology (and related) preLights
preLights, the preprint highlighting service supported by The Company of Biologists, has now reached over 150 posts since launching in late February. To further promote this platform, we are now featuring preLights content in three of The Company of Biologist’s journals – including Development. In addition, complementing the Node’s monthly trawl for preprints, we are starting a monthly series where we highlight some of the exciting developmental biology (and related) preLights articles. So, on to our first post!
Flies, fish and chicks
preLighters featured several exciting preprints using Drosophila. A modern imaging approach to study transcriptional dynamics, combined with mathematical modelling shed new light on the generation of the famous even-skipped stripes, and was covered by Erik Clark. Mathematical modelling was also key in getting at the mechanisms of dorsal closure in a preprint that Yara Sanchez and Arnaud Monnard reviewed together. Ivana Viktorinova’s preLight discussed how the wing epithelium responds to mechanical stress during morphogenesis, in which the mechanosensitive binding of the endocytic regulator p120 plays a major role. Drosophila embryos also have to tolerate other kind of stresses during their development, such as hypoxia; Sarah Bowling’s preLight featured a study that showed how the fat body of larvae senses low oxygen and modulates TORC1 in response.
Mechanical stress regulates E-cad turnover; reproduced from Figure 3. of the preprint by Iyer et al. and discussed by Ivana Viktorinova
There were plenty of exciting preprints coming from the zebrafish community, and the preLighters did their best to cover them. James Gagnon highlighted a fascinating study showing the essential role of a novel small protein for species-specific fertilization. Genes and phenotypes were studied at a much larger scale in a story – covered by Daniel Grimes – about an impressive effort to link functions to human schizophrenia-associated loci. The strength of zebrafish in modelling complex human diseases was also nicely featured in preLights posts by Hannah Brunsdon and Giuliana Clemente. Hannah highlighted how a putative human CHD gene variant enhances the phenotype of a known CHD-associated gene, and called attention to the importance of investigating the contribution of risk alleles to existing disease-associated phenotypes. Giuliana wrote about the introduction of elements of the human immune system into zebrafish, which could allow future research to investigate how immune cells contribute to cancer progression and metastasis. It turns out that immune cells, and specifically the spatial-temporal dynamics of cytokine induction, are crucial for spinal cord regeneration, as explored in Shikha Nayar’s preLight.
Wouter Masselink and Ashrifia Adomako-Ankomah both highlighted preprints that combined chick embryology methods with modern molecular approaches to gain new insights into one of developmental biology’s ‘classical’ problems: limb development. Wouter covered a study showing that the collinear activation of Hox genes is important for controlling the position of the forelimb. Ashrifia’s post dealt with the question of how the final size of the limb is determined and a preprint showing BMP signalling-dependent regulation of proliferation rates in the limb bud.
Chromatin and genomics
The chromatin biology and genomics fields were well represented on preLights this month. Claire Simon and Sophie Morgani discussed how Sox3 and Pou5f3 act as pioneer transcription factors to open chromatin for genome activation in Xenopus, while the organization of inactive chromatin in senescent cells was the focus of Carmen Adriaens’ post. Lauren Neves’ preLight reported on a remarkable study, which showed that the histone H3-H4 tetramer – a main component of the building blocks of chromatin – also has copper reductase activity. This enzymatic function of histones may have been important for the emergence of eukaryotes. Several preLights featured the ever-increasing power of RNA-seq to answer interesting biological questions. A transcriptomic study in plants, highlighted by Martin Balczerowicz, investigated the level of transcriptional noise among genetically identical species. Rob Hynds’ preLight discussed the use of single-cell transcriptomics combined with bulk proteomics to study healthy ageing in mouse lungs. Finally, an RNA-seq study also gave insight into how a remodelled chloroplast is maintained in a non-photosynthetic alga, preLighted by Ellis O’Neill.
Tools & Technologies
Novel technologies featured heavily this month, for instance Rebekah Tillotson’s preLight on CRISPR gene drives in mouse, which could help scientists create disease models with multiple mutations. Satish Bodakuntla covered a novel expansion microscopy technique that goes beyond the limits of super-resolution. The most popular type of method to highlight turned out to be optogenetics. Mahesh Karnani reflected on two preprints that made major advances in achieving optogenetic inhibition of neural activity by soma targeting, and the preLight includes insightful comments from both research teams. Patricia’s preLight discussed optogenetic manipulation in locust brains, while moving a bit away from neuronal function, Srivats Venkataramanan preLighted a study that used an optogenetic tool to control the formation of stress granules.
Activation of a light-gated chloride channel causes excitatory and inhibitory neurons to release neurotransmitter. Reproduced from Figure 1. of the preprint by Messier et al. and discussed by Mahesh Karnani
Finally, preLighters’ coverage of reproducibility (see Carmen’s earlier post and Reid Alderson’s preLight from this month) and flaws in experiments (see Fabio Liberante’s post on false detection of circular RNAs from RNA-seq) signalled the importance of preprints in rapid dissemination of studies which are extremely valuable for the community, but often more difficult to get published in journals.
Considering reading something more out of your scope? Then explore the preLights website, where you can also find out about microbes in a Mars-analogue environment, Mycobacteria on your showerheads, or drug repurposing to combat brain-eating amoebae.
Postdoctoral positions are available in the laboratory of Daniel T. Grimes, Institute of Molecular Biology and Department of Biology at the University of Oregon. The laboratory focuses on symmetry in the biological world.
We are a new lab opening in January 2019. We want to understand how some features of vertebrate bodies (like limbs and the skeleton) develop with left-right symmetry, while others (like the heart, liver, and gut) develop with striking left-right asymmetries.
We use zebrafish to address these fundamental questions of developmental biology. Our main techniques include genome editing, confocal and micro-CT imaging, genetics and genomics, single-cell sequencing, and more. Current projects include: understanding how mechanical flow signals are generated and sensed by cilia in left-right patterning, investigating Polycystin signal transduction, elucidating mechanisms by which the spine remains straight during growth. We also model human diseases of aberrant (a)symmetry including heterotaxia, scoliosis, and primary ciliary dyskinesia.
We offer the opportunity to work closely with the PI, and to help build and shape lab culture. Your career and manuscripts are important; we are committed to the mentorship and long-term success of our lab members. We also offer a high quality training in zebrafish development and genetics. Zebrafish research was founded at the University of Oregon. As such, we share a large state-of-the-art aquarium with labs that use the zebrafish for diverse science. There is no better place to work with zebrafish!
The Biology Department at the University of Oregon offers an exceptional environment and a broad range of research, great core facilities and close ties to the Knight Campus, a new $1-billion initiative that brings together engineers, biologists and computational scientists to address pressing biological questions.
We are looking for ambitious candidates who hold (or be due to complete) a relevant PhD and have evidence of excellent scholarship. Experience with confocal imaging is a bonus but previous experience with zebrafish is not essential. Candidates should be able to work independently, to implement new technologies, and be productive and collaborative.
The position is fully funded but the candidate will be expected to seek external funding opportunities during their tenure.
The candidate should email (dtgrimes@princeton.edu) with a cover letter describing their career goals, previous experience, scientific interests and reasons for applying, along with a CV. Informal inquiries are welcome (see grimes-lab.com/join-us-1).
The Novo Nordisk Foundation Centers for Stem Cell Biology (DanStem) and Protein Research (CPR) at the University of Copenhagen are jointly seeking applications for a 2-year position of Bioinformatics Specialist.
The Novo Nordisk Foundation Center for Stem Cell Biology (DanStem) addresses basic research questions in stem cell and developmental biology and has activities focused on the translation of promising basic research results into new therapeutic strategies for cancer and chronic diseases. Learn more about DanStem at https://danstem.ku.dk/.
The Novo Nordisk Foundation Center for Protein Research (CPR) promotes basic and applied research on proteins of medical relevance. Spanning in silico biology, proteomics, biochemistry and cell biology, and the unifying research theme is the identification and functional exploration of proteins and protein pathways involved in disease. Learn more about CPR at www.cpr.ku.dk.
Background
Both DanStem and CPR are vibrant, internationally diverse and ambitious research centers housing modern laboratories with state-of- the-art facilities located in close proximity to one another at the Faculty of Health and Medical Sciences, University of Copenhagen. The setting is ideally suited for seamless collaboration and exchange of expertise, material and lively scientific communication. DanStem and CPR collaboratively operate three research platforms in imaging, flow cytometry and genomics. These platforms are staffed with experts who provide advanced training and tailored project planning and assistance. The Genomics Platform provides high-throughput sequencing services with personalized support, covering applications from classical genomics to single-cell transcriptomics and has generated a demand for bioinformatics support. Therefore, the Centers intend to develop a new informatics platform to full fill the need of bioinformatics and computational approaches to data analysis enhancing the scientific synergies between the two centers.
We are now seeking a highly motivated and qualified bioinformatician with specialty in genomics to work within the Informatics Platform at DanStem and CPR.
Responsibilities
The Bioinformatics Specialist will provide bioinformatics support, develop new pipelines, and organize practical training to assist DanStem and CPR researchers with analysis and interpretation of primarily genomics data. The bioinformatician will work within the newly developing Informatics Platform, as well as with the dedicated sequencing specialist to provide tailored service and support for multiple research projects of broad biomedical scope throughout the two research centers.
This Bioinformatics Specialist is expected to:
Develop tailored bioinformatics approaches and provide advice and assistance to DanStem and CPR researchers in their analyses, focusing in particular on the planning, processing, analysis and publication of transcriptomic and epigenomic next-generation sequencing data.
Prepare pipelines and scripts for use by DanStem and CPR researchers.
Provide training and design new approaches to analysis.
Cooperate and collaborate with DanStem and CPR researchers.
Cooperate with platform personnel and contribute to platform management.
Network actively with the global bioinformatics community and stay up-to-date with current trends in the field.
Similar to our other platform specialists, the Bioinformatics Specialist will sit at the nexus of the two centers. To ensure good support and guidance, the NGS Bioinformatics Specialist will work closely with the Big Data Management Platform at CPR, which handles data from other technology platforms, as well as among research groups that cover many systems-level aspects of biology and medicine, including the integration of molecular level and healthcare data.
Qualifications, competences and experience
Candidates are expected to have a Master’s or PhD degree in bioinformatics, computational genomics or biology, computer science and data analysis, or a biological or natural sciences discipline, with at least four years of experience in bioinformatics for genomics. In addition, we are seeking a candidate with the following competences and experiences:
Fluent programming skills in R/Bioconductor.
Experience with next generation sequencing data.
Strong statistics skills.
Knowledge of UNIX-like operating system.
Experience in working with biologists and an understanding of molecular biology and genomics.
Excellent oral and written communication skills and fluency in English.
Successful experiences in project management, including multi-tasking.
A service-minded and team player attitude.
Employment conditions
We offer
State-of-the-art computational infrastructure for data analysis, including access to cluster computing at the Danish National Supercomputer (Computerome) and excellent IT and systems administration support.
Stimulating and multifaceted research environment of high scientific and societal impact.
Possibility for continued education and training, especially to develop further as an expert in single cell bioinformatics.
With an initial duration until the end of 2020 with possibility of extension, employment can begin in November 2018 or upon agreement with the chosen candidate. The place of work is at DanStem, University of Copenhagen, Blegdamsvej 3B, Copenhagen. The position, with formal employment title of Senior consultant at the University of Copenhagen, will be in accordance with provisions of the collective agreement between the Danish Confederation of Professional Associations (AC/PROSA) and the Danish Ministry of Finance. To the salary is added a monthly contribution to a pension fund according to the collective agreement. A supplement could be negotiated, depending on the candidate’s experiences and qualification.
Questions
Informal inquiries can be directed to Chief Operating Officer Gretchen Repasky.
Your online application must be submitted in English by clicking ‘Apply now’ below.
Please note that only online applications will be accepted.
The application must include a statement of motivation, curriculum vitae, and copies of relevant diplomas.
Application deadline: September 2, 2018.
Only applications received in time and consisting of the documents stated above will be considered.
University of Copenhagen and The Centers highly value diversity and welcome applications from all qualified candidates, regardless of personal background and status.
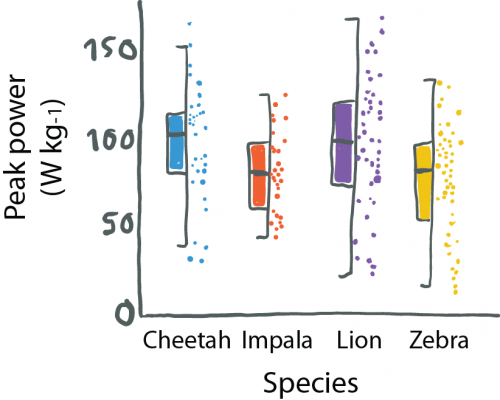
When reading about co-evolution of prey and predators, I stumbled across a cute new plot type: a half boxplot, half dot plot to show data distributions.
Half boxplot, half data plot. Figure re-drawn from Wilson et al. 2018 (doi: 10.1038/nature25479).
Wilson used this plot to simultaneously visualize summaries about their data (center, spread) and the actual data points. This allows us, the audience, to learn a lot about their results. That cheetahs are maybe binomially distributed and have outliers, or that zebras show a curious clustering.
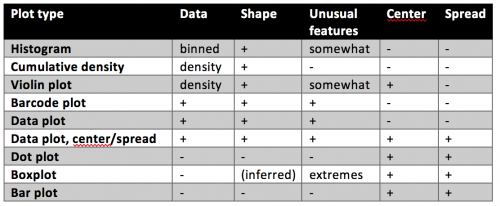
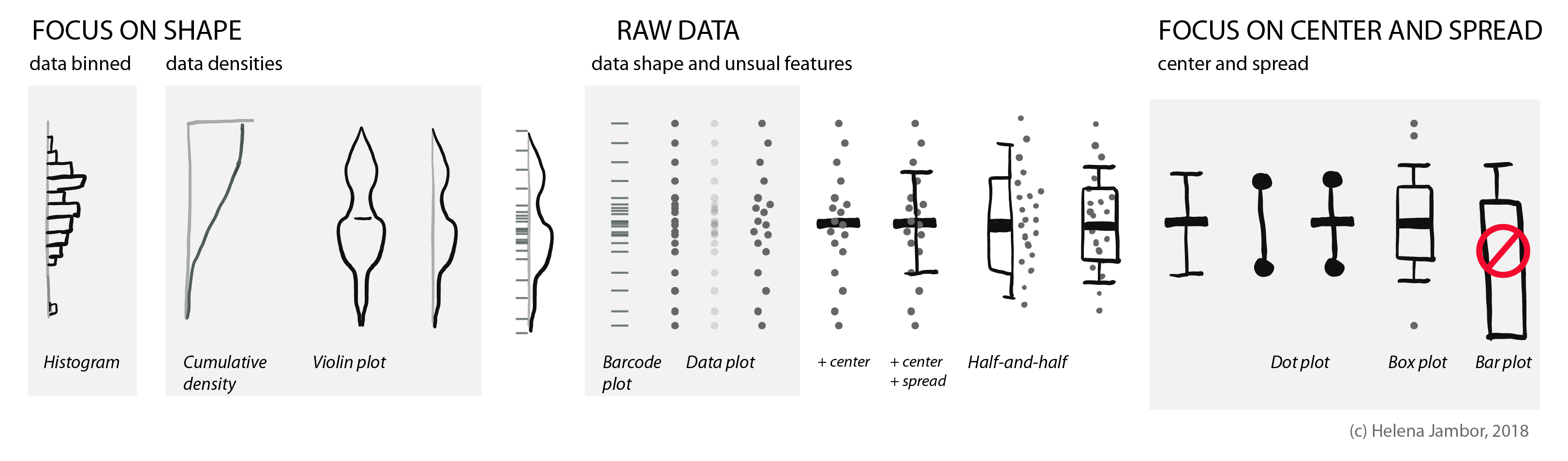
Your quick guide to distribution plots:
The half-and-half, aka dox-plot (a friend), led me to explore which visuals are commonly used for showing distributions.
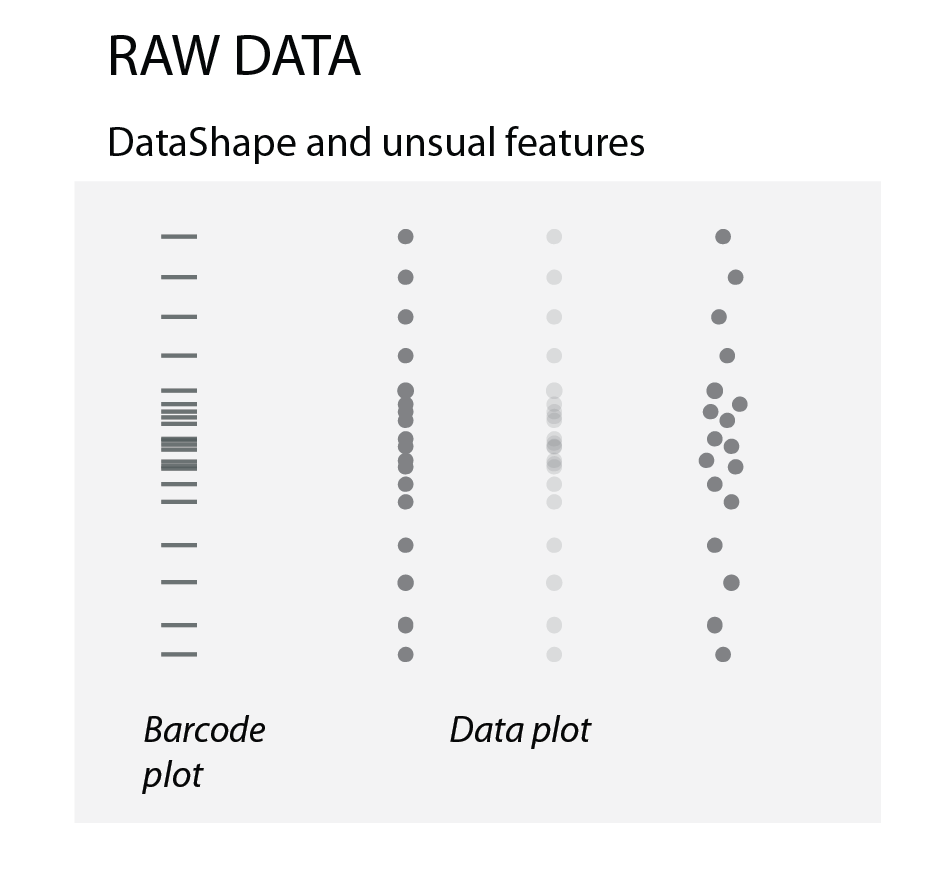
Raw data
To show how the raw data is distributed, we simply use dots or bars (as in barcode plots). When there are overlapping data points, we can use transparency or “jittering”. Jittering is distributing the data points in a given area, for increased clarity: the y-position remains the same, the x- position becomes random.
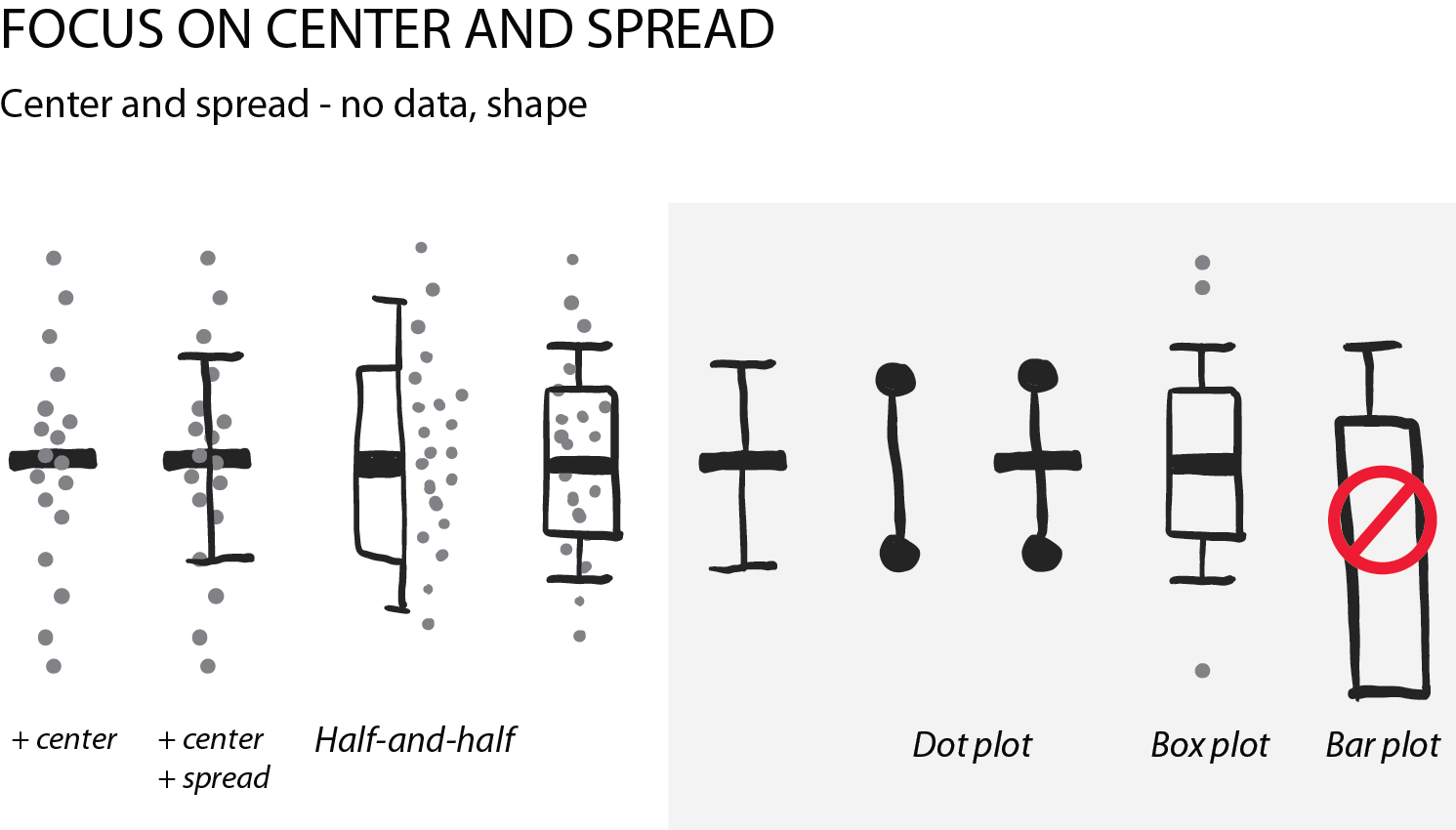
Summarizing the data: center and spread
Often, we are interested in summarizing statistics to judge and compare data. By convention the center (median) is indicated with a horizontal bar and the spread (variance, standard deviation) with vertical whiskers. Common plot types for this are the “star-wars rebel fighter”, the dot plot and boxplot. Bar plot used to be widely used, but are now banned by most journals for concealing most relevant information, so they are here only for completeness (see previous post). Very often these days I see boxplots that are overlaid with the data points – this works really well for up to 100 data points and is easy to implement with most software.
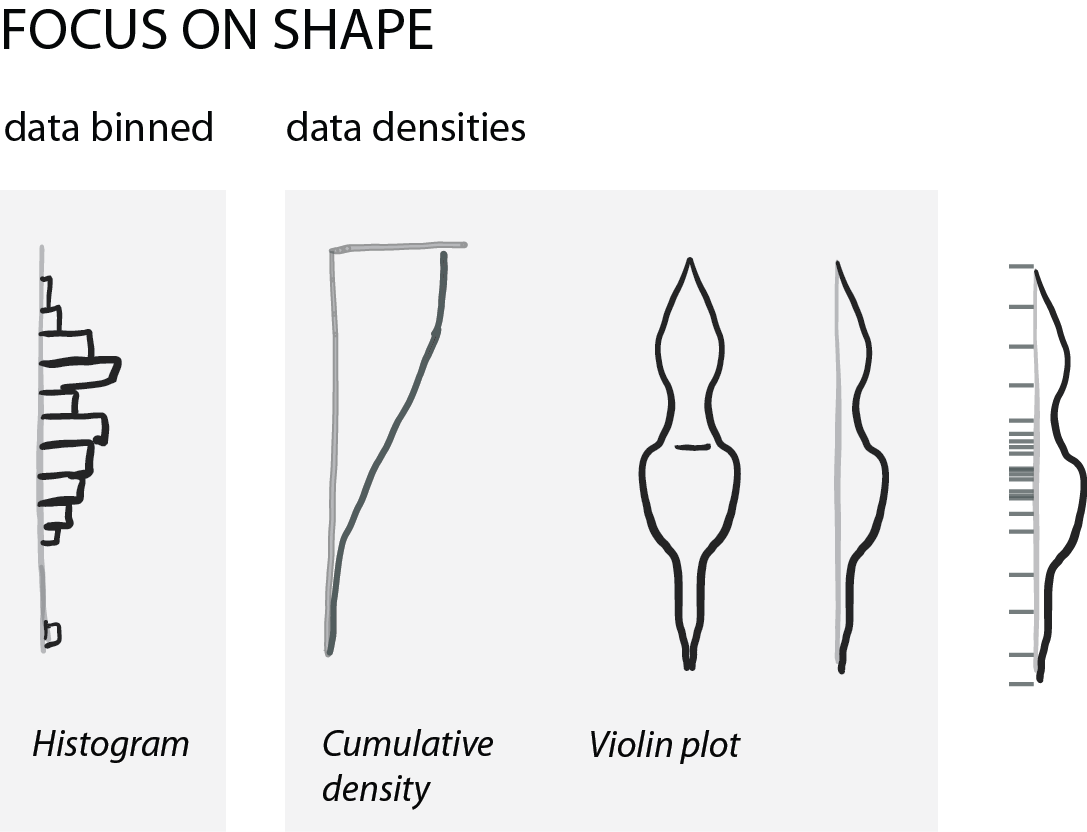
Show shape and unusual features
For normally distributed data, the center and the spread are highly informative. However, in life science we often have bimodal distributions, clusters, or gaps. Then boxplots become very insufficient and might even conceal interesting aspects (if not outright be misleading).
For faithfully showing distributions, histograms have a long history. Here, one has to be very careful with choosing bin sizes: too large or too small bins can greatly distort the histogram shape and result in a misleading chart. Choosing bin sizes is a science in itself, for details see wikipedia – but basically, it again depends on the data shape and sampling depth.
An alternative to histograms are density plots. Density plots show how data are distributed. They become very useful for large data sets. For large data sets individual points can’t be visualized anymore and the eye can’t anymore judge spread intuitively. A rather recent but so far “happy marriage” is the violin plot. Violin plots are a fusion of the boxplot and its summary statistics, with the density/shape of the data (Hintze and Nelson, 1998 doi: 10.1080/00031305.1998.10480559).
 (No Ratings Yet)
(No Ratings Yet) (1 votes)
(1 votes)