The deadline to apply for the 2018 Mouse Development, Stem Cells & Cancer course at Cold Spring Harbor Laboratory (CSHL) is March 15th. If you don’t know much about the course or are on the fence about applying, I want to give you some background about my experiences from 2017, in hopes that I can convince you to apply, too.
The mouse course was founded in 1983 by superstar mouse wranglers Frank Costantini, Brigid Hogan, and Elizabeth Lacy. They recognized the power of emerging techniques to generate transgenic mice and knew they had to create an opportunity to pass these skills on to scientists of all levels and from around the world. Every year, the mouse course brings together 14 students at various stages of their careers and from diverse backgrounds to blast through an intense curriculum packed with hands-on lab work. The course is hosted by the historic Cold Spring Harbor Laboratory, established in 1890: the home of many major scientific discoveries that have shaped modern biology.
In June 2017, I was fortunate to take the mouse course. During my 3 weeks at the CSHL campus, I learned a host of experimental techniques and made tons of new professional contacts and friends. Exposure to these new tools and conversations with the many faculty speakers brought fresh insight into problems I face in my current research, giving me ideas for future projects and collaborations that will benefit me for the rest of my career. I emerged from the mouse course a better scientist equipped, not just with new skills, but the belief I can learn whatever I set out to learn.
Applying for the course
Who should apply for the mouse course? You! Graduate students at all levels, postdoctoral fellows, and new PIs. We had all 3 categories in our class, and we came from different backgrounds within the life sciences: genetics, neuroscience, developmental biology, cancer biology, reproductive biology, and medicine, among others. I am an early graduate student, as were several other participants. The 14 of us had different levels of experience with lab and mouse work, but everyone had the chance to learn and try each new skill, and even the more seasoned students picked up new tricks.
[An aside: If you’re worried you might not have enough experience to apply, consider my case. I had virtually no experience with most techniques taught during the course, and (have to admit), I have a fear of handling mice. As long as you’re willing to grit your teeth and really throw yourself into the course, any level of background experience (or mouse-phobia) is ok.]
For each applicant, the course has different albeit overlapping benefits. For example, junior graduate students can establish a solid experimental foundation for their graduate research. Senior graduate students and new postdocs moving into fresh projects in mouse labs can use the course to quickly gain experience with a new model system. Finally, senior postdocs and new PIs can learn more about the experimental techniques and equipment they’ll need for their new labs, not to mention learning how to design mouse protocols and experiments essential for development of their research programs. In sum: if you want to get ahead in mouse research, you should apply, regardless of your current position along whatever scientific trajectory you are following.
With an intensive, 3-week long course like this, you might find reasons to talk yourself out of applying. Maybe you worry about being away from your regular work schedule for 3 weeks. Sure, 3 weeks away can be disruptive, but in my opinion it’s worth it, and you can plan around it. The mouse course only happens once a year, and your lab will still be there when you come back! Plus, you’ll be returning with way more expertise and new ideas than when you left. If you’re worried about the cost of the course, remember that many institutions offer travel awards and funds to cover the costs. CSHL also provides generous financial aid to students who need it, so there’s no need to worry about funding, and financial concerns should not stop you from applying. Finally, if you’re pretty convinced the mouse course is a good idea, but you don’t know whether this is the right year, or maybe next year would be more convenient, here’s the truth: it will never be perfectly convenient (because science is busy!). That said, applying to the mouse course this year can provide you with the expertise and networking opportunities that will strongly affect how you tackle and succeed next year. So don’t wait.
Taking the course
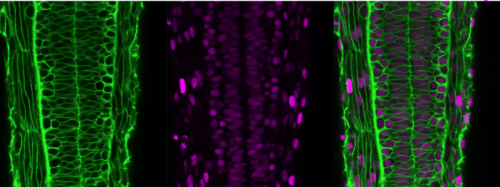
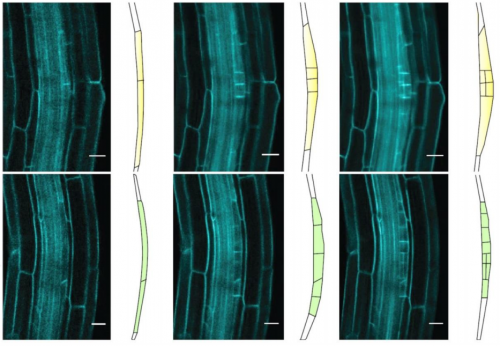
The mouse course is jam-packed with experiments, some lasting several days, and all requiring specific lab supplies and equipment. The course instructors and teaching assistants lovingly and painstakingly organize all the protocols into a tight schedule that flows smoothly and follows evolving themes over the course of the 3 weeks. At the beginning of the course, the experimental focus is on the tools required to create a transgenic mouse; specifically, work with very early embryos. This section includes techniques like electroporation and microinjection of 1- and 2-cell embryos, as well as CRISPR workshops. During my course, we moved along the developmental timeline and towards tools required to manipulate and characterize older embryos, including roller bottle culture and electroporation of mid-gestation embryos, tissue sectioning and staining, and fluorescence-activated cell sorting (FACS). Aside from the lab work, there were guest lectures once or twice a day. Each speaker would cover background topics related to the day’s lab work, and then talk about new research being carried out by their group(s). Usually, the guest speaker would also join us in the lab, and sometimes even show off their skills by demonstrating experiments and giving students hands-on help.
The course also included great ways to connect with other students and faculty. Students presented their work in a mini-seminar early on in the course, which allowed us to get to know each other and understand our various backgrounds and scientific inclinations. We enjoyed weekly social events organized either by the instructors or the students, as well, providing a perfect place to unwind (and sing karaoke, if that’s your thing). One of my favorite parts of the course was the opportunity to meet the guest lecturers. Students would sign up to take speakers out for their “first drink” at the bar; basically, this was a chance to sit down and get to know each other in an informal setting. These get-togethers at the campus pub were a fantastic way to relax after a long day in the lab, not to mention giving us a chance to chat about anything from science and careers to English bulldogs and pink fairy armadillos. (Don’t ask.)
After the course (and some final reasons why you should apply if you aren’t already convinced)
After my 3 weeks at CSHL, I was mentally and physically exhausted, but excited about my work in a way I hadn’t felt before. I left the course equipped with new skills and heaps of reading materials and protocols, all of which I was enthused to share with my labmates back home. In the months since the course, I’ve had the chance to apply protocols taught by the instructors and to follow up on ideas that came up in conversations with guest lecturers. I’ve even spent some idle time dreaming about and planning out a potential post-doc project (which is still several years away for me), based on discussions that took place during the course. The many benefits I reaped overall extended beyond just my own feelings about research; the course was also a place I could start to build a network of scientific contacts and friends. At conferences since my time at CSHL, I have encountered so many familiar faces and reconnected with fellow students, teaching assistants, and guest lecturers. My experiences and friendships from the mouse course have helped me become a part of and feel at home within a larger scientific community.
So: if you want to add an injection of energy and excitement to your scientific research and career that will last for years (and why wouldn’t you?!), then apply. You won’t regret it.
Our latest monthly trawl for developmental biology (and other cool) preprints. Let us know if we missed anything.
Here at the Company of Biologists we are very excited about our soon to launch biology preprints highlighting service, preLights. Before the site goes live, you can sign up for email updates here and follow preLights on Twitter here.
This month was notable for its plant content (see the many beautiful images below), plus a bumper ‘Research practice’ section with discussion of gender, ethics, education, and the future of US science.
The preprints were hosted on bioRxiv, PeerJ, andarXiv. Use these links to get to the section you want:
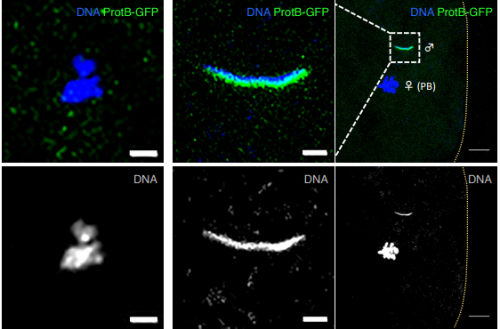
Pcdh18a-positive tip cells instruct notochord formation in zebrafish. Bernadett Bosze, Benjamin Mattes, Claude Sinner, Kathrin Stricker, Victor Gourain, Thomas Thumberger, Sham Tlili, Sabrina Weber, Joachim Wittbrodt, Timothy E Saunders, Uwe Straehle, Alexander Schug, Steffen Scholpp
GSK3 Controls Migration of the Neural Crest Lineage. Sandra G Gonzalez Malagon, Anna Lopez Munoz, Daniel Doro, Triona Bolger, Evan Poon, Elizabeth Tucker, Hadeel Adel Al-Lami, Matthias Krause, Christopher Phiel, Louis Chesler, Karen J Liu
Mapping and dynamics of regulatory DNA during seed development. Alessandra M Sullivan, Andrej A Arsovski, Agnieszka Thompson, Richard Sandstrom, Robert E Thurman, Shane Neph, Audra K Johnson, Shawn T Sullivan, Peter J Sabo, Fidencio V Neri III, Molly Weaver, Morgan Diegel, Jennifer L Nemhauser, John A Stamatoyannopoulos, Kerry L Bubb, Christine Queitsch
The contribution of non-canonical splicing mutations to severe dominant developmental disorders. Jenny Lord, Giuseppe Gallone, Patrick J. Short, Jeremy F. McRae, Holly Ironfield, Elizabeth H. Wynn, Sebastian S. Gerety, Liu He, Bronwyn Kerr, Diana S. Johnson, Emma McCann, Esther Kinning, Frances Flinter, I. Karen Temple, Jill Clayton-Smith, Meriel McEntagart, Sally Ann Lynch, Shelagh Joss, Sofia Douzgou, Tabib Dabir, Virginia Clowes, Vivienne P. M. McConnell, Wayne Lam, Caroline F. Wright, David R. FitzPatrick, Helen V. Firth, Jeffrey C. Barrett, Matthew E. Hurles, on behalf of the DDD study
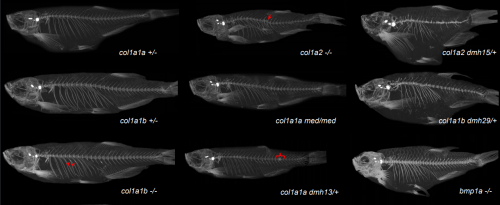
Zebrafish type I collagen mutants faithfully recapitulate human type I collagenopathies. Charlotte Gistelinck, Ronald Y Kwon, Fransiska Malfait, Sofie Symoens, Matthew P Harris, Katrin Henke, Shannon Fisher, Patrick Sips, Brecht Guillemyn, Jan Willem Beck, Petra Vermassen, Hanna De Saffel, MaryAnn Weis, Anne De Paepe, David R Eyre, Andy Willaert, Paul Coucke
A high-quality sequence of Rosa chinensis to elucidate genome structure and ornamental traits. Laurence Hibrand, Tom Ruttink, Latifa Hamama, Ilya Kirov, Deepika Lakhwani, Ning-Ning Zhou, Peter Bourke, Nicolas Daccord, Leen Leus, Dietmar Schulz, Henri Van deGeest, Thamara Hesselink, Katrijn Van Laere, Sandrine Balzergue, Tatiana Thouroude, Annie Chastellier, Julien Jeauffre, Linda Voisine, Sylvain Gaillard, Theo Borm, Paul Arens, Roeland Voorrips, Chris Maliepaard, Enzo Neu, Marcus Linde, Marie-Christine Le Paslier, Aurelie Berard, Remi Bounon, Jeremy Clotault, Nathalie Choisne, Hadi Quesneville, Koji Kawamura, Sebastien Aubourg, Soulaiman Sakr, Rene Smulder, Elio Schijlen, Etienne Bucher, Thomas Debener, Jan De Riek, Fabrice Foucher
The genome of the water strider Gerris buenoi reveals expansions of gene repertoires associated with adaptations to life on the water.David Armisen, Rajendhran Rajakumar, Markus Friedrich, Joshua B Benoit, Hugh M Robertson, Kristen A Panfilio, Seung-Joon Ahn, Monica F Poelchau, Hsu Chao, Huyen Dinh, HarshaVardhan Doddapaneni, Shannon Dugan-Perez, Richard A Gibbs, Daniel ST Hughes, Yi Han, Sandra L Lee, Shwetha C Murali, Donna M muzny, Jiaxin Qu, Kim C Worley, Monica Munoz-Torres, Ehab Abouheif, Francois Bonneton, Travis Chen, Christopher Childers, Andrew G Cridge, Antonin JJ Crumiere, Amelie Decaras, Elise M Didion, Elizabeth Duncan, Elena N Elpidina, Marie-Julie Fave, Cedric Finet, Chris GC Jacobs, Alys Jarvela, Emily J Jennings, Jeffery W Jones, Maryna P Lesoway, Mackenzie Lovegrove, Alexander Martynov, Brenda Oppert, Angelica Lilico-Ouachour, Arjuna Rajakumar, Peter N Refki, Andrew J Rosendale, Maria Emilia Santos, William Toubiana, Maurijn van der Zee, Iris M Vargas Jentzsch, Aidamalia Vargas Lowman, Severine Viala, Stephen Richards, Abderrahman Khila
Embracing heterogeneity: Building the Tree of Life and the future of phylogenomics. Gustavo A Bravo, Alexandre Antonelli, Christine D Bacon, Krzysztof Bartoszek, Mozes Blom, Stella Huynh, Graham Jones, L. Lacey Knowles, Sangeet Lamichhaney, Thomas Marcussen, Hélène Morlon, Luay Nakhleh, Bengt Oxelman, Bernard Pfeil, Alexander Schliep, Niklas Wahlberg, Fernanda Werneck, John Wiedenhoeft, Sandi Willows-Munro, Scott V Edwards
Observing the Cell in Its Native State: Imaging Subcellular Dynamics in Multicellular Organisms.Tsung-li Liu, Srigokul Upadhyayula, Daniel E Milkie, Ved Singh, Kai Wang, Ian A Swinburne, Kishore R Mosaliganti, Zach M Collins, Tom W Hiscock, Jamien Shea, Abraham Q Kohrman, Taylor N Medwig, Daphne Dambournet, Ryan Forster, Brian Cunniff, Yuan Ruan, Hanako Yashiro, Steffen Scholpp, Elliot M Meyerowitz, Dirk Hockemeyer, David G Drubin, Benjamin L Martin, David Q Matus, Minoru Koyama, Sean G Megason, Tom Kirchhausen, Eric Betzig
Digital museum of retinal ganglion cells with dense anatomy and physiology. J. Alexander Bae, Shang Mu, Jinseop S. Kim, Nicholas L. Turner, Ignacio Tartavull, Nico Kemnitz, Chris S. Jordan, Alex D. Norton, William M. Silversmith, Rachel Prentki, Marissa Sorek, Celia David, Devon L. Jones, Doug Bland, Amy L. R. Sterling, Jungman Park, Kevin L. Briggman, H. Sebastian Seung, the EyeWirers
Resolving the Full Spectrum of Human Genome Variation using Linked-Reads. Patrick Marks, Sarah Garcia, Alvaro Martinez Barrio, Kamila Belhocine, Jorge Bernate, Rajiv Bharadwaj, Keith Bjornson, Claudia Catalanotti, Josh Delaney, Adrian Fehr, Brendan Galvin, Haynes Heaton, Jill Herschleb, Christopher Hindson, Esty Holt, Cassandra B. Jabara, Susanna Jett, Nikka Keivanfar, Sofia Kyriazopoulou-Panagiotopoulou, Monkol Lek, Bill Lin, Adam Lowe, Shazia Mahamdallie, Shamoni Maheshwari, Tony Makarewicz, Jamie Marshall, Francesca Meschi, Chris O’keefe, Heather Ordonez, Pranav Patel, Andrew Price, Ariel Royall, Elise Ruark, Sheila Seal, Michael Schnall-Levin, Preyas Shah, Stephen Williams, Indira Wu, Andrew Wei Xu, Nazneen Rahman, Daniel MacArthur, Deanna M. Church
Equivalent high-resolution identification of neuronal cell types with single-nucleus and single-cell RNA-sequencing. Trygve E Bakken, Rebecca D Hodge, Jeremy M Miller, Zizhen Yao, Thuc N Nguyen, Brian Aevermann, Eliza Barkan, Darren Bertagnolli, Tamara Casper, Nick Dee, Emma Garren, Jeff Goldy, Lucas T Gray, Matthew Kroll, Roger S Lasken, Kanan Lathia, Sheana Parry, Christine Rimorin, Richard H Scheuermann, Nicholas J Schork, Soraya I Shehata, Michael Tieu, John W Phillips, Amy Bernard, Kimberly A Smith, Hongkui Zeng, Ed S Lein, Bosiljka Tasic
Mapping nonapoptotic caspase activity with a transgenic reporter in mice. Peter Nicholls, Thomas Pack, Nikhil Urs, Sunil Kumar, Gabor Turu, Evan Calabrese, Wendy Roberts, Ping Fan, Valeriy Ostapchenko, Monica Guzman, Flavio Beraldo, Vania Prado, Marco Prado, Ivan Spasojevic, Joshua Snyder, Kafui Dzirasa, G. Allan Johnson, Marc Caron
Harmonizing semantic annotations for computational models in biology. Maxwell L Neal, Matthias König, David Nickerson, Goksel Mısırlı, Reza Kalbasi, Andreas Dräger, Koray Atalag, Vijayalakshmi Chelliah, Michael Cooling, Daniel L Cook, Sharon Crook, Miguel de Alba, Samuel H Friedman, Alan Garny, John H Gennari, Padraig Gleeson, Martin Golebiewski, Michael Hucka, Nick Juty, Nicolas Le Novère, Chris Myers, Brett G Olivier, Herbert M Sauro, Martin Scharm, Jacky L Snoep, Vasundra Touré, Anil Wipat, Olaf Wolkenhauer, Dagmar Waltemath
Opportunities And Obstacles For Deep Learning In Biology And Medicine. Travers Ching, Daniel S. Himmelstein, Brett K. Beaulieu-Jones, Alexandr A. Kalinin, Brian T. Do, Gregory P. Way, Enrico Ferrero, Paul-Michael Agapow, Michael Zietz, Michael M Hoffman, Wei Xie, Gail L. Rosen, Benjamin J. Lengerich, Johnny Israeli, Jack Lanchantin, Stephen Woloszynek, Anne E. Carpenter, Avanti Shrikumar, Jinbo Xu, Evan M. Cofer, Christopher A Lavender, Srinivas C Turaga, Amr M Alexandari, Zhiyong Lu, David J. Harris, Dave DeCaprio, Yanjun Qi, Anshul Kundaje, Yifan Peng, Laura K. Wiley, Marwin H. S. Segler, Simina M Boca, S. Joshua Swamidass, Austin Huang, Anthony Gitter, Casey S. Greene
Recently, Nature published my correspondence “Dispense with redundant P values”. It highlights my concern that p-values are often calculated because “everybody does it”. This reminded me of the mechanical repetition that parrots are well-known for (footnote 1). Parroting of p-value reporting should stop and I suggest to only present a p-value in a figure if it is necessary for interpretation.
During the editing process of my contribution a specific example of a redundant p-value was removed. The reasoning was that it seemed unfair to single out only one paper. I agreed and I would like to stress that parroting of p-value reporting is not restricted to a specific paper, a specific issue of Nature or to some specific journal. It’s just that I found it very ironic that in the same issue of Nature that proposes “Five ways to fix statistics” (Leek et al., 2017) there are several clear examples of figures (in different papers) with meaningless p-values.
Since I am convinced that an example will clarify my point, I have extracted the data (see footnote 2) from one of the papers in the aforementioned issue (without disclosing the nature of the paper). I performed a t-test (two-tailed, unequal variances) as described in the paper and reproduced the p-value (footnote 3). The resulting figure is shown below on the left and closely mimics the figure of the original paper.
Clearly, there is a large difference between the ’Control’ and ‘Treated’ condition, reflecting a large effect of the treatment. To reach that conclusion, there is no need for a p-value. Moreover, the p-value does not convey any relevant information for interpretation of the figure. As such, the p-value qualifies as chartjunk (E.R. Tufte, 1983) and should be omitted. This will generate a cleaner figure (shown above on the right) that emphasizes the data.
The problems with p-values are larger than their meaningless use in figures whenever the effects are large. The way that p-values are defined has some confusing backward logic (footnote 4). Consequently, p-values are often misinterpreted (Greenland et al., 2016, Lakens, 2017) or misused as a ‘measure of credibility’ (Goodman, 2016). The misconception that p-values represent the strength of evidence is reinforced by catogerizing p-values, e.g. by using increasing number of asterisks (e.g. * P < 0.05; ** P < 0.01; *** P < 0.001). P-values cannot be used as a rating system (Wasserstein and Lazar, 2016) and categories should be avoided at all times.
To avoid the unnecessary, and at times misleading, use of p-values, the mechanical repetition of current practices should stop. Whether p-values are important for the interpretation of the figure should be a central question. Before that question can be answered, the correct definition of a p-value needs to be thoroughly understood. In addition, the correct interpretation and common misconceptions (Greenland et al., 2016) of p-values should be considered. I hope that careful reflection on the meaning of p-values will decrease their use and improve figures.
Acknowledgments: I am indebted to Marten Postma for the many discussions about statistical concepts and applied statistics, that have increased my understanding of the topic.
Footnotes
Footnote 1: The original title of my correspondence was “Prevent p-value parroting”. This title was changed to “Dispense with redundant P values” by Nature after I returned the proofs and without consulting me.
Footnote 2: The raw data that I extracted is listed below in csv format:
Footnote 3: This is an example of yet another questionable practice, i.e. calculating a p-value for a dataset with only a couple of datapoints per condition. Ironically, several examples can be found in the aforementioned issue while this matter has also been addressed previously by David Vaux (2012) in Nature (and by many others as well).
Footnote 4: The p-value is the probability of the observed data (or more extreme values), assuming that the null-hypothesis (there is zero difference between the two conditions) is true.
Do you play bass? Guitar? Keys? Sax? Brass? We’re putting together an SDB all-star jazz combo to play at the SDB meeting in Portland, and we’re looking for a couple more players! Feel free to reach out to me at david.matus@stonybrook.edu if you’re interested!
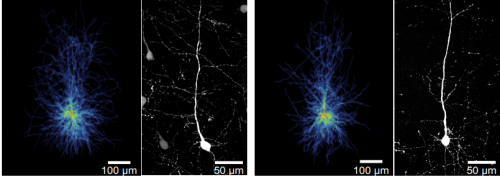
Adult neurogenesis is distinct from embryonic forms of neurogenesis in that neuronal development underlies extrinsic regulation, from physiological to pathological. This regulation targets different stages of neuronal development, between cell cycle entry and proliferation of radial glia-like stem cells to the lasting integration of the newly generated neurons. There are obvious feedback loops in that regulation of neurogenesis might alter behavior, which in turn acts back on adult neurogenesis. But how does this all work? How does regulation occur across scales? What constitutes ‘causes’ and ‘mechanisms’ in such complex system? This conference uses these key questions as entry point into discussions about how activity and development are linked in adult neurogenesis.
Free registration up for grabs!
Abcam and the Node are looking for an official meeting reporter to attend this meeting. The Reporter will be responsible for providing regular updates of interesting talks/discussions for twitter/tweets (by Abcam), plus a meeting report of their experience and the sights and sounds of the meeting (for publishing on The Node and Abcam website). Check out the article written by the last conference’smeeting reporter winner, Nambirajan Govindarajan.
To apply to be the meeting reporter, please send a short paragraph (written for a scientific audience, max. 200 words) toevents@abcam.com, telling us what makes adult neurogenesis fascinating to you! Application deadline: February 2, 2018. The winner will receive free registration to the meeting (travel and accommodation not included).
Meeting information
Organizer: Gerd Kempermann (CRTD – Technische Universität Dresden)
Confirmed speakers: Nora Abrous, Laure Bally-Cuif, Benedikt Berninger, Michael Brand, Federico Calegari, Fred H. Gage, Sebastian Jessberger, H. Georg Kuhn, Alejandro Schinder, Jason Snyder, Sandrine Thuret, and Henriette van Praag
Call for abstracts: Abstracts are invited for short talks or poster presentations and can be submitted during registration (see meeting website for more details)
Poster session at the 2015 meeting
Terms and conditions
Effective November 2017
The contest is open to persons aged 21 years or over, except employees of, and consultants to Abcam, their families, government employees and any third party
associated with administration of the contest.
The contest is free to enter and no purchase is necessary. VOID WHERE PROHIBITED BY LAW.
All entries must be submitted by February 2, 2018. Entries received after this time will not be accepted.
The winner will be selected by a judging panel and announced after the entry deadline of February 2, 2018. The winner will receive one free registration to the Adult Neurogenesis 2018 conference.
The prize is non-exchangeable, non-transferable and no cash alternative is offered.
The decision of the judges regarding any aspect of the contest is final and binding and no correspondence will be entered into about it.
By entering this competition, you are agreeing to be bound by these terms and conditions, and you are representing and warranting to Abcam that you meet
the eligibility requirements of the contest. You are also agreeing to be contacted by Abcam in relation to the competition. We will not share your details with
anyone else except the winner’s announcement.
By entering this competition, you agree to receive further information regarding Abcam products and services. You can opt out of receiving communications
from us at any time by unsubscribing from our newsletter.
Insofar as is permitted by law, Abcam reserves the right to hold void, cancel, suspend, or amend the contest where it becomes necessary to do so.
Insofar as is permitted by law, Abcam, its agents or distributors will not in any circumstances be responsible or liable to compensate the winner or accept any
liability for any loss, damage, personal injury or death occurring as a result of taking up the prize except where it is caused by the negligence of Abcam, its agents or distributors or that of their employees. Your statutory rights are not affected.
Personal data supplied during this contest maybe passed onto third party suppliers only insofar as required for fulfilment/delivery/arrangement of the prize.
This offer is void if it conflicts with the policy of the recipient.
The contest will be governed by law of England and Wales and entrants to the contest agree to submit to binding arbitration to resolve any disputes relating to the contest.
Abcam plc located at 330 Cambridge Science Park, Cambridge CB4 0FL is the sponsor of this contest.
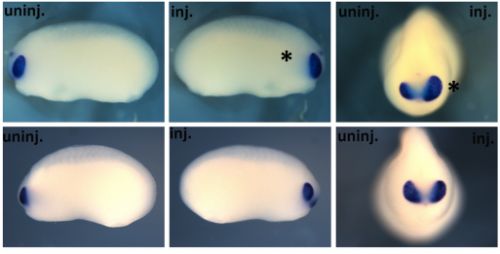
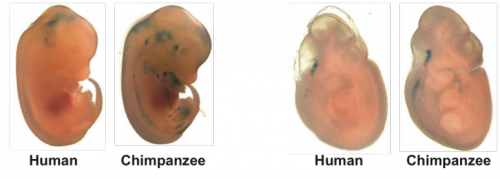
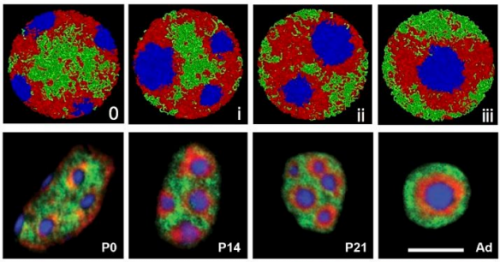
With new findings, scientists may be poised to break a long impasse in research on Huntington’s disease, a fatal hereditary disorder for which there is currently no treatment.
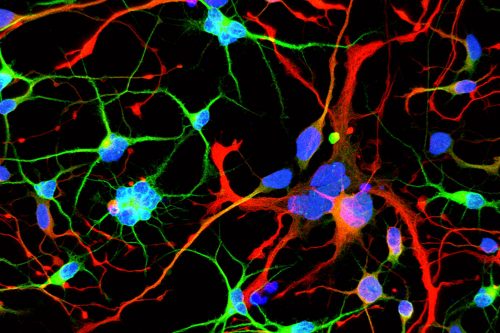
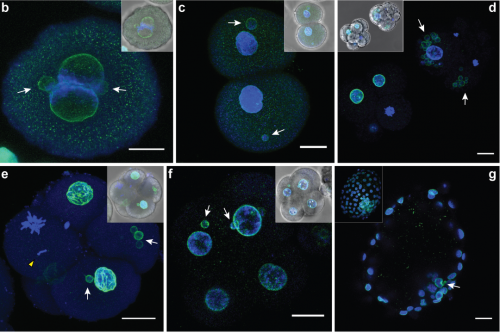
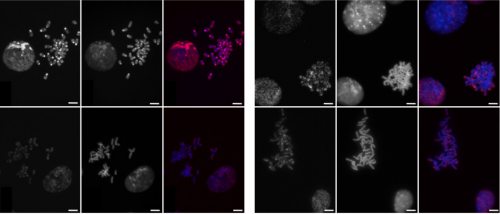
Huntington’s neurons show multiple nuclei (blue) within the same cell, and other signs of trouble, long before symptoms emerge.
One in 10,000 Americans suffer from the disease, and most begin to show symptoms in middle age as they develop jerky movements—and as these patients increasingly lose brain neurons, they slide into dementia. But the new research suggests that these symptoms may be a late manifestation of a disease that originates much earlier, in the first steps of embryonic development.
A team at Rockefeller led by Ali Brivanlou, the Robert and Harriet Heilbrunn Professor, developed a system to model Huntington’s in human embryonic stem cells for the first time. In a report published in Development, they describe early abnormalities in the way Huntington’s neurons look, and how these cells form larger structures that had not previously been associated with the disease.
“Our research supports the idea that the first domino is pushed soon after fertilization,” Brivanlou says, “and that has consequences down the line. The final domino falls decades after birth, when the symptoms are observable.”
The findings have implications for how to best approach treating the disorder, and could ultimately lead to effective therapies.
A new tool
Huntington’s is one of the few diseases with a straightforward genetic culprit: One hundred percent of people with a mutated form of the Huntingtin (HTT) gene develop the disease. The mutation takes the form of extra DNA, and causes the gene to produce a longer-than-normal protein. The DNA itself appears in the form of a repeating sequence, and the more repeats there are, the earlier the disease sets in.
Research on Huntington’s has thus far relied heavily on animal models of the disease, and has left many key questions unanswered. For example, scientists have not been able to resolve what function the HTT gene serves normally, or how its mutation creates problems in the brain.
“We started seeing things that were completely unexpected”
Suspecting that the disease works differently in humans, whose brains are much bigger and more complex than those of lab animals, Brivanlou, along with research associates Albert Ruzo and Gist Croft, developed a cell-based human system for their research. They used the gene editing technology CRISPR to engineer a series of human embryonic stem cell lines, which were identical apart from the number of DNA repeats that occurred at the ends of their HTT genes.
“We started seeing things that were completely unexpected,” says Brivanlou. “In cell lines with mutated HTT, we saw giant cells. It looked like a jungle of disorganization.”
When cells divide, they typically each retain one nuclei. However, some of these enlarged, mutated cells flaunted up to 12 nuclei—suggesting that neurogenesis, or the generation of new neurons, was affected. The disruption was directly proportional to how many repeats were present in the mutation: The more repeats there were, the more multinucleated neurons appeared.
“Our work adds to the evidence that there is an unrecognized developmental aspect to the pathology,” Brivanlou says. “Huntington’s may not be just a neurodegenerative disease, but also a neurodevelopmental disease.”
Toxic or essential?
Treatments for Huntington’s have typically focused on blocking the activity of the mutant HTT protein, the assumption being that the altered form of the protein was more active than normal, and therefore toxic to neurons. However, Brivanlou’s work shows that the brain disruption may actually be due to a lack of HTT protein activity.
To test its function, the researchers created cell lines that completely lacked the HTT protein. These cells turned out to be very similar to those with Huntington’s pathology, corroborating the idea that a lack of the protein—not an excess of it—is driving the disease.
“We should rethink our approach to treating Huntington’s”
The findings are significant, Brivanlou notes, since they indicate that existing treatments that were designed to block HTT activity may actually do more harm than good.
“We should rethink our approach to treating Huntington’s,” he says. “Both the role of the HTT protein and the timing of treatment need to be reconsidered; by the time a patient is displaying symptoms, it may be too late to medicate. We need to go back to the earliest events that trigger the chain reaction that ultimately results in disease so we can focus new therapies on the cause, not the consequences.”
The researchers hope their new cell lines will be a useful resource for studying the cellular and molecular intricacies of Huntington’s further, and suggest they may provide a model for examining other diseases of the brain that are specific to humans.
# # #
Contact:
Katherine Fenz, Media Relations Manager at The Rockefeller University
Tel: +1 212 327 7913; Email: kfenz@rockefeller.edu
About The Rockefeller University
The Rockefeller University is the world’s leading biomedical research university and is dedicated to conducting innovative, high-quality research to improve the understanding of life for the benefit of humanity. Our 82 laboratories conduct research in neuroscience, immunology, biochemistry, genomics, and many other areas, and a community of 1,800 faculty, students, postdocs, technicians, clinicians, and administrative personnel work on our 14-acre Manhattan campus. Our unique approach to science has led to some of the world’s most revolutionary and transformative contributions to biology and medicine. During Rockefeller’s 115-year history, 25 of our scientists have won Nobel Prizes, 22 have won Albert Lasker Medical Research Awards, and 20 have garnered the National Medal of Science, the highest science award given by the United States.
DanStem seeks two laboratory technicians or biomedical laboratory scientists for tasks related to a research project in stem cells and type 1-diabetes.
About DanStem
The Novo Nordisk Foundation Center for Stem Cell Biology – DanStem was established as a result of a series of international recruitments coupled with internationally recognized research groups focused on insulin producing beta cells and cancer research already located at the University of Copenhagen. DanStem addresses basic research questions in stem cell and developmental biology and has activities focused on the translation of promising basic research results into new strategies and targets for the development of new therapies for cancer and chronic diseases such as diabetes and liver failure. Learn more about DanStem at http://danstem.ku.dk/
Job description
You will be responsible for growing and expanding human stem cells and in close collaboration with the group’s researchers; you will help develop new methods and protocols for differentiation of hPSC. You will also perform basic characterization of hPSC using karyotyping, immunohistochemistry, qPCR and FACS, to ensure quality and reproducibility. In addition, you will be responsible for maintaining equipment in the hPSC laboratory.
The goal of the research project is to develop therapeutic active islet-like aggregate for future cell therapy in clinical phase 1 trials.
Your qualifications
We expect you to hold a degree as either laboratory technician or biomedical laboratory scientist and that you have several years of experience working as a technician in a research lab. Furthermore, we expect the following qualifications:
Minimum 5 years of experience in cellular work, preferably with stem cells
Experience with transfection, immunohistochemistry, qPCR and FACS
That you work systematically and are flexible
Good cooperation and communications skills as the project is carried out within a team of scientists and technicians
Good English skills in both speech and writing, as our working language is primarily English
For further information, please contact Professor Henrik Semb by e-mail semb@sund.ku.dk
Terms of salary and employment
The employments are planned to start as soon as possible upon agreement with the chosen candidates. The place of work is at DanStem, University of Copenhagen, Blegdamsvej 3B, Copenhagen. The average working hours are 37 hours per week. The positions are time limited to the end of 2020 with a possibility of further employment.
Terms of salary and employment are in accordance with the collective agreement between the Danish Ministry of Finance and the Organizations of Public Employees – the Governmental area (OAO-S) and the HK organization agreement with placement in salary group 5, or The Danish Association of Biomedical Laboratory Scientists and the Organization Agreement for Biomedical laboratory scientists.
Furthermore a salary supplement can be negotiated based on individual qualifications.
Application
Send your application electronically by clicking “Apply online” below or via this advertisement on http://employment.ku.dk/staff/. The application must include the following documents/attachments:
Motivated letter of application (max 1 page)
Curriculum vitae incl. education, experience, previous employments, language skills and other relevant skills
Certified copy of diplomas/degree certificate(s)
Letter of recommendation and/or contact details of referees
Application deadline: 20 February 2018
The University of Copenhagen wishes to reflect the diversity of society and welcomes applications from all qualified candidates regardless of personal background.
preLights is a new preprint highlights service supported by The Company of Biologists, the not-for-profit publisher which hosts the Node via one of its five journals, Development.
The site is currently being built and will be launched soon, but before then you can keep up-to-date by signing up for news here:
The Company of Biologists Gurdon Summer Studentship scheme was initiated by the BSDB in 2014 to provide highly motivated undergraduate students with an opportunity to engage in practical research during their summer vacation. Each year, ten successful applicants spend eight weeks in the research laboratories of their choices. For the last four years, the quality of projects and feedback we receive has been outstanding – as is clearly illustrated by reports from previous awardees.
The new round of applications is now open and will close Friday, 30 March 2018. Please, look at our dedicated web page for eligibility and application procedures.
Our recent paper in “Nature”[1] deconstructs molecular arguments that have been used to homologize bilaterian nerve cords. Our work illustrates well the strength of the comparative approach and the broad sampling across the animal tree of life that we use in my research group at the Sars Centre for Marine Molecular Biology.
Evo-Devo branched off as a discipline from Developmental Biology. Therefore Evo-Devo inherited concepts and narratives from Developmental Biology that dominated over those of its cousin-discipline Evolutionary Biology. This was in part because Evo-Devo began with the comparison of data from a handful of well-studied genetic models, such as Drosophila, C. elegans, and vertebrates, which were used as a quite limited representation of the animal diversity. At this time, Evo-Devo was more “Devo” than “Evo”.
I entered Evo-Devo as a trained comparative zoologist who was –and still is– fascinated by animal diversity, evolution, and the many developmental pathways animals can display. I was attracted by the new discoveries of developmental biology, but being well familiar with the comparative method, I was also skeptical about their potential evolutionary implications, given the low taxon sampling (that is, the sparse number of species investigated across a tree).
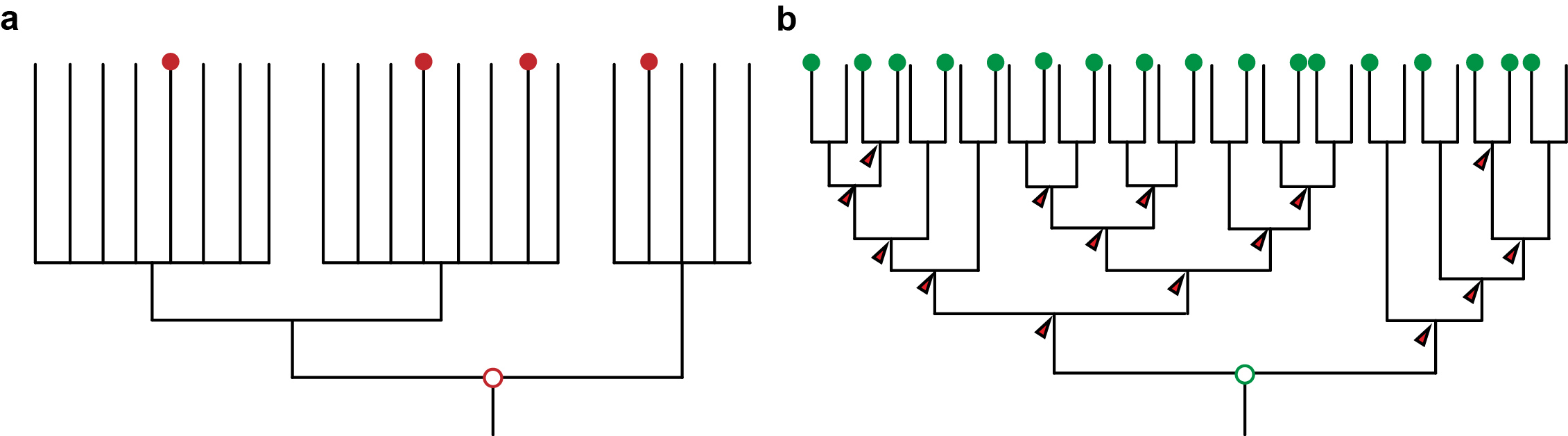
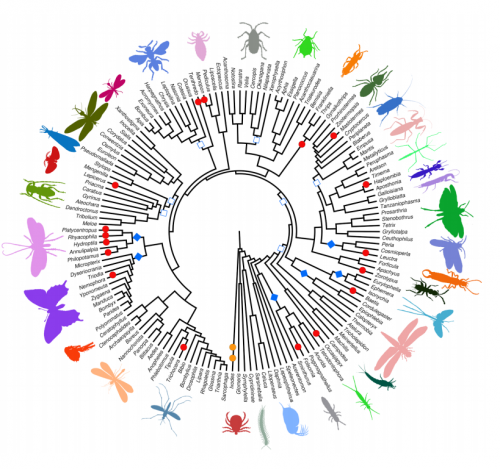
At about the same time, molecular phylogenetic methods were shaking and uprooting the Animal Tree of Life. New molecular trees provided evidence for the monophyletic Ecdysozoa [2] (e.g. nematodes and arthropods), Lophotrochozoa [3] (e.g. annelids, molluscs) and Deuterostomia (e.g. urchins and frogs). Traditional relationships, such as the close relationship of annelids and arthropods, were questioned. However, the order of the internal branches remained unresolved. Such polytomies, together with the low taxon sampling by that time, obscured evolutionary conclusions about ancestral characters in these major groups, and thus hampered the detection of evolutionary change in the animal tree of life (Figure 1a). Instead, this situation sparked anthropogenic speculations.
Evo-Devo was thus influenced by two problems: the low number of studied species, and the lack of a phylogenetic framework that allows hypotheses testing.
This situation rapidly changed in areas of the Animal Tree of Life where taxonomical sampling was denser, and where the phylogenetic relationships were better resolved – such as in the vertebrates, and partly in the arthropods. This favored hypothesis testing, and improved our understanding of morphological evolution, as seen for instance with the evolution of limbs from fins [4]. But scientists interested in squishy invertebrates had much harder times.
Advances in sequencing technologies, imaging, and molecular methods have radically changed Evo-Devo, opening up new opportunities, and becoming a continuous source of innovation in evolutionary biology. Next generation sequencing has hugely impacted phylogenomics, allowing much better resolved animal relationships, thus providing the necessary phylogenetic frame to test evolutionary hypotheses [5]. These same sequencing technologies also make it now much easier to take animals from the field and implement molecular techniques faster for their study, which enables a much denser taxonomical sampling of animal diversity. And as sampling more data points provides a better measurement in physics, sampling more species provide a better understanding of evolution (Figure 1b). A denser sampling within a resolved phylogeny allows a more solid reconstruction of ancestral characters and offers a much better picture about character changes during evolution [6]. This approach is common practice in evolutionary biology [7] – but has not yet been extensively used in Evo-Devo.
Figure 1: a) An unresolved phylogeny with only few taxa sampled (red dots) can only provide vague and untestable hypotheses. b) A well resolved phylogeny with large sampling allows the reconstruction at many more nodes and thus also insights about the evolution along the branches that connect the nodes. It also provides a framework for testing hypotheses.
The approach allows to test previous hypotheses that have been developed when low taxon sampling and/or unresolved phylogenies prevailed. For instance, our study of nine species published in “Nature” we show that the expression of the transcription factors traditionally used to call for a single condensed nerve cord in the last common ancestor of bilateral animals [8] evolved not only independently from each other, but also independently from the nervous system architecture and neuronal cell types. This implies that statements such as “this organ is so complex, it is unlikely to have evolved twice” are human presumptions, and that shared similarities might also be the result of independent adaptation processes (that is homoplasy). The comparative method strongly reduces human bias and also helps to avoid wrong conclusions [9].
The advantages of our approach not only extend to the study of organ system evolution, but simply affects all other characters that undergo evolutionary changes: tissues, cell types, developmental modes, genome architectures, gene expression [9]. Increasing the number of research organisms and the interpretation of the data on the basis of a resolved phylogeny provides a powerful toolkit for understanding evolution. It opens up the horizon for addressing new questions on when and how things changed in evolution, which is indeed where our curiosity lays.
 (4 votes)
(4 votes)









 (No Ratings Yet)
(No Ratings Yet)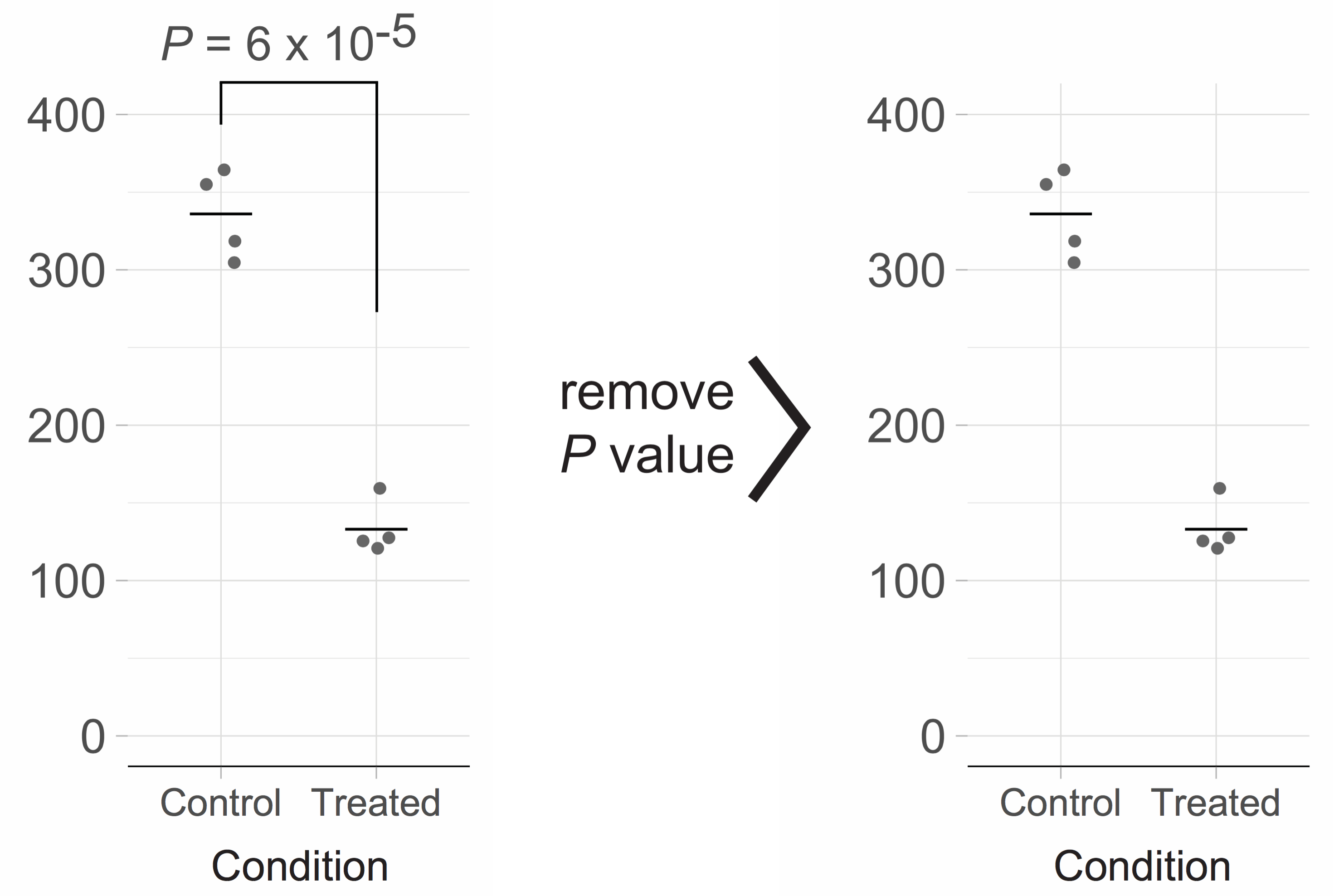
 (19 votes)
(19 votes)