The Novo Nordisk Foundation Center for Stem Cell Biology – DanStem has been established as a result of a series of international recruitments coupled with internationally recognized research groups focused on insulin producing beta cells and cancer research already located at the University of Copenhagen. DanStem addresses basic research questions in stem cell and developmental biology and has activities focused on the translation of promising basic research results into new strategies and targets for the development of new therapies for cancer and chronic diseases such as diabetes and liver failure.
The goal of this project is to engineer therapeutically active islet-like aggregates for future cell therapy phase 1 trials in Type 1 Diabetes (T1D).
Job Description:
We are looking for a postdoctoral candidate with a strong cell biological and cell signaling background in directed differentiation of human pluripotent stem cells. Experience in differentiation towards pancreatic lineages is a merit. The candidate is expected to work on the engineering of human pluripotent stem cell-derived aggregates with functional properties close to human islets of Langerhans. The functionality and therapeutic potential of the aggregates will be tested in vitro and in vivo after transplantation into mice. The candidate is expected to use state-of-the-art genetic, molecular and cell biological, and tissue engineering experimental strategies. The candidate will work together with a dedicated team of scientists and technicians who together will tackle bottle-necks towards implementing the phase 1 clinical trials in T1D.
Qualifications:
The candidate is required to hold a PhD degree in pluripotent stem cell or developmental biology. A few years of postdoctoral experience in the same area are a merit. The candidate should also have hands-on experience in human pluripotent stem cell maintenance and differentiation, 3D culture of pluripotent stem cells, various cell and molecular biological methods, flow cytometry and live-cell imaging. Finally, we are looking for applicants with a good record of peer reviewed scientific publications, grant writing skills and an interest in team work.
Employment Conditions:
The position is for 2 years with a possible extension. The employment is planned to start as soon as possible or upon agreement with the chosen candidate.
The terms of employment are set according to the Agreement between the Ministry of Finance and The Danish Confederation of Professional Associations or other relevant professional organization. The position will be at the level of postdoctoral fellow and the basic salary according to seniority is 32.700-34.400 DKK/month. A supplement could be negotiated, dependent on the candidate´s experience and qualifications. In addition, a monthly contribution of 17.1% of the salary is paid into a pension fund.
Applicants recruited from abroad are eligible for a special researcher taxation scheme. In all cases, the ability to perform the job will be the primary consideration, and thus we encourage all – regardless of their personal background and status – to apply.
The application must be submitted online and in English, by clicking on “Apply online” below.
The closing date for applications is 23.59pm, 22th January 2018.
The application must include:
Cover letter detailing the basis on which the applicant scientific qualifications meet the requirements for this position.
Curriculum vitae.
List of references (full address, incl. email and phone number)
Diplomas – all relevant certificates.
List of publications.
Application procedure:
After the expiry of the deadline for applications, the authorized recruitment manager selects applicants for assessment on the advice of the Appointments Committee. All applicants are then immediately notified whether their application has been passed for assessment by an expert assessment committee. Selected applicants are notified of the composition of the committee and each applicant has the opportunity to comment on his/her assessment. You may read about the recruitment process on http://employment.ku.dk/faculty/recruitment-process
DanStem and The University of Copenhagen wish to reflect the diversity of society and welcome applications from all qualified candidates, regardless of personal background.
Founded in 1479, the University of Copenhagen is the oldest university in Denmark. With 37,000 students and 9,000 employees, it is among the largest universities in Scandinavia and one of the highest ranking in Europe. The University consists of six faculties, which cover Health and Medical Sciences, Humanities, Law, Science, Social Sciences and Theology.
Part of the International Alliance of Research Universities (IARU), and among Europe’s top-ranking universities, the University of Copenhagen promotes research and teaching of the highest international standard. Rich in tradition and modern in outlook, the University gives students and staff the opportunity to cultivate their talent in an ambitious and informal environment. An effective organization – with good working conditions and a collaborative work culture – creates the ideal framework for a successful academic career.
Science communication (scicomm) has become a buzz term in the current science landscape. I fully support its importance and have been a scicomm “activist” for over 6 years. My initiatives promote the enormous importance of Developmental Biology as a key discipline of the biomedical sciences (see our advocacy campaign); within this context, I put specific emphasis on the use of Drosophila as a most powerful tool to advance concepts and fundamental understanding (see our recent publication).
Defining scicomm and its many different facets is not easy. In my interpretation, it means establishing dialogue (in a variety of modalities) between practicing scientists (called “scientists” from now on) and a wide range of target groups to resolve reciprocal misconceptions, learn from one another and achieve mutual benefit.
Direct engagement of scientists with the wider public is usually done at science fairs, school visits, public presentations, etc. Many of these activities tend to be short-lived one-offs that reach a limited amount of people and, at first glance, may appear to be relatively low on ‘impact.’ However, there are opportunities if we open up to dialogue! Genuine engagement with pupils, teachers or visitors at a science fair can be a sobering exercise: the responses you receive make absolutely clear, what topics and arguments come across, excite and are perceived as being important – and it is the “thumbs down” responses which should make us think about our own science! To put it bluntly: if you cannot explain your science and its importance, you either have not thought hard enough and need to refine your explanations, or you are doing the wrong thing and should consider changes in your research direction! If we use scicomm in this way, it will help to align our science with the wider society in the long term; this can be taken even a step further through citizenscience and other forms of actively involving the public in our research. Furthermore, it will provide the refined explanations and elevator pitches with which to advocate our science and engage with journalists to achieve improved and helpful press outlets. Even more, they provide profound rationales and simple narratives that will be as powerful when presenting our own science in grant applications, talks and publications.
Important aspects of scicomm lie in the hands of journalists or teachers. Scientists tend to have little influence on article or school lesson contents, although journalists reach audiences in their millions, and students at schools are the potential future scientists and will constitute and shape the future society which we would wish to embrace science. To engage in true dialogue ….
To read the full content, please see the original publication of this post on PLoS blogs.
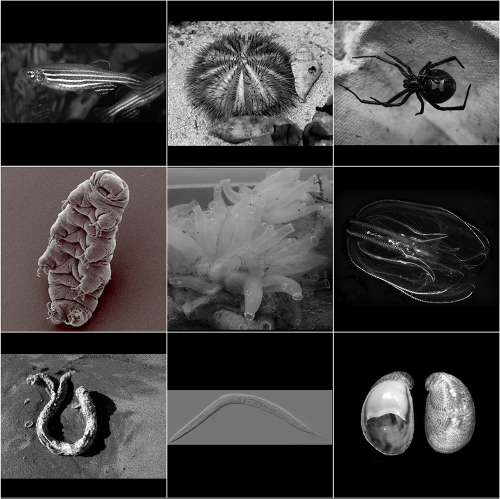
Here at the Node we are always on the lookout for beautiful developmental biology images and videos, and love our science art (see here, here, here, here and here!).
So we were excited to hear FASEB announce the winners of their 2017 BioArt competition. As well as gorgeous images (see below) there was this wonderful video – the first 24 hours of embryo development in 9 animal species. I’d recommend full screen HD/4K for full embryonic immersion!
It’s a wonderful piece of comparative embryology, maybe one for all introductory developmental biology courses! And look what they turn in to:
Image cobbled together in house. All images from Wikipedia, aside from the Chaetopterus and Crepidula that come from the always useful World Register of Marine Species (WoRMS) site
Here’s a gallery featuring the development-y winning images (click for more info):
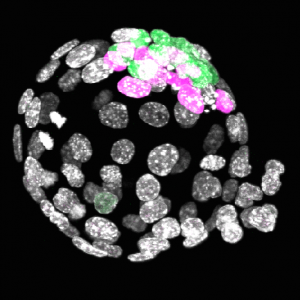
By Marina Venero Galanternik, Daniel Castranova, Tuyet Nguyen, and Brant M. Weinstein. This microscopy image shows that Fluorescent Granular Perithelial cells (FGPs, in green) are closely associated with the blood vessels (red) that surround an adult zebrafish’s brain. FGPs are novel type of cell found in both zebrafish and mammals, and researchers suspect that they play a key role in maintaining the blood–brain barrier and clearing toxic substances from the brain. Investigators from the Intramural Research Program of the NIH Eunice Kennedy Shriver National Institute of Child Health and Human Development are using the zebrafish to further our understanding of the function of FGPs. They recently discovered that FGPs are closely related to cells that form the lymphatic system, which collects, cleans, and returns fluid to the circulatory system.
By MENU Related Pages Breakthroughs and Horizons in Bioscience Scientific Contests BioArt About BioArt Submit Entry Dates, Terms & Conditions Frequently Asked Questions Current Winners Past Winners Stand Up for Science Washington Update Newsletter Receive FASEB Email Communications News Room The 2017 BioArt Winners Scroll below for the winners of the 2017 BioArt Contest. For better viewing, click the images below to enlarge them. Marina Venero Galanternik1*, Daniel Castranova1, Tuyet Nguyen2, and Brant M. Weinstein1* 1Eunice Kennedy Shriver National Institute of Child Health and Human Development, National Institutes of Health (Bethesda, MD) 2University of Maryland (College Park, MD) *Society for Developmental Biology Research Focus: Cardiovascular development This microscopy image shows that Fluorescent Granular Perithelial cells (FGPs, in green) are closely associated with the blood vessels (red) that surround an adult zebrafish’s brain. FGPs are novel type of cell found in both zebrafish and mammals, and researchers suspect that they play a key role in maintaining the blood–brain barrier and clearing toxic substances from the brain. Investigators from the Intramural Research Program of the NIH Eunice Kennedy Shriver National Institute of Child Health and Human Development are using the zebrafish to further our understanding of the function of FGPs. They recently discovered that FGPs are closely related to cells that form the lymphatic system, which collects, cleans, and returns fluid to the circulatory system. Dimitra Pouli1, Sevasti Karaliota2, Katia P. Karalis2, and Irene Georgakoudi1 1Tufts University (Medford, MA) 2Biomedical Research Foundation, Academy of Athens (Athens, Greece) Research Focus: Fat metabolism These white fat cells were imaged using a specialized technology called Coherent anti-Stokes Raman scattering (CARS). This label-free, noninvasive process uses near-infrared light to probe the vibrations of specific types of atomic bonds. The output of CARS lets scientists “see” where high concentrations of fat (lipids) are present in intact living tissue. This research team is studying the metabolic behavior of tissues with lots of white fat cells (energy-storing) versus those with many brown fat cells (energy-dissipative). Through this NIH National Institute of Biomedical Imaging and Bioengineering-supported project, they aim to expand our understanding of how different fat tissues work, which might inform new interventions for obesity, diabetes, and metabolic syndrome. João Botelho, Daniel Smith, Macarena Faunes, and Bhart-Anjan Bhullar. This alligator embryo is in the early stages of organ development or organogenesis. Fluorescent labeling highlights the nerves (green), muscles (orange), and cell nuclei (blue). At this developmental stage, several crocodylian characteristics are becoming apparent, including a long tail and the massive trigeminal nerve in the head – which makes an alligator’s face more sensitive than a human fingertip. However, it still looks very similar to a bird embryo, which is no coincidence: crocodylians and birds are each other’s closest living relatives. Their common ancestor lived over 250 million years ago and would have looked like a small dinosaur. This research team is comparing alligator and chicken development to identify differences that produce bird-like characteristics. The NSF Directorate for Biological Sciences supports these researchers’ studies of the evolution and development of bird body structures.
By Kevin A. Murach, Charlotte A. Peterson, and John J. McCarthy. In this image culture-grown muscle stem cells from a mouse have fused together to form myotubes, mimicking the formation of muscle fibers in living organisms. Fluorescent labeling reveals the myotubes’ multiple nuclei (blue) and distinct striations (red) – both are characteristic of mature muscle fibers. Some of the myotubes also display green fluorescence, which was introduced into the cells with a virus. The researchers plan to use the same viral delivery system to genetically modify the cells and assess how impairing cell fusion alters myotube growth. The NIH National Institute of Arthritis and Musculoskeletal and Skin Diseases and National Institute on Aging support their research into muscle growth, adaptation, and recovery in adults, including how muscle stem cells in modify the surrounding cellular environment to promote these activities.
By Haley O’Brien. Cloven hoofed mammals have a special arterial network inside their skulls that is used to keep their brains cooler than their bodies. Selective brain cooling helps these animals reduce water loss and avoid heat stroke. In this CT scan of an American pronghorn antelope (Antilocapra americana), a special contrast dye was used to illuminate the skull and arteries. Dr. O’Brien is investigating whether the ability to keep the brain cool has helped these animals survive warming and drying climates. The NSF Directorate for Social, Behavioral, and Economic Sciences recently funded the purchase of a x-ray micro-computed tomography (microCT) scanner at the University of Arkansas that will be used by Dr. O’Brien and other researchers in Arkansas, Oklahoma, Missouri, and Kansas to promote scientific discovery and foster academic-industry partnerships.
By Vanja Stankic and Rachel K. Miller. Cilia (yellow) are specialized hair-like structures on cells. Some cilia are motile and can beat in coordination, creating a directional fluid flow. This motion is used to propel an egg toward the uterus, circulate cerebrospinal fluid in the brain, and clear airway tracts in the respiratory system. Immotile cilia within the kidney are thought act as sensors of fluid flow. Structural and functional defects in cilia are linked to infertility, brain abnormalities, chronic respiratory problems, and kidney abnormalities. This image show skin cells from a frog (Xenopus laevis) embryo, which also have motile cilia and are commonly used as a research model for cilia development, or ciliogenesis. These NIH National Institute of Diabetes and Digestive and Kidney Diseases-funded researchers are using this frog model to study the role of ciliogenesis in kidney development.
Olga Zueva,Thomas Heinzeller, Daria Mashanova, and Vladimir Mashanov. Brittle stars and starfish have radial symmetry, with a nerve cord running down the length of each arm. This 3D model shows the nervous system within one arm segment of a brittle star (Amphipholis kochii). The colors indicate the three subdivisions of its nervous system: the ectoneural system (green); the hyponeural system (magenta); and mixed peripheral nerves (blue). This pattern of nerves is repeated in all segments throughout an arm. To create this model, the research team imaged thin sections of a brittle stars arm and used specialized software to assemble and fine-tune the model. Scientists are increasingly using brittle stars and other echinoderms to study limb regeneration, bioluminescence, and other features. Their NSF Directorate for Biological Sciences-supported work expands our fundamental knowledge of how echinoderm nervous systems are organized.
Development is delighted to announce the third in our highly successful series of events focussing on human developmental biology. Since the initiation of this series, in 2014, we have witnessed huge progress in this field, with more and more researchers turning to stem cell and organoid systems to investigate development and organogenesis in vitro, as well as increased analysis of human embryos and tissues to understand how these processes occur in vivo. Technological advances such as genome editing, single cell sequencing and improvements in tissue engineering now allow us to delve more deeply into the conserved and divergent processes underlying human development. Such knowledge is essential to underpin translational research into developmental disorders and to develop cell and tissue therapies.
This meeting brings together researchers working on a diverse set of questions, united by common challenges associated with working with human cells and tissues, and by a common goal to understand the similarities and differences between human development and that of other species. As well as talks from invited speakers and selected delegates, the meeting will also include a discussion session on the ethical and legal challenges of working with early human embryos, cells and organoid cultures – and how we as a community should address these.
We invite applications for a funded PhD position in the Department of Zoology in Central Cambridge on Downing Street with Dr Stephen Montgomery’s research group.
Project title:
Developmental basis of mushroom body expansion in Heliconius butterflies
Project summary:
Mushroom bodies (MBs) are the most enigmatic structures in the insect brain. They have ‘higher order’ functions, integrating sensory information and storing memories of past experience. MBs share a conserved ground plan, but their size and structure varies extensively across species. MB volume in Heliconius butterflies are among the highest across insects, 3-4 times larger than typical for Lepidoptera, including their most closely related genera in the wider tribe of Heliconiini. This provides a novel system for investigating the developmental mechanisms that control neural proliferation and brain component size. However, nothing is known about the developmental changes that have produced such a dramatic expansion.
What the student will be doing:
The project will involve core laboratory, microscopy and neuroanatomical techniques, insect rearing and experimental manipulation. You will construct the first time course of Heliconius brain development, from late larvae through pupation to adulthood. Using immunocytochemistry and confocal imaging you will determine key properties of MB growth trajectories, providing a template for developing hypotheses of when and how MB development in Heliconius diverged from related genera.
By adopting an evo-devo approach you will then conduct a series of comparative studies across Heliconiini to assess how increases in neuron number are produced, considering four potential mechanisms: i) an increase in the number of neural progenitor cells, ii) accelerated cell-cycle rates during neurogenesis, iii) an extension in the overall duration of neurogenesis including the possibility of adult neurogenesis, iv) reduced or delayed patterns of apoptosis among neural progenitor cells.
Finally, once key periods of developmental divergence have been identified between Heliconius and related genera, you will perform a second series of analyses to identify divergent patterns of gene regulation and expression as part of a project that aims to identify the genetic basis of MB expansion.
Some field/insectary work in Panama may be necessary/desirable. The studentship is funded by the European Research Council for 3.5 years. A student stipend of £14,553 per annum, and tuition fees will be offered to a successful candidate.
FUNDING: The studentship is funded by the European Research Council for 3.5 years. A student stipend of £14,553 per annum, and tuition fees will be offered to a successful candidate.
Students on average author 1-3 papers and produce at least three times that many conference posters***. At large meetings, such as the ASCB, thousands of posters are presented each year. While presenting posters is popular, posters sessions evoke mixed feelings: they are often late in the evening, interrupted by special workshops, held in badly lit rooms far away from the bar, and many posters are sub-par: they are crammed with details and text in small font, and presenters elaborate in great detail. Experienced conference attendees therefore excel in the brief scanning of the title while avoiding eye contact with the presenter for fear of being entangled in a never-ending run-down of experimental details.
While we can’t influence the conference organization, we can absolutely and with little effort improve the posters! Based on my survey data, I compiled the top ten tips to improve your poster:
Legible title
Make the title and your name readable from afar. This means, not too many words per title, maybe 6 to 10, in a legible font – Helvetica Neue, Verdana, Calibri or similar. Refrain also from All caps as it becomes hard to read after a few words. – If you love all caps, why not try Small Caps with capitalization instead.
Avoid abbreviations
Ideally no abbreviations in the title and as few as possible in the poster content. Only few abbreviations are so common that they became words themselves: DNA, RNA, some gene and protein names. You don’t want to turn audience away with jargon, and remember, even specialist’s conferences are attended by editors, journalists, and newcomers in the field – be welcoming to them all!
Not too much text
We read maximally 100-200 words per minute – but in posters, with scientific data, terms, and charts our reading speed will be significantly decreased. Keep that in mind – I personally am more convinced by a figure than by you explaining and interpreting it.
Clear section layout
Start at the top left and end at the bottom right. This is how we read text, and also posters! Alternative: arrange your content in 2-3 columns, similar to an article – make sure the columns are clear by leaving enough white space surrounding them! Please refrain from unconventional layouts – the chances are high that it will confuse your readers!
Figure titles instead of legends
This is easy – try moving the figure legend above the image/chart, instead of showing it below as you would in a paper. Right away, this gives you a header for that section! Explanations of the color code, which are critical to understand a figure, can be sub-headers!
Consistent color code
Absolutely keep the color code consistent across all figures! Nothing kills more time than figuring out the color code of each individual chart! Please, if your main experiment/mutant/condition is shown in “red” in the first figure, do not deviate from this in the next figure! And, of course, be color-blind friendly (no mixing red and green!)
Simple pictures and charts.
There is likely fascinating detail in your data, but not everyone wants to know all of it during a poster session. Therefore, please consider removing unnecessary details from your graphs! (Also: avoid 3D, no bar charts for distributions (#BarBarCharts) and avoid unconventional graph-types: it’s already unlikely people understand them in a paper, and less likely they feel like deciphering them in a poster session.
Poster-Etiquette: Have the elevator speech ready!
Give your audience a polite overview in 2-3 minutes that includes the big picture and key finding, but leave out experimental details. If they are interested in more, they will ask! (Also, it is convenient to have this 2-minute blurb ready in case you accidentally bump into the heroine/hero of your field in the coffee line, instead of at the poster session!)
Rehearse whenever you can!
Find 10 volunteers, not necessarily your supervisor only, to test out your 2-minute presentation; while in the lunch line, when waiting for a measurement to finish, or when cleaning the bench.
Tricks are allowed.
To get people interested in your poster, you can use tricks. Have handouts ready to take home, bring a laptop to show movies, I’ve seen people hand out sweets, and know someone that served beers – everything is allowed when trying to convince people to read your poster!
Further reading:
A really nice paper on how to give a poster presentation is here: “Producing punchy posters” by Bernard S. Brown, in Trends in Cell biology, Vol. 6, 1996. He mainly deals with text, and less with figures, but has been helpful for me for 20 years!
Three months ago I did the unthinkable and moved to England, a strange haggis-free land. Starting my BBSRC PhD Internship (introductory post here) at the Node having left my Scottish lab was an amazing opportunity to get more involved with the dev bio community.
My main task while here was the day to day running of the Node; twitter, jobs and asking authors of interesting papers to write for us, thank you to all the researchers that did! You can read one of my favourite posts on using scRNA-seq to understand cell fate conversion here. I also conducted our people behind the papers interviews and a particular favourite was on the fascinating puzzle of plant cell topology, I even attempted to read On Growth and Form after their interview!
The chance to read and explore such a wide variety of topics has been eye-opening and there is so much fascinating research being conducted on topics I didn’t know existed. Learning about so many different model systems has shaped my thinking about my future in research (any Cephalopod neurobiology labs needing a Postdoc soon?).
Alongside keeping the Node running, my second project focused on refurbishing the resources section on the Node. My goal here was to make use of my position as an active researcher to make the resources page a community area to find useful links for research, teaching and to encourage researchers to engage in outreach ad advocacy. This resources list is by no means comprehensive and we still need your input. Please get in touch to let us know what is useful to you, what needs to be added/updated and any way that the resources section can be improved!
This internship has been a wonderful insight into the developmental biology community and I am so grateful to Aidan and Katherine for the opportunity and their guidance throughout the three months. I encourage all researchers to write for the Node, get your research out there! I am now returning to my PhD, all that is left to say is so long and thanks for all the Zebrafish.
The story behind this study provides yet another example of where the pursuit of a few chance observations developed into an interesting project in its own right.
I started my postdoc in the lab of Alex Gould at The Francis Crick Institute with the objective of investigating how nutrient deprivation affects the growth of different body parts of the fruit fly larva. These experiments required precise measurements of organ sizes, and it soon became pretty clear that in order to get the most useful data, males and females had to be analysed separately. Not because the different sexes necessarily show different growth responses to starvation, but because female body parts are consistently 20-40% larger than their male counterparts. Hence, if males and females are pooled, the data have a large variance. Surprisingly, a lot of studies that have analysed larval growth seem to have ignored this sex difference in body size, officially termed sexual size dimorphism (SSD). So SSD effectively doubled my workload but it also opened up a new question: how does the sex of the larva influence its growth?
Summary of the key findings
We used two opposite angles of attack to identify the mechanism underlying larval SSD. First, in a sort of top-down approach, we asked if there were any measurable differences in growth parameters or behaviour that could give insights into underlying genetic mechanisms. Second, using a bottom-up strategy, we manipulated sex determination genes in individual cell types to see if the sex of a specific body part influenced overall body size.
For the first approach, I painstakingly weighed individual male and female larvae at different stages in order to determine their growth curves. When a larva hatches from the embryo it only weighs around 10mg, about 1/5th of the mass of a grain of salt! [1], and it grows to a final size of about ~2mg, around half the mass of a sesame seed [2], before it undergoes pupal development about 4 days later. This work established that males and females have the same size at larval hatching and that they have similar time windows of growth. Nevertheless, females begin to have a higher growth rate from about the middle of the second larval instar onwards. Interestingly, I found that the greatest difference in the fold rate of growth between the sexes occurs relatively early in larval development, during the second larval instar. This means that any pathways that control sex differences in body size should be active before or during the second larval instar.
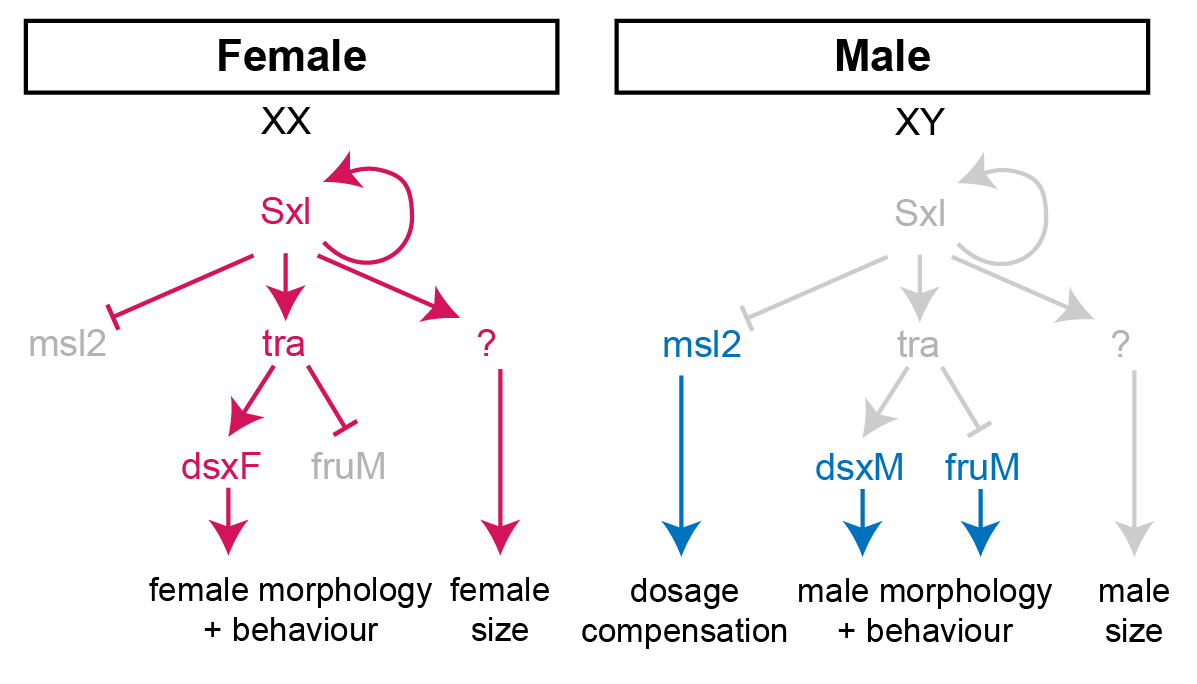
Fig 1: Diagram of the sex determination pathway in Drosophila. In females, the presence of two X-chromosome activates the expression of the splicing factor Sex-lethal (Sxl). Sxl maintains its own expression and causes sex-specific splicing of transformer (tra), such that a functional Tra protein is only produced in females. Tra is another splicing factor that causes the sex-specific splicing of the transcription factors doublesex (dsx) and fruitless (fru), which regulate sex-specific morphologies and behaviour. Sxl also represses the translation of male-specific lethal 2 (msl-2) in females, thereby limiting assembly of the X-chromosome dosage compensation complex to males.
For the second approach, we used the amazing GAL4/UAS system of Brand and Perrimon [3] to manipulate a gene called Sex-lethal (Sxl), the master regulator of fly sex determination. Sxl is normally only expressed in females, and it directs the sex-specific splicing of downstream genes in order to specify the development of female morphology and behaviour (Fig 1). Sxl mutant females are masculinised both in terms of their morphology and their body size, demonstrating that Sxl controls SSD by promoting higher female growth.
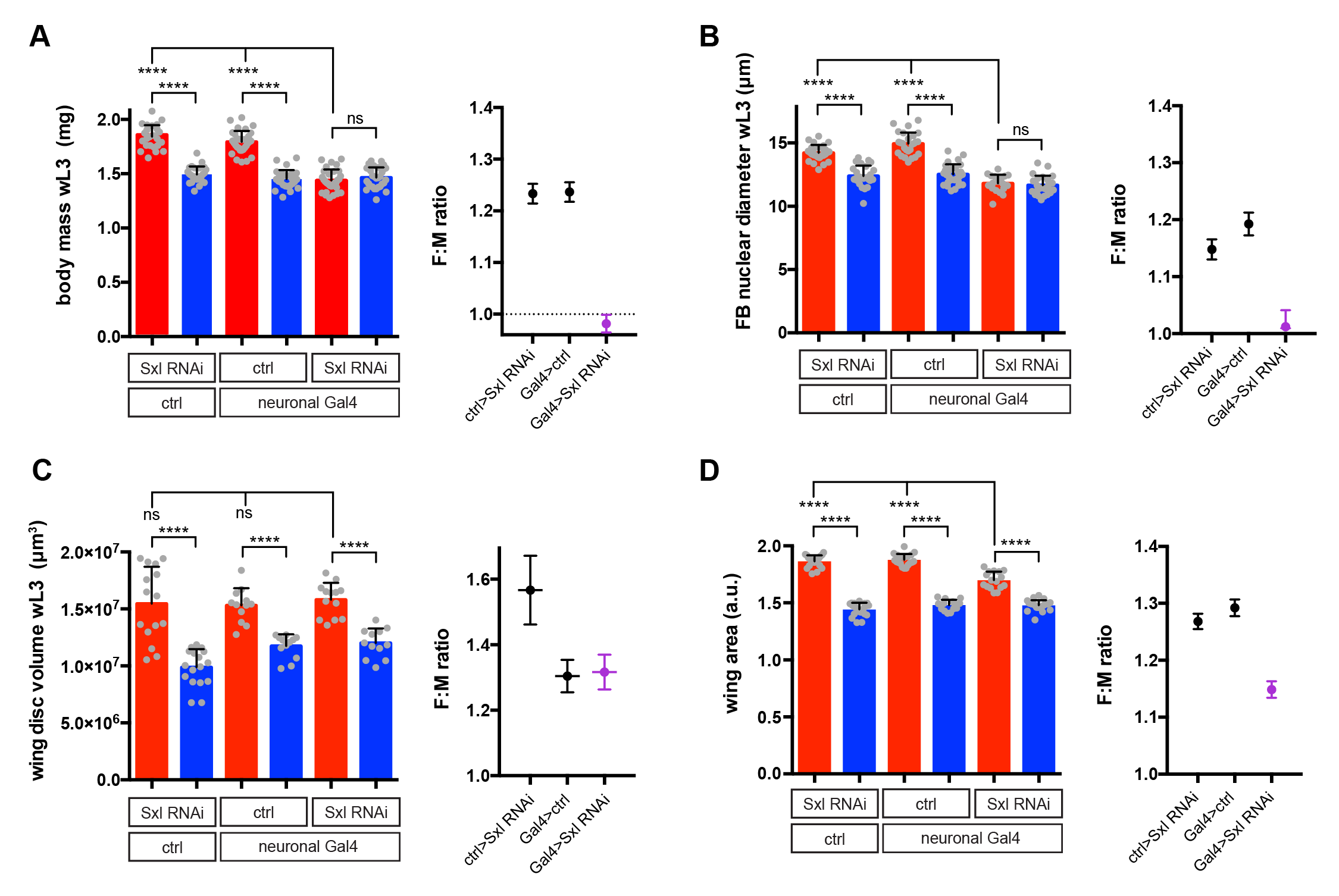
I used RNAi to inhibit Sxl expression in different tissues and, strikingly, found that knockdown with a pan-neuronal Gal4 driver reduced the body size of females to that of males (Fig 2A). In the reverse experiment, restoring Sxl expression in neurons could partially rescue the female body size of Sxl mutants. Thus, Sxl acts in neurons in a non-cell autonomous manner to boost the growth of the female body. To pinpoint the neuronal population in which Sxl functions to promote SSD, a panel of neuronal subset-specific Gal4 drivers were then screened. We found that Sxl acts additively in at least two non-overlapping subsets of neurons: 1) the insulin producing cells (IPCs), a cluster of 7 neurosecretory cells that are known to secrete several insulin-like peptides into the circulation, and 2) cells expressing the Gad1-Gal4 driver, which is active in GABAergic neurons (although we found that the overlap with GABA-expressing neurons is only partial).
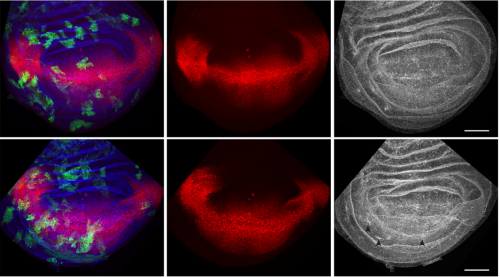
Fig 2: Effect of pan-neuronal knockdown of Sxl on sexual size dimorphism (SSD) in larval and imaginal tissues. Sxl RNAi was expressed using a pan-neuronal Gal4 driver (elav[c155]-Gal4) and the effect on SSD was measured for various body parts. Measurements were taken at the end of larval development, in wandering L3 (wL3) larvae (A-C) or in adult flies (D). The right graphs in each panel plot the female to male ratio as a quantification of SSD. Pan-neuronal Sxl RNAi abolishes SSD at the level of larval body mass (A) and the fat body nuclear diameter (B), while SSD of the wing imaginal disc remains intact (C). Pan-neuronal Sxl RNAi reduces, but does not abolish SSD of the adult wing (D).
The results of both approaches raise four tricky questions that Alex and I have been thinking about, and also investigating, with varying levels of success:
How does the new study fit with the dogma that somatic sex determination in flies is cell-autonomous?
We think the answer to this important question lies in differences between how Sxl regulates the SSD of the larval versus the adult body. On the face of it, our finding that Sxl acts in neurons to increase female body growth would seem to conflict with classic genetic studies of gynandromorphs (flies mosaic for male and female cells). These sex mosaic flies had clearly demonstrated that male/female morphologies are controlled cell-autonomously. Not only that but, in a bilateral gynandromorph, the male side is smaller than the female side [4] (Fig 3). So how can we square the classic findings with our new observations?
Fig 3: Drawing of a bilateral gynandromorph (reproduced from [4]). The drawing depicts a mosaic fly with male cells developing on the left side and female cells developing on the right side, likely as a result of a loss of one X-chromosome during early embryonic divisions. Note that the male side shows typical male morphological differentiation such as the sex-combs (dark bristled on front leg), as well as a smaller body size compared to the female side.
The first thing to note is that the presence of the cell-autonomous effects revealed by gynandromorphs does not completely rule out additional non-cell autonomous effects. In fact, such a dual SSD mechanism does seem to operate in mammals. For example, a recent mouse study suggests that gonadal hormones and direct effects of sex chromosomes combine to produce overall differences in growth and metabolism between males and females [5, 6]. Furthermore, we and others [7, 8] have found that manipulation of Sxl or its downstream target transformer does indeed have cell-autonomous effects on growth, although they are too small to account for the full extent of SSD.
Secondly, we realised that the gynandromorph studies only looked at the external structures of the adult fly. These are derived from small groups of diploid cells called imaginal discs, which grow within the larval body and only transform into adult structures during metamorphosis. In contrast, I had measured SSD for the larval body itself, the bulk of which is composed of large polyploid cells, which are degraded during metamorphosis and so do not make it to the final fly. This realization led to the testable hypothesis that SSD is regulated differently in the larval polyploid tissues versus the diploid imaginal tissues. Unlike many of our crazy hypotheses, this one turned out to stand the test of time as neuronal knockdown of Sxl abolished SSD in the fat body (a larval tissue), yet it had no effect on SSD in the wing imaginal disc (the adult wing precursor) (Fig 2B and 2C). Thus, neuronal Sxl in females specifically boosts the growth of larval not imaginal tissues during the period of juvenile development. Nevertheless, by the time the imaginal discs have transformed into the mature external structures of the adult fly, I did observe a moderate effect of neuronal Sxl knockdown on SSD (Fig 2D). To account for this, we propose that, by promoting larval tissue growth, neuronal Sxl also indirectly boosts the growth of imaginal tissues, because a larger larval body confers increased pupal resources (nutrients and/or growth factors) to sustain the growth of imaginal structures during pupal stages. An alternative explanation, which we cannot rule out for now, is that neuronal Sxl also acts during pupal stages to directly influence the size of imaginal structures.
Is there a role for insulin signalling in SSD?
We think the overall answer to this question is “yes but only for imaginal tissues” and here’s our logic for why this may be the case. Sxl functions in insulin producing cells and it is known that insulin secretion from these cells is a key regulator for growth, so an attractive hypothesis is that Sxl promotes insulin secretion in females, thereby boosting female growth. Interestingly, SSD in mammals is driven in part by gonadal hormones causing sex differences in insulin-like growth factor 1 (IGF-1) release. However, our data suggest that SSD of the larval body (i.e. the target of neuronal Sxl) is established independently of sex differences in insulin signalling. This is because, at the early stages when the sex difference in growth rates is maximal (the second instar larva), no sex differences in insulin secretion or insulin signalling could be detected. In addition, several manipulations aimed at changing insulin secretion from IPCs also had no effect on larval body SSD. And finally, mutant larvae that lack one or all of the insulin-like peptides (Ilps) produced in the insulin producing cells are very small yet their SSD remains intact. These findings suggest to us that Ilps are not required for the establishment of sex differences in growth in the early larva.
But the twist here is that there is evidence suggesting a role for insulin signalling in SSD in older larvae and in adult flies. Thus, insulin receptor mutant adult flies show a strong reduction in SSD (measured as dry body mass) [9]. At late larval stages, it has also been reported that insulin secretion from IPCs and insulin signalling is higher in females [7]. It is therefore likely that sex differences in insulin signalling in old larvae contribute to SSD in imaginal tissues, even though at this time larval body growth has largely finished. In support of this, I found that when IPCs are genetically manipulated to decrease their size and so presumably their insulin secretion, SSD is abolished in the wing imaginal disc but it remains intact in the larval body, despite the smaller absolute body masses of both sexes. Clearly, more work is needed to understand why insulin signalling appears to regulate the SSD of larval versus imaginal tissues so differently.
What is downstream of neuronal Sxl to boost female body growth?
We wish we could answer this tricky question but, unfortunately, all we can give you right now is a list of factors that are probably NOT involved.
As mentioned above, the current evidence indicates that insulin like peptides are not strong candidates for the Sxl targets in IPCs that are relevant for establishing larval body SSD. Nevertheless, it is possible that Sxl regulates the secretion of other signalling molecule(s) into the circulation, either directly from IPCs, or from cells that the IPC neurons connect with. Notably, IPCs send axons to the ring gland [10, 11], a major hemosecretory organ, which could provide such a systemic signal. The role of Sxl in the Gad1-Gal4 neurons is even less clear.
Two direct downstream targets of Sxl that have been widely studied are transformer and msl-2 (see Fig 1). Our loss of function and rescue experiments strongly suggest that neither transformer nor msl-2 play a role in the neurons regulating larval SSD. The msl-2 data also ruled out the possibility that the reduction in female body sizes we saw in Sxl loss of function experiments were due to “sickness” caused by ectopic activation of msl-2 expression and the dosage compensation machinery. It is likely that a new Sxl target is involved, but identification of this may well require unbiased sequencing experiments or other genome-wide techniques, using brains from the relevant developmental stage.
Is increased female body growth driven by increased feeding?
We have been asked this question a lot at conferences. We did measure feeding rates in early second and early third instar larvae. In both cases, sex differences in the absolute food intake rate (i.e. intake per larva) were detected, but when these were normalised for body mass, mass-specific food intake rates (i.e intake per mg of larval tissue) appear to be the same for males and females. This suggests to us that sex differences in absolute food intake are a consequence of a larger female body mass, but not the driver of it. So why then do female larvae have a higher mass-specific (i.e. fold) growth rate than males despite both sexes having similar mass-specific feeding rates? One possible explanation here is that female larvae are more efficient at converting nutrients into tissue mass than males.
It is important to highlight that there are caveats to our conclusion that mass-specific food intake is equal in male versus female larvae. This is because we measured food intake over 20-30 minutes using a blue dye uptake method that would not be sensitive enough to detect very small differences. Some clever method for measuring food consumption over much longer periods might be necessary to detect very small sex differences in mass-specific food intake. Unfortunately, this may be tricky as larvae not only eat their food but they also burrow in it, and defecate into it. However, there is another reason for thinking that the neuronal Sxl mechanism for establishing SSD does not involve sex-specific feeding. This is because increased feeding rates in females would be predicted to increase the growth of both larval and imaginal tissues, yet we observed that imaginal tissues inside the larva are blind to neuronal Sxl regulation.
Concluding remarks
Sex determination and growth regulation have been extensively studied in Drosophila for many years. It is therefore very surprising that only a handful of studies have investigated how sex influences growth in this model organism. The neuronal Sxl relay mechanism that we have recently discovered contrasts with the text book view that somatic sexual differentiation in insects is controlled in a strictly cell-autonomous fashion. Future research into the neuronal Sxl mechanism for larval SSD will be needed to identify the relevant Sxl targets and perhaps endocrine growth regulatory mechanisms that act selectively on larval polyploid tissues rather than diploid imaginal discs. Despite this, our research has already revealed glimpses that suggest that the regulatory logic of SSD in mammals and insects may be more similar than previously thought. This encouraging news opens up the possibility of using our favourite model organism to study how sex differences in growth and metabolism impact upon disease – an important emerging area of biomedical research.
References
“Grain of salt”: Bluebulb Projects’ The Measure of Things; [02.12.2017]. Available here.
Our latest monthly trawl for developmental biology (and other cool) preprints. Let us know if we missed anything.
This month in preprint news – here at The Company of Biologists, set just outside Cambridge in the UK, we’re hiring a new ‘Preprints Community Manager’. You can find out more here.It’s a great opportunity to get in to science communication and publishing, and of course preprints!
And this month in preprints, we found platypuses and pigeons, xenotransplants and reoriented axes, CRISPR tools and liquid droplets. The preprints were hosted on bioRxiv, PeerJ, andarXiv. Use these links to get to the section you want:
Human-specific NOTCH-like genes in a region linked to neurodevelopmental disorders affect cortical neurogenesis. Ian T Fiddes, Gerrald A Lodewijk, Meghan M Mooring, Colleen M Bosworth, Adam D Ewing, Gary L Mantalas, Adam M Novak, Anouk van den Bout, Alex Bishara, Jimi L Rosenkrantz, Ryan Lorig-Roach, Andrew R Field, Maximillian Haeussler, Lotte Russo, Aparna Bhaduri, Tomasz J Nowakowski, Alex A Pollen, Max L Dougherth, Xander Nuttle, Marie-Claude Addor, Simon Zwolinski, Sol Katzman, Arnold Kreigstein, Evan E Eichler, Sofie R Salama, Frank MJ Jacobs, David Haussler
Loss of Wt1 in the murine spinal cord alters interneuron composition and locomotion. Danny Schnerwitzki, Sharn Perry, Anna Ivanova, Fabio Viegas Caixeta, Paul Cramer, Sven Guenther, Kathrin Weber, Atieh Tafreshiha, Lore Becker, Ingrid L. Vargas Panesso, Thomas Klopstock, Martin Hrabe de Angelis, Manuela Schmidt, Klas Kullander, Christoph Englert
Higher-order inter-chromosomal hubs shape 3-dimensional genome organization in the nucleus. Sofia A Quinodoz, Noah Ollikainen, Barbara Tabak, Ali Palla, Jan Marten Schmidt, Elizabeth Detmar, Mason Lai, Alexander Shishkin, Prashant Bhat, Vickie Trinh, Erik Aznauryan, Pamela Russell, Christine Cheng, Marko Jovanovic, Amy Chow, Patrick McDonel, Manuel Garber, Mitchell Guttman
The Transcriptional Logic of Mammalian Neuronal Diversity. Ken Sugino, Erin Clark, Anton Schulmann, Yasuyuki Shima, Lihua Wang, David L. Hunt, Bryan M. Hooks, Dimitri Trankner, Jayaram Chandrashekar, Serge Picard, Andrew Lemire, Nelson Spruston, Adam Hantman, Sacha B. Nelson
| Stem cells, regeneration & disease modelling
Neuroepithelial rosettes from Medelnik, et al.’s preprint
T helper cells modulate intestinal stem cell renewal and differentiation. Moshe Biton, Adam Haber, Semir Beyaz, Noga Rogel, Christopher Smillie, Karthik Shekhar, Alexandra Schnell, Zuojia Chen, Chuan Wu, Jose Ordovas-Montanes, David Alvarez, Rebecca H. Herbst, Itay Tirosh, Grace Burgin, Danielle Dionne, Michael E. Xifaras, Mei Zhang, Alex K. Shalek, Ulrich H. von Andrian, Daniel B. Graham, Orit Rozenblatt-Rosen, Hai Ning Shi, Vijay Kuchroo, Omer Yilmaz, Aviv Regev, Ramnik J. Xavier
Developmental And Genetic Regulation Of The Human Cortex Transcriptome In Schizophrenia. Andrew E Jaffe, Richard E Straub, Joo Heon Shin, Ran Tao, Yuan Gao, Leonardo Collado Torres, Tony Kam-Thong, Hualin S Xi, Jie Quan, Qiang Chen, Carlo Colantuoni, William S Ulrich, Brady J Maher, Amy Deep-Soboslay, The BrainSeq Consortium, Alan Cross, Nicholas J Brandon, Jeffrey T Leek, Thomas M Hyde, Joel E Kleinman, Daniel R Weinberger
Gene expression imputation across multiple brain regions reveals schizophrenia risk throughout development. Laura M Huckins, Amanda Dobbyn, Douglas Ruderfer, Gabriel Hoffman, Weiqing Wang, Antonio F Pardinas, Veera M Rajagopal, Thomas D Als, Hoang Tan Hoang, Kiran Girdhar, James Boocock, Panagiotis Roussos, Menachem Fromer, Robin Kramer, Enrico Domenici, Eric Gamazon, Shaun Purcell, CommonMind Consortium, Schizophrenia Working Group of the Psychiatric Gen, iPSYCH-GEMS Schizophrenia Working Group, Ditte Demontis, Anders Borglum, James Walters, Michael O’Donovan, Patrick F Sullivan, Micahel Owen, Bernie Devlin, Solveig K Sieberts, Nancy Cox, Hae Kyung Im, Pamela Sklar, Eli Ayumi Stahl
Aging and neurodegeneration are associated with increased mutations in single human neurons. Michael A Lodato, Rachel E Rodin, Craig L Bohrson, Michael E Coulter, Alison R Barton, Minseok Kwon, Maxwell A Sherman, Carl M Vitzhum, Lovelace J Luquette, Chandri Yandava, Pengwei Yang, Thomas W Chittenden, Nicole E Hatem, Steven C Ryu, Mollie B Woodworth, Peter J Park, Christopher A Walsh
Germline loss of MBD4 predisposes to leukaemia due to a mutagenic cascade driven by 5mC. Mathijs A. Sanders, Edward Chew, Christoffer Flensburg, Annelieke Zeilemaker, Sarah E. Miller, Adil al Hinai, Ashish Bajel, Bram Luiken, Melissa Rijken, Tamara Mclennan, Remco M. Hoogenboezem, François G. Kavelaars, Marnie E. Blewitt, Eric M. Bindels, Warren S. Alexander, Bob Löwenberg, Andrew W. Roberts, Peter J. M. Valk, Ian Majewski
Hemimetabolous genomes reveal molecular basis of termite eusociality. Mark C. Harrison, Evelien Jongepier, Hugh M. Robertson, Nicolas Arning, Tristan Bitard-Feildel, Hsu Chao, Christopher P. Childers, Huyen Dinh, Harshavardhan Doddapaneni, Shannon Dugan, Johannes Gowin, Carolin Greiner, Yi Han, Haofu Hu, Daniel S. T. Hughes, Ann-Kathrin Huylmans, Carsten Kemena, Lukas P. M. Kremer, Sandra L. Lee, Alberto Lopez-Ezquerra, Ludovic Mallet, Jose M. Monroy-Kuhn, Annabell Moser, Shwetha C. Murali, Donna M. Muzny, Saria Otani, Maria-Dolors Piulachs, Monica Poelchau, Jiaxin Qu, Florentine Schaub, Ayako Wada-Katsumata, Kim C. Worley, Qiaolin Xie, Guillem Ylla, Michael Poulsen, Richard A. Gibbs, Coby Schal, Stephen Richards, Xavier Belles, Judith Korb, Erich Bornberg-Bauer
Platypus sampling sites from Martin, et al.’s preprint
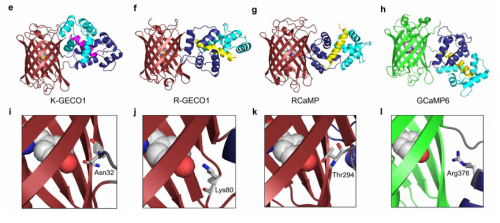
Genetically encoded calcium indicators from Shen, et al.’s preprint
A genetically encoded Ca2+ indicator based on circularly permutated sea anemone red fluorescent protein. Yi Shen, Hod Dana, Ahmed S. Abdelfattah, Ronak Patel, Jamien Shea, Rosana S. Molina, Bijal Rawal, Vladimir Rancic, Yu-Fen Chang, Lanshi Wu, Yingche Chen, Yong Qian, Matthew D. Wiens, Nathan Hambleton, Klaus Ballanyi, Thomas E. Hughes, Mikhail Drobizhev, Douglas S. Kim, Minoru Koyama, Eric R. Schreiter, Robert E. Campbell
A suite of transgenic driver and reporter mouse lines with enhanced brain cell type targeting and functionality. Tanya L Daigle, Linda Madisen, Travis A Hage, Matthew T Valley, Ulf Knoblich, Rylan S Larsen, Marc M Takeno, Lawrence Huang, Hong Gu, Rachael Larsen, Maya Mills, Alice Bosma-Moody, La’Akea Siverts, Miranda Walker, Lucas T Graybuck, Zizhen Yao, Olivia Fong, Emma Garren, Garreck Lenz, Mariya Chavarha, Julie Pendergraft, James Harrington, Karla E Hirokawa, Julie A Harris, Medea McGraw, Douglas R Ollerenshaw, Kimberly Smith, Baker A Baker, Jonathan T Ting, Susan M Sunkin, Jerome Lecoq, Michael Z Lin, Edward S Boyden, Gabe J Murphy, Nuno da Costa, Jack Waters, Lu Li, Bosiljka Tasic, Hongkui Zeng
Cross-Site Comparison of Ribosomal Depletion Kits for Illumina RNAseq Library Construction. Zachary T Herbert, Jamie P Kershner, Vincent L Butty, Jyothi Thimmapuram, Sulbha Choudhari, Yuriy O Alekseyev, Jun Fan, Jessica W Podnar, Edward Wilcox, Jenny Gipson, Allison Gillaspy, Kristen Jepsen, Sandra Splinter BonDurant, Krystalynne Morris, Maura Berkeley, Ashley LeClerc, Stephen D. Simpson, Gary Sommerville, Leslie Grimmett, Marie Adams, Stuart S. Levine
An interlaboratory study of complex variant detection. Stephen E Lincoln, Justin M Zook, Shimul Chowdhury, Shazia Mahamdallie, Andrew Fellowes, Eric W Klee, Rebecca Truty, Catherine Huang, Farol L Tomson, Megan H Cleveland, Peter M Vallone, Yan Ding, Sheila Seal, Wasanthi DeSilva, Russell K Garlick, Marc Salit, Nazneen Rahman, Stephen F Kingsmore, Swaroop Aradhya, Robert L Nussbaum, Matthew J Ferber, Brian H Shirts
Community-driven data analysis training for biology. Bérénice Batut, Saskia Hiltemann, Andrea Bagnacani, Dannon Baker, Vivek Bhardwaj, Clemens Blank, Anthony Bretaudeau, Loraine Guéguen, Martin Čech, John Chilton, Dave Clements, Olivia Doppelt-Azeroual, Anika Erxleben, Mallory Freeberg, Simon Gladman, Youri Hoogstrate, Hans-Rudolf Hotz, Torsten Houwaart, Pratik Jagtap, Delphine Lariviere, Gildas Le Corguillé, Thomas Manke, Fabien Mareuil, Fidel Ramírez, Devon Ryan, Florian Sigloch, Nicola Soranzo, Joachim Wolff, Pavankumar Videm, Markus Wolfien, Aisanjiang Wubuli, Dilmurat Yusuf, Rolf Backofen, Anton Nekrutenko, Björn Grüning
 (No Ratings Yet)
(No Ratings Yet)
 (1 votes)
(1 votes)