Applied research team in Cape Town, South Africa, is seeking quick assistance from an adventurous soul who is a specialist in the expansion and differentiation of hESCs into neural progenitors and, ideally, also islet progenitors and hematopoeitic cells. The project is part of a clinical trial and will start in early October. We will provide a stem cell lab (we don’t have a bioreactor) and will cover travel and lodging cost and provide a stipend.
We are looking for a highly skilled and motivated candidate to join our group for a PostDoc position. In the Payer lab (http://www.crg.eu/bernhard_payer), we study epigenetic reprogramming in the mammalian germ line and the effects of ageing on fertility. In this project, which will be performed in collaboration with a fertility clinic, the prospective candidate will study molecular links between ageing and fertility decline in women.
We are seeking a candidate with a strong background in Mammalian Cell Culture, Stem Cell Reprogramming and Differentiation, Epigenetics, Reproduction and Molecular Biology. Excellent candidates from other related fields will also be considered.
Work Environment
Our lab is part of the Gene Regulation, Stem Cells & Cancer Programme at the Centre for Genomic Regulation (CRG) in Barcelona, Spain (www.crg.eu). The CRG is a vibrant International Research Institute with Research Groups working in diverse fields such as Genomics, Cell and Developmental Biology, Systems Biology, Stem Cells, Cancer and Epigenetics. English is the working language.
Eligibility
Candidates can be of any nationality, but must undertake trans-national mobility and must not have resided or carried out their main activity in Spain for more than 12 months in the 3 years prior to the call deadline. Furthermore, applicants working at CRG for more than 3 months before the deadline will not be considered.
Candidates must have a PhD degree from a recognized university, plan to obtain a PhD degree by the time of employment, or have at least four years of full-time equivalent research experience. Candidates who already hold a PhD degree at the time of application are eligible to apply only if they passed their PhD exam (or equivalent) in the four years prior to the call deadline. Exceptions up to 3 years for maternity/paternity leaves and other documented career breaks will be considered.
Candidates must have at least one publication as first author (either in press or published) at the time of the deadline
Candidates must provide two letters of reference
Fellowship
36 months by the INTREPiD Fellowship programme.
Applications are accepted exclusively online through:
We now seek to appoint a Research Technician in Molecular and Cell Biology to complement our existing expertise and fill a vacant position for the final 4 years of the “Cellular thyroid hormone availability: regulation of development and tissue repair, and pathogenesis of degenerative disease” project.
The Molecular Endocrinology Laboratory employs state-of-the-art high-throughput imaging and functional phenotyping, together with next generation sequencing and bioinformatics, in a whole organism and systems biology approach. You will receive comprehensive training in order to provide up-to-date technical expertise in cell culture, investigation of molecular mechanisms and signalling pathways, and skeletal phenotyping. You will also contribute to general administration and management of the laboratory.
Closing Date: Tuesday 3 October 2017 (Midnight BST)
Development often involves the asymmetric partitioning of cellular components to daughters, and this process is crucial for successful gametogenesis. Today’s paper, published in the current issue of Development, explores the cytoskeletal mechanisms of spermatogenesis in different nematode species. We met the multi-lab team behind the work, starting with Diane Shakes (The College of William and Mary in Williamsburg, VA), and then her collaborators André Pires-daSilva(University of Warwick, UK), Gunar Fabig and Thomas Müller-Reichert (Technische Universität Dresden, Germany), and Jessica Feldman (Stanford University, CA).
Diane, can you give us your scientific biography and the main questions your lab is interested in?
Diane Shakes
DS I got my start in research as a high school senior through a special program at NASA-Ames Research Center.There I was paired with a fantastic mentor, Patricia Buckendahl, who has talent for productively incorporating novice young scientists into her research quest which at the time was to understand the fundamentals of bone metabolism and why astronauts were losing bone mass in zero gravity. As an undergraduate at Pomona College, NASA-Ames continued to be my summer research home as I explored the breadth of biology in my coursework. Ultimately, I was captured by the wonders of cell biology and the big questions of developmental biology. It was also during this time that I developed an appreciation of the special insights that can be obtained by studying unusual organisms and cell types.
As a Ph.D. student at Johns Hopkins, I joined the research group of Sam Ward. This choice not only linked me not only to the early community of C. elegans researchers but also immersed me in the exciting research that was going on at the Carnegie Institution, Department of Embryology. In Sam’s lab, I worked alongside postdoctoral fellow Steve L’Hernault to isolate and phenotypically characterize a large collection of spermatogenesis-defective mutants in C. elegans. Through these studies, I developed an appreciation for genetics, a love for microscopy, and a life-long interest in the mechanisms of cell polarity. As I neared the end of my graduate studies, Ken Kemphues had just published his foundational study on the C. elegans PAR mutants, so I was excited to join his lab for my post-doctoral studies. My project was to analyze par-5, which like the other par proteins is required to establish proper asymmetries in the 1-cell C. elegans embryo, and ultimately was found to encode 14-3-3.
When I subsequently established my own lab, first at the University of Houston and subsequently at the College of William and Mary, I decided to use my combined knowledge of C. elegans sperm and oocytes to investigate a pair of C. elegans mutants that had been reported to exhibit both maternal and paternal effect defects.
Penny Sadler
Within my own lab, Penny Sadler discovered that although affected oocytes and sperm were both arresting in metaphase of meiosis I, the sperm continued to develop post-meiotically into anucleate sperm that could nevertheless crawl and fertilize oocytes.And in a fruitful collaboration with Andy Golden and a generous supply of mutants from the Bowerman and Seydoux labs, we showed that these and other mutants with the same phenotype were temperature-sensitive alleles of the anaphase-promoting complex. These studies that came out this work stimulated my interest in the interplay between the various cellular and developmental sub-programs of gamete development and drew me back into the analysis of C. elegans spermatogenesis, particularly in the stages leading up to meiotic divisions. The next set of studies – an analysis of the spermatogenesis-specific events during meiotic prophase – were carried out in collaboration with Diana Chu whose expertise in chromatin complemented my own in the cell cycle and cytoskeleton.
In addition to on-going studies in C. elegans, my group has also started using what we know about gametogenesis in C. elegans as a basis for comparative studies in other nematodes. A phone-call from André Pires da Silva got us specifically interested in the trioecious (male/female/hermaphrodite) species Rhabditis sp. SB347 (now called Auanema rhodensis), a lab cultivable nematode with strikingly non-Mendelian sex ratios. In many ways, these cross-species comparisons are analogous to studying a very informative mutant; but in this case, they help us distinguish highly conserved, fundamental processes from those that have been subject to variation over evolutionary time.
What was known about the cytoskeletal drivers of sperm development and asymmetrical positioning in worms before your study?
DS A conserved feature of sperm development in all organisms is that, following the meiotic divisions, sperm become streamlined by discarding unnecessary cellular components. In the early 1980s, Sam Ward’s group had shown, that in C. elegans, these unnecessary components included both actin and microtubules. This is only possible because nematode sperm motility is driven by a completely different cytoskeletal protein, the major sperm protein (MSP). Subsequent experiments with actin and microtubule inhibitors suggested that actin was more important than microtubules in this process of asymmetric partitioning. Subsequently, the L’Hernault and Titus labs showed that proper partitioning required the non-conventional myosin (myosin VI). Yet, no one had ever gone back to study the stepwise progression of events that underlies this wholesale swap of the cytoskeletal system during sperm development in C. elegans. Were there aspects of the process that could be better understood in light of new studies of asymmetric partitioning? Was the unusual partitioning event in R. sp. SB347 completely novel, or an informative variant of events that happen in all nematode sperm?
André – how did your collaboration with Diane come about, and why are different nematode species such useful models for the evolution of sex and reproduction?
André Pires-daSilva
APS Sex determination is a developmental switch prone to rapid evolution, but the causes and consequences for this pattern of evolution are poorly known. The existence of species with three sexes caught my attention because they are supposed to be extremely rare. In 2004 Marie-Anne Félix published a paper mentioning the free-living nematode strain SB347 (now named Auanema rhodensis), which produces males, females and hermaphrodites. Until that time, other free-living nematode species producing three sexes were not available in culture, or were parasitic nematodes difficult to work in the laboratory. Back in 2009, I contacted Diane Shakes to help me in characterizing the cytology of SB347 spermatogenesis, because we were trying to understand why males of this species generate so few males. This was especially puzzling, since heterogametic XO males should produce XX and XO progeny in equal proportions. However, we observed mostly XX progeny only. I contacted Diane because of her expertise in cell biology of C. elegans spermatogenesis and her recent interest in comparative work.
Gunar – I understand that an interest in C elegans sperm mutants brought you, Anna and Thomas into the collaboration?
Gunar Fabig
GF As a PhD student, I am working in the lab of Thomas’ on chromosome segregation in C. elegans male meiosis. Our lab has a strong expertise in live-cell imaging and electron microscopy. So, some years ago I started to image living C. elegans males by fluorescence microscopy to analyze the dynamics of meiotic chromosome segregation. We also started to characterize wild-type spindles of various stages at the ultrastructural level using electron tomography. During the initial phase of my project, I started to think about a comparison of wild-type and mutant data to study situations of impaired chromosome segregation. At the time, I was aware that Diane had a very interesting paper together with André, in which they characterized male meiotic spindles in Rhabditis sp. SB347 (now A. rhodensis). In this paper, they reported about a skewed sex ratio that was most likely caused by a changed meiotic “program” during male chromosome segregation. So we contacted Diane and André to ask whether they would be interested in collaborating with us on the ultrastructure of male meiotic spindles in Rhabditis sp. SB347.
Thomas Müller-Reichert & Anna Schwarz
TMR At the same time Anna started in my lab to work on her Master’s thesis and I proposed to her that an EM analysis of males of this species would be a very exciting project. She did a terrific job in preparing and analyzing SB347 males. Anna discovered the interesting patterns of organelles partitioning to the respective daughter cells.
Jessica – how did you get recruited, and how did your previous work on non-centrosomal microtubules fit into the story?
Jessica Feldman
JF One of the interests of my lab is to understand the mechanisms underlying non-centrosomal microtubule organization. C. elegans is a particularly good model in which to study this question as microtubule organizing center (MTOC) activity is completely reassigned from the centrosome in dividing cells to another site following mitotic exit. I had been exploring this switch in MTOC activity in a number of different cell types in C. elegans and started to focus on the germline, where MTOC activity is at the plasma membrane of non-dividing germ cells, and at the centrosome of mitotically dividing germ cells or meiotically dividing spermatocytes. I started to film this transition and found that microtubules and microtubule minus end proteins remarkably appeared to move from the centrosome of dividing spermatocytes to the residual body. This behavior appeared to mimic a similar movement of microtubules and microtubule regulators that I had previously seen in embryonic intestinal epithelial cells during their polarization, but due to the geometry of the movement in spermatocytes was much easier to visualize there. I presented this work at a conference that Diane was also attending. I think my live imaging data helped shed light on some of the observations Diane had been making about microtubules in fixed samples. In an incredibly gracious act, Diane contacted me to see if I would like to incorporate my data into this manuscript.
Can you give us the key results of the paper in a paragraph?
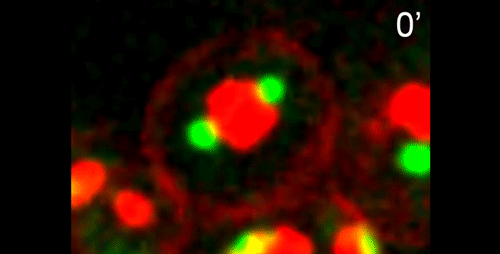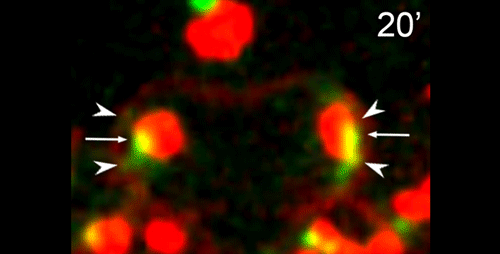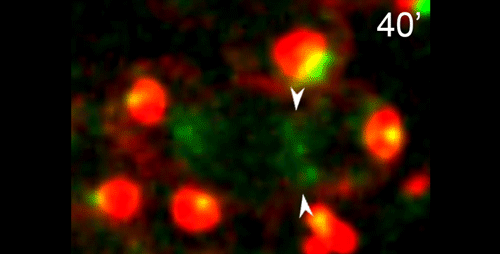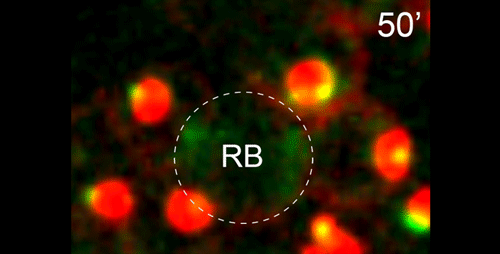
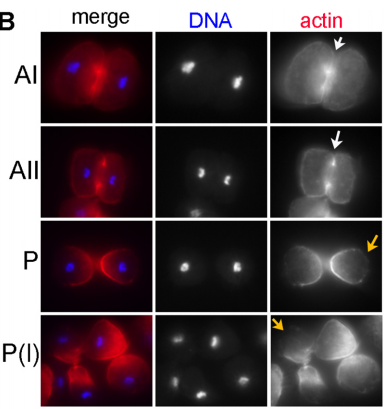
DS We knew from our earlier paper that the meiotic divisions of R. sp. SB347 male spermatocytes yielded a 50:50 mix of functional X-bearing sperm and residual bodies containing the other chromosomal complement. However we didn’t understand much about the details of the process or whether this pattern was an oddity of a single species. In fact, no one had described the stepwise process by which C. elegans sperm partition into residual bodies, not only unneeded cellular organelles but also their entire pool of actin and microtubules. In this study, we address all of these questions using the combined approaches of immunocytology, transmission electron microscopy, and live-imaging of GFP constructs. We show the first time that in C. elegans, the microtubules redistribute with their gamma-tubulin ring complexes intact from the centrosome to the sperm-residual body boundary in a process that resembles microtubule shifts in differentiating cells. At the same time, in a variation of normal cell division, actin reorganizes through a combination of cortical ring expansion and clearance from the poles. Relative to these cytoskeletal changes, organelles appear to partition in at least two phases; most partition just after the completion of anaphase chromosome segregation while others partition as the sperm detach from the residual body. In the much smaller spermatocytes of both R. sp. SB347 and its near relatives, the cytoskeletal remodeling events are restricted to the pole of the X-bearing chromosome set. Consequently, a partitioning process that is normally bipolar with two haploid sperm generating a central residual body becomes unipolar and generates one functional sperm and one DNA-containing residual body. Intriguing, this unipolar partitioning process also occurs in the XX spermatocytes of SB347 hermaphrodites. In contrast, partitioning is bipolar in the large spermatocytes of R. sp. SB347’s closest known male/female relative. Taken together, this study reveals two major insights. First, the process by which nematode sperm discard actin and tubulin into their residual bodies may be variations of common cellular and developmental processes. Second, constraints related to spermatocyte downsizing may have contributed to the evolution of a sperm cell equivalents of female polar bodies.
Fixed male gonad from C. elegans, from Figure 2 in the paper
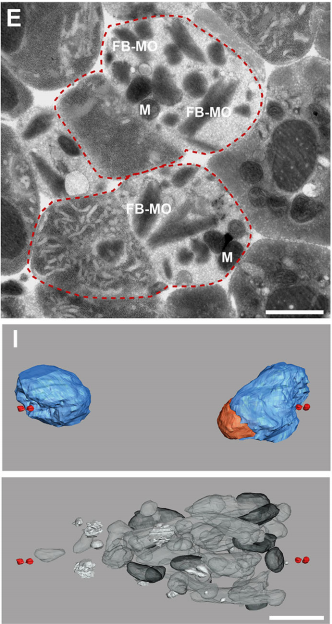
What did the TEM bring to the story?
GF & TM Spermatocytes had to be visualized in whole worms, so we had to perform serial sectioning for this project. Anna mastered this without any difficulties. The beauty about electron microscopy is that one can get very detailed images of cellular organization (e.g. about centrioles, microtubules or the Golgi apparatus). For electron tomography, we prepared whole males of Rhabditis sp. SB347 by applying high-pressure freezing. Males were ultrarapidly frozen to liquid nitrogen temperature, freeze-substituted and embedded them in Epoxy resin. We took overview images of serial sections of several worms and counted the number of different organelles in numerous cells as worms contain many spermatocytes. This information enabled us to quantify the distribution of organelles during various stages of meiotic cell divisions in males.
Thin section EM and 3D models from serial electron tomographic reconstructions, from Figure 3 in the paper
What are the key open questions about the role of microtubules in the asymmetric cell division that makes nematode spermatocytes?
JF To me the most interesting questions about the microtubules in nematode spermatocytes are, 1) how is the centrosome inactivated as an MTOC?, 2) how are microtubules transported to the RB?, 3) what organizes microtubules inside the RB?, and 4) in Rhabditis sp. SB347, what controls the selective inactivation of MTOC activity at only one of the centrosomes? The answers to these questions will not only teach us about spermatogenesis, but will also answer major questions in the field of microtubule organization.
γ-tubulin localization during the separation phase of spermatogenesis in C. elegans. Movie 1 from the paper
What do you think the main evolutionary implications of the work are?
APS Three-sexed species are interesting, because they are probably evolutionary transitions between male/female and male/hermaphrodite mating system as known in C. elegans. They may help us understand in how mating systems evolve, a long-standing question in Evolutionary Biology. In this paper, we showed the mechanisms of ‘how’ males generate non-functional nullo-X sperm. This is one of the few examples in the literature describing cellular mechanisms of how heterogametic animals generate progeny of a single karyotype. There are examples for other nematodes and animals in other phyla in which crosses between XX and XO individuals generate mostly only XX progeny. Perhaps they use similar mechanisms as SB347, and this paper provides the foundation for testing this hypothesis. At the moment, however, we do not understand ‘why’ SB347 males get rid of their ‘male-making sperm’. According to evolutionary theory, this would happen if sibling matings are common. Unfortunately, we do not know much about the ecology of SB347 to test this.
DS From my perspective as a cell and developmental biologist, I think that this work highlights how a conserved set of cellular and developmental sub-programs can be co-opted for different outcomes. This study suggests that highly unusual process of partitioning microtubules into a residual body is not a novelty of nematode spermatogenesis but rather a variation of a common process during which differentiating cells lose their centrosomal microtubules as they establish a distinct population of non-centrosomal microtubules. Similarly, we show that the step-wise progression by which nematode sperm partition actin into the residual body exhibits many similarities to actin remodeling during the transition from anaphase to cytokinesis. Thus I predict that much of the underlying molecular machinery will be evolutionarily conserved with the notable exception of a novel regulator or altered feedback loop. Yet because these processes within nematode sperm are indeed a bit extreme, their analysis will yield important insights into related cellular and developmental processes.
Actin changes in R. axei spermatocytes, from Figure 5 in the paper
What next for the Shakes lab?
DS Our on-going analysis of R. sp. SB347 continues to surprise and delight us. There is still much to be learned about the molecular mechanisms of this partitioning event as well as other oddities of SB347 meiosis. For example, we also have a paper coming out this month in Developmental Biology, in which we describe how this clade evolved a distinct solution for acquiring hermaphrodite self-fertility, namely the simultaneous rather than sequential production of oocytes and sperm. So in the R. sp. SB347 realm, we have many interesting avenues to explore. At the same time, the C. elegans focused cohort in my lab are hard at work gleaning new insights from the existing collection of spermatogenesis mutants, some of which have been languishing in our liquid nitrogen tanks since my graduate days.
Any plans for future collaborations?
Yes, Andre and Diane’s lab are collaborating on a project that describes differences in meiosis in SB347 males, females and hermaphrodites, and the labs of Andre, Diane and Thomas are collaborating on a project to identify the signal that determines the directionality of the asymmetric division of the SB347 male spermatocytes.
Anything else you would like to add?
Ethan Winter
DS One voice that is missing in this interview is the lead author Ethan Winter, a former undergraduate honors student in my lab who helped spearhead this project through his cytological studies of R. sp. SB347 and its near relatives. Ethan continued on to Harvard to pursue a Ph.D. in Chemical Biology. Not only was he brilliant, hard-working, and tenacious, but he was also kind, patient, and possessed a wonderful quirky sense of humor. We all predicted that he was on track to develop into a beloved professor. Sadly, Ethan passed away last October (2016). I hope that this paper serves at least in a small way as a tribute to his scientific accomplishments and his too-short life.
We are seeking a highly motivated and collaborative Laboratory Research Scientist in the area of human embryology and stem cell biology to join Dr. Kathy Niakan’s laboratory. The lab has identified several signalling pathways that may be operational in the human embryo to regulate the establishment or maintenance of pluripotent epiblast progenitor cells that can be coaxed to self-renew indefinitely as embryonic stem cells in vitro. The role will involve further characterising the function of these signalling pathways in the human embryo and testing their sufficiency to establish alternative human embryonic stem cells.
The successful candidate is likely to be collaborative, energetic, focused, and productive individual. Excellent organisational, analytical, and communication skills are essential.
Dr Niakan’s laboratory focuses on understanding the mechanisms of lineage specification in human embryos and the derivation of novel human stem cells. The post holder will report directly to the Group Leader, Kathy Niakan. Details of research projects currently being undertaken can be seen at: http://www.crick.ac.uk/kathy-niakan
PROJECT SUMMARY
The pluripotent epiblast of the early human embryo has the unique potential to give rise to the entire fetus in vivo and can self-renew indefinitely as embryonic stem cells (hESCs) in vitro. Understanding how this lineage is established is of fundamental biological importance and has significant clinical implications for both infertility treatment and the use of hESCs to treat various diseases. We have identified several components of key signaling pathways that are highly expressed in the epiblast, and whose activity leads to the proliferation of these pluripotent cells in vivo. Based on our preliminary data, we have been awarded an Insight to Innovate Grant to follow up these observations, in collaboration with commercial organisations.
The aim of this project is to further characterise how these factors regulate human pluripotency and embryogenesis. We also seek to establish novel culture conditions for human pluripotent stem cells by modulating these signaling pathways during stem cell derivation. In collaboration with our commercial partners, we will evaluate if these conditions better maintain pluripotency of existing hESCs, and how this may translate to improved derivation of induced pluripotent stem cells (iPSCs) or more efficient directed differentiation protocols.
Please note: all offers of employment are subject to successful security screening and continuous eligibility to work in the United Kingdom.
A postdoctoral research position is available starting in the first half of 2018 for a biologist to work with Asst. Prof. Timothy Saunders’ group at the Mechanobiology Institute, Singapore (http://labs.mbi.nus.edu.sg/mod/). The Saunders lab studies the fundamental processes shaping organs and tissues during development.
One major focus in the lab is myogenesis in the developing Zebrafish embryo. We are part of a major five-year grant focused on understanding non-canonical roles of receptor tyrosine kinases in cell regulation, including in vivo. The project will involve live-imaging myogenesis on confocal and light-sheet microscopes and developing detailed four-dimensional maps of myotome development. Reagents, including optogenetic and novel live-markers, will need to be created as part of the project.
Candidates should have extensive experience in at least two of: (1) Zebrafish genetics and general fish capabilities; (2) Biochemistry and construct design; and (3) Quantitative imaging methods and image analysis. The candidate must be prepared to learn the necessary skills to perform this challenging project.
The project is in collaboration with Prof. Philip Ingham, at the Living Systems Institute, Exeter, UK and Nanyang Technological University, Singapore. Opportunities are available for lab placements in Prof. Ingham’s lab as part of the project. This offers an opportunity for a dedicated researcher to develop a truly interdisciplinary collaboration.
Interested candidates should contact Timothy Saunders (dbsste@nus.edu.sg), including a CV.
Location: Institut Curie, located in the center of Paris, is an internationally renowned institution bringing together physicists, chemists, biologists, bioinformaticians and clinicians.
Position:
Jean-Leon Maitre, head of the “Mechanics of mammalian development” team (science.institut-curie.org/team-maitre/), is seeking a motivated postdoc with a strong interest in interdisciplinaryresearch.
The candidate will study the morphogenetic events occurring before implantation of the embryo, which requires an approach at the interface between biology and physics (Maître et al, Nat. Cell Biol., 2015; Maître et al, Nature, 2016; Maître, Biol. Cell, 2017). The candidate’s work will include developmental biology techniques with recovery, culture and manipulation of mouse embryos; biophysical techniques such as high-resolution microscopy and micropipette manipulation of embryos; data and image analysis.
Skills:
Prior experience with mouse, advanced microscopy, molecular biology and/or image analysis will be extremely valuable, but on-the-job training will be additionally provided. The ideal candidate should feel comfortable working in an interdisciplinary and international environment.
The position is funded by the ERC for 24 months initially.
Interested candidates should contact jean-leon.maitre@curie.fr
A post by Jasmin Imran Alsous, on work done in collaboration with Paul Villoutreix and Alexander M. Berezhkovskii in the Shvartsman lab.
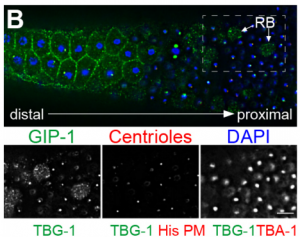
I started working on Drosophila egg chambers from the day I joined the Shvartsman lab. Egg chambers are small clusters of cells that eventually develop into mature oocytes in the abdomen of the female fruit fly. Perhaps not surprisingly, because the development of gametes, oocytes (or sperm), within such clusters is a conserved feature from insects to mammals, there is much to learn from probing their development within these simpler organisms. As I was taught how to dissect out the egg chambers and take images of the ovarioles – these are strings of egg chambers lined in a chronologically ordered array (Figure 1B) – I knew that regardless of what my project(s) turned out to be, it would have to involve these clusters: their structure was simple and images of the fluorescently labeled egg chambers were really beautiful. Both features meant I would not quickly tire of collecting data and could explain my research to non-biologists and non-scientists in general with little difficulty.
Interesting Phenomena in Developing Egg Chambers
Most questions we asked in the lab about how these egg chambers develop arose naturally from simply observing these structure as they formed and grew. First, the egg chamber is a cluster of exactly 16 germline cells that are enveloped by an epithelium. These 16 cells arise from a single founder cell that undergoes four synchronous and incomplete divisions: the cells remain connected through bridges, called ring canals, that allow for intercellular transport and communication (Figure 1A). One of these cells becomes the oocyte, and the other 15 develop into endoreplicating nurse cells that synthesize RNA, proteins, and organelles; these are then transported to the oocyte. Because the divisions are stereotypic, each of the 16 cells is uniquely identifiable, so one of the earliest questions we asked was whether the cells are packed or spatially arranged in a stereotypic manner also (this became the subject of another project). Second, as the egg chamber develops and grows, it does so non-uniformly, i.e. the tremendous gain in volume, by roughly 4 orders of magnitude, is distributed unequally among the 16 cells (Figure 1B). Interestingly, previous studies dating back to the late 50s had made note of this, and I was amazed at how much was known about the egg chamber already – its structure, its cell type composition, the specialized functionalities of the oocyte and nurse cells, its various stages of development – and frankly, a lot of the questions I was interested in were either posed in one way or another in these initial studies or inspired by them. However, without a proper sample size and comprehensive quantification, which would have been impossible without today’s imaging technologies, it was unclear whether there was a pattern to this nonuniformity, and by which mechanism it arose. The project was then to answer these two questions, and it eventually culminated in our recent Current Biology paper, “Collective Growth in a Small Cell Network”.
Figure 1. Differential Growth in the Drosophila Egg Chamber [1]. (A) Schematic representation of the four rounds of divisions that give rise to the egg chamber. The cells (nodes) remain connected by ring canals (edges). One of the cells with 4 ring canals becomes the oocyte; the other 15 become nurse cells. (B) Volume renderings of two ovarioles with egg chambers arranged chronologically from youngest (left) to oldest (right). Membranes (gray) and nuclei (red) are fluorescently labeled. Each egg chamber is a cluster of 16 germline cells connected as shown in (A) and is encapsulated by an epithelium. Throughout oogenesis, egg chambers grow by ~4 orders of magnitude, with unequal distribution of volume among the germline cells (yellow arrowheads). Scale bar, 10 um.
Differential Growth: Examples & Mechanisms
This phenomenon, of parts in growing objects growing at different rates, is not an unfamiliar one. Perhaps some of the clearest examples of that are the disproportionately large deer or elk antlers and fiddler crab claws (Figure 2A, B). These structures are evidently growing at faster rates than the animal’s other body parts. Another example closer to home is a human baby during postnatal development: born with a relatively large head and small limbs, the toddler develops into an adult with a proportionally smaller head and larger limbs (Figure 2C).
Figure 2. Familiar Examples of Differential Growth. (A) A fiddler crab with a disproportionately large claw [2]. (B) An elk with outsized antlers [3]. (C) Postnatal development of human babies: the final shape of the adult is unobtainable by pure dilation of the toddler [4].
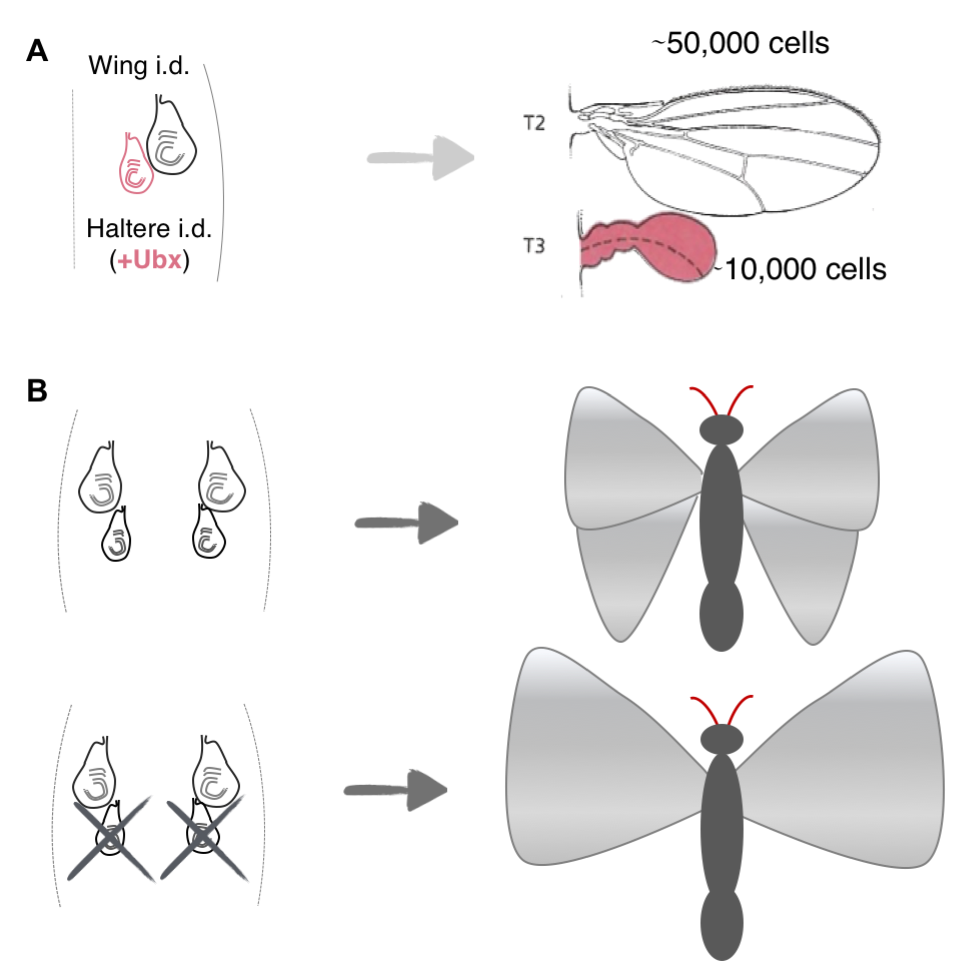
How such spatiotemporal patterns arise in growing organisms have fascinated researchers since Huxley and Thompson’s times [5, 6], and recently, studies have come up with potential mechanisms to explain these emergent collective growth patterns in model systems. For example, studies have shown that the size discrepancy between the Drosophila wings and halteres, which are a second and smaller pair of appendages (~fivefold fewer cells) used for balance and steering, can be explained by intrinsic differences, i.e. expression of the gene ultrabithorax (ubx) in the imaginal discs in the larva that give rise to the halteres, but not the ones that give rise to the wings (Figure 3A) [7]. Another elegant study showed that the difference in size between the fore and hind wings on butterflies, P. coenia, can be explained by competition for resources among the growing wing appendages (Figure 3, top). This was best exemplified by experiments that surgically removed the patch of cells that give rise to the hind wings: in such butterflies, the researchers noted a compensatory increase in the size of the forewings (Figure 3, bottom) [8]. Although these model systems are considerably simpler than whole organisms, they are still relatively complex: both systems comprise thousands of cells and the developing structures are not independent, but are parts of a whole.
Figure 3. Mechanisms for Spatiotemporal Growth Patterns in Model Systems. (A) Schematic showing the imaginal discs in the larva (left) that give rise to the wings (gray) and the halteres (magenta). The halteres are much smaller than the wings in the adult fly (right) due to expression of the gene ultrabithorax in all the cells of their imaginal discs. (B) Top: Schematic showing the imaginal discs in the caterpillars of P. coenia (left) that give rise to the fore-and hindwings in the adult butterfly (right). Bottom: Surgical removal of the imaginal discs that give rise to the hindwings result in a compensatory increase in the size of the forewings.
A Correlation & An Observation
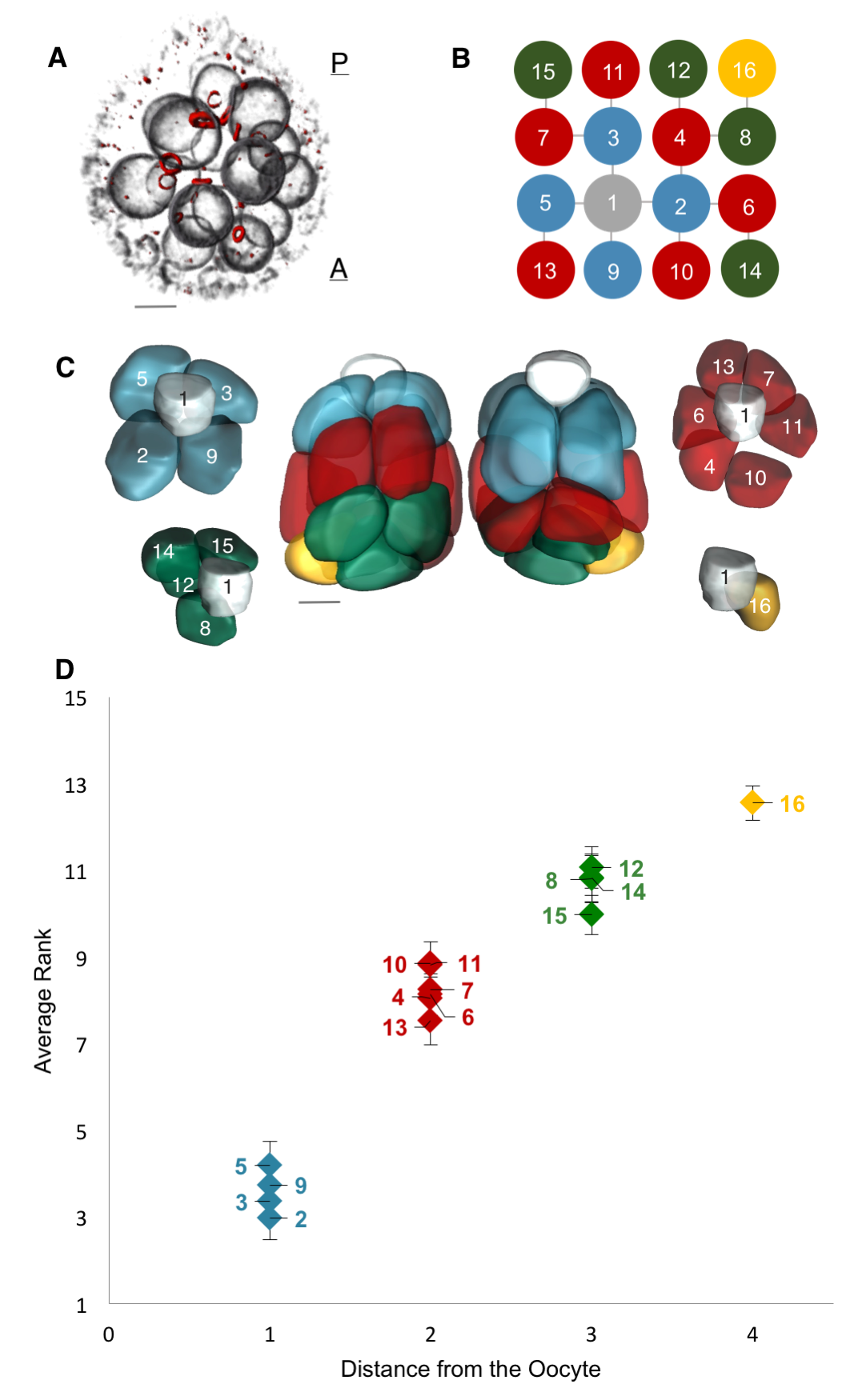
Motivated by these examples, we set out to quantify the nonuniformity of cell sizes in the growing egg chamber, and if possible, to describe and mechanistically explain the emergent pattern. To that end, we collected a large number of 3D confocal images of egg chambers that were immunostained to reveal their cell membranes, nuclei and ring canals. Using the stereotypic structure of the cell lineage tree, and starting with the oocyte, labeled as cell 1, we identified and annotated each of the 16 cells (Figure 4A, B). Furthermore, using nuclear and membrane volume measurements of reconstructed egg chambers, we had previously found that nuclear and cell volume correlate strongly, so for our subsequent measurements, we used nuclear volume as a proxy for cell volume – a much more easily measurable parameter than cell volumes: obtaining cell volumes required reconstructing the 3D shapes of each cell in the egg chamber, which involved drawing contours that traced the outlines of the cells’ membranes – a painstakingly slow process because the cells are irregularly shaped and thus far the process has not been automated. These reconstructions also revealed that the nurse cells are spatially organized into layers dictated by the number of ring canals separating any given nurse cell from the oocyte: arranged from posterior (closest) to anterior (furthest) of the oocyte, the numbers of nurse cells that the layers contain are 4, 6, 4, and 1 (Figure 4C).
Figure 4. A Layered Arrangement of Cells and an Emergent Pattern of Cell Sizes [1]. (A) Volume-rendered egg chamber showing its 3D structure, with fluorescently labeled nuclear envelopes (gray) and ring canals (red). The oocyte lies at the most posterior location of the egg chamber. (B) The ring canal tree, with nodes (cells) colored based on the number of edges (ring canals) from the oocyte (cell 1, gray). The tree’s invariant structure renders each germline cell uniquely identifiable. (C) Color-coded membrane-based reconstruction of a young egg chamber showing front and back views. Also shown is the layered spatial organization of nurse cells separated from the oocyte by one ring canal (blue), by two ring canals (red), by three ring canals (green), and by four ring canals (yellow), away from the oocyte. (D) A plot of each nurse cell’s average nuclear volume rank (1 = largest) as a function of distance from the oocyte across egg chambers sampled from all stages of oogenesis prior to stage 10 (n = 41). Nurse cells exhibit differential growth, and four groups of nurse cell sizes emerge that correlate with the spatial organization of the cells illustrated in (C). Error bars indicate SE. Scale bars in (A) and (C), 10 um.
A Pattern of Cell Sizes
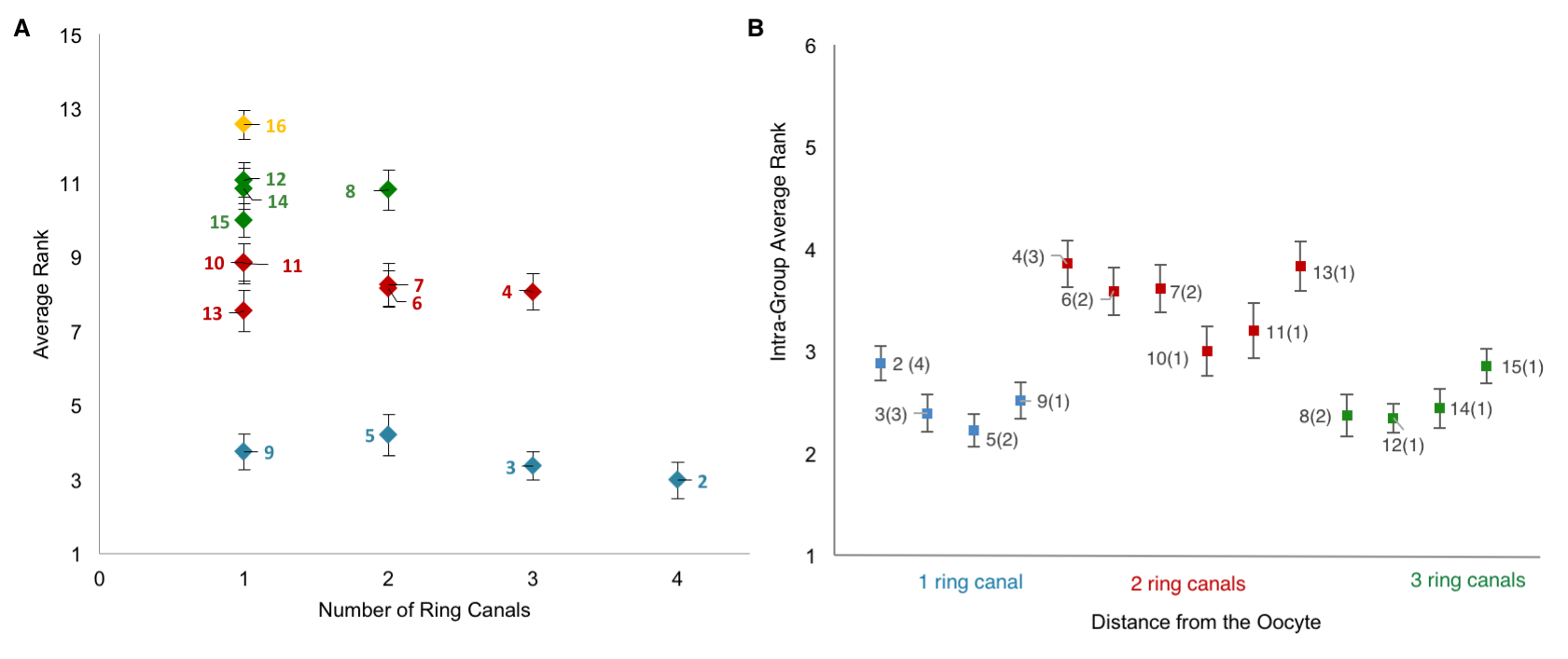
Collecting the data was a two-step process. First, we uniquely identified and labeled each of the cells in each collected 3D image of an egg chamber. Second, we collected nuclear volume measurements for each of those cells. To normalize for variations in growth among different egg chambers, we then ranked each nurse cell within each egg chamber by its nuclear volume. This gave us a vector of 15 rank entries, labeled 1–15, with 1 being largest and 15 smallest, and allowed us to calculate an average rank for each cell across all egg chambers. Once we had that data, it was unclear how to proceed or what correlations to look for: cell size as a function of ring canal number? Or cell size as a function of distance from the oocyte as defined on the lineage tree? Both factors were possible and had been proposed as potentially affecting cell size to some degree. After much plotting, it became clear that distance to the oocyte on the lineage tree was the primary factor affecting cell size (Figure 4D). Namely, we found that cells closer to the oocyte rank more highly and that, naturally, four groups of cell sizes emerge that correlate with their layered spatial organization relative to the oocyte. Interestingly, the number of ring canals each cell has appeared to not have much of an influence on cell size, within or across groups (Figure 5A, B). More importantly, we found that this pattern of cell sizes emerges from uniform initial conditions, only after the 16 cells are enveloped by the epithelium. These measurements, although critical, were more difficult to make: the divisions that give rise to the cyst take place inside a germarium, which is a structure at the vary anterior tip of the ovariole. This structure contains the stem cell niche and is extremely packed and crowded: identifying separate cysts and labeling the cells from 1-16 required high resolution images in x, y and z, a lot of zooming in and forgoing any sense of urgency.
Figure 5. Number of Ring Canals is Not the Primary Factor Affecting Cell Size [1]. A) A plot of each cell’s average rank as a function its ring canal number. B) A plot of intra-group average rank for each of the 16 germline cells; cells within a group were ranked relative to each other. There is no specific nurse cell nuclear volume distribution within a group that can be explained by the number of ring canals each cell has (in parenthesis). Error bars indicate SE (n=41).
Origin of This Pattern
Having established the pattern of cell sizes, our next step was to come up with the simplest mathematical model that can both explain our observations and make predictions. This process of developing and writing out the model was perhaps the most exciting, but the most iterative: the model was written and re-written more times than I can remember, mainly because at each iteration, we found that the model could be simplified further. In fact, the final version that appears in the paper is a model that exhibits how differential growth arises already in a 2-cell system, and how groups of cell sizes arise in a 4-cell system; these results are easily generalizable to the 16-cell system and there was little need to write out the equations for all 16 cells in the main body of the paper (these made their cumbersome way to the supplementary material).
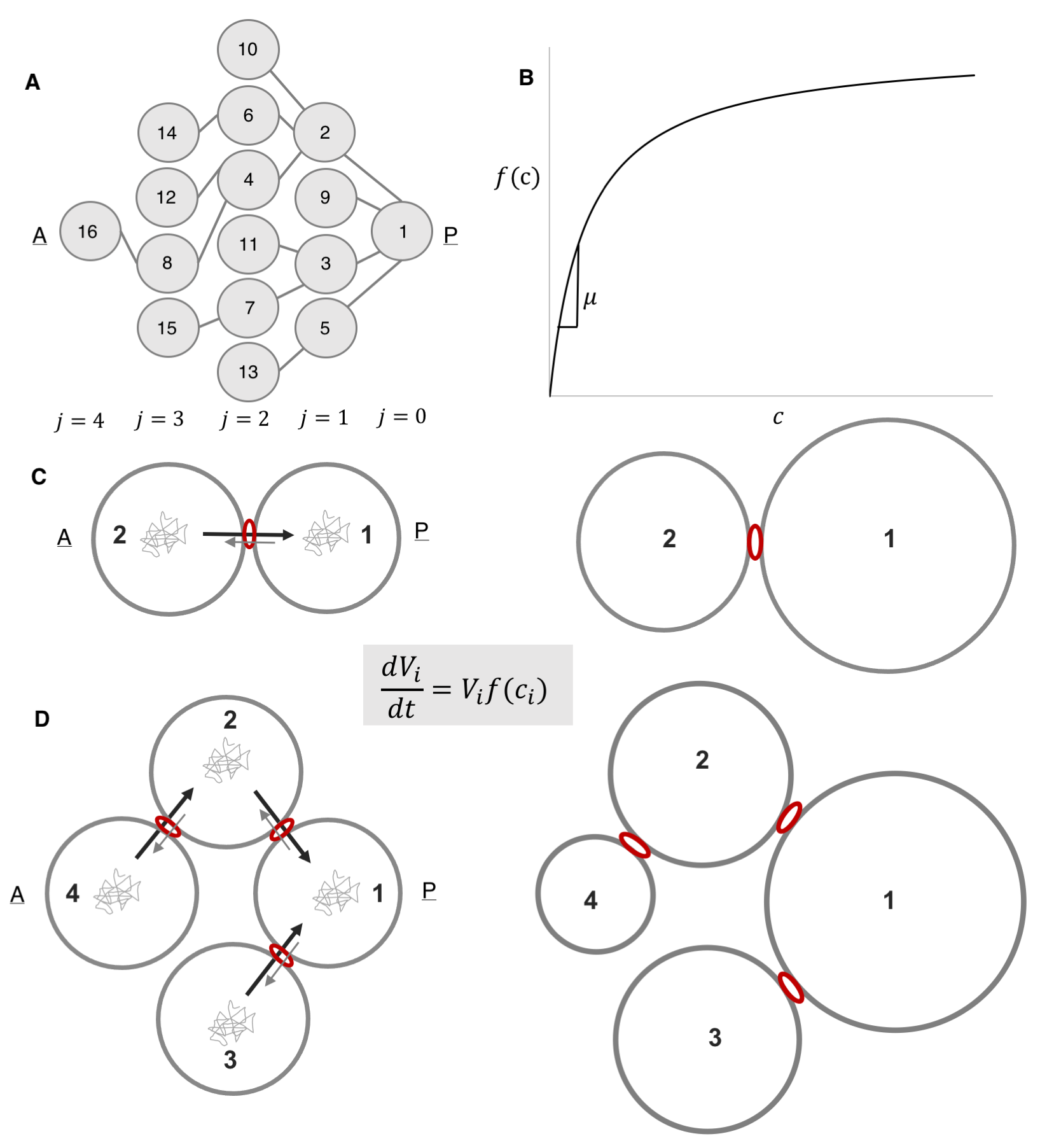
We started with a coarse-grained model whose variables corresponded to cell volumes, which increase according to Monod-like kinetics: proportional to the product of the current cell volume of cell and a monotonically increasing function of the intracellular concentration of some limiting component (Figure 7B). Intuitively, differences in the cellular concentrations of the limiting factor result in different specific growth rates among the cells. Notably, we were unsure as to what that limiting component was: was it a nutrient, a protein, an organelle? Were there several limiting components? The study alluded to in figure 3B that established that the size discrepancy between the fore and hind wings in the butterfly was due to competition for resources remained “deliberately vague” as to the nature of the shared components, so we felt emboldened to do just the same. Figuring out the nature of that component became an item on the future works agenda.
We then postulated that this component is exchanged among the cells and that this exchange is polarized, such that the probability of being transported through a ring canal connecting any two cells is higher in the direction of the more posterior cell, i.e. the cell closer to the oocyte. Here we also defined the parameter v, which quantifies the asymmetry of transport through the ring canals such that when transport is polarized in the posterior direction, v<1. Notably, this bias in transport across the ring canals is made possible by a polarized network of microtubules that is set up following specification of the oocyte and that extends from the oocyte to the nurse cells through the ring canals, connecting all cells within the germline cyst (Figure 6A) [9, 10]. Furthermore, several particle tracking studies in developing egg chambers showed that particles diffuse randomly in the cytoplasm of the cell, and while particles can diffuse bi-directionally across a ring canal, once at a ring canal, these particles exhibit a posterior bias in their translocation (Figure 6B) [11, 12, 13, 14].
Figure 6. Inter- and Intra-cellular Transport. (A) Schematic of an egg chamber illustrating the network of microtubules (MTs) that extends from the oocyte to the nurse cells through the ring canals (modified from [9]) (B) Particles diffuse randomly in the cytoplasm of the cell, and while particles can diffuse bi-directionally across a ring canal (gray spots), once at a ring canal, these particles exhibit a posterior bias in their translocation (green arrows).
We also assumed that concentrations within the cell cluster equilibrate rapidly and can therefore be considered at steady state on the much longer timescale of growth of the cyst. This assumption was based on the difference between our estimate of the time it takes a diffusing particle to reach a circular window of some radius on the wall of a cell with some volume, and the time scale for growth. The former was estimated to be around 10 minutes; the latter, ~ 3 days. Note that it helped that our advisor and our collaborator (SYS and AMB) had co-authored a paper on the kinetics of escape through a small hole – one of the main results of that paper was an analytical expression for the rate constant of escape, k, in terms of the problem’s parameters: the diffusion constant, D,of the particle, the hole radius, a, and the cavity volume V : k=4Da/V [15].
We then wrote out equations illustrating how the fluxes between any two connected cells balance, and given that the fluxes can be expressed in terms of a rate constant for diffusion multiplied by the cellular concentration of our limiting component, this gave us relationships that related (1) the concentrations between any two anterior and posterior cells separated by either a single ring canal and (2) the intercellular concentration of any anterior cell relative to the oocyte, cell 1. Based on these assumptions alone, the model explained both the observed pattern of differential growth and the emergence of groups in wild type egg chambers. Specifically, when transport is polarized toward the founder cell, i.e. v<1, the 2-cell system gives us differential growth (Figure 7C), and the 4-cell system gives us both differential growth and emergence of (three) groups (Figure 7D). These results were easily generalizable to the 16-cell cyst.
Figure 7. A Biophysical Model for Differential Growth and Emergence of Groups [1]. (A) Schematic representation of the ring canal tree arranged to highlight the anterior nurse cells’ spatial organization as it relates to the posterior oocyte (A, anterior; P, posterior). Each cell (node) is numbered (1–16), and each layer j is labeled (B) A plot of the specific growth rate, f(c), as a function of concentration, c, showing the linear and saturated regimes. (C) Schematic representation of a 2-cell cluster whose cells are connected by a ring canal. Each cell i, of volume V, grows according to the growth law shown. Diffusing particles of concentration, c, in the cells’ cytoplasm that arrive at the ring canal have a higher probability of being transported from an anterior (A) cell to the more posterior (P) cell than in the opposite direction. The result is that two cells, initially of uniform size, will grow at unequal rates, with cell 1 (posterior) getting larger than cell 2 (anterior). (D) Schematic representation of a 4-cell cluster whose cells are connected by ring canals. Allowing for polarized transport in the posterior direction, the 4-cell cluster model leads both to differential growth and to the emergence of 3 groups of cell sizes that correlate with the spatial arrangement of the layers relative to cell 1.
Our model also made a prediction, namely, that in the absence of a polarized transport network and polarized transport (v=1), cells in the growing cluster would be of the same size. We therefore examined mutants that fail to set up a polarized microtubule cytoskeleton for transport. Luckily, there are several such mutants available but we chose egalitarian (egl) in which no oocyte is specified, the formation and maintenance of a polarized microtubule network are disrupted, and all 16 cells develop into polyploid nurse cells [9]. We showed that in such mutants, the group-wise arrangement of cell sizes observed in wild type egg chambers was absent: the average size ranks of cells from virtually all groups overlap. These results suggested that the relative absence of both differential growth and emergence of groups of cell sizes is attributable to the absence of a polarized transport within the cluster.
Allometric scaling
We were pretty happy with the model: it was simple, had few assumptions and could explain our observations in both wild type and mutant egg chambers. But we wondered whether we had extracted all of what the model had to offer, and we had not. By making an additional assumption, namely that we were in the linear regime of the proposed growth law (shown in Figure 7B), we could extract a quantitative relationship between the volumes of cells belonging to different layers in the cell network in terms of already-defined problem parameters.
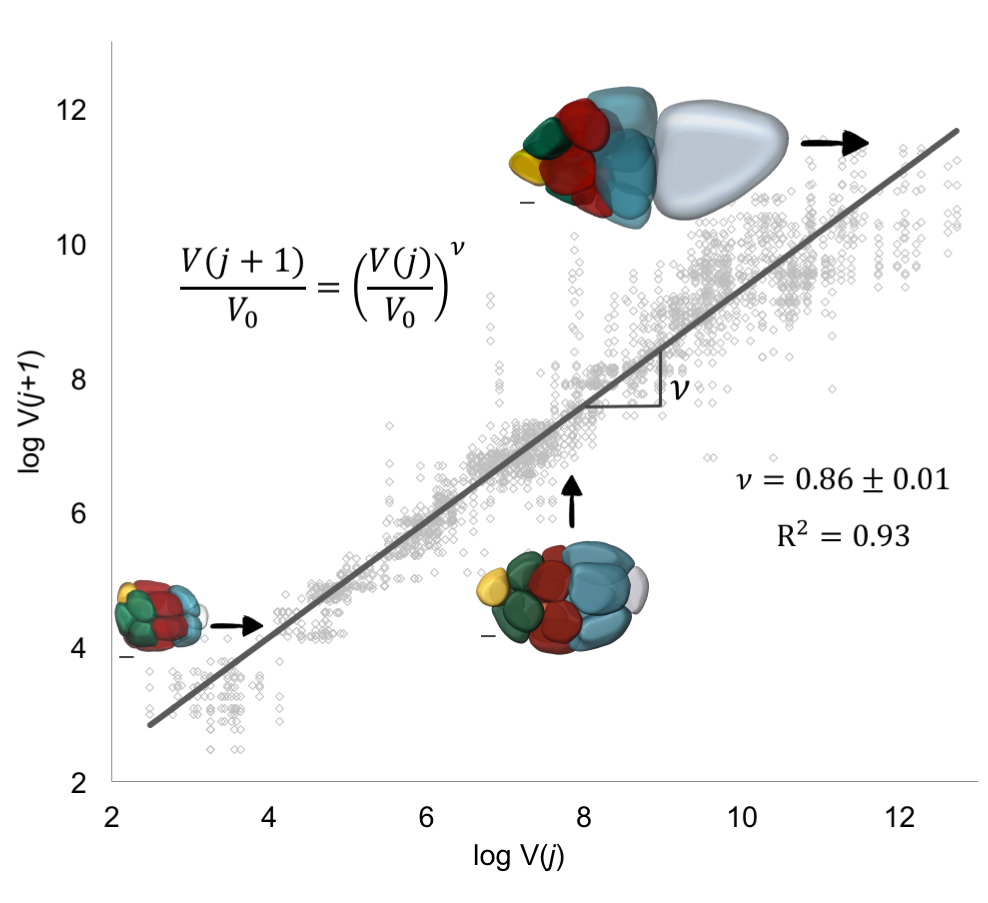
Figure 8. Allometric Scaling in Collective Cell Growth [1]. To extract v, we fit the logarithmic form of the equation shown in the figure to data pairs of nuclear volumes of nurse cells from consecutive layers j and j + 1. The extracted value of v is consistent with biased transport toward the posterior oocyte. The insets show representative color-coded membrane-based reconstruction of a young, roughly spherical egg chamber (bottom left), an egg chamber beginning to elongate (middle), and an older ellipsoidal egg chamber (top right). Scale bars, 10 um.
This result now predicted a specific quantitative pattern for a thus far qualitative observation, namely, that the pattern of decreasing cell sizes with distance from the oocyte at any moment in time can be quantified with an allometric-type relationship. The only parameter here was v, which linked a process occurring at the molecular scale, transport of an intracellular limiting component, to an observable phenotype, cell size. To estimate v, took the natural logarithm of both sides of the equation in the previous paragraph: the value of v was then given by the slope of the line that best fits through pairs of nuclear volumes of nurse cells from consecutive layers j and j + 1; each egg chamber has 52 such pairs (Figure 8). This lead to v =0.86±0.01. For this value of , which is less than 1, transport is polarized (biased towards the oocyte), and a growing egg chamber will have four groups of nurse cells that exhibit a posterior-to-anterior gradient of decreasing cell sizes, which is consistent with what we observed experimentally.
We were particularly happy with this result because in addition to deriving the quantitative relation between volumes of cells from different layers, we also extracted the parameter characterizing the extent of polarized transport within the cluster, thus linking a subcellular molecular process to an emergent pattern of collective cell growth. It is not often that one can link processes that span different scales in biology, regardless of how phenomenological the starting model is, and being able to come back full circle to Huxley’s allometries is cool.
This paper was perhaps one of the most rewarding project I had been involved in so far – mainly because we finally answered basic questions about growing egg chambers that we had been interested in for quite some time, and because the model that explained our results was relatively simple and could be dissected and understood to the end. Perhaps the biggest challenge in writing up the paper was deciding how to most clearly explain our experimental observations and theoretical results to a broad community of cell, developmental, and quantitative biologists, as well as applied mathematicians: we wanted to avoid writing a cumbersome paper but not at the risk of imprecision. However, having several lab mates, collaborators and, of course, co-authors from various backgrounds who read and re-read the manuscript streamlined that process and ensured that we used the most correct yet simplest terminology, so thanks friends.
[11] Bohrmann, J., and Biber, K. (1994). Cytoskeleton-dependent transport of cytoplasmic particles in previtellogenic to mid-vitellogenic ovarian follicles of Drosophila: time-lapse analysis using video-enhanced contrast microscopy. J. Cell Sci. 107, 849–858.
[13] Snee, M.J., and Macdonald, P.M. (2004). Live imaging of nuage and polar granules: evidence against a precursor-product relationship and a novel role for Oskar in stabilization of polar granule components. J. Cell Sci. 117, 2109–2120.
[14] Shimada, Y., Burn, K.M., Niwa, R., and Cooley, L. (2011). Reversible response of protein localization and microtubule organization to nutrient stress during Drosophila early oogenesis. Dev. Biol. 355, 250–262.
Here are the highlights from the current issue of Development:
Separating nematode species by spermatogenesis
During spermatogenesis, unnecessary organelles and cytoplasmic components are shed from developing sperm in order to streamline them for optimal motility. These components are partitioned into structures known as ‘residual bodies’, which subsequently separate from the sperm and are lost. In the nematode worm C. elegans, this partitioning occurs immediately after the meiotic divisions, thus accelerating the process of sperm production. On p. 3253, Diane Shakes and colleagues exploit the interspecies diversity in spermatogenesis among nematodes to reveal how cellular components become partitioned. Focussing on the role of microtubules and actin, they characterise the process in C. elegans and in another nematode species, Rhabditis sp. SB347, and find important mechanistic variations between the two. In Rhabditis, which produces small spermatocytes, two rather than four sperm are generated during meiosis; the rest of the genetic material is partitioned into residual bodies. Interestingly, this mirrors oocyte production and resulting polar-body formation in females. These results provide insight into both the conserved and divergent mechanisms that underpin partitioning during spermatogenesis, and reveal how these segregation mechanisms can be modulated to achieve differences between species.
Sending signals through SMADs: how blood flow shapes arteries
During organogenesis, mechanical forces can induce transcriptional and cytoskeletal changes in cells that help shape tissues as they develop. However, the mechanisms allowing cells to sense and respond to these mechanical signals are poorly understood. In the cardiovascular system, endothelial cells, which line the arteries, are able to change their shape in response to high or low blood flow through an artery, resulting in a change to the vessel’s diameter. On p. 3241, Kristy Red-Horse and colleagues investigate how mechanical signals are transduced by endothelial cells to regulate the size of arteries, and show that SMAD4 signalling mediates this response. When SMAD4 is deleted in mice, coronary artery size is increased, subsequent to the onset of blood flow. They also show that in human coronary artery endothelial cells cultured in vitro, knockdown of SMAD4 leads to defects in flow-guided cell elongation and migration. Additionally, they find that these cells undergo increased proliferation when exposed to shear stress. Their data directly link BMP/SMAD signalling to endothelial changes in response to mechanical force. These results increase our understanding of how forces can regulate tissue development during embryogenesis, and might be important in developing treatments for human vascular pathologies.
Opening up: new insights into chromatin decondensation
During heat shock, cells respond to temperature stress by opening their chromatin, allowing the transcription of genes that enable them to cope with the sudden change in the environment. How chromatin becomes decondensed to permit active transcription during this process is not clear. Recently, a model based on experiments in Drosophila has been proposed, whereby histone H2Av is deposited and subsequently phosphorylated by JIL-1 kinase, followed by recruitment of poly(ADP-ribose) polymerase 1 (PARP-1). PolyADP-ribosylation of chromatin then takes place, which loosens its structure. This permits phosphorylation of serine 10 in the tail of histone H3 (H3S10p), again by JIL-1 kinase, which is required for the function of the transcriptional machinery. On p. 3232, Kristen Johansen and colleagues test this model using null mutants and find that H2Av phosphorylation and chromatin opening can occur in the absence of JIL-1 kinase, and that H3S10p still occurs in a PARP-1 knockdown mutant. In light of these findings, the proposed model breaks down. Instead, the authors find that PARP-1 can be recruited by H3S10p independently of H2Av, providing insight into an alternative mechanism for opening up of chromatin structure to permit active transcription in Drosophila.
Histone demethylase functions in fertility
The acquisition and removal of epigenetic marks can help modulate gene expression during development by altering chromatin structure. Di- or tri-methylation of histone H3 on lysines 9 and 36 (H3K9/36) is associated with gene repression and silencing, since these marks induce a closed chromatin state. Kdm4a is an enzyme that demethylates H3K9/36, and functions to prevent the build-up of methyl groups at these sites in order to maintain active transcription. On p. 3264, Kristian Helin and colleagues now reveal a role for Kdm4a in female fertility. They show that this histone demethylase is expressed in all tissues of the female reproductive system, including the oocyte. While mice lacking the enzyme are able to ovulate and fertilise embryos normally, pregnancies fail because the embryos do not implant in most cases. Although the morphology of the reproductive tract is not altered in these animals, gene expression analysis shows that genes important for uterine receptivity are downregulated. Furthermore, maternal Kdm4a is also required in the oocyte to generate viable embryos, with knockout embryos arresting within the first few days after fertilisation. These results provide insight into how chromatin regulation through epigenetic factors can impact on physiological processes, including fertility.
PLUS…
An interview with George Daley
George Daley is Dean of the Faculty of Medicine, Professor of Biological Chemistry and Molecular Pharmacology, and Caroline Shields Walker Professor of Medicine at Harvard Medical School. A former Howard Hughes Medical Institute Investigator and President of the International Society for Stem Cell Research (ISSCR) from 2007-2008, his lab works on the biology and clinical application of stem cells, with a particular focus on hematopoiesis. He was awarded the Public Service Award at the ISSCR 2017 meeting in Boston, where we caught up with him to discuss his move from the lab to the clinic and back again, his quest to derive human hematopoietic stem cells in vitro, and his advocacy for science in uncertain political times. Read the Spotlight article.
Metabolism in time and space – exploring the frontier of developmental biology
In May 2017, a diverse group of scientists assembled at the EMBO/EMBL Symposium ‘Metabolism in Time and Space’ to discuss how metabolism influences cellular and developmental processes. The speakers not only described how metabolic flux adapts to the energetic needs of a developing organism, but also emphasized that metabolism can directly regulate developmental progression. Overall, this interdisciplinary meeting provided a valuable forum to explore the interface between developmental biology and metabolism. Read the Meeting Review by Alena Krejci and Jason Tennessen.
The enigma of embryonic diapause
Embryonic diapause – a period of embryonic suspension at the blastocyst stage – is a fascinating phenomenon that occurs in over 130 species of mammals, ranging from bears and badgers to mice and marsupials. It might even occur in humans. During diapause, there is minimal cell division and greatly reduced metabolism, and development is put on hold. Yet there are no ill effects for the pregnancy when it eventually continues. Multiple factors can induce diapause, including seasonal supplies of food, temperature, photoperiod and lactation. The successful reactivation and continuation of pregnancy then requires a viable embryo, a receptive uterus and effective molecular communication between the two. In their Primer article, Marilyn Renfree and Jane Fenelon provide an overview of the process of diapause, focusing on recent molecular data.
The goal of this project is to engineer therapeutically active islet-like aggregates for future cell therapy phase 1 trials in Type 1 Diabetes (T1D).
Job Description:
We are looking for a postdoctoral candidate with a strong cell biological and cell signaling background in directed differentiation of human pluripotent stem cell biology. Experience in differentiation towards pancreatic lineages is a merit. The candidate is expected to work on the engineering of human pluripotent stem cell-derived islet of Langerhans-like aggregates with functional properties close to human islets. The functionality and therapeutic potential of the aggregates will be tested in vitro and in vivo in mice. The candidate is expected to use state-of-the-art genetics, molecular and cell biological, and tissue engineering experimental strategies. The candidate will work together with a dedicated team of scientists and technicians who together will tackle bottle-necks towards implementing the phase 1 clinical trials in T1D.
Qualifications:
The candidate is required to hold a PhD degree in pluripotent stem cell/developmental biology. A few years of postdoctoral experience in the same areas is a merit. The candidate should also have hands on experience in human pluripotent stem cell maintenance and differentiation, 3D culture of pluripotent stem cells, various cell and molecular biological methods, flow cytometry and live-cell imaging. Finally, we are looking for applicants with a good record of peer reviewed scientific publications, grant writing skills and an interest in team work.
Employment Conditions:
The position is for 2 years with a possible extension. The employment is planned to start as soon as possible or upon agreement with the chosen candidate.
The terms of employment are set according to the Agreement between the Ministry of Finance and The Danish Confederation of Professional Associations or other relevant professional organization. The position will be at the level of postdoctoral fellow and the basic salary according to seniority is 32.700-34.400 DKK/month. A supplement could be negotiated, dependent on the candidate´s experiences and qualifications. In addition, a monthly contribution of 17.1% of the salary is paid into a pension fund.
Applicants recruited from abroad may be eligible for a special researcher taxation scheme. In all cases, the ability to perform the job will be the primary consideration, and thus we encourage all – regardless of their personal background and status – to apply.
The application must be submitted online and in English, by clicking on “Apply online” below.
The closing date for applications is 23.59pm, November 1st 2017.
The application must include:
Cover letter detailing the basis on which the applicant scientific qualifications meet the requirements for this position.
Curriculum vitae.
List of references (full address, incl. email and phone number)
Diplomas – all relevant certificates.
List of publications.
Application procedure:
After the expiry of the deadline for applications, the authorized recruitment manager selects applicants for assessment on the advice of the Appointments Committee. All applicants are then immediately notified whether their application has been passed for assessment by an expert assessment committee. Selected applicants are notified of the composition of the committee and each applicant has the opportunity to comment on his/her assessment. You may read about the recruitment process on http://employment.ku.dk/faculty/recruitment-process
DanStem and The University of Copenhagen wish to reflect the diversity of society and welcome applications from all qualified candidates, regardless of personal background.
Founded in 1479, the University of Copenhagen is the oldest university in Denmark. With 37,000 students and 9,000 employees, it is among the largest universities in Scandinavia and one of the highest ranking in Europe. The University consists of six faculties, which cover Health and Medical Sciences, Humanities, Law, Science, Social Sciences and Theology.
 (No Ratings Yet)
(No Ratings Yet)






 (2 votes)
(2 votes)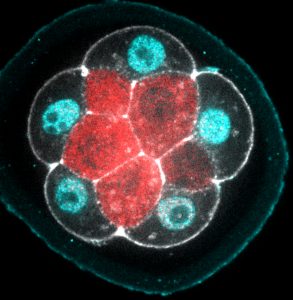







 George Daley is Dean of the Faculty of Medicine, Professor of Biological Chemistry and Molecular Pharmacology, and Caroline Shields Walker Professor of Medicine at Harvard Medical School. A former Howard Hughes Medical Institute Investigator and President of the International Society for Stem Cell Research (ISSCR) from 2007-2008, his lab works on the biology and clinical application of stem cells, with a particular focus on hematopoiesis. He was awarded the Public Service Award at the ISSCR 2017 meeting in Boston, where we caught up with him to discuss his move from the lab to the clinic and back again, his quest to derive human hematopoietic stem cells in vitro, and his advocacy for science in uncertain political times. Read the
George Daley is Dean of the Faculty of Medicine, Professor of Biological Chemistry and Molecular Pharmacology, and Caroline Shields Walker Professor of Medicine at Harvard Medical School. A former Howard Hughes Medical Institute Investigator and President of the International Society for Stem Cell Research (ISSCR) from 2007-2008, his lab works on the biology and clinical application of stem cells, with a particular focus on hematopoiesis. He was awarded the Public Service Award at the ISSCR 2017 meeting in Boston, where we caught up with him to discuss his move from the lab to the clinic and back again, his quest to derive human hematopoietic stem cells in vitro, and his advocacy for science in uncertain political times. Read the  Embryonic diapause – a period of embryonic suspension at the blastocyst stage – is a fascinating phenomenon that occurs in over 130 species of mammals, ranging from bears and badgers to mice and marsupials. It might even occur in humans. During diapause, there is minimal cell division and greatly reduced metabolism, and development is put on hold. Yet there are no ill effects for the pregnancy when it eventually continues. Multiple factors can induce diapause, including seasonal supplies of food, temperature, photoperiod and lactation. The successful reactivation and continuation of pregnancy then requires a viable embryo, a receptive uterus and effective molecular communication between the two. In their
Embryonic diapause – a period of embryonic suspension at the blastocyst stage – is a fascinating phenomenon that occurs in over 130 species of mammals, ranging from bears and badgers to mice and marsupials. It might even occur in humans. During diapause, there is minimal cell division and greatly reduced metabolism, and development is put on hold. Yet there are no ill effects for the pregnancy when it eventually continues. Multiple factors can induce diapause, including seasonal supplies of food, temperature, photoperiod and lactation. The successful reactivation and continuation of pregnancy then requires a viable embryo, a receptive uterus and effective molecular communication between the two. In their