Organs develop from a few founding cells that divide to produce descendants. In a recent study published in the journal Nature Communications, the group of Professor Anne Grapin-Botton at DanStem shows that some of the cells in the community forming the pancreas will make large family trees while others will generate a few daughters or will remain single.
In addition, the founder cells also show heterogeneous abilities to produce daughters that make digestive enzymes or cells that make hormones that regulate blood sugar.
This fundamental finding could also be true for other organs and could be exploited to select the most powerful “organ making cells” in vitro. Selecting such powerful cells assumes that the ability to generate large families is predetermined and that such cells can be identified by certain markers.
Investigating a battery of markers, Hjalte list Larsen, the primary investigator of the study, showed that one gene activity discriminates the cells that make small families composed only of hormone-making cells. Surprisingly, half of the organ-founder cells were of this type, although the hormonal cells form only 1% of the adult organ. ”We wondered why so many of these hormone-making cells were set aside so early. We think that it is important to make many of these cells early because they nurture their neighbors, enabling them to divide extensively.” says Anne Grapin-Botton.
In the video: The cells of the two embryonic pancreas buds (round) are marked in bright red while the lighter red shows their connection to the intestine. The two green cells in the pancreatic bud (top) are two daughters made by one mother cell.
This study also illustrates how powerful the collaboration between biologists and physicists can be. This was the aim of Grundforskningsfonden when they promoted the STEMPHYS project enabling these interactions. Ala Trusina and her team from the Niels Bohr Institute could indeed use computer modelling to reveal that many decisions observed are not predictable and are likely triggered by a communication signal released by some cells, with a probability that it reaches another cell. The ‘ball’ is back on the biologists’ side to find it, to realize the promise of producing organs or specific cells types for regenerative purposes in a petri dish!
An exciting role has come up to be a part of a pioneering biomedical research institute, dedicated to innovation and science. Dr Frickel’s laboratory focuses on elucidating the immune mechanisms targeting the parasite Toxoplasma gondii, as well as deciphering pathways mediated by immune-activated large GTPases. Dr Frickel is seeking a talented and motivated postdoc to further develop this study in human stem cell-derived macrophages and dendritic cells, as a complement to studying these pathways in conventional macrophage cell lines.
Infection with the parasite Toxoplasma gondii causes the production of the cytokine gamma interferon (IFN) that in turn leads to the upregulation of a multitude of host defence pathways. Specifically, the lab is interested in ubiquitin-mediated pathogen control (Clough et al, 2016, PLoS Pathogens) and restriction of pathogens by guanylate binding proteins (Johnston et al, 2016, Cellular Microbiology). We are now looking to extend these findings to macrophages as these present the most relevant cell type for Toxoplasma and other infectious agents. We have generated iPSC-derived macrophages as a tool to study IFN-dependent host defence mechanisms. This project will examine how relevant ubiquitin-mediated pathways are to Toxoplasma control in these macrophages and/or how guanylate binding proteins mediate immune defences.
The Francis Crick Institute is a biomedical discovery institute dedicated to understanding the fundamental biology underlying health and disease. Its work is helping to understand why disease develops and to translate discoveries into new ways to prevent, diagnose and treat illnesses such as cancer, heart disease, stroke, infections, and neurodegenerative diseases.
An independent organisation, its founding partners are the Medical Research Council (MRC), Cancer Research UK, Wellcome, UCL (University College London), Imperial College London and King’s College London.
The Crick was formed in 2015, and in 2016 it moved into a brand new state-of-the-art building in central London which brings together 1500 scientists and support staff working collaboratively across disciplines, making it the biggest biomedical research facility under a single roof in Europe.
The Francis Crick Institute will be world-class with a strong national role. Its distinctive vision for excellence includes commitments to collaboration; to developing emerging talent and exporting it the rest of the UK; to public engagement; and to helping turn discoveries into treatments as quickly as possible to improve lives and strengthen the economy.
The closing date for applications is Sunday 29th October 2017 at 23:30.
Applications are invited for a Research Assistant/Associate within the Division of Infection & Immunity at UCL in Dr Gillian Tomlinson’s laboratory.
We are seeking a highly motivated individual interested in integrating cutting-edge human and zebrafish models to study the immunopathogenesis of tuberculosis. The post is funded by a Medical Research Council Clinician Scientist Fellowship entitled “Tuning the immune response in tuberculosis”, and combines a human experimental tuberculosis challenge model with studies using Mycobacterium marinum infection of zebrafish to identify and validate host factors that calibrate a favourable immune response in tuberculosis.
The post-holder will be supervised by Dr Gillian Tomlinson based in the Cruciform Building at UCL. Dr Tomlinson works closely with Dr Mahdad Noursadeghi’s and Professor Benny Chain’s group’s which study host immune responses to infectious diseases at genome‑wide level with a particular focus on tuberculosis (www.innate2adaptive.com). The zebrafish work will be supported by the fully managed world class research aquarium at UCL.
The post is available until 1st March 2020, subject to satisfactory probationary and annual appraisals. There is an established track record for department post-doctoral staff gaining personal fellowships. Independently minded and talented investigators will be encouraged and supported in seeking such fellowship support.
Key requirements
Applicants must have an MSc (or equivalent degree) and/or a PhD (or equivalent degree) in a relevant subject.
Candidates must be competent in molecular and cellular biology techniques including RNA and DNA extraction, qPCR, PCR, cloning and tissue culture.
I recently participated in “Skype a Scientist” – a program self-described as one that “matches scientists with classrooms” and “give(s) students the opportunity to get to know a ‘real scientist’”. Pretty accurate. Basically, if a scientist signs up, she will be matched with a k-12 classroom somewhere around the world. After some coordination with the classroom teacher, she will get the chance to describe her work to a group of eager youngsters in about half an hour to an hour. Needless to say, it is an opportunity worth exploring.
For those of us unsettled by science’s recently more pronounced credibility crisis, marked by uninformed, hence unhealthy skepticism, we can take comfort in the fact that this program is exactly what is needed to (1) spread the word – that science is interesting, thorough and important, and (2) pique the next generation’s interest in science. The underlying importance of this initiative aside, there are some incredibly gratifying reasons to participate.
First, you get to figure out how to explain your work to kids and why you are doing it – which is not easy. This is the stuff of grant-writing ! I work on understanding growth dynamics in developing fruit fly egg chambers, and when I presented to third graders, I was asked questions such as what can we learn from the fruit fly that would help us humans? Try answering that without using the word ‘conserved’. It is no secret that scientist often find it challenging to explain their work to others outside their field, and there is hardly a better exercise in distilling one’s work to the absolute basics than “Skype a Scientist”.Second, you get to inform kids and get them excited about novel and potentially game changing work, and perhaps even to express a desire to pursue a career in science. I often think of scientists as individuals who wander through the world and ask questions about their environment – the whys and the hows. In that sense, kids are already scientists: indeed, there is a hardly a group of individuals more curious about their surroundings, and if we can find ways to nourish and support their interests, then we are doing each other and the pursuit of science a great favor.
So if you find half an hour to spare, please do participate !
New research conducted at the stem cell centre, DanStem, at the University of Copenhagen shows that insulin is a key determinant of embryonic stem cell potency in mammals. When large amounts of Insulin are around, stem cells retain their ability to make all the cell types in the body. However, too little insulin leads to embryonic stem cells being transformed into a new type of stem cell, one that can make tissues that support foetal development and helps make the different internal organs. As embryonic stem cells come from embryos around the time they implant into the mother, this study suggests that maternal insulin and diet maybe be important for the earliest stages of pregnancy. This study also points to new ways that stem cells can be made and differentiated to help treat degenerative diseases.
Insulin was found to be a new factor that is important for the identity of pluripotent stem cells, cells able to make all the cells in the body. A new study conducted at the Novo Nordisk Foundation Center for Stem Cell Biology, DanStem, at the University of Copenhagen reveals that insulin acts in unexpected ways to stabalise stem cells in the laboratory and on embryos cultured in a dish.
“As this type of stem cell is like the cells of the early embryo, it suggests that insulin could also be important for human development implying maternal diet and insulin levels could effect the earliest stages of a healthy pregnancy.
Professor Joshua Brickman
‘We were exploring how stem cells respond to signals produced by other cells, instructions that tell them to develop into cells that are specialized for organ and intestine formation, the endoderm. But then, when we added these factors to the food (or media) normally used to grow stem cells in, we were surprised that these signals could instruct stem cells to stay as stem cells. By comparing different medias we discovered a key difference, Insulin. With Insulin, they stay as stem cells, but with out it they make a special type of endoderm. As this type of stem cell is like the cells of the early embryo, it suggests that insulin could also be important for human development implying maternal diet and insulin levels could effect the earliest stages of a healthy pregnancy, says head of the study Professor Joshua Brickman from DanStem. At the same time, he stresses that some of these ideas are still only conjecture and a lot of work will now need to be done to understand the relationship between maternal insulin, implantation and early development before researchers can make concrete recommendations.
Transmitter Substances Play a Main Role in the Discovery The researchers in the study have examined in detail two types of stem cell which resemble each other. One kind – the embryonic stem cells, also called pluripotent stem cells – have the ability to support the development of the entire foetus. These cells can develop into any types of cell in the body. The other kind is the extraembryonic cells, which produce a type of endoderm that makes tissues that support foetal development known as the yolk sac and also helps make the internal organs, for example the intestinal system.
Mouse embryo that has been injected with embryonic stem cells grown in Insulin showing these cells can make all cell types. The embryos are at the stage when they are making all the different basic cell types of the future body, gastrulation. The stem cells were labelled genetically so they fluoresce in red under the microscope.
“The mechanisms we have uncovered are very interesting. This suggests that different amounts of insulin cause cells to respond differently to the same signals. So far we have only done tests on mice, but the next step is to examine whether the same mechanisms are found in humans.
Post.doc. Kathryn Anderson
The researchers have studied two transmitter or signal substances (Nodal and Wnt), which play a main role in stem cell development. Here they discovered that the transmitter substances were able to promote cell division in the endoderm and, at the same time, supported cell division among the pluripotent cells, but that they could choose which cell they supported based on what insulin was telling them to do. If the researchers removed insulin, the pluripotent stem cells stopped dividing and endoderm cells replaced them. When Insulin was there, the pluripotent cells grew and retained the ability to become any cell in the body.
‘The mechanisms we have uncovered are very interesting. This suggests that different amounts of insulin cause cells to respond differently to the same signals. So far we have only done tests on mice, but the next step is to examine whether the same mechanisms are found in humans’, says post.doc. Kathryn Anderson.
Applications are invited for a Postdoctoral Research Scientist to join Dr Duncan Sparrow’s laboratory in the Department of Physiology, Anatomy & Genetics to work on a project focused on investigating the environmental causes of congenital heart. This post will be funded for up to 5 years by the British Heart Foundation. You must hold, or be near completion of, a PhD/DPhil in Molecular Biology, Biomedical Sciences, Genetics or a related science. You must have experience working with animal models of development and/or cardiovascular diseases, preferably experience with mice. You will have a high level of technical competence in developmental, molecular and/or cell biology methods and techniques, for example mouse embryology and histology, fluorescence microscopy, immunohistochemistry, in situ hybridisation, ability to communicate your findings in a scholarly manner in writing as well as orally in English. Must have, or be capable of obtaining, a Home Office PILB (or better) to conduct required procedures. The closing date for applications is midday on 23 October, with interviews likely to be held the week beginning 30 October
For further information, please contact Dr Duncan Sparrow (duncan.sparrow@dpag.ox.ac.uk)
To apply, go to: https://www.recruit.ox.ac.uk/pls/hrisliverecruit/erq_jobspec_version_4.jobspec?p_id=131117
George Daley is Dean of the Faculty of Medicine, Professor of Biological Chemistry and Molecular Pharmacology, and Caroline Shields Walker Professor of Medicine at Harvard Medical School. A former Howard Hughes Medical Institute Investigator and President of the International Society for Stem Cell Research (ISSCR) from 2007-2008, his lab works on the biology and clinical application of stem cells, with a particular focus on hematopoiesis. He was awarded the Public Service Award at the ISSCR 2017 meeting in Boston, where we caught up with him to discuss his move from the lab to the clinic and back again, his quest to derive human hematopoietic stem cells in vitro, and his advocacy for science in uncertain political times.
You’re here to receive the ISSCR Public Service Award, which recognises your role in the formation of the ISSCR guidelines for stem cell research. How did you come to be involved with these guidelines, and what does the award mean to you?
My involvement with the guidelines began around 2005, when we had been living through years of debate about embryonic stem cells (ESCs), and the US National Academy of Sciences (NAS) had established a set of professional guidelines for this work. Although these had some influence internationally, many in the ISSCR leadership felt that there were subtle but meaningful restraints in the US that were unique because of the pressured political environment at the time, and that if the issues were vetted more broadly, global practices might be a bit more liberal. So I appealed to the ISSCR leadership to convene a taskforce that would examine the NAS guidelines and cast them in a broader perspective for international audiences and constituents, and I ended up chairing this ISSCR guidelines taskforce. Ultimately, we endorsed the broad principles of the NAS guidelines but proposed subtle differences as well, for example around payment for egg donation: our language was less restrictive, and this let particular communities evolve their own responses. Subsequently, the ISSCR ethics and policy committee has written a white paper outlining the ethical frameworks on which payments for these kinds of donations can be ethically justified.
Then again, around 2007 it was clear that the issue of premature clinical translation of science and unproven interventions was becoming a real problem. I had been elected ISSCR president, and decided to convene and serve on the clinical translation guidelines taskforce – these deliberations produced a framework for thinking about the legitimate way to translate basic science, and about how to preserve prospects for innovation and flexibility on behalf of practitioners. In the years after the 2006 and 2008 guidelines, the field produced induced pluripotent stem cells (iPSCs) and, by 2014, people started wondering if the existing guidelines were out of date. So I acted as the board liaison to yet another taskforce chaired by Jonathan Kimmelman, which led to the 2016 guidelines. In the end, I’m accepting the Public Service Award on behalf of the many dozens of folks who worked on these guidelines with me for the last 11 years.
The ISSCR’s 2016 Guidelines for Stem Cell Research and Clinical Translation, available here
In such a fast-moving field, how do the ISSCR guidelines keep pace?
It was specified in the guidelines that they should be re-evaluated periodically – we didn’t specify how frequently, but in response to changing technology. What is challenging about working on those kinds of guidelines is that your goal is to articulate timeless principles – independence, peer review, autonomy of the individual, those kinds of things – so that you don’t necessarily have to tie the guidelines to particular technologies, which we know will evolve. When we wrote the 2006 guidelines, we anticipated various kinds of pluripotent stem cells, without exactly knowing that iPSCs would emerge. In the latest version, we knew CRISPR gene editing was possible, but since we can’t anticipate even more transformative future technologies, we framed a set of principles around the broader concerns of intervening in the germline, whether mitochondrial or nuclear. Like a constitution that is well written, the guidelines can provide principles that can be reinterpreted in different eras but, like the constitution of the US, they are supposed to be subject to amendment.
Let’s go back to the beginning: was there anything that got you into science in the first place?
I always wanted to be a scholar. I was a good student, I liked to read, to think, to solve problems. So I always thought I would end up in academia, but along the way I took a number of crazy turns. When I came to Harvard as an undergraduate, I was as interested in politics as science. I quickly became disillusioned by the anonymity of the large, introductory science courses, so I searched the course catalogue for the smallest class at Harvard and found a freshman seminar on spiders given by a professor and curator of arachnology at the Museum of Comparative Zoology that was limited to two people. I thought that I’d get a lot of attention there, but when I went to interview the professor said he’d already filled the class! It turned out that the two who had already signed up were identical twins! I argued to the professor that he’d never be able to tell them apart, so he should take me as a third; he thought this was quite funny, so let me in. In studying a particular class of animals I was introduced to taxonomy and the underlying principles of evolutionary biology. Meanwhile, I also became very interested in the philosophy of science and evolution, and initially declared my major in philosophy.
I got interested in laboratory science also quite by happenstance. As a freshman on financial aid, I had a work study job washing dishes in one of the dining halls. When I came back for the sophomore year, I figured I would find a more interesting job washing dishes, so this time I began washing glassware in a lab. Very quickly thereafter I ingratiated myself into the lab: once I’d done the beakers and test tubes I would be looking over the shoulders of the scientists to see what they were doing. That was a lot of fun, and I really loved the research angle – you were asking a question that no one else had ever answered. Research wasn’t just being told this is they way it is – you had to discover what the principles were.
How did you first get interested in stem cells?
I had a wonderful early supervisor and mentor, a postdoc named Beth Luna, who told me that if I was really interested in doing graduate work in biology, then I would need more intensive coursework, so I started taking more biology classes, and then ultimately switched my major from philosophy to biology. Then, the summer after my junior year, I decided to spend some time with a clinician just to see what being a doctor was like (initially more to prove to myself I didn’t really want to do it!). I spent time with a neurologist at Massachusetts General and the experience struck me as very compelling – the problems of human biology were deeply entrenched in medicine. This convinced me to apply to the MD PhD programme.
In grad school I ended up working on a problem that had been introduced to me in my hematology class: the molecular basis of chronic myeloid leukaemia (CML). I went to work for David Baltimore, the world authority on ABL, the gene that was disrupted in CML. That was really deep cancer and leukaemia biology, and I ended up making a mouse model of the disease that validated the BCR-ABL fusion gene as the driving molecular lesion in CML and as a target for treatment.
But even then I realised that there were a lot of leukaemias that didn’t have a defined molecular lesion, and that the broad platform for treating leukaemias – if one didn’t have targeted chemotherapy – was bone marrow transplantation. I was heavily influenced by a visiting professor from the UK, John Goldman, who was making his reputation expanding the use of unrelated donors for marrow transplants. We spent a lot of time thinking about hematopoietic stem cells (HSCs), the challenges of finding matched donors, and whether one could either generate a universal ʻoff-the-shelf’ donor cell, or an autologous cell that you could make from some non-leukaemic tissue. I was thinking about stem cell transplants when, once again by happenstance, Martin Evans, who had isolated mouse ESCs, was on sabbatical in the next lab. He introduced me to a paper from Rolf Kemler’s group describing the methodology of in vitrodifferentiation of ESCs to make embryoid bodies – one of the prominent differentiated tissues was blood. So even as a graduate student – and unbeknownst to David – I started culturing ESCs and seeing whether I could coax them to make blood. And I could, and this led to the notion that I could use ESCs as a platform to understand blood development.
But then I had to decide whether I was going stay in science or go back and finish my training in clinical medicine; because I had done work in a human leukaemia I decided to finish medical school and a medical residency and hematology/oncology fellowship. I was asked to spend an extra year as Chief Resident, which delayed my return to the lab but was an amazingly deep and wonderful experience. When I finally came back to the lab, my goal was to see if we could use ESCs as a platform for investigating the mechanisms of blood development. My scientific focus over the last 20-plus years has really been using mouse models in parallel with in vitro models comprising human and mouse ESCs and iPSCs to get at the core pathways driving blood development.
At the end of a recent paper you describe your ultimate goal in this line of research as ‘the derivation of bona fide transgene-free HSCs for applications in research and therapy’. How far are you from achieving that goal?
When I first laid out this goal in my proposal for becoming a Whitehead Fellow back in 1995, my hypothesis was that because pluripotent stem cells give rise to blood, they must transit through an HSC intermediate to get there; the dogma was that all blood cells arose from an HSC. That turned out to be wrong – the HSC isn’t there in the dish, unless you go through a different set of morphogen patterning to make a different type of mesoderm. That’s where developmental biology comes in – to inform directed differentiation in vitro.
The cells that we’ve made recently come from a two-step strategy: morphogen-directed differentiation gets us to what is probably the equivalent of lateral plate mesoderm, from which hemogenic endothelium and HSCs arise; then we direct cell fate specification with transcription factors in the manner of Yamanaka. The cells behave according to principles by which you define HSCs: they self-renew, engage multi-lineage engraftment over a long term (although not as long as a native cell), and they can do this on a single-cell basis. However, functionally we know our iPSC-derived HSCs are way below the standards and performance of freshly isolated human HSCs, and molecularly there are significant differences. I think we can continue to refine our strategy and get ever closer to a fully functional cell; then, the question becomes how native and how normal does it have to be? Does it need to be identical to a native HSC? This sort of goes back to philosophy again – it’s not really science!
Not only does developmental biology inform stem cells, but it also works the other way round
How important has developmental biology been for your research, and how do you see the relationship between developmental and stem cell biology?
I think all of stem cell biology is built upon the foundations of developmental biology. In the absence of the principles by which embryonic polarity is laid down, morphogenesis is induced and gastrulation is regulated, attempts at directed differentiation of pluripotent stem cells in a dish would be pure empiricism, pure tinkering, and not particularly intellectually satisfying. A large amount of directed differentiation is just that anyway, which is part of its well-justified criticism. But what’s so distinctive about stem cell biology is that you are observing every single step of the differentiation pathway – there are very few developmental systems that allow you that degree of direct observation and insight, especially when you get to more complex organisms. We work very closely with Len Zon, in part because the transparent zebrafish embryo allows direct visualisation and experimental manipulation of the earliest stages of commitment to both primitive and definitive hematopoiesis. That’s been enormously powerful, but we’ve had to then make the leap back to mouse and, crucially, to human; many of the principles are conserved, but the detailed mechanisms and specific gene sets might not be.
I’d like to make the point that not only does developmental biology inform stem cells, but it also works the other way round. In our recent work we started with a screen in vitro, taking iPSC-derived progenitors that make primitive forms of hematopoietic cells and asking whether we could identify epigenetic barriers to multipotency. Indeed, we identified a histone-modifying enzyme that broadly inhibits multipotency in embryonic blood precursors. Then, in order to really understand how this negative regulation works, we went back into a developmental system, the mouse. I think the real interest of this work is what it reveals about the regulation of the emergence of hematopoiesis in the embryo.
Back at the start of my lab, I saw an unmet medical need: there were, and still are, lots of patients who could benefit from bone marrow transplants but are never offered the chance, either because they didn’t have a suitable donor or due to the enormous toxicity of the transplant because of immunological dissimilarities. I have for many years thought that we could make a platform for combining gene therapy and cell replacement in a way that could open up a curative marrow transplant approach to dozens of diseases. But I always knew that clinical success would be a very high bar, and so what has really driven me, and the reason I’ve been able to sustain my lab for 20 years on the same model, is that it’s been a fabulous platform for learning about the pathways and mechanisms that contribute to blood formation in the embryo, and blood lineage development.
You were president of the ISSCR a decade ago. What have been the most significant changes in the field in that period, and where do you see the next ten years going?
When I was president, the revolution in reprogramming was just happening. It’s really interesting to see how that evolved: in the later part of the 1990s and early 2000s, many of us were thinking about reprogramming via nuclear transfer as a mechanism for generating customised or individualised patient-derived cells. There were even many who appreciated that genes such as Oct4 and Nanog were almost certainly involved in reprogramming. But Yamanaka’s great insight was that you could identify a manageable set of transcription factors that would change a cell’s fate, contrary to the expectation that reprogramming would be so complex that it could never just involve a small set of factors. There’s been an enormous amount of progress since then, and the number of disease models that have benefitted from reprogramming is staggering.
I think that over the last decade we’ve come to a much deeper understanding of the regulation of the genome, the mechanisms of gene regulation, but also chromatin organisation. Side by side with this, the technological advance that I think is so exciting and likely to dominate the next ten years is organoid biology. It’s just so exciting to see tissues forming because the information is preordained in the cells themselves, but perhaps not surprising given what we know about anatomy and historical experiments on tissue assembly after dissociation in animals such as sponges.
You have been a stem cell researcher and vocal science advocate since the years of George W. Bush. How did stem cell research cope when federal funding for human ESCs was barred?
It was discouraging and profoundly upsetting to be part of a scientific community that was seeing the limitless potential of a new field opening up and then to have it embroiled in a deeply divisive, fundamentally ideological debate around the nature of the human embryo and subject to a form of scientific censorship – the denial of federal funding. Science advocacy was not something I’d planned to get involved in, but the knowledge base that I had and the areas in which I was working made certain individuals in the political sphere seek me out for support and advocacy. I was a junior person at the time and several senior mentors were worried about the effect on my scientific career of being seen as too public; a lot of people told me to keep my head down, focus on the science. It was a struggle and I often wondered about this balance, and did turn down numerous opportunities to speak out more aggressively. But I think it really is a core responsibility of scientists to not simply be withdrawn behind their lab doors but to be willing to speak out on behalf of their science. It’s become obvious once again under the new administration that we have to stand up and justify the value of what we do in the context of lots of other competing priorities.
It really is a core responsibility of scientists … to be willing to speak out on behalf of their science
How do you see the prospects for state-funded research six months into the new administration?
The current President’s suggested budget included a roughly 20% cut in the NIH budget, and cuts for many other areas of discretionary scientific funding. Congress is unlikely to allow these cuts to stand, given that they have a broader base and interest in the importance of scientific research as a foundation not just for the future of human health but as an economic driver, a real creator of jobs, a way of priming the pump of biotechnology and the pharmaceutical industry. So I’m hopeful that the US’s traditional support for scientific research will continue. The other crucial element is that biotechnology is an area of international competitiveness, and the US is arguably the unchallenged leader. But without continued support, that leadership is subject to disintegration, and with the rise of China as a force in science, and the amount of investment they have committed to, international competition can be seen as an existential threat to the US. Trump is nothing if not hugely competitive, and this is an area where we certainly don’t want to lose out.
This year you started as Dean of Harvard Medical School (HMS). What do you hope to achieve during your tenure?
It’s an exhilarating and challenging new role. Before I took it up, I was Principal Investigator of a Howard Hughes-funded lab, a director of a small clinical division of bone marrow transplantation, a teacher of an undergraduate and a medical school course, a consultant to biotechnology and founder of a number of biotech companies. I had a very diverse professional life as it was, but I stayed within a fairly narrow realm of my own expertise. Becoming Dean has forced me to get up to speed with a tremendous array of different areas, both scientific and biomedical, and to some degree political. The Dean of HMS is seen as one of the prominent spokespersons for US medicine broadly defined, so I have to think about things like the economic aspects of healthcare delivery. HMS plays an enormous role internationally in global public health; we’ve got constituencies that are setting the pace in fundamental discovery, we have a community that is organised around translating biology into new medicines and new treatments, and we’re partners with biotech and pharma, which is increasingly concentrated in Boston. Clinical development and delivery of healthcare is an aspect that I now have to think about and influence. It has been exciting, and I hope that I can marshal my deep respect for science and my excitement and enthusiasm for clinical translation, together with a heightened sense of service as a clinician, to help with HMS’s fundamental mission to relieve the considerable suffering that disease inflicts on society.
In 2011 you were awarded the HMS A. Clifford Barger Excellence in Mentoring Award. Is there a ‘Daley’ style of mentorship, and who were your own important mentors in research?
I have to say that was one of my proudest awards, and in fact I was delighted as Dean to preside over this year’s awards just the past week.
I’ve had a phenomenal group of wonderful mentors who have taken an interest in me and helped me to develop throughout my career. I mentioned how important Beth Luna was very early on, and later there were people such as John Goldmann, Sam Lux and David Nathan, physicians who worked with me, and John Potts, who is in his eighties but who still meets and shares wisdom with me.
David Baltimore’s style of mentorship very much influenced the kind of mentor I want to be – the way he organised his lab and ran group meetings, demanding high standards and yet welcoming open candid critical debate about science. He emphasised the importance of the question and not just the technique, which fostered a sense of innovation: in chasing questions you marshalled whatever technology you needed, absorbing established techniques and developing new ones. Like David’s, the best scientific labs have a horizontal structure, so that everyone speaks equally and individually to assert a quest for the truth; that was very influential on me. So I have tried over the years to establish a climate in the lab where people are inspired to ask big questions, are empowered by adequate resources to chase those questions in a relatively unfettered way, and are open to collaboration. I also want to encourage a climate of having fun: we do a lot of social events and sing a lot of karaoke; if I can get up in front of a group, belt out a song and make an ass of myself, I feel it gives others a certain willingness to step forward and to take risks.
Finally: is there anything Development readers would be surprised to find out about you?
Outside of the lab, I’m passionate about cooking and collecting wine: I’m a foodie. It grew directly from travelling for science – going around Europe and being hosted by people who were eager to feed me their local fare and ply me with their local libation, whether wine or beer. My wife and I came together around cooking and we do a lot of that. One has to eat, so it’s a fairly efficient way of exploring new cuisines and experiences while still keeping your time commitments.
In the future diabetics might benefit from getting insulin-regulating beta cells transplanted into their body because their own beta cells are destroyed or less functional. However, according to new stem cell research at the University of Copenhagen, the current way of producing beta cells from stem cells has significant shortfalls. The beta cells produced have some features resembling alpha cells.
Beta cells release insulin in your blood, but when you suffer from Type 1 diabetes, you hardly have any of them left in your body. This is because the immune system attacks the beta cells.
The role of insulin is to reduce and regulate the blood sugar level when it is too high. People with diabetes do not have this function, and therefore need insulin injections in order to regulate their blood sugar levels.
Researchers are trying to produce beta cells artificially with the purpose of transplanting them to diabetic patients to regulate their blood sugar. A new research result from the University of Copenhagen and Novo Nordisk recently published in the scientific journal Stem Cell Reports provides a better understanding of how to improve the production of beta cells from human embryonic stem cells.
“At the moment, we can make stem cells develop into something that resembles proper beta cells. Our research shows that the current method produces cells that resemble alpha cells a little too much. However, the research has given us a better understanding of the steps stem cells go through when they develop into beta cells. In fact, we also show that the cells can develop along different paths, and still end up making the same type of beta cells,” says Anne Grapin-Botton, professor at the Novo Nordisk Foundation Center for Stem Cell Biology, DanStem.
Cells individually examined
The researchers have based their work on human pluripotent stem cells, which are able to evolve into any cell type in the body. Using known methods, the scientists analysed about 600 different cells on their path to beta cell differentiation and individually examined the cells to find out how much they molecularly resemble the beta cells.
In doing so, the researchers acquired important new knowledge about the way in which the cells develop and which genes play a role in this development. Notably, it was important that the genes NXK6.1 and MNX1 were activated for the cells to become beta cells in the end.
“This study takes an in-depth look at the molecular mechanisms on the cell level. We are not looking at what the average cells do, as other scientists have previously done – we are looking at all the individual cells. We are doing so in the hope that we can prevent cells from developing in the ‘wrong direction’. This work sheds light on the paths which the cells take in their development and how we human beings develop in the womb,” says Anne Grapin-Botton.
Start but do not complete the process
Alpha cells have the opposite function of beta cells. They must ensure that the body secretes the peptide hormone glucagon into the blood when the blood sugar level is too low. While the alpha cells cause the blood sugar level to rise, the beta cells ensure that it falls. And when the produced cells resemble the alpha cells too much, they are not optimal for treating diabetics.
“The cells definitely start the process of becoming either alpha or beta cells, but they don’t complete it. Here, we need to carry on researching to learn even more about how we can optimise the last step in the development of beta cells,” explains Anne Grapin-Botton.
The study was conducted in cooperation with project leader Christian Honoré from Novo Nordisk, and is supported by Innovation Fund Denmark, the Danish National Research Foundation and the Novo Nordisk Foundation.
Applications are invited for a Research Assistant position in the group of Prof Daniel St Johnston at the Gurdon Institute, University of Cambridge (http://www.gurdon.cam.ac.uk/research/stjohnston). The BBSRC-funded project aims to determine how epithelial cells organize apical-basal arrays of microtubules and how this is controlled by cortical polarity factors. Responsibilities will include establishing a method for the biochemical isolation of microtubule organizing protein complexes and their analysis by mass spectroscopy in collaboration with the Cambridge Centre for Proteomics and the generation of transgenic and mutant flies using CRISPR/Cas9.
Applicants must have a Bachelors or Masters level degree in a relevant area of Biology or equivalent experience. Expertise in protein purification, molecular biology and/or Drosophila transgenesis would be an advantage, although training can be provided where necessary. The post does not require a PhD qualification.
Our paper, like so many scientific findings, was brought about by a beer – or more specifically a discussion over a beer.
“I had a beer with David (Drechsel)” Jochen (Rink) said to me after one of our weekly scientific social events at the MPI-CBG. Over their beers they had discussed the challenges we were having imaging planaria and David had suggested we use Iodixanol as a supplement to increase the refractive index of mounting media – a suggestion which proved to be key to our success. While this particular problem and solution were quite straightforward, getting to the point where we could even recognize the problem and thus seek a solution involved a few more people (and a few more beers!) This is the story of how our paper, which describes a simple and straight forward method to correct for spherical aberrations of live tissues thus enabling significantly improved image resolution and quality in deeper tissue layers, came to be.
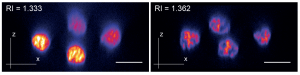
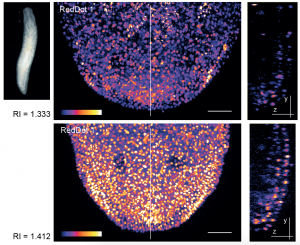
Tuning the refractive index of zebrafish cell culture media to RI 1.362 leads to improved signal to noise ratios along the z-axis. (Boothe et al, eLife 2017, https://elifesciences.org/articles/27240)
During my Ph.D. training in cell biology at the University of British Columbia in the laboratory of Jim (Johnson) I became used to thinking of microscopy as a tool rather than a challenge. This changed when I started my postdoctoral work, which focused on imaging cell dynamics during regeneration in planarian flatworms, in Jochen’s lab. Jochen warned me that this would “not be an easy” endeavor and, given how few tools were available for this relatively rare model system, I believed him. Once I started the imaging experiments and realized it was not possible to see any nuclear structure beyond the outermost cell layer however, I started to understand just how difficult the task I had taken on was. I took advantage of the lab’s species collection to obtain and test an unpigmented planarian species and exhausted the resources of our well-equipped light microscopy facility but the epithelium still seemed to act like a black-out curtain. At this point in time I did not have a well-developed understanding of optics in complex tissues however with the help of MPI-CBGs Moritz (Kreysing) and his student Alfonso (Garcia) I began to understand why a lack of pigment does not automatically mean that a tissue becomes optically clear. One of Moritz’ projects had seen a similar “black-out” effect when imaging dense retinal tissue and he had been able to overcome this challenge and image the deeper tissue layers by tuning the refractive index of their mounting media. While this was a bit different from what we were looking to do, as their work focused on fixed tissues, their results did suggest that the problem we were having could be caused by a significant difference in the refractive indexes of the planarian tissue and the aqueous mounting medium we were using at this time, and that if this were the case it might be possible to improve our images by tuning the refractive index of our mounting media. What we needed to do seemed clear however we still faced a challenge – while refractive index adjustment is a core component of state of the art clearing techniques in fixed tissues we had to find a component to tune the refractive index of our mounting medium to that of the sample without harming the live specimen.
I started by trying obvious candidates such as glycerol and sucrose but their high osmolality created a lethal environment for planaria. Halocarbonoils, which we tried next, worked well in fixed planaria but their hydrophobic nature made it impractical for an aquatic model system. After many trials and even more errors, I found myself at an institute social event sharing a beer and discussing these failures with Lennart (Hilbert). Lennart was studying DNA structure by live super-resolution microscopy in Nadine’s (Vastenhouw) lab at the time, and used BSA for refractive index tuning. While BSA was a promising candidate for planaria, the rather low refractive index tuning range and the viscosity and stickiness of saturated BSA solutions limited its usefulness. Lennart also found these limitations frustrating and so, while his approach at that time could not solve my problem, I did find myself a companion in the search for a live compatible refractive index tuning media supplement.
The postdocs had rounds of beers and discussions week after week but it was not until (of course) the PI got involved that we had our next breakthrough. At the time of Jochen’s forementioned beer with David we were focusing on epithelial cell dynamics because this was really the only cell type we could image in planaria due to the “black-out” effect. David, who was leading the MPI-CBG’s protein expression facility at the time, suggested over that famous beer we should give Iodixanol a try. Like many others he knew Iodixanol as a density gradient medium. While it was widely used for cell or cell organelle isolation, David was also aware though that the stock solution has a rather high refractive index which meant it might be able to meet our particular needs.
Refractive index tuning of planarian culture media by Iodixanol supplementation leads to a significant improvement of nuclei detection in deeper tissue layers. (Boothe et al, eLife 2017, https://elifesciences.org/articles/27240)
After performing some initial tests it became clear that Iodixanol was indeed the reagent we had been looking for. We were able to image past the first cell layer in planaria and significantly improved the live imaging quality in these specimens. Since we were not aware of any compound with similar properties we were eager to test it in other model systems. The diversity of MPI-CBG provided us with access to a number of different model systems and so we tried to improve live imaging in zebrafish embryos. Although this organism is thought to be easy to image, I knew of Lennart’s frustrations thanks to our earlier beers and was happy to share that there might be something which would help. Since imaging zebrafish was so far already of high quality Iodoxanol supplementation did not lead to the clear “day and night” effect we could observe with planaria. As I am a very self-critical (some would even call it pessimistic) experimentalist I started to doubt the broad applicability of Iodixanol but luckily Lennart has a more optimistic approach to things and just said: “Of course it works, it’s Physics.” – and sure enough it did.
In the end it was the diversity of the institute and the inter-lab interactions which led us to this story. We are happy that together we could establish with Iodixanol a compound which can now compensate for spherical aberrations in vivo and we are eager to hear how it works for the community – ideally over another case of beer!
 (No Ratings Yet)
(No Ratings Yet) (7 votes)
(7 votes) Mouse embryo that has been injected with embryonic stem cells grown in Insulin showing these cells can make all cell types. The embryos are at the stage when they are making all the different basic cell types of the future body, gastrulation. The stem cells were labelled genetically so they fluoresce in red under the microscope.
Mouse embryo that has been injected with embryonic stem cells grown in Insulin showing these cells can make all cell types. The embryos are at the stage when they are making all the different basic cell types of the future body, gastrulation. The stem cells were labelled genetically so they fluoresce in red under the microscope.