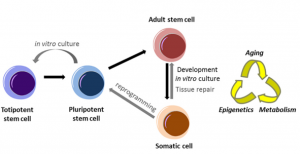
As a medical doctor specialising in respiratory diseases, with limited experience in real lab-based research, I was disappointed how little could be done for patients with end-stage lung disease (ESLD). Although respiratory diseases account for the third highest mortality of non-infectious disease deaths worldwide and lung cancer is the most common cancer (World Health Organization, 2011), treatment is largely supportive and lung tissue destruction is irreversible.
I started my PhD with Emma Rawlins at the Gurdon Institute, University of Cambridge, as part of the Wellcome PhD Programme for Clinicians by looking at human induced pluripotent stem cells (hiPSCs). These seemed to provide an opportunity to apply the discoveries of developmental biology to treat disease (Takahashi and Yamanaka, 2006). Differentiating patient-specific iPSCs into any relevant cell type could potentially be used for disease modelling and drug screening, and also bring us a step closer to cell-based therapies including personalised organogenesis. Research on murine lung development as a model for human lung development has been crucial to current protocols for differentiating human iPSCs into lung (Firth et al., 2014; Firth et al., 2015; Ghaedi et al., 2013; Huang et al., 2015; Huang et al., 2013; Longmire et al., 2012; Mou et al., 2012; Wong et al., 2012; Wong et al., 2015). I initially started work on iPSC directed differentiation to lung and wanted to use human embryonic lungs simply for validation purposes. However, it became evident very quickly that little developmental biology had been done on human embryonic lungs and it was not clear how closely the published step-wise iPSC differentiation protocols actually reflect human embryonic and foetal lung development.
Characterising the different progenitor populations in both the developing mouse and human embryonic lung seemed to me an essential part of regenerative medicine. The molecular understanding of developing human lungs was inadequate, as human lung development had not been characterised using cells derived from the human embryonic lung itself, but only using iPS-derived lung cells.
Mouse lung development
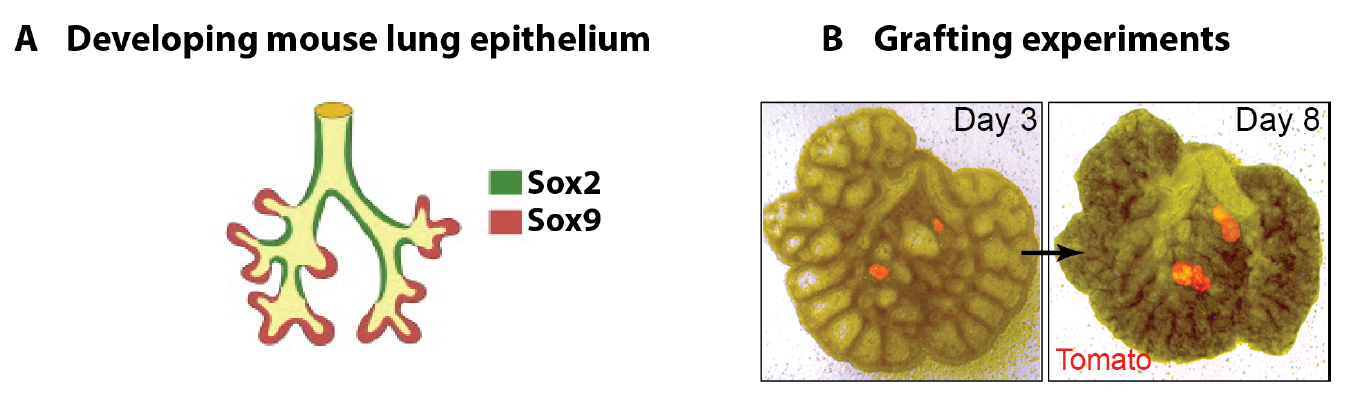
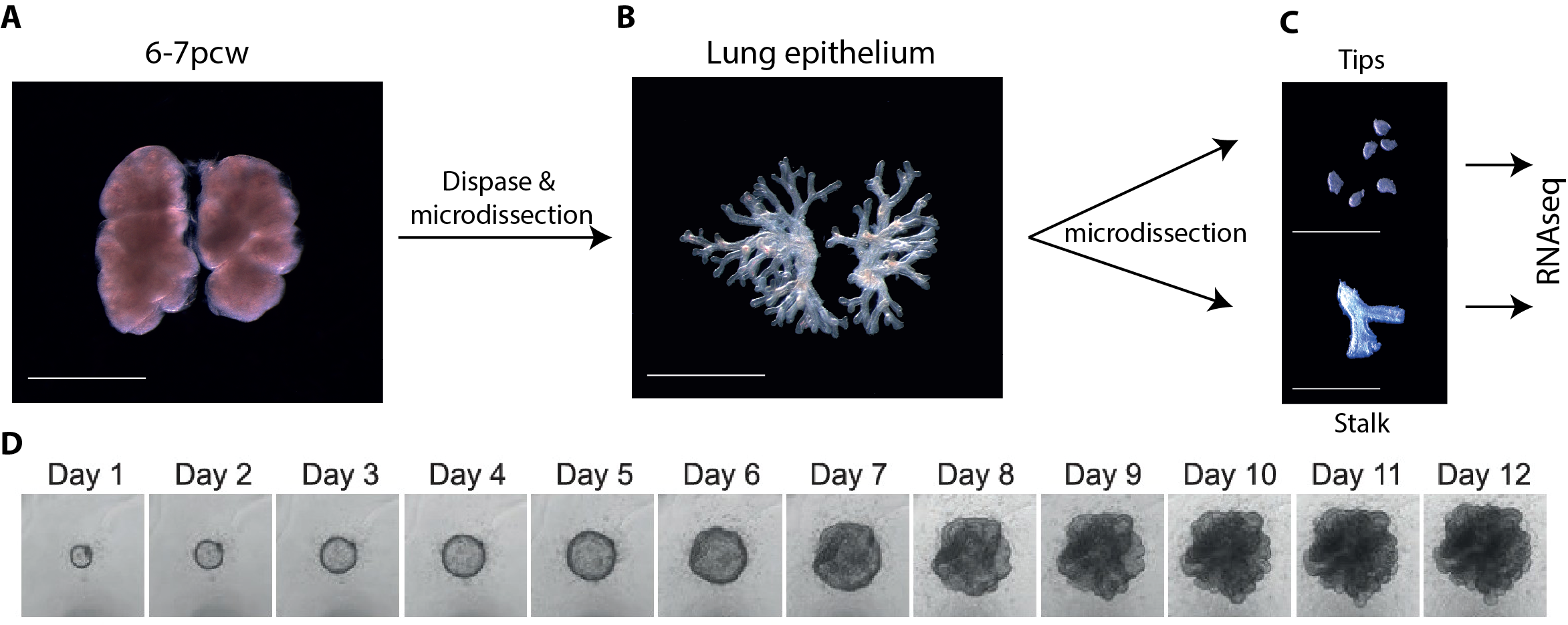
In parallel to my human work, I also worked on mouse lung development since it has been so extensively used as a model for human lung development. Considering the various anatomical and molecular differences already known between mouse and human lungs, I felt it was important to understand both mouse and human lung development more fully in order to ascertain to what extent the mouse findings could indeed be applied to human lungs. In both species, the lung develops as a blind-ending tube which branches multiple times, a process which results in the production of gas-delivering bronchioles and gas-exchanging alveoli. We know from lineage tracing experiments that the most important epithelial progenitor population in the developing mouse lung is found in the branching tips, giving rise to all the various epithelial cell types in the adult lung, both bronchiolar and alveolar cells (Rawlins et al., 2009). In mice, these epithelial tip cells are classically Sox9+ and initially give rise to the neighbouring Sox2+ stalk cells which are bronchiolar progenitors and give rise to the future airway (Fig.1 A). Using technically challenging grafting experiments I showed, among other things, that extrinsic signalling is the key in determining the tip and stalk progenitor cell fate, but that a significant degree of plasticity is present and that this gradually decreases throughout development (Fig.1 B) (Laresgoiti et al., 2016). I got a lot of practice in dissecting the epithelium free from any mesenchyme, which was very useful later on for the human work (Fig. 3B).
The finding that the environment is crucial in determining distal tip progenitor fate suggested that it would be possible to define in vitro culture conditions for human embryonic lung epithelium, both for the self-renewal of tip cells and their differentiation towards alveolar and bronchiolar lineages. This was an exciting realization as successful identification of culture conditions for developing human lungs would make molecular genetics possible.
Figure 1 A. Branching mouse lung is characterised by Sox9+ tips and Sox2+ stalks (reproduced from (Wang et al., 2013). B. Epithelial tip progenitors were microdissected from donor E12.5 (A) or E16.5 (B) Tomato (Rosa26RmTmG/+) lungs and grafted into the mesenchyme of unlabelled E12.5 hosts. Hosts/grafts were cultured for 8 days. Tip grafts grew (shown), integrated into the host lung and formed a lumen (not shown).
Main challenges of working on human embryonic lungs
One of the main limitations of working with human tissue is the lack of continuous tissue supply. This changed quickly and unexpectedly when I went to one of the Cambridge pubs to discuss my research plans and get some advice from a post-doctoral friend. He mentioned the “TransEuro” trial going on at the University of Cambridge – a clinical trial in which brain tissue from terminations of pregnancies is used for neurosurgical transplantation into treatment-resistant Parkinson’s patients. This meant that developing lung tissue was regularly discarded without any scientific benefit. This seemed to be the ideal source of human lung tissue for validation of my attempts at iPS-derived directed differentiation, but it quickly became clear what a huge potential an in-depth molecular characterisation of human embryonic lungs would have for the entire lung developmental and stem cell biology community. Additionally, the Joint MRC/Wellcome Human Developmental Biology Resource (www.HDBR.org) coordinated tissue harvesting from all over the UK via London and Newcastle. This meant that we had lung tissue provided to us multiple times per month from all over the country. The developmental age ranged from 5 to 20 weeks’ developmental age (7-22 weeks’ gestational age) and the samples used had no known genetic abnormalities.
Another challenge was that tissue used for sequencing and in vitro culture had to be as fresh as possible. Initially, we did not want to take any chances and dissection usually occurred as soon as possible after tissue was obtained, which was often late in the day. Experiments frequently continued until the early hours of the morning. I must say I was grateful once it became clear that overnight storage in specifically designed embryonic tissue storage medium did not affect gene expression, reproducibility and viability!
Since this was an exciting new area with huge potential, I was very grateful for the help I received from two amazing undergraduate/masters students, Oriol Caritg and Dawei Sun, who assisted me with my human work, as well as another PhD student, Jo Johnson.
Main achievements
Molecular comparison of human and mouse embryonic lungs
Molecular characterisation of human embryonic and foetal lungs using immunostaining, qRT-PCR and RNAseq transcriptome analysis revealed both major similarities and differences between mouse and human lung epithelial progenitors (Nikolić et al., 2017). Interestingly, 96% of orthologous genes that were expressed in human tips were also present in mouse.
The most important difference was the discovery that the classical stalk marker SOX2 is expressed in the human SOX9+ distal tip progenitors (Fig. 2), in stark contrast to the expression pattern in mice. We showed that this is a true interspecies difference and cannot be explained by relative age. This is an exciting finding, since Sox9 has been used as a classical mouse tip marker and Sox2 as a classical mouse stalk marker, with a clear demarcation between them (Chang et al., 2013; Liu and Hogan, 2002). My work suggests that SOX2 cannot be used as a unique stalk marker in human embryonic lungs and for validation during iPS-derived directed differentiation. Furthermore, there were more subtle differences in tip signalling pathways between mouse and human (Figure 3). The same signalling pathways were present, but these were wired in fundamentally different ways. For example, BMP2 and BMP7 were highly enriched in human tips, whereas Bmp4 was enriched in the mouse (Bellusci et al., 1996); IHH was expressed in human lungs whereas mouse lungs express Shh (Bellusci et al., 1997).
Figure 2. In the early stages of lung development, Sox2 is only expressed in the stalk in mouse (E11-15), whereas in human SOX2 expression extends into the tip (weeks 5-16).
On the one hand these data provide support for the use of mouse as a model for human lung development, but on the other hand they question its applicability in view of the many genetic differences, the functional consequences of which are not yet known. Therefore, to study the functional consequences of manipulating these genes, a genetically modifiable system for studying human lung development is required.
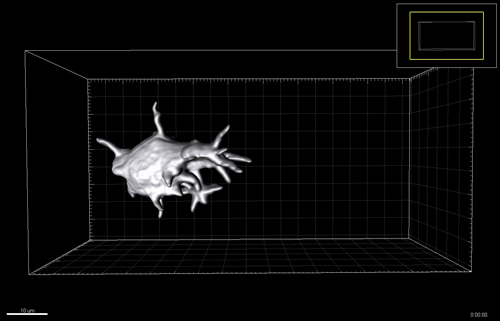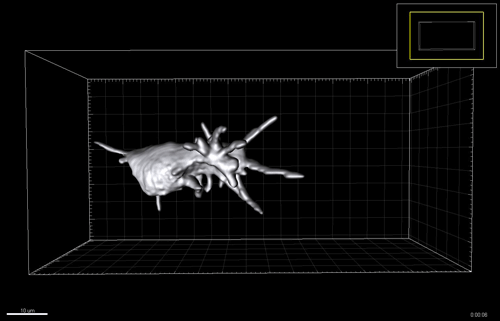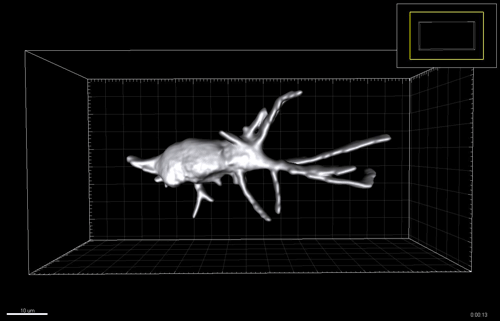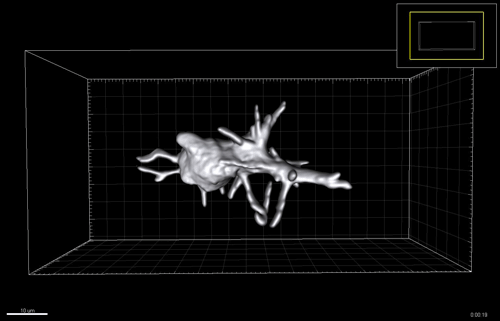
Figure 3 A-C. 6-7 post-conception week (pcw) human embryonic lung was incubated in dispase for 2 minutes, upon which the epithelium was microdissected free from any macroscopically obvious mesenchyme. The epithelium was further microdissected into tips and stalk, and then processed for RNAseq. D. Bright field images of one tip taken every 24 hours for 12 days cultured in self-renewing media (R-SPONDIN1, EGF, Noggin, FGF10, FGF7, CHIR99021, SB431542).
Self-renewing 3D organoid culture of human tip progenitors
We therefore developed culture conditions which have the ability to expand human embryonic lung epithelial stem cells as an indefinitely self-renewing, expanding population of undifferentiated cells with a organoid forming efficiency of 100% (Figure 3). The concept is similar to capturing the inner cells mass of a blastocyst as a self-renewing embryonic stem cell line. Our lung organoids could also be frozen and thawed, and the technical advice obtained from Meritxell Huch’s lab within the Gurdon Institute was immensely helpful. This now allows the in vitro study of human embryonic lung development without the need to obtain fresh tissue for every experiment.
However, in mouse, the same culture conditions were not sufficient to maintain mouse lung epithelial progenitor cells after several passages. This suggests that acquiring a self-renewing state is possible with further optimisation of the culture conditions, but the required signalling mechanisms are likely to be different. This was not surprising based on the transcriptome comparison between mouse and human tip progenitors.
RNAseq of cultured tips and stalks further validated my self-renewing culture conditions, since key genes are shared by cultured tip, cultured stalk and fresh tip cells without expression of differentiation markers. This suggests that the in vitro conditions mimic the in vivo conditions very closely.
Furthermore, single cell efficiency was high at about 35% and unpublished results showed that we were able to genetically manipulate human epithelial tip progenitors using CRISPR-Cas9. Knocking out SOX9 led to loss-of-self-renewing state phenotype, showing that SOX9 expression is essential for organoid maintenance. What this means is that we can now study human lung progenitor self-renewal in vitro in a genetically modifiable system which is extremely efficient without introducing chromosomal abnormalities, and which reproduces what is happening in vivo. This opens up a whole new research field of studying the mechanism of self-renewal and differentiation.
In vivo and in vitro differentiation as proof for a self-renewing state
One of the most important proofs that we had actually achieved a self-renewing state was the ability to differentiate our expanded organoid cells towards both bronchiolar and alveolar lineages. If real stem cells do it, then our expanded ones should do it do. Indeed, human embryonic epithelial lung progenitors, which had been expanded as 3D organoids, could be differentiated in vitro towards both alveolar and bronchiolar lineages. Similarly, xenotransplantation of organoid cells mixed with E13.5 dissociated mouse lungs, into the mouse kidney capsule led to efficient bronchiolar differentiation and the appearance of ciliated cells, mucus cells and basal cells, although alveolar differentiation was much less efficient. This strongly supported our hypothesis that a self-renewing state had been achieved.
We have also performed xenotransplantation into bleomycin-injured adult mouse lungs which showed highly efficient integration and proliferation of intratracheally administered single organoid cells. It also showed that the organoid cells can differentiate into bronchiolar cells, suggesting that lung identity is retained within the organoids. Such efficient engraftment has not been achieved previously. This is an exciting new model with many future possibilities.
Impact and future directions
In conclusion, the platform that we have developed for in vitro analysis of the embryonic human lung will be the first system available for functional experiments on human embryonic lungs. It will allow human embryonic lung development to be studied using modern molecular genetic techniques. The fact that we can genetically modify our organoid system also means that we can use CRISPR-Cas9 to identify novel genes required for alveolar and bronchiolar differentiation. This has been done in pancreatic development using iPS-derived cells (McGrath et al., 2015). However, our approach would be more reliable, as we would be doing this with real lung progenitors, albeit expanded ones, rather than iPS-derived ones. This research has the long-term potential to be transformative for lung regeneration by disease modelling, specifically by determining the role of genetic variants associated with lung diseases, by developing improved therapies for premature neonatal lung maturation, and also for in vitro differentiation of iPS cells. It has been claimed that iPSCs offer unparalleled opportunities to model lung development and disease using human cells. I would argue that our culture system using real human embryonic lung cells provides a more authentic and reliable platform to model lung development and disease.
From a clinical point of view, I hope that the work will make a difference to at least three groups of patients:
Patients with end-stage lung disease: our work will guide iPS-derived directed differentiation, which could then be used to recellularise a decellularised lung or lobar scaffold to be used for lung transplantation without the need for immunosuppression.
Premature neonates with lung failure due to immaturity: the 3D organoid culture system we developed will allow the study of late foetal development using real lung progenitors rather than iPS-derived ones. Understanding how the lung matures in these final stages of lung development will be crucial to improve survival for babies born prematurely.
Patients with rare congenital lung conditions. Gene editing in our 3D culture system will enable disease modelling of conditions such as surfactant protein B and C deficiencies.
Publishing first and fast versus publishing “big”
Every scientist would like their work to be recognised as important and groundbreaking by those in their field. However, as far as career progression is concerned, the journal chosen often appears to be far more important than the actual quality of the work. It seems to me that journal impact factor is used as a substitute for quality by those who are not experts in a particular field, or who do not understand the impact of the work. In my experience, important factors in choosing a journal include the speed of the review process, the availability of person-power to do the revision work, the awareness of competitors, and related guesswork of what stage those competitors have reached and whether their work is yet of a standard suitable for publication. During our paper revision period, I was in full-time clinical training and I was reliant on the goodwill of my colleagues for the revision experiments, especially Oriol Caritg and Quitz Jeng. On reviewing all these aspects, we chose a journal known for its efficient and fair review process rather than focusing simply on journal impact factor. Put differently, we chose to publish first and fast, rather than attempting to publish “big”. I remain hopeful that our work (as well as that of others) will be judged purely by its contents and the impact it has on the field, rather than by the impact factor of the journal in which it was published.
Marko Nikolić
Wellcome Trust/CRUK Gurdon Institute & Wellcome Trust/MRC Stem Cell Institute,
Liu, Y. and Hogan, B. L. M. (2002). Differential gene expression in the distal tip endoderm of the embryonic mouse lung. Gene Expr. Patterns2, 229–233.
Takahashi, K. and Yamanaka, S. (2006). Induction of pluripotent stem cells from mouse embryonic and adult fibroblast cultures by defined factors. Cell126, 663–676.
Our latest monthly trawl for developmental biology (and other cool) preprints. See last year’s introductory post for background, and let us know if we missed anything
It all went very meta this month with Matthew Cobb’s PeerJ preprint about a forgotten experiment in preprints from the 1960s. The story was highlighted in Science, a journal, Cobb explains, whose editor disparaged the initial preprint experiment in the 60s. Elsewhere, doubts were raised about last month’s headline-making human embryo CRISPR paper, as highlighted on the Niche (which also highlighted Shoukhrat Mitalipov’s response to the prepint).
“T47D_rep2 and b1913e6c1_51720e9cf were two Hi-C samples. They were born and processed at the same time, yet their fates were very different. The life of b1913e6c1_51720e9cf was simple and fruitful, while that of T47D_rep2 was full of accidents and sorrow…”
The preprints were hosted on bioRxiv, PeerJ and arXiv. Use these links to get to the section you want:
Resolution of Reprogramming Transition States by Single Cell RNA-Sequencing. Lin Guo, Xiaoshan Wang, Mingwei Gao, Lihui Lin, Junqi Kuang, Yuanbang Mai, Fang Wu, He Liu, Jiaqi Yang, Shilong Chu, Hong Song, Yujian Liu, Jiadong Liu, Jinyong Wang, Guangjin Pan, Andrew P. Hutchins, Jing Liu, Jiekai Chen, Duanqing Pei
Inhibition of granulocyte ROS production by opioids prevents regeneration. Elodie Labit, Lise Rabiller, Christophe Guissard, Mireille Andre, Christine Rampon, Corinne Barreau, Beatrice Cousin, Audrey Carriere, Margaux Raffin, Gilles Mithieux, Mohamad Ala Eddine, Bernard Pipy, Anne Lorsignol, Sophie Vriz, Cecile Dromard, Louis Casteilla
Tracking ectopic vertebra formation in injured zebrafish, from Lopez-Baez, et al.’s preprint
Hemimetabolous genomes reveal molecular basis of termite eusociality. Mark C. Harrison, Evelien Jongepier, Hugh M. Robertson, Nicolas Arning, Tristan Bitard-Feildel, Hsu Chao, Christopher P. Childers, Huyen Dinh, Harshavardhan Doddapaneni, Shannon Dugan, Johannes Gowin, Carolin Greiner, Yi Han, Haofu Hu, Daniel S. T. Hughes, Ann-Kathrin Huylmans, Carsten Kemena, Lukas P. M. Kremer, Sandra L. Lee, Alberto Lopez-Ezquerra, Ludovic Mallet, Jose M. Monroy-Kuhn, Annabell Moser, Shwetha C. Murali, Donna M. Muzny, Saria Otani, Maria-Dolors Piulachs, Monica Poelchau, Jiaxin Qu, Florentine Schaub, Ayako Wada-Katsumata, Kim C. Worley, Qiaolin Xie, Guillem Ylla, Michael Poulsen, Richard A. Gibbs, Coby Schal, Stephen Richards, Xavier Belles, Judith Korb, Erich Bornberg-Bauer
Structural and functional diversity of a dense sample of retinal ganglion cells. J. Alexander Bae, Shang Mu, Jinseop S. Kim, Nicholas L. Turner, Ignacio Tartavull, Nico Kemnitz, Chris S. Jordan, Alex D. Norton, William M. Silversmith, Rachel Prentki, Marissa Sorek, Celia David, Devon L. Jones, Doug Bland, Amy L. R. Sterling, Jungman Park, Kevin L. Briggman, H. Sebastian Seung, the EyeWirers
An improved ATAC-seq protocol reduces background and enables interrogation of frozen tissues. M. Ryan Corces, Alexandro E. Trevino, Emily G. Hamilton, Peyton G. Greenside, Nicholas A. Sinnott-Armstrong, Sam Vesuna, Ansuman T. Satpathy, Adam J. Rubin, Kathleen S. Montine, Beijing Wu, Arwa Kathiria, Seung Woo Cho, Maxwell R. Mumbach, Ava C. Carter, Maya Kasowski, Lisa A. Orloff, Viviana I. Risca, Anshul Kundaje, Paul A. Khavari, Thomas J. Montine, William J. Greenleaf, Howard Y. Chang
Active ribosome profiling with RiboLace. Massimiliano Clamer, Toma Tebaldi, Fabio Lauria, Paola Bernabo, Rodolfo F. Gomez-Biagi, Elena Perenthaler, Daniele Gubert, Laura Pasquardini, Graziano Guella, Ewout J. N. Groen, Thomas H. Gillingwater, Alessandro Quattrone, Gabriella Viero
A Large-Scale Binding and Functional Map of Human RNA Binding Proteins. Eric L Van Nostrand, Peter Freese, Gabriel A Pratt, Xiaofeng Wang, Xintao Wei, Steven M Blue, Daniel Dominguez, Neal A L Cody, Sara Olson, Balaji Sundararaman, Rui Xiao, Lijun Zhan, Cassandra Bazile, Louis Philip Benoit Bouvrette, Jiayu Chen, Michael O Duff, Keri Garcia, Chelsea Gelboin-Burkhart, Abigail Hochman, Nicole J Lambert, Hairi Li, Thai B Nguyen, Tsultrim Palden, Ines Rabano, Shashank Sathe, Rebecca Stanton, Ashley L Louie, Stefan Aigner, Julie Bergalet, Bing Zhou, Amanda Su, Ruth Wang, Brian A Yee, Xiang-Dong Fu, Eric Lecuyer, Christopher B Burge, Brenton Graveley, Gene W Yeo
SEA: The Small RNA Expression Atlas. Raza-Ur Rahman, Abdul Sattar, Maksims Fiosins, Abhivyakti Gautam, Daniel Sumner Magruder, Joern Bethune, Sumit Madan, Juliane Fluck, Stefan Bonn
Enabling cross-study analysis of RNA-Sequencing data. Qingguo Wang, Joshua Armenia, Chao Zhang, Alexander V Penson, Ed Reznik, Liguo Zhang, Thais Minet, Angelica Ochoa, Benjamin E Gross, Christine A Iacobuzio-Donahue, Doron Betel, Barry S Taylor, Jianjiong Gao, Nikolaus Schultz
A Data Citation Roadmap for Scientific Publishers. Helena Cousijn, Amye Kenall, Emma Ganley, Melissa Harrison,David Kernohan, Thomas Lemberger, Fiona Murphy, Patrick Polischuk, Simone Taylor, Maryann Martone, Timothy Clark
Bioinformatics Core Competencies for Undergraduate Life Sciences Education. Melissa A. Wilson Sayres, Charles Hauser, Michael Sierk, Srebrenka Robic, Anne G. Rosenwald, Todd M. Smith, Eric W. Triplett, Jason J. Williams, Elizabeth Dinsdale, William Morgan, James M. Burnette III, Sam S. Donovan, Jennifer C. Drew, Sarah C. R. Elgin, Edison R. Fowlks, Sebastian Galindo-Gonzalez, Anya L. Goodman, Neal F. Grandgenett, Carlos C. Goller, John Jungck, Jeffrey D. Newman, William R. Pearson, Elizabeth Ryder, Rafael Tosado-Acevedo, William Tapprich, Tammy C. Tobin, Arlín Toro-Martínez, Lonnie R. Welch, Robin Wright, David Ebenbach, Kimberly C. Olney, Mindy McWilliams, Mark A. Pauley
A position for a Postdoctoral Fellow is available in the Department of Development and Stem Cells, in the lab of Bill Keyes, IGBMC, Strasbourg, France (www.igbmc.fr/keyes). Previously, the group discovered roles for cellular senescence during embryonic development, and instructing cell plasticity and stemness in tissue regeneration and cancer. We are seeking a postdoctoral candidate to continue and develop these projects. As such, preference will be given to candidates with a proven record in developmental biology or with animal models of regeneration. These projects will involve the use of tissue culture and animal models coupled with high-throughput genomic analysis and molecular biology approaches.
For further information on the project and the work from the lab in general, see recent publications: Keyes et al, Cell Stem Cell, 2011, 8(2) 164-176; Doles et al, Genes & Development, 2012, 26(19): 2144-53; Storer et al, Cell, 2013, 155(5), 1119–1130; Ritschka et al, Genes & Development, 2017, 31(2):172-183
Candidate requirements
Ph.D or M.D./Ph.D. (already obtained or soon-to-be), with at least one first author publication.
Highly motivated person with strong interest in science research.
Interactive person, with ability to work independently.
Good communication skills, and fluency in English is required.
Previous experience in developmental biology will be heavily favored. Experience in molecular biology is desirable.
Work Environment
The candidate will join an international group of scientists, working on highly competitive topics. The candidate will benefit from access to a modern well-equipped laboratory, as well as access to the IGBMC’s renowned Core Facilities and support.
Conditions
Starting date: January 15th, 2018
Salary: salary is available for one year, but additional funding is sought, and the candidate will be supported to apply for competitive external fellowships.
Application procedure
Deadline: October 1st, 2017
All applications must include a CV with a letter describing your motivation and research interests (past and present), and the contact information for 2-3 referees.
Please submit your application to the following email addresses: bill.keyes@igbmc.fr
The Developmental Biology Unit studies the development of multicellular organisms. Research in the unit covers all levels, from the cellular to the whole organism, and is highly interdisciplinary, combining a wide range of approaches and innovative techniques. EMBL brings together the most talented biologists empowering them to explore bold new areas of biological inquiry.
Research in the Developmental Biology Unit is firmly embedded within the overall EMBL research environment, with extensive in-house collaborations and support from cutting-edge facilities, including genomics, transgenics, metabolomics, mass- spectrometry, and microscopy. Interdisciplinary research at the interface between development and metabolism, tissue mechanics, single-cell genomics, ecology, evolution, or theoretical biology is welcome, as are candidates using classical genetic models or new developmental model organisms.
The successful candidate should have a strong motivation to work in the multidisciplinary and collaborative environment of EMBL, grasping the opportunity to interact with many other research groups. In general, EMBL appoints group leaders early in their career and offers a highly collaborative and uniquely international culture. It fosters top quality, interdisciplinary research by promoting a vibrant environment consisting of independent research groups with access to outstanding graduate students and postdoctoral fellows. Significant core funding and limited teaching responsibilities allow EMBL group leaders to undertake ambitious, farsighted research programs and provides a scientific environment that is particularly attractive for researchers with families.
Further information about the position can be obtained from the Head of Unit Anne Ephrussi (anne.ephrussi@embl.de).
Further information about research in the Developmental Unit and at EMBL can be found at www.embl.org.
Application closing date: 8 October 2017. Interviews are planned for 18, 19 and 20 December 2017.
An initial contract of 5 years will be offered to the successful candidate. This is foreseen to be extended to a maximum of 9 years, subject to an external review.
APPLICATION INSTRUCTIONS
Please apply online through www.embl.org/jobs and include a cover letter, CV and a concise description of research interests and future research plans. Please also arrange for 3 letters of recommendation to be emailed to: references@embl.de at the latest by 1 October 2017.
ADDITIONAL INFORMATION
EMBL is an inclusive, equal opportunity employer offering attractive conditions and benefits appropriate to an international research organisation with a very collegial and family friendly working environment.
The remuneration package comprise a competitive salary, a comprehensive pension scheme, medical, educational and other social benefits, as well as financial support for relocation and installation, including your family, and the availability of an excellent child care facility on campus.
EMBL is committed to achieving gender balance and strongly encourages applications from all under-represented groups. Appointment will be based on merit alone. Further details on Group Leader appointments can be found under www.embl.org/gl_faq.
The European Molecular Biology Laboratory (EMBL) is one of the highest ranked scientific research organisations in the world. The Headquarters Laboratory is located in Heidelberg (Germany), with additional sites in Barcelona (Spain), Grenoble (France), Hamburg (Germany), Hinxton near Cambridge (UK) and Rome (Italy).
The Zaidel-Bar group studies Actomyosin Contractility and Tissue Morphogenesis from single proteins to the organism level.
After seven successful years at the Mechanobiology Institute in Singapore the lab is moving to Tel-Aviv university in October 2017. This is an exciting opportunity for highly motivated young scientists to join as MSc, PhD or Postdoctoral fellows.
If you are interested in working on a project related to the regulation of actomyosin during cell and tissue morphogenesis (http://celladhesionlab.com/) send a letter of motivation and your CV to zaidelbar@gmail.com.
Meeting Report: Congress of the International Society for Developmental Biology (ISDB 2017)
Singapore, the Lion City, attracted hundreds of developmental biologists from around the world to attend the 18th Congress of the International Society for Developmental Biology (ISDB 2017), June 18-22, at the National University of Singapore (NUS). The meeting takes place every 4 years, and gathers those fascinated by the early steps in life. I had the intention to attend one of the ISDB meetings since my early PhD months, but for different reasons it ended up not being possible – this year (already as a post-doc), I managed to join the international community in ‘the Garden of Asia’, and really enjoyed a vibrant and diverse program.
The opening ceremony kicked-off to welcome everyone with the traditional Lion dance. The main local host (and Singapore enthusiast) Phil Ingham, together with other members of the vibrant scientific community in Singapore, put together a phenomenal program that included 6 Nobel laureate lectures and many talks covering a variety of topics and (model) organisms. Based on my notes from the meeting, I will give an overview of the diverse sessions and highlight some talks – thus, my own personal bias to select certain topics will likely emerge.
The D’Arcy Thompson Symposium
The first day included the D’Arcy Thompson Symposium, celebrating 100 years since the publication of the landmark ‘On Growth and Form’ (a topic that was already highlighted in previous posts on the Node). The symposium, naturally, focused on physical and quantitative approaches in developmental biology. It was a great way to start the meeting, reminding us all about the benefits of talking to scientists in other field and join efforts to address important questions from different angles. Speakers included James Briscoe (The Francis Crick Institute, London, UK), employing experimental and computational modelling of transcription networks in neural tube cell fate specification; Thomas Gregor (Princeton University, USA), visualizing long-range promoter-enhancer interactions and transcription regulation in the fly embryo; Hiroshi Hamada (RIKEN CDB, Japan), addressing symmetry breaking in the mouse embryo; Tim Saunders (Singapore), exploring the zebrafish myotome as a model in addition to his work in the fly embryo (and who co-edited, with Phil Ingham, a special issue of MoD that also celebrated the topics covered in the symposium); Olivier Hammant (ENS Lyon, France), exploring the effects of forces and tension on tissue shape; and Carl-Philip Heisenberg (IST, Austria), who also focuses on how forces are generated and propagated in tissues using zebrafish as a model.
Nobel Lectures
We were all lucky to have a stellar selection of speakers and participants in all sessions of the meeting, and the organizers could additionally go the extra mile and gather 6 Nobel laureates all under the same roof (an organizational achievement in itself, as we know agendas do not often allow it). Unfortunately, we learned the one of the lectures had to be cancelled due to health issues (Sydney Brenner), but all other laureates gave fantastic talks: Christiane Nüsslein-Volhard (from fly and zebrafish early development to skin colours), Eric Betzig (beautiful movies of unmatched resolution that will surely inspire generations of developmental biologists), John Gurdon (from somatic cell nuclear transfer to cellular memory and histone modifications), Martin Chalfie (Hox genes as guarantors of cell fate specification in C. elegans), and Eric Wieschaus (Interpretation of the bicoid gradient and the role of chromatin accessibility).
On a personal account, as my current projects lie at the intersection of developmental biology and chromatin regulation, I have particularly enjoyed two of the talks that focused on these aspects. Sir John Gurdon, always with an inspiring clarity, showed us recent data about resistance to reprogramming by oocyte factors, not only identifying genes resistant to transcriptional reactivation but also by using chromatin modifiers to erase several histone modifications. Eric Wieschaus, in a talk that I have particularly enjoyed, explored how the maternal bicoid gradient is differentially interpreted by the genome of cells in different regions of the embryo. One of the key aspects is the regulation of chromatin accessibility, and one of the conclusions is that chromatin structure, rather than the underlying sequence, is what determines the concentration-dependent cohort of bicoid enhancers, with different degrees of affinity along the embryonic anterior-posterior axis. It was nice to hear another example of how chromatin remodelling is crucial during early development.
The Ross Harrison Medal Lecture
The prize is awarded every four years at the ISDB and is named after the American biologist who pioneered experimental embryology, and ‘applied tissue culture techniques to the study of embryonic tissue development and axon outgrowth’. The prize was awarded this year to Claudio Stern (UCL, London), who gave a talk on the fascinating links between twinning and the pathways that regulate asymmetry. With a well-balanced mix between the classical experiments that are always inspiring, and modern studies on human genetics, Claudio covered a range of the research in his lab over several years. I found it particularly interesting to see how topics connected from the early days to the current times, with very curious aspects that are more than coincidence!…
In general, the meeting had a wonderful mix of topics as well as model systems. From well-established model organisms to exquisite animals that are now becoming more amenable to use experimentally, of course due to the potentials of genome editing brought by the CRISPR/Cas9 technologies. In particular, I enjoyed the work from Antonia Monteiro’s lab (Singapore) on the evo-devo aspects of Butterfly eyespot formation and wing patterning; an amazing variety of fly species and other diptera resulted in a phenomenal talk by Claude Desplan (NYU) on the visual system and its evolution. It was also cool to hear about using Emu embryos as models (Craig Smith, Australia), or even about turtle shell development (Shigeru Kuratani, Japan)!
To highlight a few more speakers, I was marvelled with the identification of multinucleated cells during regeneration in the zebrafish heart (Ken Poss, Duke University, USA), which are temporarily generated and migrate to cover the wound. Norbert Perrimon (Harvard) explored inter-organ communication, giving several examples of how tissues send signals to each other within the organism, making us all aware that the processes we may be interfering with can have impact at distant target organs or tissues. François Schweisguth (Institut Pasteur, Paris) employed experimental and modelling approaches to beautifully tackle a long-lasting question about patterning: how stereotyped sensory organ patterns are self-organized (through Notch dynamics) in the developing fly notum. Finally, Denis Duboule (EPFL, Switzerland) gave an overview of limb development from the point of view of chromatin organization and genome topology. Given my own interests, I found it fascinating as work from his lab could uncover how enhancer regions may activate certain Hox genes in the forelimb or the digits, starting to reveal how development is orchestrated in connection with the structural organization of the genome in topological-associated domains (TADs).
There would certainly be more to tell, and I have really enjoyed the opportunity to meet some of my scientific heroes and openly discuss science with old colleagues and meet new students/post-docs from all over the world! It was also great for me to meet the Portuguese community (and make me feel at home even half a world away), who were also representing the Portuguese Developmental Biology Society (SPBD).
I can only look forward to the next ISDB, in 4 years’ time…! We already know it will take place in Vancouver, Canada (Summer 2021), and it will surely be a calendar highlight for all of us developmental biologists! Also, a new ISDB President will be in charge over the next 4 years up to the meeting – Angela Nieto (Alicante, Spain) is the newly elected ISDB President. Angela has committed to increase the interactions among developmental biologists around the world, as well as making funding agencies and the lay public aware of the importance of basic research in these fields to further the understanding of mechanisms that will be relevant other fields. This is certainly a point of view that I also share, so we can all hope Angela has a successful term!
To promote Developmental Biology and bring across the importance of fundamental research in this field, we all should have our elevator pitches ready at all times – whenever there is an opportunity to engage with audiences about our discipline – be it with the public, students, fellow scientists or policy makers. This is a first attempt at providing a concise rationale and ideas that can be woven into such conversations. The arguments presented can and must still be improved and complemented. Therefore, we would like to invite you to send in your thoughts and ideas, potential corrections, suggestions for improvement, additions, new arguments and/or potential links to supporting resources – all with the aim of further strengthening the message that we need to bring across (please, send to Andreas.Prokop@manchester.ac.uk). In particular those working in evo-devo or plant development, please come forward and make your contributions by helping us explain the importance of your subjects and extend this resource for the benefit of all.
If you would like to use some of the ideas currently presented, please download PowerPoint slides with the advocacy information here. If you are already active in science communication or plan to do so, please, also have a look at our public engagement outreach collection which can be found on the BSDB site and in revised form on The Node, at a special issue on science communication in “Seminars in Cell & Developmental Biology” (Box 5) and a recent PLoS Blog post.
If you are a Developmental Biology society, lab, researcher or aficionado and have a website: are you actively advocating the importance of our discipline? Feel free to copy out or use ideas and materials presented here. Vice versa, please share your links, your improved, complementary or additional arguments, and your advocacy strategies and resources. We aim to improve advocacy strategies across the globe and form a powerful network communicating the importance of Developmental Biology!
Why should we engage in Developmental Biology?
Developmental Biology enquires about the fundamental processes that underpin the fertilisation of an egg cell and its step-by-step transformation into the fascinating complexity of a whole organism (Box 1).
Box 1:
Some definitions of Developmental Biology
Developmental Biology is the study of the processes by which organs grow and develop. Modern developmental biology studies the genetic control of cell growth, differentiation and morphogenesis, which is the process that gives rise to tissues, organs and anatomy, but also regeneration and ageing (after L. Wolpert)
Developmental biology is the study of the process by which animals and plants grow and develop, and is synonymous with ontogeny (Wikipedia).
Developmental Biology is the causal analysis of the cellular mechanisms that drive processes of growth, pattern formation and morphogenesis (A. Martínez Arias)
Further advocacy articles
Development special advocacy articles
Maartens, A., Prokop, A., Brown, K., Pourquié, O. (2018). Advocating developmental biology (Editorial). Development 145 — [LINK]
Ebisuya, M., Briscoe, J. (2018). What does time mean in development? Development 145 — [LINK]
Wiese, K. E., Nusse, R., van Amerongen, R. (2018). Wnt signalling: conquering complexity. Development 145 — [LINK]
Philip W. Ingham. (2018). From Drosophila segmentation to human cancer therapy. Development 145 — [LINK]
Prokop, A. (2018). What is Developmental Biology – and why is it important? Open Access Govern, in press — [LINK]
Duronio, R. J., O’Farrell, P. H., Sluder, G., Su, T. T. (2017). Sophisticated lessons from simple organisms: appreciating the value of curiosity-driven research. Disease Models & Mechanisms 10, 1381-1389 — [LINK] – comment by A Martínez Arias [LINK2]
Gilbert, S. F. (2017). Developmental biology, the stem cell of biological disciplines. PLoS Biology 15, e2003691 — [LINK] – comment by A Martínez Arias [LINK2]
Zhang, F. (2017). A new age of discovery for biology. In “The World in 2018”. The Economist — [LINK]
St Johnston, D. (2015). The renaissance of Developmental Biology. PLoS Biol 13, e1002149 — [LINK]
Wassarman, P. M. (2016) Essays on Developmental Biology. Current Topics in Developmental Biology — [PartA] [PartB]
Hines, P. J., Marx, J., Parks, S. (1994). Frontiers in development. Science 266, 561-564 — [LINK]
Crick, F. (1977). Developmental biology. In “The encyclopedia of ignorance – life sciences” (R. Duncan, M. Weston-Smith, Eds.), Vol. 2, pp. 299-303. Pergamon Press, Oxford, New York, Toronto, Sydney, Paris, Frankfurt — [LINK]
At first sight, Developmental Biology could be viewed as an academic discipline driven by mere curiosity and, hence, to be of little relevance to the big challenges of population health or sustainability. On the contrary, Developmental Biology – along with Physiology[1] – is arguably the most important biological discipline we have. Here we will explain and substantiate this statement[2].
(1)Developmental defects in humans are very abundant (Box 2). By studying the underlying mechanisms and causes, Developmental Biology addresses the key challenge of population health. Sustaining food resources is another major global challenge, and Developmental Biology can provide key strategies to improving crop and plant cultivation (see Mathan et al., 2016, Development 143, 3283ff. — LINK; further arguments will follow – please help us by contributing your ideas!).
Box 2: Statements from the literature illustrating the abundance of developmental defects in humans
The frequency at which all classes of developmental defects occur is thought to be … exceeding half of initial pregnancies.
Major developmental defects … occur in approximately 3% of live births.
In 1995, major developmental defects accounted for approximately 70% of neonatal deaths (occurring before 1 month of age) and 22% of the 6,500 infant deaths (before 15 months of age) in the US.
Approximately 30% of admissions to pediatric hospitals are for health problems associated with such defects.
source:Scientific Frontiers in Developmental Toxicology and Risk Assessment, 2000, National Academic Press, Washington DC, pp.354; edited by the National Research Council (LINK)
(2) Developmental Biology (like Physiology) is asking fundamental questions at the level of whole organisms, organs or tissues (Box 3). Notably, this is the level at which diseases become manifest. For this reason, Developmental Biology has been, and continues to be, most effective in delivering explanations for diseases or medically relevant processes including infertility, neonatal death, birth defects (e.g. deformation, body growth abnormalities, developmental brain disorders, blindness, deafness), cancer, wound healing, tissue regeneration (regenerative medicine including stem cell biology), etc.
Box 3: Fundamental questions asked by developmental biologists – and how they translate into biomedical application
What processes lead to fertilisation and the initiation of development? How can we overcome infertility and childlessness?
How do single fertilised egg cells, or later on groups of progenitor cells, generate the enormous cellular diversity of an organism and its organs and tissues? How do stem/progenitor cells generate whole tissues or organs – for example in regeneration or tissue engineering, and how does wound healing work?
How do cells, which originate from common ancestors and contain the same genetic information, adopt different fates? How do cells change their identities and behaviours – for example in cancer?
How do tissues and their cells know when to stop growing? How can cells evade growth control – for example in tumour growth?
How is the formation of different cells/tissues coordinated in space and time? What are the patho-mechanisms underlying birth disorders?
(3) By asking fundamental questions at the level of organisms, organs and tissues, Developmental Biology-related research is a generator of new ideas and concepts (Box 4). These concepts essentially underpin the modern biomedical sciences and include cell signalling, tissue and body patterning, growth regulation, cell migration or morphogenesis; they form the basis for contemporary research into stem cells, cancer, wound healing, regeneration or ageing.
(4) Developmental Biology is exciting and powerful because it reaches across the different levels of biological complexity and explanation; phenomena at the level of organisms, organs or tissues can ultimately be understood only by tracing them back to events at the level of genes and cells. Consequently, Developmental Biology embraces disciplines such as genetics, molecular biology, (stem) cell biology, biochemistry, biophysics as well as evolutionary biology.
(5) Developmental Biology capitalises on the principle of evolutionary conservation of genes, mechanisms and concepts by making informed and strategic uses of suitable model organisms, down to experimentally and genetically amenable invertebrates. The use of invertebrate model organisms provides an efficient and powerful strategy to generate new ideas, concepts and understanding which can then be tested (and often validated) in higher organisms, eventually leading to medical applications in humans (LINK). This discovery pipeline has been a central driver for the enormous contributions that Developmental Biology has made and continues to make to the biomedical sciences.
Box 4: A metaphor explaining how Developmental Biology works
Understanding a combustion engine requires investigating its single parts, such as the sparking plug, cylinder or crank shaft. However, for a developmental biologist it is not sufficient to find out how each of these parts works, but this new knowledge needs to be linked back into the mechanistic framework that constitutes the entire engine and explains its function. Linking detailed findings back to the bigger question of understanding the engine is an important validation and filter step that reveals whether the detailed findings are actually relevant and make deeper sense. Only through such systematic and holistic “reverse enginieering” can the necessary systemic and conceptual understanding be achieved which is required to diagnose and eventually repair a faulty engine.
By executing all research work with the ultimate aim of understanding the bigger question (e.g. how to make an organ, appendix or entire body), Developmental Biology has become such an important contributor of concepts and understanding in the field of the biomedical and medical sciences.
As an interesting side note: already in the late 18th century, Johann Wolfgang von Goethe understood that it is not enough to “have the pieces in your hand” but that they need to be connected and “woven” together to “recognise and describe the living”.
These are only some thoughts (still incomplete and in need of further optimisation) about the importance of Developmental Biology as a discipline. If we want to improve the standing of our discipline, we MUST embrace these ideas with passion, help to complement and further improve them and, most importantly, use them whenever possible and adequate to advocate our discipline with self-confidence and enthusiasm. To do this with impact, we need to acknowledge that communicating science is a difficult task which requires belief, love for our subject, stamina, and efficient strategies that enable us to engage with a wide spectrum of target audiences. Such target audiences include the general public and schools, fellow scientists and clinicians, as well as politicians and other decision makers. But we need to be realistic and accept that most members of these target audiences will, by default, show little interest to engage with us. Therefore, intelligent and strategic long-term approaches – ideally shared within networks of scientists – likely provide the most promising path of engagement – as is explained in greater detail in the editorial of a special issue on science communication (published in Sem Cell Dev Biol; Box 5). For those who take an interest, this special issue provides many ideas of how long-term strategies for the communication of biomedical research can be implemented successfully – and can then even become rewarding for our own science and career. Please, also have a look at our link list of public engagement outreach resources which will provide you with further ideas and useful support materials.
Box 5:
Science communication in the field of fundamental biomedical research
The aim of this special issue is to provide concise and accessible advice on how to engage effectively, as well as share valuable practice gained from successful long-term science communication initiatives – thus providing ideas to all those who want to participate in dialogue with the public, policy makers or other scientists, or are participating already. This issue is primarily written for scientists working in the field of the biomedical sciences (and beyond), but will hopefully also be seen as a helpful resource for academic & professional science communicators. To appeal to both groups and hopefully stimulate impact-enhancing interdisciplinary collaborations, the issue is an unprecedented experiment written at the interface of both disciplines. The enormous opportunities of long-term approaches and the formation of interdisciplinary science communication networks are explained in great detail in our editorial. Further explanations about this issue were given in the recent NCCPE and PLoS|BLOGS posts.
———————————–
Cover art by Matt Girling. See figure legend in the editorial.
———————————–
1. EDITORIAL: Science communication in the field of fundamental biomedical research (Sam Illingworth, Andreas Prokop) – This article provides a critical assessment of the current state of science communication in the field of fundamental biomedical science, including lists of existing science communication initiatives, as well as a conceptual overview of the articles in the special issue [PDF] [see also].
2. Delivering effective science communication: advice from a professional science communicator (Sam Illingworth) – From his experience as Senior lecturer in Science communication, Sam Illingworth writes about important considerations for all who want to participate in active science communication. His tips and tricks for science fairs, school visits or many other forms of dialogue are provided in plain language. [PDF] [YouTube]
3. The nuts and bolts of evaluating science outreach (Suzanne Spicer) – From her experience as specialist for science communication at the Manchester Museum, Suzanne Spicer writes about the importance and practical implementation of project/event evaluation as an important means to improve quality and demonstrate impact. [PDF]
4. EuroStemCell: A European infrastructure for communication and engagement with stem cell research (Jan Barfoot, Kate Doherty, C. Clare Blackburn) – EuroStemCell is the flagship of non-commercial science communication in the area of the biomedical sciences, centred around their website with over a million annual visitors. The initiative was born out of an EU grant consortium and has developed into an interdisciplinary network that promotes responsible dialogue about stem cells, also through developing outreach resource and providing training. [PDF]
5. The Manchester Fly Facility: Implementing an objective-driven long-term science communication initiative (Sanjai Patel, Andreas Prokop) – The Manchester Fly Facility is an excellent example of an initiative communicating the importance of model organism biology. They promote Drosophila research through improved research training, school projects, science fair programs, and the development of publicly shared resources and strategies. Furthermore, they address Drosophila researchers to stimulate wider participation in science communication. [PDF]
6. Building dialogues between clinical and biomedical research through cross-species collaborations (Hsiao-Tuan Chao, Lucy Liu, Hugo J. Bellen) – Clinicians and biologists do not necessarily share a common scientific culture and way of thinking, but they share the common scientific goal of understanding and treating diseases. To this end, they can complement each other in powerful ways. This article by the Bellen lab provides helpful tools and successful strategies towards interdisciplinary collaboration between biologists working on model organisms and clinicians. [PDF]
7. DrosAfrica: Establishing a Drosophila community in Africa (Maria Dolores Martín-Bermudo, Luka Gebel, Isabel M. Palacios, I. M.) – DrosAfrica is an excellent example of how cost-effective, yet cutting edge research – as it is possible with Drosophila – can be used to free resources and promote active science and science education in disadvantaged countries. This article describes the origins of the DrosAfrica initiative and provides detailed insights into how their activities are organised. [PDF] [YouTube]
8. Engaging with primary schools: supporting the delivery of the new curriculum in evolution and inheritance (Paula Kover, Emily Hogge) – The “Teaching Evolution for Primary Children” initiative is an excellent example of successful dialogue with schools. The key strategy explained here is to research curricular and teacher requirements first, to then decide on suitable contents and develop useful resources in close collaboration with schools. This ensures influence of scientists on the correctness and quality of content and the influence of teachers on the way of presentation – in sum an excellent way to engage audiences at young age with science. [PDF]
9. The droso4schools project: long-term scientist-teacher collaborations to promote science communication and education in schools (Sanjai Patel, Sophie DeMaine, Joshua Heafield, Lynne Bianchi, Andreas Prokop) – Analogous to the previous article, the droso4schools initiative by the Manchester Fly Facility seeks true dialogue with teachers in that students are placed as teaching assistants in schools to learn about teaching strategies and teacher requirements. These experiences are then turned into sample lessons that use Drosophila as a teaching tool to explain curriculum-relevant concepts of biology, spiced up with micro experiments that are possible with flies even in classrooms. [PDF]
10. Science Communication at Scientific Societies (Jeanne Braha) – As former member of the AAAS science communication team, Jeanne Braha explains how societies can help their communities to improve science communication through offering advice, providing resources and training, connecting scientists to audiences, making information available to the public, and awarding prizes for outstanding science communication work. [PDF]
11. The Node and beyond – using social media in cell and developmental biology (Catarina Vicente, Aidan Maartens, Katherine Brown) – This article describes The Node as a unique communication platform for biomedical researchers, in particular from the fields of Cell and Developmental Biology. The Node does not only collate event and advocacy information, publish job vacancies and report about science-relevant topics, but it is also a blog site with international outreach where members of the science community can share news, thoughts or experiences. As such it acts as a modern electronic newsletter. [PDF]
————————–
[1] In the sense used here, Physiology comprises disciplines like immunology or functional studies in the field of neurobiology
[2] Arguments and examples given so far concern studies of animal development, and those for plant development will follow soon
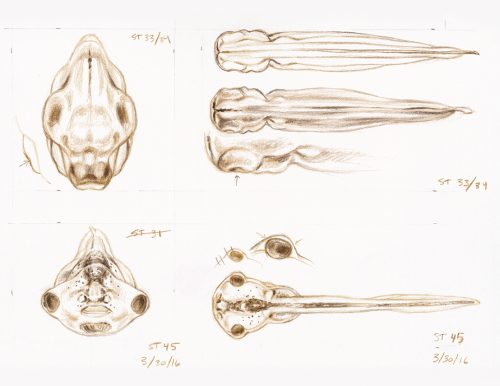
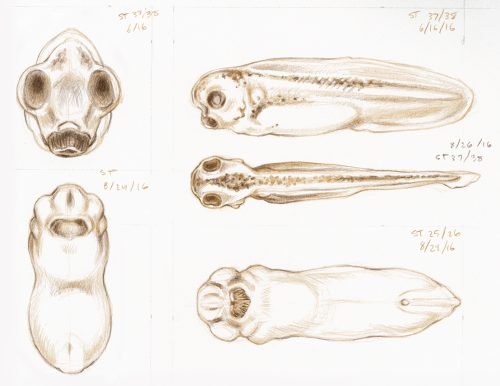
Illustrator Natalya Zahn on the role of observation and visual interpretation in her work creating an addendum to Nieuwkoop and Faber’s classic Normal Table of Xenopus laevis
As an artist of science and nature subjects, I’m often asked what makes the work I do better than a photograph. It makes perfect sense to imagine that a direct photographic capture of an object would offer the very most accurate description of that object – and photography certainly is a brilliant format for capturing detail. What that imagination fails to take into account is that a camera must capture everything that it sees in any given shot. Depending on the complexity or ambiguity of the subject, a highly detailed photograph may just as easily overwhelm or confuse a viewer, rather than offer clarification. The beauty of illustration is that it can precisely isolate specific forms or features of a subject, omitting irrelevant details and making clear essential attributes. It is this quality of simplified clarification, produced through execution and interpretation, that can, under the right circumstances, make scientific illustration a more successful form of visual communication than photography.
In early 2016 I was commissioned by Dr. Michael Levin and Dr. Dany Adams, at the Tufts Center for Regenerative and Developmental Biology, to create a new series of embryo development illustrations for Xenopus laevis, in the style of Nieuwkoop and Faber’s classic Normal Table, originally illustrated by J. J. Prijs in 1956. The new illustrations would be similar enough in drawing quality so as to seamlessly fit into the existing atlas, but they would feature views of embryos and tadpoles that had not previously been generated (including anterior and ventral views that highlight craniofacial changes).
My work began with many hours observing and sketching live specimens under a Nikon SMZ1500 stereomicroscope; multiple individuals from each stage were used as models. Photographic captures were made using digital micrograph, for later reference, but even high-quality photos can include areas of vague detail. The subtle surface contours in early stage embryos, and the intricacy of numerous visible layers of translucent anatomy in later tadpole stages, made the value of snapshots in time a distant second to my live study. By personally observing numerous embryos of the same stage (taking variation and anomaly into account), and periodically adjusting the angle of my view (to illuminate areas of ambiguity), I was able to completely understand each developmental stage in three dimensions, which brought significantly higher accuracy to my interpretation and rendering of each two-dimensional illustration.
Preliminary Xenopus sketches, drawn in colored pencil from live embryo specimens under a microscope (artist’s notations visible).
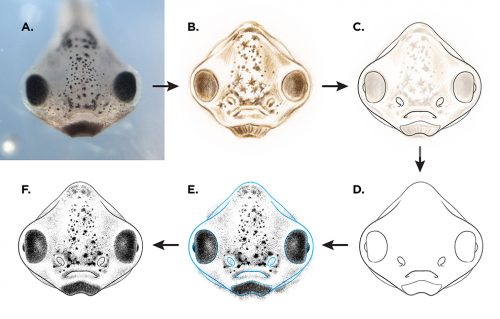
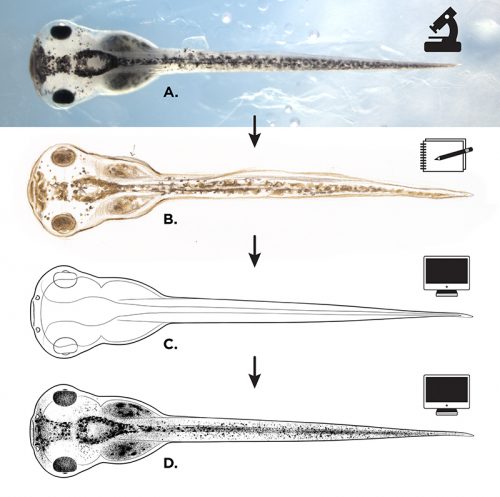
Once I was satisfied with the form and detail of my sketches, I imported each drawing into Adobe Illustrator. Drawing in a vector-based application allowed me to generate the final illustration outlines in extremely clean and consistent line weights. Minimal shading was added in Photoshop, though many of the illustrations are available for use in both shaded and outline-only versions (the later potentially being useful to those interested in adding their own visual notation to an illustration).
My hope for this body of work is that it becomes an invaluable standardized reference for the scientific community, just as Mr. Prijs’ beloved Xenopus illustrations have been for the last half century. Micrographs have only improved since the 1950s but the enduring nature of the drawings within the Nieuwkoop and Faber classic Normal Table is a testament to the enormous teaching power and utility of well executed illustration.
Illustration process for the “The Zahn Drawings”; A: live embryo reference (observed and photographed), B: pencil sketch, C & D: vector line drawing in Adobe Illustrator, E: shading in Photoshop, F: merging of line and shading layers to create finished illustration.
Illustration process for the “The Zahn Drawings”; A: live embryo reference (observed and photographed), B: pencil sketch, C: vector line drawing in Adobe Illustrator, D: shading and merging of layers to create finished illustration.
Natalya Zahn is a Boston-based illustrator, designer and visual story-teller. Deeply inspired by animals and nature, her award-winning work explores the intersection of art, design and science and has been featured by National Geographic, Longwood Botanical Gardens, the San Diego Zoo, and MIT Media Lab, among many others. She is skilled in both traditional media and digital techniques, and works fluently across commercial, educational, editorial, and corporate industries.
Here are the highlights from the current issue of Development:
AdamTS-A keeps the CNS in shape
The Drosophila central nervous system (CNS) is covered by a thick basement membrane that mediates interactions with glial cells and governs the shape of the tissue. Basement membranes are formed from a meshwork of secreted extracellular matrix (ECM) proteins and must be continually remodelled to accommodate growth during development. Metalloproteinases, such as those in the AdamTS family, can break down ECM proteins in the basement membrane and thus allow for tissue expansion. On p. 3102, James Skeath and colleagues identify one protein in the family, AdamTS-A, that is critical for both maintaining the structural integrity of the Drosophila CNS and keeping cell lineages anchored in the tissue. When the function of this protein is reduced, neurons escape the CNS and invade the peripheral tissues of the developing larva. AdamTS-A acts through protease-dependent and possibly protease-independent mechanisms to retain neurons within the developing CNS, and regulates tissue stiffness by restricting Collagen IV/Viking accumulation in the basement membrane. These findings demonstrate the crucial role for extracellular proteases in tissue development and highlight the importance of the basement membrane in shaping the nervous system.
Morphogens and microcolonies: directing stem cell fate
Morphogens are long-range signals that provide cells in the embryo with positional information, helping to ensure cells differentiate appropriately for their position along the body axes. In vivo, and in vitro when cell colonies are large, the function of a single morphogen is difficult to study in isolation. This is because interaction between cells and crosstalk from other signalling factors can confound results. On p. 3042, Aryeh Warmflash and colleagues show that upon stimulation with BMP4, which induces a mixture of fates in large colonies of human embryonic stem cells (hESCs), colonies of just one to eight cells instead acquire trophectoderm-like fate uniformly. They show that above a threshold concentration, BMP4 acts as an ‘on-off’ switch to specify trophectoderm-like cells in these microcolonies, and that a community effect ensures all cells acquire the same fate. In contrast, the mixed cell fates in larger colonies result from the activity of a secondary signal, Nodal, which is produced endogenously by the cells. These findings provide insight into how cells in culture might be directed to specify a single lineage more reproducibly, and demonstrate how the same morphogen signals can be used repeatedly in development to achieve different outcomes, depending on their context.
Pax8+ secretory cells: progenitors of the oviductal epithelium
The mammalian oviduct is lined by a pseudostratified epithelium comprising two main cell types: secretory cells and ciliated cells. Like many other epithelia, it is thought to also contain a population of stem cell-like progenitors, which are important to maintain tissue homeostasis. Such progenitors are hypothesised to actively divide, to facilitate epithelial regeneration upon wounding and to ensure the tissue remains healthy. However, these cells have remained unidentified. On p. 3031, Pradeep Tanwar and colleagues use lineage tracing in mice to identify the oviductal progenitors as secretory cells expressing the Pax8 marker gene. They show that these cells actively divide in the oviductal epithelium, and that this cell population is expanded in humans who are predisposed to ovarian cancer. Like progenitor populations in other tissues, the Pax8+ cells respond to canonical Wnt signalling, which governs their differentiation into ciliated cells and simultaneously maintains the stem cell-like population. These results are a step forward in understanding tissue homeostasis in the oviduct, and may provide insight into how cell proliferation is regulated and subsequently becomes dysregulated in ovarian cancer.
PLUS…
Meeting Review
Alecia-Jane Twigger andChristina H. Scheel report from an EMBO meeting on advances in stem cells and regenerative medicine
Reviews
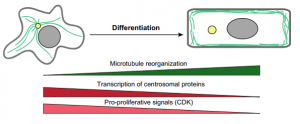
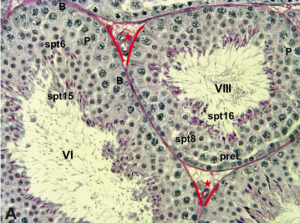
Andrew Muroyama andTerry Lechler summarize current knowledge of how microtubule organization and dynamics change upon cellular differentiation, and Dirk G. de Rooij provides an overview of the organization and timing of spermatogenesis.
Laboratory of Protein Metabolism in Development and Aging
International Institute of Molecular and Cell Biology in Warsaw
is seeking a talented Postdoctoral fellow
Location: Warsaw, a vibrant city with an international academic research environment. International Institute of Molecular and Cell Biology (www.iimcb.gov.pl) – one of the most dynamic and top ranked Polish research institutes.
Job description: Laboratory of Protein Metabolism in Development and Aging, which will be opening on August 2017, is seeking highly motivated and talented Postdoc to join young team investigating the protein homeostasis in development and aging. We use both genetic, molecular and biochemical approaches, primarily in the C. elegans, to study proteolytic networks. Postdoc fellowship is funded in frame of National Science Centre OPUS grant.
Summary: Organismal development or environmental stimuli challenge the homeostatic protein balance (proteostasis) of individual cells, tissues or the entire organism. The ubiquitin proteasome system (UPS) is a key determinant of proteostasis as it regulates the turnover of damaged proteins supporting cellular protein homeostasis and thereby maintains the proteome during stress and aging. Our long-term objective is to understand the mechanistic and developmental aspects of protein degradation pathways defined by combinations of particular ubiquitin ligases (E3). The identification of stress and aging-induced signals that coordinate the interplay between specific E3s will offer intriguingly new mechanistic insights how proteolytic networks are fine-tuned to maintain the cellular proteome and support development and longevity.
Qualifications:
• PhD (or be close to completion) in Molecular Biology, Cell Biology, Protein Chemistry, Genetics or a related discipline;
• experience in C. elegans or cell culture is an advantage;
• experience in Next Generation Sequencing techniques (RNA-Seq, ChIP-Seq) and genomic engineering is an advantage;
• good writing and oral communication skills in English, and competence in scientific writing.
How to apply:
Please send your application including CV, motivation letter and the list of publications to the e-mail address: wpokrzywa@iimcb.gov.pl, until 20th September 2017. Thanking all applicants for their interest, we will contact only selected candidates for an interview.
Please include in your application the following statement: “In accordance with the personal data protection act from the 29th of August 1997, I hereby agree to process and to store my personal data by the Institution for recruitment purposes”.
The recruitment procedure fulfills the National Science Centre’s regulations on granting the scholarships to young scientists.
Selected publications:
Riga T*, Pokrzywa W*, Kevei E, Akyuz M, Vishnu Balaji, Svenja Adrian, Hoehfeld J, Hoppe T. (2017). The ubiquitin ligase CHIP integrates proteostasis and aging by regulation of insulin receptor turnover. Cell. 169: 470-482.
Ackermann L., Schell M., Pokrzywa W., Gartner A., Schumacher B., Hoppe T. (2016). E4 ubiquitin ligase specific degradation hubs coordinate DNA double strand break repair and apoptosis. Nat Struct Mol Biol. 23: 995-1002
Kaushik S, and Cuervo AM (2015). Proteostasis and aging. Nat Med. 21, 1406-15
Frumkin A, Dror S, Pokrzywa W, Bar-Lavan Y, Karady I, Hoppe T, Ben-Zvi A. (2014). Challenging muscle homeostasis uncovers novel chaperone interactions in Caenorhabditis elegans. Front Mol Biosci., doi: 10.3389
van Oosten-Hawle P, and Morimoto RI (2014). Organismal proteostasis: role of cell-nonautonomous regulation and transcellular chaperone signaling. Genes & Dev. 28: 1533-43
Segref A, Kevei E, Pokrzywa W, Mansfeld J, Schmeisser K, Livnat-Levanon N, Ensenauer R, Glickman M.H, Ristow M, Hoppe T. (2014). Pathogenesis of human mitochondrial diseases is modulated by reduced activity of the ubiquitin/proteasome-system. Cell Metab. 4:642-652
Pokrzywa W. and Hoppe T. (2013). Chaperoning myosin assembly in muscle formation and aging. Worm. 2:e25644
Gazda L*, Pokrzywa W*, Hellerschmied D, Loewe T, Forné I, Mueller-Planitz F, Hoppe T, Clausen T. (2013). The myosin chaperone UNC-45 is organized in tandem modules to support myofilaments formation in C. elegans. Cell. 1, 183-195.
Kuhlbrodt K, Janiesch PC, Kevei E, Segref A, Barikbin R, and Hoppe T (2011). The Machado-Joseph disease deubiquitylase ATX-3 couples longevity and proteostasis. Nat Cell Biol. 13, 273-81

 (1 votes)
(1 votes)





 (No Ratings Yet)
(No Ratings Yet)















 (10 votes)
(10 votes)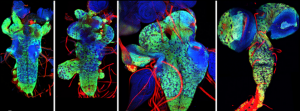 On p.
On p. 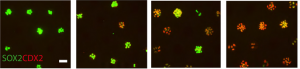 On p.
On p. 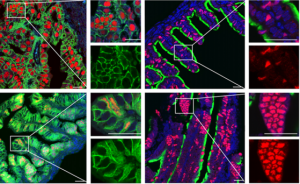 On p.
On p.