One of the research topics in Michel Milinkovitch’s laboratory (https://www.lanevol.org) at the University of Geneva (Switzerland) is to understand how squamates (lizards and snakes) generate such a tremendous variety of colours and colour patterns.
Colours
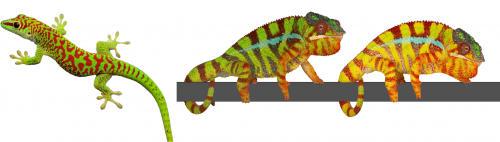
The colour of a lizard’s patch of skin is generally the result of the combination among structural and pigmentary elements found in various types of chromatophores(1-5). Pigmentary colours are produced by brown/black melanins in melanophores, as well as by yellow and red pteridines/carotenoids in xanthophores and erythrophores, respectively. On the other hand, structural colours are produced by light interference in iridophore cells containing layers of guanine nanocrystals(5): the wavelengths specifically reflected by these periodic structures is a function of the mean distance between successive layers of guanine nanocrystals — the longer the distance, the longer the wavelengths that are reflected. For example, many species of reptiles and amphibians are green despite that their skin does not contain any green pigment! They produce their chlorophyll-matching colour in a more sophisticated way(4): a layer of iridophores selectively reflects most of the incoming green and blue wavelengths but a layer of yellow pigments absorbs blue. As all other wavelengths of the visible range (yellows, oranges, and reds) go through the skin (and are absorbed by deeper tissues), the only colour that bounces back from the skin is pure bright green(4), as in day geckos (Fig. 1a).
The Milinkovitch lab also discovered that chameleons change colour by manipulating structural colours rather than by dispersion/aggregation of pigment-containing organelles within chromatophores. Indeed, combining microscopy, videography, RGB photometry and photonic band-gap modelling, they showed that chameleons shift colour through active tuning of a 3D lattice of guanine nanocrystals within a superficial layer of dermal iridophores(5). In other words, chameleons manipulate light interference by changing the distance among their nanocrystals of guanine. Take an adult male panther chameleon. In its cryptic state, it is green for the same reason as the Phelsuma lizards: its dermal iridophores reflect green and blue, and the latter is being absorbed by a yellow pigment. But if another mature male enters its territory, the two animals increase the distance between the nanocrystals within their iridophores … such that they both turn yellow or orange or red to become as visible as possible and impress each other (Fig. 1b and link to YouTube videos). This usually suffices for one of the two males to give up…otherwise they will start a physical fight.
These studies provided some new answers but also opened many new questions. How do iridophores generate and spatially organise nanocrystals? What is the cellular mechanism involved in the tuning of the distance among nanocrystals in iridophores of chameleons when they change colour? Milinkovitch is now teaming up with biochemists/cell biologists such as Marcos Gonzalez-Gaitan for investigating these questions.
Fig. 1 (a- left) A day gecko (Phelsuma grandis); (b-right): A male panther chameleon (Furcifer pardalis) changing colour.
Colour change in chameleons YouTube playlist.
Colour Patterns
Animals display colours, but they often additionally exhibit colour patterns, i.e., symmetry-breaking regularities (stripes, spots, tessellations, meanders, and labyrinths) that result from short-range and long-range interactions among chromatophores(6-11). At the macroscopic scale, these dynamical processes obey reaction-diffusion (RD) equations discovered by the mathematician Alan Turing(12-14). Strikingly, the formation of skin colour in the ocellated lizard (Timon lepidus) seems to conflict with this RD framework as skin scales, rather than individual chromatophore cells, establish the pattern. Indeed, the brown juvenile lizard gradually transforms its skin colour as it ages to reach an intricate adult labyrinthine pattern where each scale is either green or black (Fig. 2).
Figure 2: The colour pattern changes drastically in about three years from (top) the juvenile to (bottom) the adult. The inset shows a juvenile on the same scale as the adult. Scale bar = 11 mm.
But why would the pattern form at the level of scales, rather than at the level of biological cells? This is the questions that the Milinkovitch team solved recently, as reported recently in the journal Nature(15).
To tackle this question, two PhD students, Liana Manukyan (computer scientist) and Sophie Montandon (developmental biologist), followed individual lizards during three to four years of their development from hatchlings crawling out of the egg to fully mature animals. For multiple time points, they reconstructed the geometry and colour of the network of scales on three animals by using R2OBBIE-3D (YouTube playlist), a very high resolution robotic system(16) developed previously in the Milinkovitch laboratory by a physicist PhD student: Antonio Martins.
R2OBBIE-3D YouTube playlist
R2OBBIE-3D allows to reconstruct the 3D geometry and colour texture of objects up to 1.5 meters with a resolution of … 15 microns ! The Swiss team then set up a software pipeline that allowed them to automatically detect scales on each animal at each time point and match these networks. This was not a trivial task because the size of the animal, the positions of its body parts, and its skin pattern all change from scan to scan. Fortunately, the number of scales is invariant for a given individual throughout its life. This analysis indicated that the brown juvenile scales change to green or black then, surprisingly, continue flipping colour (between green and black) during the life of the animal (Fig. 3).
Figure 3: Close-ups of the same individual as in Fig 2 illustrating the pattern time evolution. Orange arrows show two examples of colour switching between two time points.
This very strange observation prompted Milinkovitch to suggest that the skin scale network forms a so-called ‘Cellular Automaton’. This esoteric computing system was invented in 1948 by the mathematician John von Neumann. Cellular automata are lattices of elements in which each element changes its state (here, its colour, green or black) depending on the states of neighbouring elements. The elements are called cells but are not meant to represent biological cells; in the case of the lizards, they correspond to individual skin scales. These abstract automata were extensively used to investigate computing systems and to model natural phenomena, but the Geneva team discovered what seems to be the first case of a genuine 2D automaton appearing in a living organism. Analyses of the four years of colour change allowed to confirm Milinkovitch’s hypothesis: the scales were indeed flipping colour depending of the colours of their neighbour scales. Computer simulations implementing the discovered mathematical rule generated colour patterns that could not be distinguished from the patterns of real lizards.
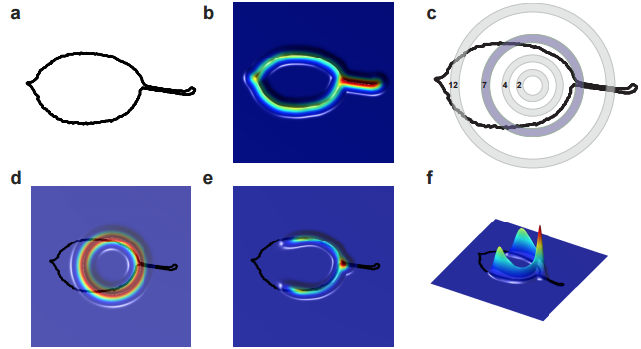
How could the interactions among pigment cells, described by Turing equations, generate a von Neumann automaton exactly superposed to the skin scales? The skin of a lizard is not flat: it is very thin between scales and much thicker at the center of them. Given that Turing’s mechanisms involves movements of cells, or the diffusion of signals produced by cells, Milinkovitch understood that this variation of skin thickness could impact on the Turing’s mechanism. Liana Manukyan, but also Anamarija Fofonjka, a third PhD student in Milinkovitch’s team, then performed computer simulations and saw a cellular automaton behaviour emerge, demonstrating that the development of Cellular Automata as computational systems is not just an abstract concept developed by John von Neumann, but also corresponds to a natural process generated by biological evolution.
However, the automaton behaviour was imperfect as the mathematics behind Turing’s mechanism and von Neumann automaton are very different. Milinkovitch called in Stanislav Smirnov, Professor of mathematics at the University of Geneva. Stanislav was awarded in 2010 the Fields Medal (the equivalent of the Nobel Price in Mathematics). Before long, Smirnov derived a so-called discretisation of Turing’s equations that would constitute a formal link with von Neumann’s automaton. Anamarija Fofonjka implemented Smirnov new equations in computer simulations, obtaining a system that had become un-differentiable from a von Neumann automaton (Fig. 4). The highly multidisciplinary team of researchers had closed the loop in this amazing journey, from biology to physics to mathematics.
Figure 4. A CA behaviour rapidly emerges from the continuous RD process when diffusion coefficients are reduced by a factor 1-P > 0.8 in the inter-scale regions (from left to right, 1-P=0.2, 0.6, 0.8, 0.9, 0.95, 0.99).
Hiroshi Hamada is the Director of the RIKEN Center for Developmental Biology (CDB) in Kobe, Japan. His lab focusses on the establishment of left-right asymmetry in the mouse embryo, and the role of cilia in the symmetry-breaking event. Hiroshi’s work has been recognised by various awards, including the Keio Medical Science Prize in 2014, and election as an EMBO Associate Member in 2016. We met with Hiroshi on a recent visit to the CDB, to talk about his career and current interests, and the prospects for developmental biology in Japan.
How did you become interested in science generally and developmental biology in particular?
I was interested in science from when I was really young. I don’t know why, but I used to like to read science books and in particular I was interested in basic medical science, not in clinical medicine. So I thought I wanted to be a basic research scientist. But my interests in developmental biology are relatively recent. When I started in research, I wanted to contribute to medicine, and because cancer was the most serious disease at that point (as well as now), that was what I wanted to study. But, to start, I thought I needed to get into molecular biology (which didn’t really exist as a field at the time), so as a graduate student I studied the structure of RNA – I was a biochemist – and then I learned a lot of cloning techniques when they came along. These were very useful when I moved to the NIH for my postdoc to work on cell transformation, to look at oncogenic mutations.
You first established your own lab in Newfoundland, Canada. How did that come about?
At that time, I wanted to be more independent, so I looked for a job in the USA rather than moving back to Japan. I had a relatively decent publication record and I thought I would have a good chance of choosing where I wanted to go. But I interviewed at several places in the USA and Canada, and Newfoundland was the only place I got an offer.
And how was it there?
It was a wonderful place – the people were very kind and the university was very decent. Most of the people there were more senior and were focussing mainly on education, so they said that because I had a grant I could focus on the research and didn’t have to take on other responsibilities. So I was very happy there.
But then you did eventually go back to Japan?
Yes, but that was not my plan. I really enjoyed Newfoundland, but I had two young kids and Newfoundland wasn’t the best place to be for their education. So I was planning to move to another university in Canada, where I had an offer. Then my supervisor from Japan called me – he was planning to retire in 5 years, and his associate professor had been promoted and moved elsewhere, so he needed someone to take over from him. I decided to return to Japan, first to Tokyo and then later to Osaka.
How did you find the differences in the science culture between North America and Japan?
When I first moved back, the facilities in Japan weren’t as good – even compared with Newfoundland. Now, of course, things are much better here, but at the time all the equipment and so on was quite old. And of course there were huge differences in the culture… I felt almost half-Western by then, so I found it very different when I moved back. Fortunately, my professor ensured that I could do my own research and be independent and kindly assigned the best graduate students to me.
So when did you actually become a developmental biologist?
When I was in Canada, I started working on embryonal carcinoma (EC) cells, and I was interested in differentiation – these cells differentiated very nicely. One of the best EC lines, P19 cells, was established by Michael McBurney at University of Ottawa, and he showed me how to induce the differentiation. So I took that system to Newfoundland, and I tried to answer several questions, such as what determined the undifferentiated state and what triggered differentiation into neural cells.
We tried to identify the genes specifically expressed in undifferentiated cells, and the transcription factors that recognised the undifferentiated cell-specific enhancers. This second approach was very successful – we used enhancer trapping to identify an enhancer active in undifferentiated cells. And then when I came back to Japan, we identified the specific transcription factor that bound this enhancer – which turned out to be Oct4, though we originally called it Oct3. We and two other groups all reported the same transcription factor around the same time; two of us called it Oct3, and Hans Schöler called it Oct4, which was the name that eventually stuck.
You then discovered Lefty, also in a tissue culture screen, and this led you into working on axis establishment and left-right (LR) asymmetry, which has been your focus for the past two decades. What do you think are the big open questions in the field?
I think we have been able to find out some of the important principles underlying LR axis determination, but there are still many important unanswered questions. For example, we know that fluid flow is crucial – the rotation of motile cilia in the node generates directional flow – but we don’t really know how embryos sense this flow. The first molecular asymmetry we see is the degradation of a particular mRNA on one side of the node, but it’s not clear how this is induced. Of course, we know that immotile cilia are involved in flow-sensing, but how do they do this? Calcium signalling seems to be involved, but the pathway still isn’t clear and this is an active area of research for us.
Another question is what determines the direction in which the cilia rotate, which in turn sets up left versus right. We know there must be inputs from the anterior-posterior and dorsal-ventral patterning systems, and also that there must be some chirality in the system to set up the asymmetry. But there is a lot still to figure out for us to understand why the cilia always rotate in the clockwise direction. Also, it seems like the node cilia are the only ones in the body that rotate as opposed to beat, but why is that?
Your research encompasses a broad range of scales – from the molecular architecture of cilia to the developmental consequences of disrupting LR determination in mice. How do you stay abreast of the latest techniques required to investigate the problem at such diverse levels?
For myself, I think that the question always comes first. I don’t think about what I can do with a particular technique, I think about the question and then try to work out what kind of techniques are necessary to address it. And then of course I have to find the people who can use that technique – either through collaboration or people in my lab. Fortunately, I’ve had good people come to my lab, including people with engineering backgrounds, who’ve been very good at analysing the flow and the mechanics. I didn’t try to recruit them, but they somehow found me, and that’s been great.
I understand you’ve also become interested in mathematical and computational approaches to investigating axis establishment. How do you think such approaches can contribute?
Initially, I wasn’t really sure what mathematical modelling or computational science can do for real biology. But I was exposed to the work of my friend Shigeru Kondo, who is interested in Turing patterns and reaction-diffusion models in pigmentation. Then, when I was studying the very dynamic patterns of gene expression during LR patterning, we did lots of experiments but they didn’t really get at the principle of what’s going on. We thought that reaction-diffusion might be involved, so we decided to try and model it. And this has really helped, and I realised what modelling can do for you. When it works, it really clarifies things in a way that lab experiments couldn’t. Personally, I’m not good at mathematics and physics, but I collaborate with people who understand this, and I find it very useful.
You took over as Director of the CDB (probably the most prestigious developmental biology institute in Japan) a couple of years ago, in the wake of the problems with and fallout from the STAP cell papers. How is the institute moving forwards from that difficult period and what do you hope to achieve as director?
Masatoshi Takeichi was stepping down as Director, and partly because I was on the advisory council of the CDB and was familiar with the institute, they asked me if I would take over. And I have a great respect for the CDB and the people, including Takeichi-sensei, and how they run the institute, which has always been very fair. For example, they always make sure to select the best people for positions rather than choosing their friends, which is of course very important. And everything they do is like this. So I knew that this was a very important institute and we couldn’t see it run into difficulty or disappear – we had to support it, and I hope I have done that as Director.
I see my job mainly as to rejuvenate things – several people have moved on and we have recruited and keep recruiting some very good new people, so we are looking to the future. One thing we want to do is to extend the scope of the CDB a bit. Traditionally, we have mainly focussed on embryonic development, but we now see development as something that continues through life. And so we want to recruit people working on maintenance and homeostasis – adult stem cells – as well as regeneration and ageing.
Traditionally, we have mainly focussed on embryonic development, but we now see development as something that continues through life
More broadly, Japanese science has a long and prestigious history in developmental biology, and is now at the forefront of stem cell research, which of course depends on insights from developmental biology. How do you see the balance between basic and translational research in Japan, particularly in terms of funding?
Fortunately RIKEN headquarters appreciate the importance of developmental biology and they are committed to supporting the field. But we cannot go on with just basic developmental biology – we have to have a balance between basic and more translational science, and this is sensible. Fortunately for us, Masayo Takahashi, who has pioneered transplantation of induced pluripotent stem cell-derived cells into human patients, is based here at the CDB. She likes to be here because she knows that the basic science is important and that the reason she can do translational work is because of the basic foundations. There has to be interaction between basic and translational science, and that can be a little bit difficult, but we have to make sure we bring these people together and I am trying to promote this.
More broadly, I think that there is a larger proportion of grant money in Japan going to stem cell and other translational research, and basic scientists are having a hard time, but I think that’s true everywhere. However, it is essential to maintain the diversity in research, because, as history tells us, big breakthroughs do not always come from top-down projects but from unexpected directions.
What advice would you give to young scientists starting out their career in developmental biology?
Try to find an interesting question, and one that you think is important; then, stick with it, be patient and don’t compromise to easier questions. It might take a long time, but I think you have to do it. I guess the current situation makes it difficult for young people – who have to publish – so maybe I’m saying something unrealistic. But what I used to do was to have side projects that were maybe easier questions, but always returned to the main theme.
Try to find an interesting question, and one that you think is important; then, stick with it, be patient and don’t compromise to easier questions
Away from the lab, what might Development readers be surprised to find out about you?
I very much enjoyed my time in Newfoundland – it was a great place with lots of small fishery towns and wonderful wildlife, and I would love to go back. They have a big Irish population, and I discovered Irish music there, which I really like. The first time I visited, when I went for the job interview, I went to my host’s house for dinner. And I could hear some Irish music playing in the background. As a result, I am probably one of the few Japanese people who love traditional Irish music.
Our latest monthly trawl for developmental biology (and other cool) preprints. See last year’s introductory post for background, and let us know if we missed anything
This month featured a host of preprints on plants, stem cells, connectomics and modelling. Plus the first preprint from Nobel laureate and recent Development interviewee Eric Wieschaus, an introduction to the concept of the Human Cell Atlas, and a plea from Science Editor-in-Chief Marcia McNutt and colleagues for transparency in author contributions. The preprints were hosted on bioRxiv, F1000Research, PeerJ and arXiv.
Organization Of The Drosophila Larval Visual Circuit. Ivan Larderet, Pauline Fritsch, Nanae Gendre, Larisa Maier, Rick D. Fetter, Casey Schneider-Mizell, James Truman, Marta Zlatic, Albert Cardona, Simon Sprecher
Kenyon cells from a first instar fly larvae in Eichler, et al.’s preprint
The Complete Connectome Of A Learning And Memory Center In An Insect Brain. Katharina Eichler, Ashok Litwin-Kumar, Feng Li, Youngser Park, Ingrid Andrade, Casey M. Schneider-Mizell, Timo Saumweber, Annina Huser, Claire Eschbach, Bertram Gerber, Richard D. Fetter, James W. Truman, Carey E. Priebe, L. F. Abbott, Andreas S. Thum, Marta Zlatic, Albert Cardona
A Complete Electron Microscopy Volume Of The Brain Of Adult Drosophila melanogaster. Zhihao Zheng, J. Scott Lauritzen, Eric Perlman, Camenzind G. Robinson, Matthew Nichols, Daniel Milkie, Omar Torrens, John Price, Corey B. Fisher, Nadiya Sharifi, Steven A. Calle-Schuler, Lucia Kmecova, Iqbal J. Ali, Bill Karsh, Eric T. Trautman, John Bogovic, Philipp Hanslovsky, Gregory S. X. E. Jefferis, Michael Kazhdan, Khaled Khairy, Stephan Saalfeld, Richard D. Fetter, Davi D. Bock
Whole-Brain Serial-Section Electron Microscopy In Larval Zebrafish. David Grant Colburn Hildebrand, Marcelo Cicconet, Russel Miguel Torres, Woohyuk Choi, Tran Minh Quan, Jungmin Moon, Arthur Willis Wetzel, Andrew Scott Champion, Brett Jesse Graham, Owen Randlett, George Scott Plummer, Ruben Portugues, Isaac Henry Bianco, Stephan Saalfeld, Alex David Baden, Kunal Lillaney, Randal Burns, Joshua Tzvi Vogelstein, Alexander Franz Schier, Wei-Chung Allen Lee, Won-Ki Jeong, Jeff William Lichtman, Florian Engert
Highly Efficient Maternal-Fetal Zika Virus Transmission in Pregnant Rhesus Macaques. Sydney M Nguyen, Kathleen M Antony, Dawn M Dudley, Sarah Kohn, Heather Simmons, Bryce Wolfe, M Shahriar Salamat, Leandro BC Teixeira, Gregory J Wiepz, Troy H Thoong, Matthew T Aliota, Andrea M Weiler, Gabrielle L Barry, Kim L Weisgrau, Logan J Vosler, Mariel S Mohns, Meghan E Breitbach, Laurel M Stewart, Mustafa N Rasheed, Christina M Newman, Michael E Graham, Oliver E Wieben, Patrick A Turski, Kevin M Johnson, Jennifer Post, Jennifer M Hayes, Nancy Schultz-Darken, Michele L Schotzko, Josh A Eudailey, Sallie R Permar, Eva G Rakasz, Emma L Mohr, Saverio Capuano III, Alice F Tarantal, Jorge E Osorio, Shelby L O’Connor, Thomas C Friedrich, David H O’Connor, Thaddeus G Golos
Olfactory Receptors Are Required For Social Behavior And Neural Plasticity In Ants, As Evidenced By CRISPR-Mediated Gene Knockout. Hua Yan, Comzit Opachaloemphan, Giacomo Mancini, Huan Yang, Matthew Gallitto, Jakub Mlejnek, Kevin Haight, Majid Ghaninia, Lucy Huo, Alexandra Leibholz, Jesse Slone, Xiaofan Zhou, Maria Traficante, Clint A. Penick, Kelly Dolezal, Kaustubh Gokhale, Kelsey Stevens, Ingrid Fetter-Pruneda, Roberto Bonasio, Laurence J. Zwiebel, Shelley Berger, Juergen Liebig, Danny Reinberg, Claude Desplan
Development of a high-density, ~2M SNP genotyping array and 670k SNP imputation array for the domestic horse. Robert J Schaefer, Mikkel Schubert, Ernest Bailey, Danika L. Bannasch, Eric Barrey, Gila Kahila Bar-Gal, Gottfried Brem, Samantha A. Brooks, Ottmar Distl, Ruedi Fries, Carrie J. Finno, Vinzenz Gerber, Bianca Haase, Vidhya Jagannathan, Ted Kalbfleisch, Tosso Leeb, Gabriella Lindgren, Maria Susana Lopes, Nuria Mach, Artur da Câmara Machado, James N. MacLeod, Annette McCoy, Julia Metzger, Cecilia Penedo, Sagi Polani, Stefan Rieder, Imke Tammen, Jens Tetens, Georg Thaller, Andrea Verini-Supplizi, Claire M. Wade, Barbara Wallner, Ludovic Orlando, James R. Mickelson, Molly E. McCue
The Human Cell Atlas. Aviv Regev, Sarah Teichmann, Eric S. Lander, Ido Amit, Christophe Benoist, Ewan Birney, Bernd Bodenmiller, Peter Campbell, Piero Carninci, Menna Clatworthy, Hans Clevers, Bart Deplancke, Ian Dunham, James Eberwine, Roland Eils, Wolfgang Enard, Andrew Farmer, Lars Fugger, Berthold Gottgens, Nir Hacohen, Muzlifah Haniffa, Martin Hemberg, Seung K. Kim, Paul Klenerman, Arnold Kriegstein, Ed Lein, Sten Linnarsson, Joakim Lundeberg, Partha Majumder, John Marioni, Miriam Merad, Musa Mhlanga, Martijn Nawijn, Mihai Netea, Garry Nolan, Dana Pe’er, Anthony Philipakis, Chris P. Ponting, Stephen R. Quake, Wolf Reik, Orit Rozenblatt-Rosen, Joshua R. Sanes, Rahul Satija, Ton Shumacher, Alex K. Shalek, Ehud Shapiro, Padmanee Sharma, Jay Shin, Oliver Stegle, Michael Stratton, Michael J. T. Stubbington, Alexander van Oudenaarden, Allon Wagner, Fiona M. Watt, Jonathan S. Weissman, Barbara Wold, Ramnik J. Xavier, Nir Yosef
TIRF-M images and associated kymographs from Tan, et al.’s preprint
Loss Of PTEN Promotes Formation Of Signaling-Specific Clathrin-Coated Pits. Luciana K. Rosselli-Murai, Joel A. Yates, Sei Yoshida, Julia T. Bourg, Kenneth K. Y. Ho, Megan White, Julia Prisby, Xinyu Tan, Megan Altemus, Liwei Bao, Zhi-Fen Wu, Sarah L. Veatch, Joel A. Swanson, Sofia D. Merajver, Allen P. Liu
Drosophila embryos and aspect ratios, from Atia, et al.’s preprint
Universal geometric constraints during epithelial jamming. Lior Atia, Dapeng Bi, Yasha Sharma, Jennifer A. Mitchel, Bomi Gweon, Stephan Koehler, Stephen J. DeCamp, Bo Lan, Rebecca Hirsch, Adrian F. Pegoraro, Kyu Ha Lee, Jacqueline Starr, David A. Weitz, Adam C. Martin, Jin-Ah Park, James P. Butler, Jeffrey J. Fredberg
Aberrant Cortical Activity In Multiple GCaMP6-Expressing Transgenic Mouse Lines. Nicholas A. Steinmetz, Christina Buetfering, Jerome Lecoq, Christian R. Lee, Andrew J. Peters,Elina A. K. Jacobs, Philip Coen, Douglas R. Ollerenshaw, Matthew T. Valley, Saskia E. J. de Vries, Marina Garrett, Jun Zhuang, Peter A. Groblewski, Sahar Manavi, Jesse Miles, Casey White, Eric Lee, Fiona Griffin, Joshua D. Larkin, Kate Roll, Sissy Cross, Thuyanh V. Nguyen, Rachael Larsen, Julie Pendergraft, Tanya Daigle, Bosiljka Tasic, Carol L. Thompson, Jack Waters, Shawn Olsen, David J. Margolis,Hongkui Zeng, Michael Hausser, Matteo Carandini, Kenneth D. Harris
SCENIC: Single-Cell Regulatory Network Inference And Clustering. Sara Aibar, Carmen Bravo González-Blas, Thomas Moerman, Jasper Wouters, Vân Anh Huynh-Thu, Hana Imrichová, Zeynep Kalender Atak, Gert Hulselmans, Michael Dewaele, Florian Rambow, Pierre Geurts, Jan Aerts, Jean-Christophe Marine, Joost van den Oord, Stein Aerts
Accuracy, Reproducibility And Bias Of Next Generation Sequencing For Quantitative Small RNA Profiling: A Multiple Protocol Study Across Multiple Laboratories. Maria D. Giraldez, Ryan M. Spengler, Alton Etheridge, Paula Maria Godoy, Andrea J. Barczak, Srimeenakshi Srinivasan, Peter L. De Hoff, Kahraman Tanriverdi, Amanda Courtright, Shulin Lu, Joseph Khoory, Renee Rubio, David Baxter, Tom A. P. Driedonks, Hank P. J. Buermans, Esther N. M. Nolte-‘t Hoen, Hui Jiang, Kai Wang, Ionita Ghiran, Yaoyu Wang, Kendall Van Keuren-Jensen, Jane E. Freedman, Prescott G. Woodruff, Louise C. Laurent, David J. Erle, David J. Galas, Muneesh Tewari
Opportunities And Obstacles For Deep Learning In Biology And Medicine. Travers Ching, Daniel S. Himmelstein, Brett K. Beaulieu-Jones, Alexandr A. Kalinin, Brian T. Do, Gregory P. Way, Enrico Ferrero, Paul-Michael Agapow, Wei Xie, Gail L. Rosen, Benjamin J. Lengerich, Johnny Israeli, Jack Lanchantin,Stephen Woloszynek, Anne E. Carpenter, Avanti Shrikumar, Jinbo Xu, Evan M. Cofer, David J. Harris, Dave DeCaprio, Yanjun Qi, Anshul Kundaje,Yifan Peng, Laura K. Wiley, Marwin H. S. Segler, Anthony Gitter, Casey S. Greene
A persistent lack of International representation on editorial boards in biology. Johanna Espin, Sebastian Palmas-Perez, Farah Carrasco-Rueda, Kristina Riemer, Pablo Allen, Nathan Berkebile, Kirsten Hecht, Renita Kay Kastner-Wilcox, Mauricio Nunez-Regueiro, Candice Prince, Maria Constanza Rios-Marin, Erica P Ross, Bhagatveer Singha, Tia Tyler, Judit Ungvari Martin, Mariana Villegas, Tara Cataldo, Emilio Bruna
Lee lab has been studying on cell specification process using human pluripotent stem cells, particularly peripheral nervous system and skeletal muscle cells.
We are looking for highly motivated postdoc(s) who have experience on optogenetic studies. Recently, by using CRY2-PHR or LOV2 domains, we started to explore how light illumination can spatially and temporally modulate human stem cell fates, which can lead us to have unprecedented precise control as well as to interrogate/measure necessary/sufficient input amounts to elicit a certain biological event.
Previous experience on human pluripotent stem cells is not necessarily required (but basic cell culture experience will be great) and we value more on the ‘non-stem cell’ expertise, such as molecular cloning. Please send your application (CV and three reference contact info) to Gabsang Lee (leelabjob@gmail.com).
A position (#123028) is available immediately for a Research Technician/Faculty Specialist to contribute to our studies in neural crest and placodes. The Technician will conduct research, assist in the training of students, and take part in the management of the laboratory of Dr. Lisa Taneyhill at the University of Maryland. Laboratory skills should include the ability to perform various molecular biology and biochemical assays. Experience with microscopy, chick embryology, and tissue culture is desirable. For more information on the lab, please see http://www.ansc.umd.edu/people/lisa-taneyhill. Qualifications: A Bachelor’s degree (B.A. or B.S.) in a related field and prior laboratory research experience is essential. Fluency in spoken and written English is required. Compensation: Salaries are highly competitive, negotiable and commensurate with qualifications. Fringe benefits offered. Applicants must apply through eTerp at https://ejobs.umd.edu. Applications will be accepted until a suitable candidate is identified.
The Woods Hole Embryology Course, which will celebrate its 124th birthday this year, is a continual source of beautiful images (and videos) of development. Since 2011 the Node has run a competition for the community to pick the best images from a given year – the winning pictures become immortalised as Development covers.
Below you will find 4 images from Round 2 of the 2015 course (Round 1 was won by Theodora Koromila’s lunar chicken embryo), and a poll to vote in at the end. You can see full size versions by clicking on the images.
The poll is set up to allow only one vote per person, and closes at 13.00 BST on Wednesday 21st June.
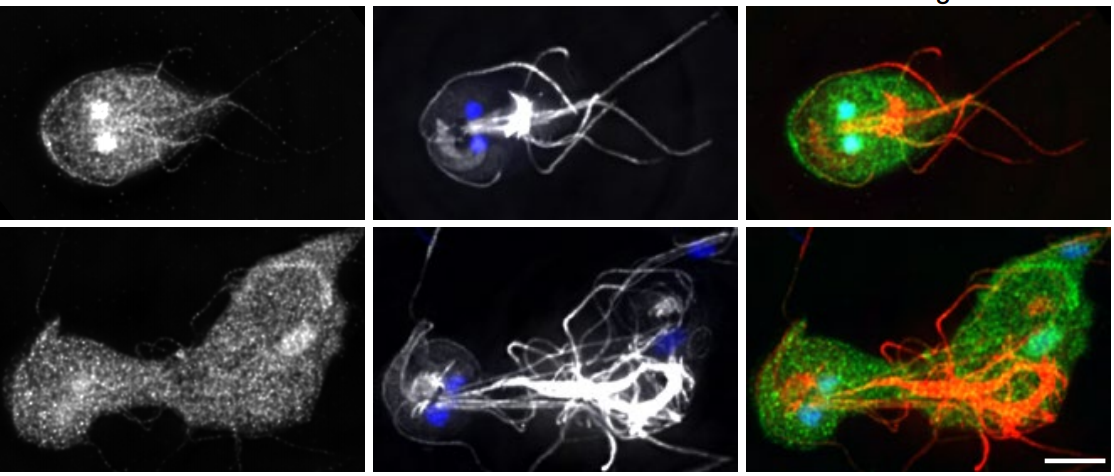
Drosophila
Drosophila melanogaster embryo, stage 17, lateral view with anterior up, ventral to the left. Yellow – anti-HRP, all neuron cell bodies and axons; Pink – anti-Fasciclin II, motoneuron projections; Blue – anti-Tropomyosin, muscles; Green – DAPI, nuclei.
Amjad Askary
University of Southern California, USA
Longhua Guo
Stowers Institute, USA
Maike Getwan
University of Hohenheim, Germany
Nick Shikuma
Cal Tech, USA
Crab
Juvenile crab collected in a plankton tow.
Chiara Sinigaglia
Observatoire Océanologique de Villefranche sur Mer/ CNRS, France
Parhyale
Adult amphipod, Parhyale hawaiensis. Pink – DsRed expression in muscle. Yellow – FITC dextran injected into hemolymph to visualize the circulatory system. Blue – autofluorescence of cuticle.
Longhua Guo
Stowers Institute, USA
Skate
Skate (Leucoraja erinacea) embryo stained with Alcian Blue to reveal cartilage.
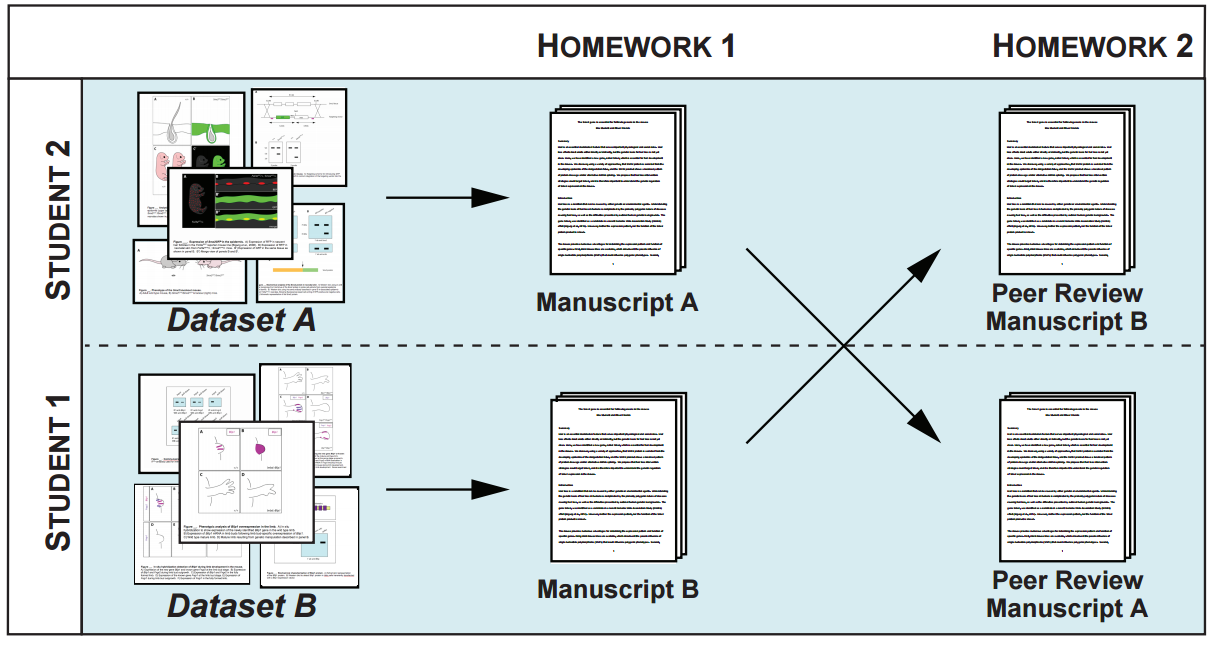
Like most things in life, research requires funding and scholarships. This becomes increasingly apparent as researchers progress through their career. At some point, everyone has to jump on the funding treadmill. For many researchers, the first experience of a grant application process may be applying for postgraduate scholarships for their masters/PhD. While I never pursued this option myself, as a postdoc, I now see it from the other side of the fence.
Writing a grant for your dream lab
It is a good opportunity for PIs to employ someone for 3-4 years at no cost. So if you’re an undergrad looking to apply for such a grant, expect pro forma replies from people only too happy to apply with you. If you already have funding, most PIs will happily host you even if your CV is hand written on the back of a napkin. If you are a young, clever, enthusiastic scientist make no mistake, you are a sought-after commodity so shop around.
Postdoctoral funding is a little different but the same general dynamics apply, particularly with regard to postdoctoral fellowships. What you need to be mindful of is that the application process may be a lot more gruelling and competitive. It is important that you gauge carefully what your chances of success are. Remember, a PI may agree to put their name on your application but the grant writing may be left entirely up to you. It can be a big commitment, particularly when you are in the midst of finishing up your PhD and writing your thesis.
This can be a particularly stressful and unnerving time as you don’t know what the future will hold. It can be difficult put together a competitive grant proposal under such conditions so it is not for the faint hearted. If you are lucky you might just get your dream fellowship and a flying start to your career. If you are less fortunate the PI you apply with, impressed by your efforts, might keep you in mind when they next secure funding. However, if you are unlucky, you get nothing but you learn from the experience always look at the positives.
Post-Doc and PhD student grant writing tips and advice
It is, admittedly, a lot easier to focus on writing a research proposal when you know where the next paycheque is coming from. For this reason, many choose to secure a postdoctoral position before pursuing fellowship funding – a bird in the hand is worth two in the bush. This is probably a less stressful approach than applying for funding during the latter stages of your PhD. However, don’t forget that the clock is ticking from the moment you finish your viva – you will not be eligible for certain early stage’ fellowships after you clock up certain amount of experience e.g. Sir Henry Wellcome Postdoctoral Fellowships (ineligible after one year of postdoctoral experience).
My first postdoc was in the same lab where I did my PhD. In reality, this was more of a mop-up’ period for my doctoral work; several manuscripts were submitted, all of which were eventually published, so paper-wise it was a productive time. My advice to anyone considering this option is don’t do it for too long – identify short-term achievable goals and make sure you have an exit strategy. Unless there is something truly amazing happening in the lab, there are a number of reasons you should probably move on after your PhD. Firstly, moving to a different lab is a new experience and will force you outside of your comfort zone, to meet new people, learn new skills and develop fresh ideas. Secondly, if you want to apply for a fellowship, there are often mobility clauses written into fellowship grants which preclude staying in the same lab where you completed your PhD e.g. Marie Curie Intra-European Fellowships for career development (IEF).
Early stage researcher grants
Aside from fellowships, there are several other funding opportunities for postdoctoral researchers. These generally take the form of larger collaborative grants where you, as an early stage researcher, may not be the lead applicant. If successful, these grants may cover your costs for a number of years so success can buy you breathing room and time to focus on your research. These grants can effectively represent a fellowship from your perspective although their raison d’etre is likely to be delivery of a research goal rather than a postdoc’s career development.
After a six month stint, I waved goodbye to my old lab in 2011. As I was finishing up, finished my ELISAs I applied with my current PI for a postdoctoral fellowship but was unsuccessful. In spite of this I moved to this lab in 2012 as funding became available and so began my first postdoctoral post outside my alma mater. The first couple of months were given to setting up the lab – things like ordering equipment and organising paper work, getting to know people in the department/university, supervising undergrads and then (eventually) getting on with my experiments. However, aside from research, the main focus has been on grant writing.
Drafting scientific grants
I have written eight grants and am currently drafting my ninth; one was successful, another three are still under review and five were unsuccessful. It’s a learning process. There is no magic formula for writing grants and even if you write the best proposal imaginable, success can be decided by factors beyond your control. Nevertheless, to begin with, the guidelines and eligibility criteria need to be gone over with a fine-tooth comb. Read the notes from the funding bodies as carefully as possible and check out the success rates to see exactly what your chances are before you even begin writing. Make sure you know exactly who you are applying to and what their mandate is. You may need to tailor your research aims to fit the call but remember that if successful you’ll have to deliver, so your aims must be genuinely achievable. You might have to consider drafting in a collaborator who has a strong track record in a particular field to strengthen your proposal. I would also advise getting in contact with researchers who have written successful grant applications (sometimes they are listed online or you might know somebody who has been successful). If you want to see some examples of high quality proposals, check out the NIH website where you can download some successful R01 grant applications. These are high-quality applications and a good benchmark for any proposal.
Importantly, don’t forget that there are no marks for effort if you are unsuccessful. Aside from grant writing you need to be generating quality research and clocking up the publications. You can’t spend all your time writing grants it’s a question of balance. For what it’s worth, I suggest using grant writing as a vehicle to develop your ideas and plan your research. It’s amazing how great an idea can seem until you put it down on paper. Writing really helps crystallise your thoughts.
Successful grant applications
While grant applications may seem a lottery at times, don’t forget that chance favours only the prepared mind. This is as true of funding applications as it is of science. Although grant writing takes time and includes elements which are not purely scientific, it is probably the best chance you have of taking control of your career and driving your research in the direction you want.
The programme (http://www.grc.org/programs.aspx?id=11170) covers a wide a range of exciting subjects such gene regulatory networks, mechanics, stem cells, regeneration, organoids, and evolution. The conference will be preceded by a two-day symposium designed exclusively for students and post-docs.
As some readers will already be aware, we have recently introduced a new ‘format-free’ submission policy. We’ve been delighted by the early feedback on this – from what we’re hearing, this has been a popular move and will help make life easier for authors submitting to Development. But what do we mean by format-free and how does it differ from our earlier policy?
Now, when you submit your paper to Development, you don’t need to worry about specific formatting requirements for the journal – we don’t care if your references are not in Development style (they can even be numbered), whether your materials and methods section comes after the introduction or the discussion, or whether you’ve provided the figures in the format that we need for final publication. We hope this should make things easier for all authors, but particularly for those submitting to Development after their paper has been considered elsewhere; while we like to believe that all our authors select Development as their first choice of venue for publication of their work, we are realistic and recognise that at least a few of you might already have tried another journal first! In general, we don’t see the value in asking you to reformat (or just format) your paper in our house style before you know whether it is likely to be accepted for publication with us. Instead, we want to remove as many of the hurdles to submission as we can and make the whole process as quick and smooth as possible.
So what do we need at initial submission? The most important requirement we still retain, and one that we recognise will not be universally popular, is a length restriction. Research articles over 7000 words and Research reports over 3000 words (excluding title page, abstract, reference list and, now, materials and methods – more on which below) will be returned to authors with a request to shorten the paper to within this limit. We make this a requirement because we believe that length limits serve a valuable purpose – to ensure that a paper remains relatively concise and accessible to the reader. And we have chosen to enforce this guideline at initial submission because, in our experience, papers tend only to get longer during the revision process, meaning that it will become even more difficult to meet these limits at a later stage in the process. In exceptional cases, and following consultation with the handling editor, we may be able to consider papers that exceed this length, but we generally believe that it should be possible to write your paper in a way that does not run over this limit – and that this will make the paper a better read upon eventual publication.
We will also return your paper before sending it to the editor if text or figures are unreadable following conversion to PDF (although this is rare), and we may also ask you for a smaller PDF if the file is too large to be easily handled by editors and referees. In addition, we may have to delay assigning your paper to an editor if we can’t confirm the identity of your co-authors. You might be aware that a few journals (fortunately not us) have encountered problems with corresponding authors submitting papers with fake email addresses for their co-authors, allowing them to circumvent the normal checks that ensure that all co-authors are aware of and approve the paper and its submission. Therefore, where non-institutional (e.g. Gmail) addresses are provided, we will query these with the corresponding author and request either institutional email addresses and/or ORCID IDs. We are sure you understand that it is important we make sure all authors are kept fully informed of the status of their work, and hence why this is an essential check to keep at first submission.
With these changes, we hope to make initial submission to Development as easy as we can. In fact, we were already operating on a largely format-free basis before the announcement of this policy, but we have further relaxed our guidelines with this latest set of changes. We will, however, ask that you ensure your paper complies with our formatting guidelines at revision stage – should your work meet with positive assessment from our editors and referees. At this point, we will also require you to fill in our submission checklist – confirming that your paper complies with various policies and best practise guidelines – to provide high-resolution versions of the figures that our graphics team can process for publication, and to tell us about your funding bodies. Given that, according to recent statistics, we accept over 95% of papers where we have invited a revision, we hope that you won’t mind taking the extra time to format your paper at this stage, when you know the chances of eventual acceptance are very high.
The other significant change we have made, as alluded to above, is to remove the materials and methods section from our word count. The aim here is twofold. First, we want to give you a little more flexibility with article length – the total word limit remains the same even though we now exclude the materials and methods. Second, and more importantly, we recognise the importance of this section of the paper and want to encourage authors to provide appropriate details of all experimental protocols. Length limits often mean that methods sections simply cite previous papers, which cite even earlier papers, so that a reader can find it impossible to figure out how an experiment has been conducted. We would prefer that methods be provided in greater detail, allowing readers to fully understand the protocols. Where materials and methods are particularly lengthy, we will still encourage some of this information – additional details that are primarily of interest to the real expert in the field or to those wishing to replicate the experiments – to be provided in the supplementary information, but again we will not enforce this at initial submission and can work with the authors to make appropriate changes at revision stages.
Together, we hope these changes will make the submission process for authors – whether you are submitting to Development as first choice (which of course we hope most of you do!), or have already been elsewhere – a quicker and easier process. As always, we will continue to review these policies as we go forwards, and we welcome your feedback.
Here are the highlights from the current issue of Development:
Getting MAD in meiosis
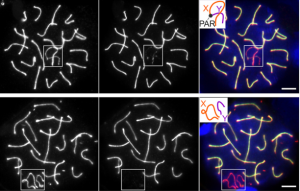
In meiosis I, homologous chromosomes must pair and form crossovers to ensure appropriate chromosome alignment and segregation. During this process, as in mitosis, the spindle assembly checkpoint (SAC) functions to detect misaligned chromosomes at metaphase. This delays anaphase to enable error correction and induces apoptosis when errors cannot be corrected. Surprisingly, previous studies have suggested that the core SAC component MAD2 might be dispensable during spermatogenesis in mice, but Imrul Faisal and Liisa Kauppi now re-investigate the role of MAD2 in male meiosis (p. 1988). Using mouse models in which either all chromosomes (Mlh1 mutants) or just the sex chromosomes (Spo11β-only mice) show defective crossover formation, the authors look at the consequences of Mad2 heterozygosity on spermatocyte apoptosis and aneuploidy. While Mad2 heterozygosity does not rescue sterility of Mlh1 mutants, suggesting that the checkpoint is still active in this context, it does partially rescue apoptosis in the Spo11β-only mouse, resulting in low-level sperm aneuploidy. These data provide the first evidence that MAD2 is important for efficient SAC activation in spermatocytes, and suggest that cells with relatively mild chromosome crossover defects are more sensitive to MAD2 levels than those with severe defects.
Growing a labyrinth with G9a
Defects in placental growth and patterning can have severe consequences for foetal health, and can cause intrauterine growth restriction (IUGR). However, relatively little is understood about the mechanisms regulating placental development, particularly the later phase of maturation when the blood vessels of the so-called labyrinth elongate and elaborate. On p. 1976, Paul Delgado-Olguin and colleagues show that the histone methyltransferase G9a is required for placental maturation. Endothelial-specific knockout of G9a in mice has no effect on early placental development, but mutants show severe defects in labyrinth size and structure after mid-gestation, owing to reduced proliferation of endothelial cells. Intriguingly, the authors provide evidence for non-autonomous regulation of trophoblast cell proliferation, which is upregulated in the endothelial-specific knockout. G9a conditional mutants show reduced expression of Notch pathway effectors (previously implicated in regulation of placental maturation), and the placental vessel phenotype can be rescued by activation of the Notch pathway. Thus, G9a is a key regulator of placental maturation in mice, regulating the balance of endothelial versus trophoblast proliferation. Notably, this mechanism may also apply in human, since G9a and Notch pathway components show altered expression in samples from IUGR pregnancies.
Wheat domestication: Q is the answer
Domestication of wheat has involved a number of phenotypic changes from wild isolates. Notably, domesticated varieties possess a subcompact spike and a loss of the tough glumes that protect the grain, leading to a free-threshing phenotype. Several genetic determinants of these characteristics have been identified, including the Q gene, which encodes an AP2-like transcription factor. Cultivated wheats generally have the hypermorphic Q allele, whereas wild varieties have the q variant, associated with lower AP2 activity. It is known that AP2 factors can be regulated by the miR172 miRNA, and that Q bears a mutation in the miR172 binding site, but how this putative regulation affects the phenotypes associated with domestication has been unclear.
Two papers in this issue of Development address the regulation of Q by miR172. Steve Swain and colleagues (p. 1959) isolate a new allele of Q, Q’, which results in higher protein abundance due to impaired miR172-mediated targeting. Through analysis of Q’ and induced revertants, the authors show that higher levels of AP2 activity are associated with the formation of ectopic florets in place of glumes – an apparent homeotic transformation in the spike. Similar results are also presented by Jorge Dubcovsky and co-workers (p. 1966), who further demonstrate that reduced miR172-mediated degradation of Q is largely responsible for the free-threshing and other phenotypes associated with domesticated varieties. The relative levels of miR172 and Q define spikelet morphology, with higher Q or lower miR172 activity being associated with glume-to-floret transition and free-threshing character, and lower Q activity with the opposite changes. Moreover, a gradient of miR172:Q levels along the spike is associated with a gradient of homeotic changes.
Together, these two papers convincingly demonstrate that tight regulation of Q by miR172 is important for the acquisition of free-threshing character in domesticated wheat varieties, and help to resolve prior controversies as to the mechanism underlying the Q phenotype. These studies also add to our understanding of how AP2 factors regulate floral patterning in plants.
Video Highlight
On p2070, Brant Weinstein and colleagues describe generation of a zebrafish transgenic line expressing GFP in lymphatic vessels allows visualization of the developing lymphatic network, demonstrating a stereotyped, stepwise assembly.
Hiroshi Hamada, director of the RIKEN Center for Developmental Biology, talks about his career in Japan and North America, his fascination with left-right axis determination and his love of Irish music.
This Review discusses the diverse epithelial cell behaviours involved in small neurosensory organ development, using dental placodes, hair follicles, taste buds, lung neuroendocrine cells and lateral line neuromasts as examples.


 o tackle this question, two PhD students, Liana Manukyan (computer scientist) and Sophie Montandon (developmental biologist), followed individual lizards during three to four years of their development from hatchlings crawling out of the egg to fully mature animals. For multiple time points, they reconstructed the geometry and colour of the network of scales on three animals by using R2OBBIE-3D (YouTube playlist), a very high resolution robotic system(16) developed previously in the Milinkovitch laboratory by a physicist PhD student: Antonio Martins.
o tackle this question, two PhD students, Liana Manukyan (computer scientist) and Sophie Montandon (developmental biologist), followed individual lizards during three to four years of their development from hatchlings crawling out of the egg to fully mature animals. For multiple time points, they reconstructed the geometry and colour of the network of scales on three animals by using R2OBBIE-3D (YouTube playlist), a very high resolution robotic system(16) developed previously in the Milinkovitch laboratory by a physicist PhD student: Antonio Martins.

 (3 votes)
(3 votes)


 (No Ratings Yet)
(No Ratings Yet)



 Surprisingly, previous studies have suggested that the core SAC component MAD2 might be dispensable during spermatogenesis in mice, but Imrul Faisal and Liisa Kauppi now re-investigate the role of MAD2 in male meiosis (p.
Surprisingly, previous studies have suggested that the core SAC component MAD2 might be dispensable during spermatogenesis in mice, but Imrul Faisal and Liisa Kauppi now re-investigate the role of MAD2 in male meiosis (p.  On p.
On p.  Several genetic determinants of these characteristics have been identified, including the Q gene, which encodes an AP2-like transcription factor. Cultivated wheats generally have the hypermorphic Q allele, whereas wild varieties have the q variant, associated with lower AP2 activity. It is known that AP2 factors can be regulated by the miR172 miRNA, and that Q bears a mutation in the miR172 binding site, but how this putative regulation affects the phenotypes associated with domestication has been unclear.
Several genetic determinants of these characteristics have been identified, including the Q gene, which encodes an AP2-like transcription factor. Cultivated wheats generally have the hypermorphic Q allele, whereas wild varieties have the q variant, associated with lower AP2 activity. It is known that AP2 factors can be regulated by the miR172 miRNA, and that Q bears a mutation in the miR172 binding site, but how this putative regulation affects the phenotypes associated with domestication has been unclear.