Hanieh Falahati, Lewis–Sigler Institute for Integrative Genomics, Princeton University.
Eric Wieschaus, Howard Hughes Medical Institute, Department of Molecular Biology, Princeton University.
The whole universe is moving toward disorder; this is the second law of thermodynamics in simple terms. Yet, living organisms have found a way to keep themselves organized by spending energy. Cells need this organization in order to provide specialized microenvironments for different cellular functions. Two types of intracellular organizations can be distinguished in biological systems: The first type is membrane-bound organelles such as ER, or lysosome. The composition of these organelles is maintain by spending energy and active transport of molecules across a membrane. The second type of intracellular organization comes from membrane-less organelles such as nucleoli, histone locus bodies, and stress granules that are high-concentration assemblies of different proteins and RNAs. A puzzling question is that without a membrane, how are cells able to form and maintain these organelles.
A historical perspective
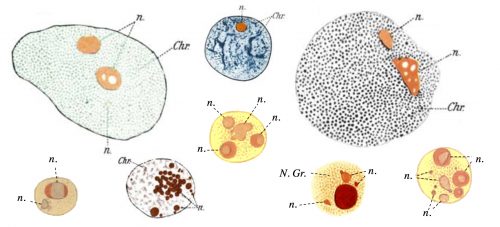
The question of how membrane-less organelles form immediately captured my attention the first time I heard about it in an introductory meeting with my doctoral advisor, Prof. Eric Wieschaus. Of course this question has fascinated many scientists since the initial observation of the nucleolus, the quintessential membrane-less organelle, in the 18th century. In 1898, Montgomery performed a very comprehensive study of the nucleoli of different cell types, hand-drawn in 346 figures (a sample is shown in Fig. 1), and concluded that the nucleolus forms when a substance from the cytoplasm enters the nucleus and is deposited there “in the form of masses of varying dimensions, which may be either globular or irregular in shape, according as they are fluid or viscid in consistency”. He further describes that nucleoli form via “coalescence of numerous small portions of nucleolar substance”, consistent with its fluidity (Montgomery 1898). However, this model faded away with the advancement in cell biology and genetics which showed that the nucleolus forms around ribosomal DNA repeats and is the site of active transcription and processing of ribosomal RNA, and ribosomal biogenesis. By the end of the 20th century, nucleolus was commonly thought to form actively, in the process of making ribosomes, until a number of influential works from Hyman and Rosen groups redirected the focus to the liquid nature of the membrane-less organelles (Brangwynne et al. 2009; Li et al. 2012). Based on this liquid property, it was proposed that formation of membrane-less organelles is a spontaneous liquid-liquid phase separation (LLPS), similar to the separation of oil from water.
Figure 1. Hand drawing of nuclei with nucleoli (n) of different cell types drawn by Montgomery in 1898. N. Gr.: nuclear granule. Chr.: Chromatin.
By the time I joined the Wieschaus lab at Princeton, two main hypotheses existed for the formation of membrane-less organelles: 1. The LLPS model, in which the components of these organelles assemble spontaneously in a thermodynamically-driven process due to their favorable intermolecular interactions, and form a liquid phase; 2. The active assembly model which suggests that these organelles form as a result of an active process, which happens inside the cells or inside the organelles. Such active processes are reactions carried out by enzymes that couple these reactions to an energy source such as ATP. The evidence for the LLPS model comes from experiments showing that many of the proteins which localize to the membrane-less organelles have the physical properties to phase separate in vitro. However, in most cases the phase separation is only observed at concentrations much higher than cellular levels. Therefore, a pressing question is whether the physical properties of these components would also drive their spontaneous assembly in vivo, or active processes are responsible for their congregation. Accordingly, we decided to develop an in vivo test that would allow us to distinguish between the LLPS and the active mechanism, and apply it to study nucleolus assembly as a model for the formation of membrane-less organelles.
Working out a path – How to test the phase separation model in vivo?
Developing such an in vivo test was not trivial, since compared to in vitro systems, living cells are far more limited in the ways they can be manipulated. But considering the expertise of my advisor, Eric Wieschaus, in manipulating Drosophila embryos, his lab was the best place to do this type of research. It was not the only reason for us to study this question in fly embryos: Development in Drosophila starts without a nucleolus, and the nucleolus forms at a particular developmental stage (Video 1). This allows us to manipulate the system and study its effect on the nucleolus assembly. In addition, nucleolus forms during the 13 synchronous divisions at the beginning of development, therefore at each time-point there are multiple nuclei at the exact same developmental stage, which helps with statistics. Finally, during these 13 division the nuclei share the cytoplasm, which means that all the nuclei have access to the same concentration of most molecules. These features make fly embryos an ideal system for studying nucleolus assembly.
Video 1. Nuclei of D. melanogaster embryos start development without a nucleolus, and form the nucleolus (bright foci) for the first time at nuclear cycle 13.
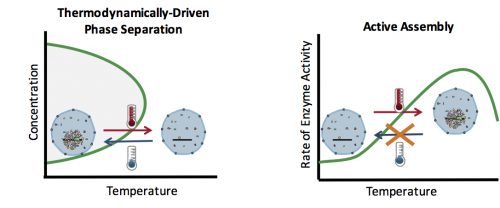
How can you test whether the formation of an organelle in vivo is a thermodynamically driven LLPS, or an active assembly process? In general, LLPS processes are affected by two factors: concentration and temperature (Fig. 2). Increasing the concentration results in an increase in the size of assemblies: The more oil you add to the water, the bigger the oil droplets. However, active processes are also affected similarly with concentration: The higher the concentration of the reactants, the more products (assemblies) are formed. Therefore, changes in the concentration cannot distinguish between an LLPS and an active assembly. On the contrary, temperature often has a differential effect on these two processes: In general, lowering the temperature causes more condensation and enhances phase separation, but slows down active enzymatic reactions. This means that by quantifying the effect of various temperatures on the assembly formation, one can distinguish between an LLPS and an active assembly model.
Figure 2. (Left) Phase separation processes are affected by concentration and temperature. Increasing the concentration and decreasing the temperature results in the formation of larger assemblies. (Right) Active processes are carried out by enzymes which couple them to an energy source. Therefore, the rate of these processes is determined by the rate of enzyme activity, which is reduced at lower temperatures.
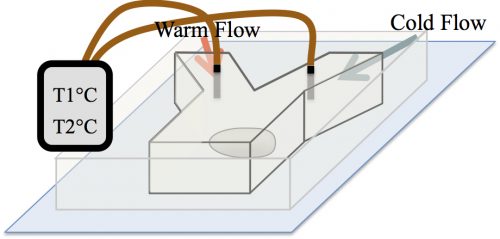
At the time, our strategy to examine the effect of temperature on the formation of assemblies was received with skepticism among the community. The main concern was whether in the temperature range tolerable by biological systems, an effect would be detectable. A reassurance, however, was an exquisite work by Sarah Veatch, who showed that plasma membrane vesicles isolated from living cells exhibit phase transitions between 15 and 25°C (Veatch et al. 2008). Subsequently, we employed a microfluidic device that allowed us to control the temperature of a fly embryo between 6 and 31°C (Fig. 3). We used this microfluidic device to examine the effect of temperature on the first time formation of assemblies, and also to test the reversibility of assembly formation, for six different nucleolar proteins.
Figure 3. A microfluidic device was used to control the temperature of an embryo in a range of 6 to 31°C by flowing in warm or cold water.
The unexpected result of two independent mechanisms
Much to our surprise, the results of our test showed that two independent mechanisms govern the assembly of different subsets of nucleolar proteins: While four of the studied proteins depict properties of LLPS, the two others are recruited actively. At lower temperatures, the phase separating proteins assemble at an earlier developmental time, and dissolve upon heating (Video 2). The latter is consistent with the reversibility of thermodynamic processes. On the contrary, the formation of active proteins is inhibited at low temperatures, and is irreversible, an exclusive property for active processes.
Video 2. The nuclei of intact fly embryos are subjected to temperature changes in the surrounding fluid. As the temperature is shifted from low to high, the phase separated proteins dissolve, as can be seen in the disappearance of the bright spots.
rDNA: Come together, right now, over me
Our unexpected result that two independent mechanisms govern nucleolus assembly in vivo raised a new question: How are these two different mechanisms coordinated to form a single organelle? In a previous paper, we had shown that transcription of rDNA dictates the spatiotemporal precision in the formation of assemblies by the phase separating proteins (Falahati et al. 2016). Interestingly, we were also able to show that rDNA is necessary for the recruitment of the two actively assembling proteins. This suggests that rDNA can function as a coordinator between the two independent mechanism. In addition, it highlights the fact that in vivo, even the assembly of phase separating proteins is regulated by active processes such as transcription.
We are very enthusiastic about the publication of this paper as it is a significant advancement in the field both from a methodological and mechanistic standpoint. We took an interdisciplinary approach and developed an invaluable tool for closing the gap between the current knowledge of the in vitro self-assembly and the formation of membrane-less organelles in vivo at its full complexity. From a mechanistic perspective, the large number of proteins studied here allowed us to unravel the unappreciated complexity in the formation of intracellular assemblies, as our results show the presence of at least two independent mechanisms for the recruitment of nucleolar proteins. We are confident that the method and the results introduced in this paper will set a framework to better understand how normal or pathological intracellular assemblies form.
Our latest monthly trawl for developmental biology (and other cool) preprints. See June’s introductory post for background, and let us know if we missed anything
Another month, another bumper crop of preprints covering everything from calcium waves in the fly to the first mutant ant line, plant superhero genes to rat embryonic stem cells. Plus, as usual, a whole bunch of tools and resources (including a 3D printed micropipete!), and right at the end some enigmatic bacterial structures.
The preprints were hosted predominantly on bioRxiv, though we also found a handful on arXiv and PeerJ.
Multiple roles of PIWIL1 in mouse neocorticogenesis. Barbara Viljetic, Liyang Diao, Jixia Liu, Zeljka Krsnik, Sagara H. R. Wijeratne, Ryan Kristopovich, Marina Dutre-Clarke, Matthew L. Kraushar, Jimin Song, Jinchuan Xing, Kevin Chen, Mladen-Roko Rasin
Laser ablation tomography image of a nodal root from a maize plant, from Bucksch, et al.
Morphological plant modeling: Unleashing geometric and topological potential within the plant sciences. Alexander Bucksch, Acheampong Atta-Boateng, Akomian Fortune Azihou, Mathilde Balduzzi, Dorjsuren Battogtokh, Aly Baumgartner, Brad Binder, Siobhan Braybrook, Cynthia Chang, Viktoriya Coneva, Thomas DeWitt, Alexander Fletcher, Malia Gehan, Diego Hernan Diaz Martinez, Lilan Hong, Anjali Iyer-Pascuzzi, Laura Klein, Samuel Leiboff, Mao Li, Jonathan Lynch, Alexis Maizel, Julin Maloof, RJ Cody Markelz, Ciera Martinez, Laura Miller, Washington Mio, Wojtek Palubicki, Hendrik Poorter, Christophe Pradal, Charles Price, Eetu Puttonen, John Reese, Ruben Rellan-Alvarez, Edgar Spalding, Erin Sparks, Chris Topp, Joseph Williams, Daniel Chitwood
Tex19.1 Restricts LINE-1 Mobilisation in Mouse Embryonic Stem Cells. Marie MacLennan, Marta Garcia-Canadas, Judith Reichmann, Elena Khazina, Carmen Salvador-Palomeque, Abigail Mann, Paula Peressini, Laura Sanchez, Christopher J Playfoot, David Read, Chao-Chun Hung, Ragnhild Eskeland, Richard R Meehan, Oliver Weichenrieder, Jose Luis Garcia- Perez, Ian R Adams
Sagittal sections of mandibular molars from Feng, et al.
Functional Annotation of Chemical Libraries across Diverse Biological Processes. Jeff S Piotrowski, Sheena C Li, Raamesh Deshpande, Scott W Simpkins, Justin Nelson, Yoko Yashiroda, Jacqueline Barber, Hamid Safizadeh, Erin Wilson, Hiroki Okada, Abraham Gebre, Karen Kubo, Nikko Torres, Marissa LeBlanc, Kerry Andrusiak, Reika Okamoto, Mami Yoshimura, Eva DeRango-Adem, Jolanda van Leeuwen, Katsuhiko Shirahige, Anastasia Baryshnikova, Grant W Brown, Hiroyuki Hirano, Michael Costanzo, Brenda Andrews, Yoshikazu Ohya, Hiroyuki Osada, Minoru Yoshida, Chad L Myers, Charles Boone
TOWARDS COORDINATED INTERNATIONAL SUPPORT OF CORE DATA RESOURCES FOR THE LIFE SCIENCES. Warwick Anderson, Rolf Apweiler, Alex Bateman, Guntram A Bauer, Helen Berman, Judith A Blake, Niklas Blomberg, Stephen K Burley, Guy Cochrane, Valentina Di Francesco, Tim Donohue, Christine Durinx, Alfred Game, Eric Green, Takashi Gojobori, Peter Goodhand, Ada Hamosh, Henning Hermjakob, Minoru Kanehisa, Robert Kiley, Johanna McEntyre, Rowan McKibbin, Satoru Miyano, Barbara Pauly, Norbert Perrimon, Mark A Ragan, Geoffrey Richards, Yik-Ying Teo, Monte Westerfield, Eric Westhof, Paul F Lasko
Uncharacterized bacterial structures revealed by electron cryotomography. Megan J Dobro, Catherine M Oikonomou, Aidan Piper, John Cohen, Kylie Guo, Taylor Jensen, Jahan Tadayon, Joseph Donermeyer, Yeram Park, Benjamin A Solis, Andreas Kjaer, Andrew I Jewett, Alasdair W McDowall, Songye Chen, Yi-Wei Chang, Jian Shi, Poorna Subramanian, Cristina V Iancu, Zhuo Li, Ariane Briegel, Elitza I Tocheva, Martin Pilhofer, Grant J Jensen
A postdoctoral research fellow position is available in the laboratory of Gregor Andelfinger at the CHU Sainte Justine Research Center, Montréal, Québec, Canada. Our work focuses on the genetic origins of cardiovascular diseases in the young and includes both genomics and functional approaches. In this project, the successful candidate will apply genomic and molecular biology approaches to develop disease models. Postdoctoral fellows will have a unique opportunity to do interdisciplinary research at all interfaces of translational medicine and publish their results in high impact journals.
The candidate will work in collaboration with all lab members and is expected to independently perform human and mouse genetic studies, as well as in vitro molecular analyses. Experience with a wide variety of techniques (developmental studies, histology, confocal microscopy, cell culture assays, RNAseq) is desired. Experience with genome editing techniques (CRISPR) and/or and bioinformatics analysis is an asset.
The successful candidate must have a recent Ph.D. degree in life sciences, molecular cell biology, or a related field. We are seeking highly motivated and well published researchers with excellent oral, written, and interpersonal communication skills.
Montreal is a vibrant bilingual city with a high quality of life. With a greater metropolitan area of 3.5 M, Montreal offers all scientific and cultural benefits of a large North American agglomeration with a European flair and beautiful surroundings.
The Andrew Yoo lab at WUSTL is looking for a lab manager/technician. The Yoo lab uses microRNA-mediated direct reprogramming of fibroblasts to neurons to study aging and neurodegenerative diseases.
PhD position opened in Switzerland for a developmental or cell biologist
Development & Evolution of structural colours in vertebrates
In the context of a multidisciplinary study combining cell biology, evolutionary developmental biology and physics, we offer one PhD student position for an outstanding, highly motivated, and creative experimental wet-lab biologist with strong skills in developmental biology or cell biology.
A major research project in Michel Milinkovitch’s group (Laboratory of Artificial & Natural Evolution; www.lanevol.org) at the University of Geneva (UNIGE) is to better understand the molecular developmental mechanisms generating a diversity of skin colours and patterns in reptiles.
One aim is to understand the development & evolution of surface gratings and intracellular nano-structures generating colours through the physical phenomenon of light interference. The successful candidates will use state-of-the-art microscopy and biochemical/genetic manipulations in both the zebrafish and chameleons to understand the development and evolution of surface gratings and intracellular photonic crystals.
Candidates must have a Master in biology or biochemistry. Skills and experience with developmental biology and/or cell biology are mandatory. The successful candidate will have a genuine interest for organismal biology and will appreciate interactions with developmental biologists, evolutionary biologists, physicists and computer scientists. The PhD student will be localised in the Milinkovitch lab (http://www.lanevol.org) and will have very regular interactions/collaborations with members of Marcos Gonzalez-Gaitan’ lab (https://cms.unige.ch/sciences/biochimie/-Marcos-Gonzalez-Gaitan-Lab-.html).
The University of Geneva is world-renowned for its research in Biology and Physics and is among the top 1% best universities in the world.
PhD students are remunerated according to the standards of UNIGE, which are very generous when compared to other international programs.
Geneva is an international city occupying a privileged geographical situation with its beautiful lake and the close-by Alps.
Refs: Saenko et al.Precise colocalization of interacting structural and pigmentary elements generates extensive color pattern variation in Phelsuma lizards.BMC Biology 2013, 11: 105; Teyssier et al.Photonic Crystals Cause Active Colour Change in Chameleons.Nature Communications 6: 6368 (2015); Tzika et al. Reptilian Transcriptomes v2.0: An Extensive Resource for Sauropsida Genomics and Transcriptomics. Genome Biol. Evol. 7: 1827-1841 (2015); Ullate-Agote et al.The genome sequence of the corn snake (Pantherophis guttatus), a valuable resource for EvoDevo studies in squamates.Int. J. Dev. Biol. 58: 881-888 (2014); Saenko et al.Amelanism in the corn snake is associated with the insertion of an LTR-retrotransposon in the OCA2 gene.Scientific Reports 5, 17118 (2015); Di-Poï & Milinkovitch. The Anatomical Placode in Reptile Scale Morphogenesis Indicates Shared Ancestry Among Skin Appendages in Amniotes. Science Advances 2, e1600708 (2016).
Candidates must send their application — in the form of a single PDF file including a brief letter of interest, a CV, as well as contact information (not support letters) of two persons of reference — to: Prof. Michel Milinkovitch (michel.Milinkovitch@unige.ch).
We are looking for a postdoc interested in studying neural development in tunicates, the sister group to the vertebrates. The larval nervous system of the tunicate Ciona has only 177 neurons and its entire “connectome” has been recently mapped (Ryan et al., eLife 2016). The Ciona genome is also highly compact and easily manipulated using CRISPR/Cas9 (Stolfi et al., Development 2014). This tractability offers an unprecedented opportunity to understand gene networks underlying the development of every single neuron in a chordate nervous system.
We have several potential projects centered on understanding the link between transcriptional regulation and cell behavior during the development of the Ciona nervous system. Specific cells of interest include homologs of vertebrate spinal cord neurons (Stolfi et al., Development 2011) and neural crest-derived sensory neurons (Stolfi et al., Nature 2015). Projects will involve isolating neural progenitors from transgenic Ciona embryos, profiling their transcriptome and chromatin states using next-generation sequencing, and targeting candidate regulators and effectors using CRISPR/Cas9.
Our lab is located at the Georgia Institute of Technology in Atlanta, USA. Georgia Tech offers first-rate research facilities and a highly collaborative research environment with traditional strengths in engineering, computing, and math. We are dedicated to promoting equity of under-represented minorities in academia.
Position highlights: The Grant Strategist will support the productivity of Zuckerman Institute scientists by editing and consulting on grants, particularly sections on research strategy, aims, and narratives; acquire and maintain sound knowledge of the Institute and the research priorities; use that knowledge and understanding to seek and recommend grants for individual as well as multi-investigator grants within and between disciplines.
The Grant Strategist will serve as a project manager to pursue, identify and promote funding opportunities for multi-PI grants. The Grant Strategist will also consult with and keep abreast of the current granting environment to provide suggestions to individual researchers on specific grant opportunities.
Position requirements: Ph.D. in the biological sciences or related field, including extensive writing experience; or advanced degree in journalism with deep expertise in science. Minimum of four (4) years of successful science grant writing experience or related experience in science communication field is required. Extensive knowledge of scientific granting processes, particularly in the biological sciences; successful grant writing or consulting record is vital.
When blood goes bad, a replacement is often needed. Each year, thousands of patients in the US receive bone marrow transplants to treat life-threatening diseases like blood cancer.
But in some cases, the transplant itself can become deadly.
The problem is not necessarily the one most people think of: rejection of the transplanted material. Instead, the donor hematopoietic stem cells (HSCs), delivered to repopulate the whole blood system, sometimes produce immune cells that attack anything foreign. In these cases, the transplant rejects the patient. This condition, called graft-versus-host disease (GVDH), is common in transplant patients and often fatal.
Developing a reliable method to make transplantable stem cells that would avoid GVHD is of considerable interest. That’s why we and others have been interested in learning how to direct the differentiation of HSCs in vitro from pluripotent precursor cells.
Intuitively, a way to make new HSCs would be to replicate the same instructions used to make them in the developing embryo. But first we have to identify the signals that make up those instructions. During embryonic development, HSCs are born from the descending aorta, the primitive vessel that carries blood to the lower half of the body. Decades of research have uncovered numerous signaling inputs that direct formation of the descending aorta, and its subsequent conversion to the first HSCs. However, we are still defining the complete set of signals, and how they are regulated and integrated.
We set out to examine this coordination and identify novel factors that might play critical roles in HSC development. We used zebrafish because their blood development is nearly identical to that of humans. We noticed a common theme: signaling pathways required for hematopoietic stem cell formation also frequently regulate vessel patterning. Specifically, well-established signaling pathways, like the Vegf, Notch, and Wnt pathways, play a role in both processes.
In the trunk of the embryo, smaller blood vessels called “intersegmental vessels” sprout from the developing descending aorta at about the same time that HSCs are born. We became intrigued by published results identifying a new signaling pathway necessary for the sprouting of intersegmental vessels from the aorta (Gore et al., 2011). This pathway involves R-spondin-1 (Rspo1), a secreted factor that augments Wnt signaling. Intriguingly, Gore et al. also showed rspo1 was expressed in the dorsal aorta during the time of HSC specification, the point when blood stem cells start to assume their future identity. We wondered if this pathway might also be necessary for HSC specification.
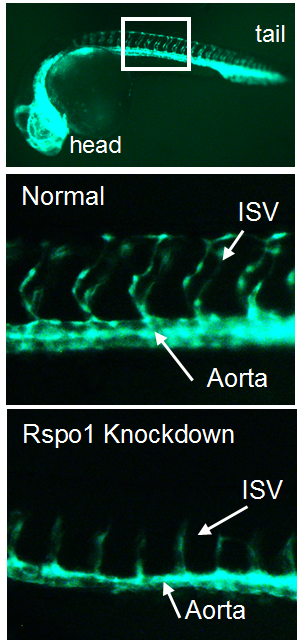
Figure 1. Transgenic zebrafish with GFP-labelled blood vessels. Rspo1 knockdown causes loss of blood vessel growth as seen by decreased intersegmental vessel (ISV) growth, providing a visual means of seeing that Rspo1 is inactivated.
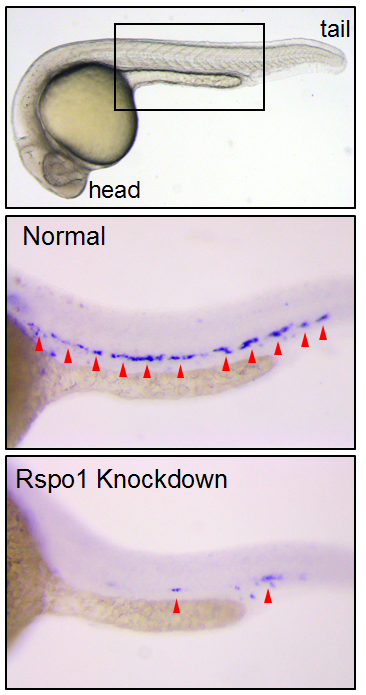
Our first goal was to determine if Rspo1 is required in HSC development. We knocked down Rspo1 in zebrafish embryos using antisense oligonucleotides and asked what would happen to HSCs. Strikingly, markers of HSC development were visibly decreased at all time points we examined. We subsequently confirmed the rspo1 requirement in an established zebrafish hypomorphic mutant.
Figure 2. HSCs marked by runx1 in 24 hours post fertilization zebrafish embryos. Rspo1 knockdown causes loss of HSCs. Red arrowheads indicate HSCs.
So Rspo1 is required for HSCs to form, but how does it work? To answer this question, we looked at the activity of other pathways already known to play a role in HSC specification to see if we could detect any changes in their activity. Our results identified two of these – the Wnt16 and Vegfa pathways – that showed alterations in expression of their downstream components.
Our findings had some surprises. Most notably, although vegfa expression was lower when Rspo1 was knocked down, a key target of Vegf signaling, notch1b, was unaffected. Our finding suggested that there might be a Notch1-independent role for Vegf in specifying hematopoietic stem cells.
As we were trying to define how this might work, the group of Roger Patient (Monteiro et al., 2016) identified precisely such a pathway. We determined that Rspo1 acted via this Notch-independent pathway, and discovered that a particular Vegfa splice variant was involved.
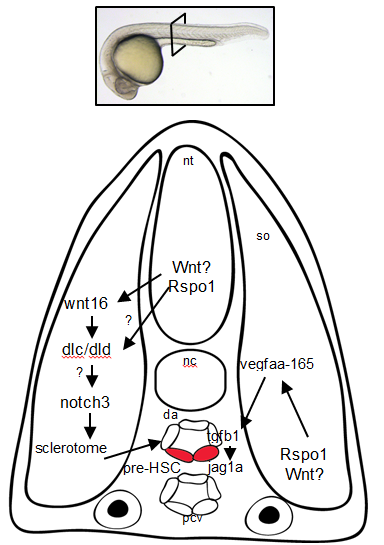
Figure 3. Rspo1 signaling pathways needed for the development of HSCs. Cross-section of a 24hr old zebrafish embryo.
Overall, these findings point to Rspo1 as a new master regulator of blood stem cells that controls two pathways both needed for the birth of these stem cells. In the future, we hope that understanding the full set of signals and their integration will provide the key to unlocking our ability to make fully functional hematopoietic stem cells in the lab. Eventually we will have the tools to create designer treatments for leukemia and other blood disorders while making GVHD a thing of the past.
Here are the highlights from the new issue of Development:
Adding a new layer of complexity to pre-eclampsia
Pre-eclampsia (PE) is a pregnancy complication associated with abnormal formation of the placenta. To date, most studies of PE have focussed on cytotrophoblasts (CTBs) within the villous placenta (the chorion frondosum); the deficient invasion of these cells into the uterine wall is thought to lead to abnormal placentation and hence PE. Here, on p. 767, Susan Fisher and colleagues reveal that CTBs within a different region of the human placenta – the smooth chorion – are implicated in severe PE. They first report that the CTB layer in the smooth chorion is expanded in severe cases of PE and is less organised. These morphological changes are accompanied by changes in the expression profiles of smooth chorion CTBs. In particular, smooth chorion CTBs exhibit enhanced expression of HLA-G, integrin α4 and E-cadherin, all of which are factors that extravillous CTBs normally modulate as they invade the uterine wall. The researchers further show that severe PE is associated with global gene expression changes in smooth chorion CTBs that are distinct from the transcriptional responses of villous and extravillous CTBs to severe PE, which they previously described. Overall, these findings suggest that smooth chorion CTBs play a greater role in placentation and pregnancy outcome than previously appreciated.
Integrins worm their way into brain regeneration
Tissue regeneration involves a number of cellular processes, including proliferation, differentiation, migration and patterning, but it is not clear how all of these processes are coordinated to allow the correct generation and assembly of cells during regeneration. Using the planarian flatworm Schmidtea mediterranea, which displays remarkable regenerative capacity and can regenerate any missing body part, two new studies reveal a key role for integrins in regulating tissue organisation during brain regeneration.
In the first study (p. 784), Nicolle Bonar and Christian Petersen use RNAi to show that β1-integrin regulates cell number and tissue organisation during regeneration following decapitation; in the absence of β1-integrin, brain tissue is disorganised and ectopic cell aggregates form in the head region. By contrast, global patterning is largely unaffected. The authors further show that β1-integrinRNAi animals exhibit an initial delay in regeneration but that this is followed by tissue overproduction. Finally, they report that integrin signalling, likely via a β1/α-2 complex, is required for the proper localisation of neoblasts and progenitor cells during regeneration. Together, these findings suggest that integrin signalling acts to recruit and localise progenitor cells following injury, thereby promoting the correct organisation of regenerating planarian tissue.
In the second paper (p. 795), Florian Seebeck, Kerstin Bartscherer and colleagues demonstrate that β1-integrin RNAi animals, as well as α-integrin-2 RNAi animals, exhibit impaired regeneration following amputation. They show that, in β1-integrin RNAi animals, the newly formed tissues – including the muscle and gut – display structural defects. The researchers also reveal that β1-integrin is required for neoblast migration towards the wound site. Finally, they report that β1-integrin RNAi causes the formation of ectopic neural spheres within the regenerating brain region that are composed of various neuronal cell types and that undergo continuous growth. Overall, these results suggest that integrins are required for the formation of organised tissues and for restricting neurogenesis during planarian regeneration.
Apoptosis: a delayed gut reaction to bacteria
The intestine is a tissue that is known to undergo regeneration, both continuously as part of tissue homeostasis and in response to damage – for example, that induced by bacterial aggression. While many studies have examined how the gut responds to large amounts of pathogenic or opportunistic bacteria, it is unclear how low levels of bacteria might influence gut homeostasis. Here, on p. 808, Armel Gallet and co-workers tackle this issue. They report that small amounts of the opportunistic Gram-positive bacterium Bacillus thuringiensis var. kurstaki induce a mild early stress response mediated by JNK signalling in the Drosophila midgut. This, in turn, induces the proliferation of intestinal stem cells and leads to the accumulation and overcrowding of differentiated intestinal cells (enterocytes). The authors further report that low amounts of ingested bacteria do not trigger apoptosis, whereas larger amounts do. However, they find that a wave of apoptosis is observed days after infection and acts to eliminate the excess enterocytes. Finally, they demonstrate that the Hippo pathway functions cell-autonomously to trigger the removal of supernumerary enterocytes. These findings lead the authors to propose that the mechanisms involved in the response to the ingestion of low amounts of opportunistic bacteria are different to those mediating the ʻregenerative cell deathʼ that occurs following a stronger aggression.
Nucleogenesis gets active
Neurons within the central nervous system can assemble into clusters, termed nuclei, that house neurons with similar synaptic inputs, outputs and function. This process of nucleogenesis, which is crucial for correct circuit formation, is poorly understood. Now, on p. 830, Sarah Guthrie and colleagues show that the correct assembly of developing motor neurons into nuclei in the chick brainstem requires interplay between spontaneous activity, type II cadherins and gap junctions. Using the genetically encoded calcium indicator GCaMP6, they first show that facial motor neurons exhibit activity patterns that change over the course of nucleogenesis. These patterns can be disrupted by perturbing the expression of the type II cadherin Cad20 or the gap junction protein Cx43. The authors further demonstrate that the inhibition of spontaneous activity (using calcium channel inhibitors) results in neuronal disaggregation, and also causes a reduction in the levels of Cad13, another type II cadherin, suggesting the presence of a feedback loop. In summary, these observations suggest that a network of interactions between cadherins, gap junctions and spontaneous activity governs nucleogenesis.
PLUS:
Creating to understand – developmental biology meets engineering in Paris
In November 2016, developmental biologists, synthetic biologists and engineers gathered in Paris for a meeting called ‘Engineering the embryo’. The participants shared an interest in exploring how synthetic systems can reveal new principles of embryonic development, and how the in vitro manipulation and modeling of development using stem cells can be used to integrate ideas and expertise from physics, developmental biology and tissue engineering. In their Meeting Review, Anna Kicheva andNicolas Rivron provide a summary of this meeting and highlight the challenges arising at the intersection of these fields.
Krüppel-like factors in mammalian stem cells and development
Krüppel-like factors (KLFs) are a family of zinc-finger transcription factors that regulate diverse processes such as cell proliferation, differentiation, development and regeneration. Several KLFs are also crucial for maintaining pluripotency and, hence, have been linked to reprogramming and regenerative medicine approaches. In their Primer, Agnieszka Bialkowska, Vincent Yang andSandeep Mallipattu review key functions for KLFs in mammalian embryogenesis, stem cells and regeneration.
Plasticity in the lung: making and breaking cell identity
In recent years, lineage tracing studies have identified distinct epithelial stem and progenitor cell populations in the lung. These cells, together with their differentiated progeny, maintain a stable identity during steady state conditions, but can display remarkable lineage plasticity following injury. In their Review, Purushothama Tata andJayaraj Rajagopal summarize our current understanding of the different cell lineages of the adult mammalian lung and discuss how these populations respond to injury.
Reviews Editor: biomedical and life sciences journals
Based in Cambridge, UK
Applications are sought for the role of Reviews Editor for The Company of Biologists’ journals: Development, Journal of Cell Science, Journal of Experimental Biology, Disease Models & Mechanisms and Biology Open. The role will be offered for an initial term of three years.
Joining a long-established and successful publishing company, this is an exciting opportunity to make a significant contribution to our highly respected biomedical and life science journals. The role will initially focus on Disease Models & Mechanisms, a growing Open Access journal in the fast-moving field of translational research.
The front sections of the journals include commissioned reviews and poster articles, thought-provoking editorials and interviews with leaders in the field.
Applicants will have a PhD or MD in a relevant area. Post-doctoral and/or previous editorial experience is desirable, although we will provide full training. The successful candidate will work alongside an established publishing team in our Cambridge office.
Core responsibilities include:
• Commissioning, handling peer review and developmental editing of material for the front section of the journals.
• Travelling to international scientific conferences and research institutes, representing the journals, keeping abreast of the latest research and making contacts in the community.
• Spotting newsworthy articles, writing informative press releases and handling any media enquiries.
• Interviewing high-profile scientists.
• Contributing to our social media output.
• Creative involvement in the journals’ development and marketing activities.
Additional responsibilities may be provided for the right candidate.
Essential requirements for the job are enthusiasm, commitment, judgement and integrity. Candidates should have excellent interpersonal skills and confidence, and excellent oral and written communication skills. The successful candidate will have a broad interest in science, the scientific community and publishing.
The position is full time and will be based in The Company of Biologists’ attractive modern offices on the outskirts of Cambridge, UK. It offers an attractive salary and benefits.
The Company of Biologists (biologists.com) exists to support biologists and inspire advances in biology. At the heart of what we do are our five specialist journals –Development, Journal of Cell Science, Journal of Experimental Biology, Disease Models & Mechanisms and Biology Open – two of them fully open access. All are edited by expert researchers in the field, and all articles are subjected to rigorous peer review. We take great pride in the experience of our editorial team and the quality of the work we publish. We believe that the profits from publishing the hard work of biologists should support scientific discovery and help develop future scientists. Our grants help support societies, meetings and individuals. Our workshops and meetings give the opportunity to network and collaborate.
Applicants should send a CV by email to recruitment@biologists.com along with a covering letter that states their current salary and any limitations regards possible start date, and summarises their relevant experience and why they are enthusiastic about this opportunity.
Applicants should be eligible to work in the UK and should be able to travel internationally. Applications should be received by 20th March 2017.
 (2 votes)
(2 votes)



 (No Ratings Yet)
(No Ratings Yet)

 In the first study (p.
In the first study (p. 


 Krüppel-like factors (KLFs) are a family of zinc-finger transcription factors that regulate diverse processes such as cell proliferation, differentiation, development and regeneration. Several KLFs are also crucial for maintaining pluripotency and, hence, have been linked to reprogramming and regenerative medicine approaches. In their
Krüppel-like factors (KLFs) are a family of zinc-finger transcription factors that regulate diverse processes such as cell proliferation, differentiation, development and regeneration. Several KLFs are also crucial for maintaining pluripotency and, hence, have been linked to reprogramming and regenerative medicine approaches. In their  In recent years, lineage tracing studies have identified distinct epithelial stem and progenitor cell populations in the lung. These cells, together with their differentiated progeny, maintain a stable identity during steady state conditions, but can display remarkable lineage plasticity following injury. In their
In recent years, lineage tracing studies have identified distinct epithelial stem and progenitor cell populations in the lung. These cells, together with their differentiated progeny, maintain a stable identity during steady state conditions, but can display remarkable lineage plasticity following injury. In their