Applications are invited for a Research Assistant/Associate within the Division of Infection & Immunity at University College London in Dr Gillian Tomlinson’s laboratory.
We are seeking a highly motivated individual interested in integrating cutting-edge human and zebrafish models to study the immunopathogenesis of tuberculosis. The post is funded by a Medical Research Council Clinician Scientist Fellowship entitled “Tuning the immune response in tuberculosis”, and combines a human experimental tuberculosis challenge model with studies using Mycobacterium marinum infection of zebrafish to identify and validate host factors that calibrate a favourable immune response in tuberculosis.
The post-holder will be supervised by Dr Gillian Tomlinson based in the Cruciform Building at UCL. Dr Tomlinson works within Dr Mahdad Noursadeghi’s and Professor Benny Chain’s group’s which study host immune responses to infectious diseases at genome‑wide level with a particular focus on tuberculosis (www.innate2adaptive.com). The zebrafish work will be supported by the fully managed world class research aquarium at UCL.
The post is available until 1st March 2020, subject to satisfactory probationary and annual appraisals. There is an established track record for department post-doctoral staff gaining personal fellowships. Independently minded and talented investigators will be encouraged and supported in seeking such fellowship support.
Key requirements
Applicants must have an MSc (or equivalent degree) and/or a PhD (or equivalent degree) in a relevant subject.
Candidates must have experience of working with zebrafish including microinjection and in vivo imaging.
Evidence of significant scientific contribution including publications and presentations at conferences is also essential.
Established by the British Society for Developmental Biology (BSDB) in 2014, The Gurdon/The Company of Biologists Summer Studentship scheme provides financial support to allow highly motivated undergraduate students an opportunity to engage in practical research during their summer vacation. Each year, ten successful applicants spend eight weeks in the research laboratories of their choices, and the feedback we receive is outstanding.
Our final report from the 2016 class comes from Christopher Taylor, who undertook his studentship with Karim Sorefan in Sheffield.
A dual hormone response in Arabidopsis thaliana
The population problem
Human population growth is not a new issue. For much of mankind’s history, the species has existed in small, dispersed populations. It is only in the last 100 years where the population has truly boomed with over a fourfold increase in the number of mouths to feed (Fig 1). Demand for food is predicted to increase by 50% by 2050 and already 1 in 7 people are starving, a figure only predicted to increase.
The problems of this unsustainable growth are exacerbated by two issues:
Climate change – in certain areas agricultural productivity is severely reduced by rising temperatures and an increase in extreme events.
Geography – areas with greatest population growth coincide with areas most likely to be impacted by climate change. These include less economically developed countries such as those in the Sub Saharan belt.
Fig 1: The post-industrial ‘population spike’ highlights just how rapidly the human population has boomed in such a short space of time.
Evidently there is an urgent need for a greener revolution.
A solution?
Plants are nature’s biochemical engineers, producing everything they need to grow, survive and reproduce from practically thin air (and a little help from the soil). Plants provide the food we eat, the food our food eats and the food that supports the very ecosystems we depend on. ‘Improving nature’s inefficiencies’ by exploiting plants’ existing biochemical pathways is one way of increasing food production in the finite space we have.
The production, transportation and signalling of plant hormones (phytohormones) allows plants to orchestrate growth and development and importantly for a changing climate, facilitate a response to environmental cues. The classic example of auxin’s role in phototropism may spring to mind. Here auxin exhibits a negatively phototropic response, moving away from the prevailing light direction. This leads to an accumulation of auxin molecules in the side furthest from the light causing acidification of the cell walls and allowing for cell elongation and plant growth. Ultimately this produces a plant (or stem) whose curvature is maximised for intercepting incoming light.
Exploiting these pathways already has agricultural applications. Think of taking a plant cutting, dabbing the base in auxin and growing a whole new plant.
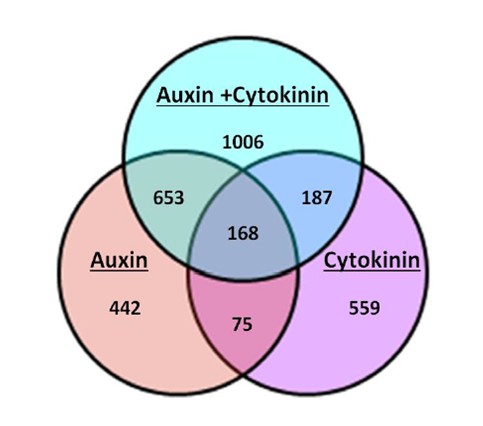
My work in the Sorefan lab in the University of Sheffield’s department of molecular biology and biotechnology (MBB), was to investigate the role of a novel hormonal response produced by auxin and cytokinin. Previous microarray data by the lab revealed an interesting occurrence when Arabidopsis thaliana seedlings were grown under conditions of elevated auxin (IAA) and cytokinin (BAP) concurrently. When grown under a dual hormone combination, the seedlings showed a significant change in the number and types of genes that were either significantly up or downregulated in the microarray. This response was larger than either responses of auxin or cytokinin alone and has been termed the ‘dual hormone response’ (Fig 2).
Fig 2: A Venn diagram illustrating the number of genes regulated by auxin alone, cytokinin alone, a combination of auxin and cytokinin and their overlaps. Nearly twice as many genes have significantly altered expression levels when auxin and cytokinin are combined compared to either hormone alone. (Image credit: James Thackery, Sorefan lab)
Functions of the dual hormone response
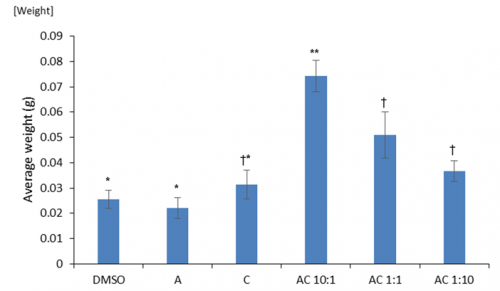
Prior to environmental stresses we tested if there were any general positive growth responses of the dual hormone response (DHR). We investigated the effects of differing ratios of auxin to cytokinin on cotyledon weight. Cotyledons are embryonic leaves that are among the first structures to emerge following germination and their weight is an accurate proxy of seedling health. We found significantly greater cotyledon mass under DHR conditions particularly with ratios showing high concentrations of auxin (Fig 3). Interestingly, this was significantly higher than the effects of just auxin alone, implying that the exogenous application of even a small amount of cytokinin may produce positive benefits.
Fig 3: The effects auxin alone (A), cytokinin alone (C) and combinations of auxin and cytokinin (AC various ratios) on cotyledon weight compared to a DMSO control. Means that share a symbol (*, ** or †) do not statistically differ from one another.
Comparing our lab’s dual hormone microarray data with that of other labs highlighted approximately 11.2% of our identified genes may show some response to heat stress conditions. I produced cDNA from seedlings grown under different combinations of hormone treatments and designed primers for the heat stress responsive genes identified by these microarrays. Using multiple quantitative real-time polymerase chain reactions (qRT-PCR), I confirmed that a number of these heat-responsive genes were also significantly affected by the dual hormone response.
I then conducted an experiment to investigate the phenotypic effects of heat shock on seedlings grown under DHR conditions (Fig 4).
Fig 4: Heat shock experiment before (left image) and after (right image) showing bleaching of the cotyledons as the chlorophyll degrades.
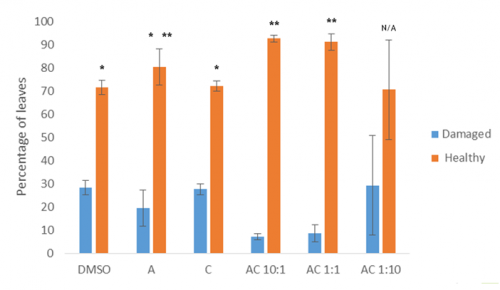
Preliminary data from this experiment suggests that the dual hormone response may convey some resistance to heat stress, as combinations of the two hormones results in significantly fewer damaged / unhealthy leaves than the control or either single hormone alone (Fig 5).
Fig 5: Percentage of leaves that were either damaged or healthy following heat stress under varying hormonal conditions.
Future work aims to quantify chlorophyll more effectively as our own analyses were confounded by the production of purple anthocyanin pigments, which are often produced as a response to stress.
These data may therefore allow us to apply dual hormone responses to agriculture. The dual hormone treated plants were larger and appear to be somewhat de-sensitized to heat stress, which may allow us to manipulate crop plants to convey some heat resistance to them. Knowing the optimal hormone ratios is important for applying this information. Manipulating plants in this way is a promising route for producing ‘climate ready’ crop species and ensuring that we can continue to feed a world of 7 or 10 billion in a vastly deteriorating climate.
Department of Experimental and Health Sciences, Universitat Pompeu Fabra, Barcelona, Spain
Reconstructing the lineage relationships and dynamic event histories of individual cells within their native context is central to understanding how the wide diversity of cell types develops during the construction of an organ. This is a long-standing challenge in biology, because up to now most efforts have been devoted to understand the genetic requirements for cell specification.
The inner ear is an attractive model to take on this challenge as it contains a manageable number of cell types, which develop rapidly on an organized schedule to generate the functional units of the mature organ – the sensory patches, containing supporting cells and hair cells innervated by the sensory neurons (Video 1).
Video 1: Innervated sensory patches in the embryonic inner ear. Animation of an embryo displaying the sensory patches with differentiated cellular neurosensory elements in green at 48hpf. Differentiated hair cells are innervated by sensory neurons of the SAG, which shows the typical segregation into anterior and posterior portion alongside with the segregated projections to the hindbrain.
During the last decades we learnt how neuronal vs. sensory specification was achieved, which signals from the surrounding tissues regionalized the otic vesicle along axes, and that the gradual restriction of cell fates over time was due to a multistep process. However, how sensory and neuronal progenitors behave throughout patterning, proliferation, and morphogenesis remained elusive, and it was difficult to reconcile some of the phenotypes observed in the signaling targeted mutants (Raft and Groves, 2015), due to the limited comprehension of how developmental gene regulatory networks are integrated. We thought that in vivo cellular data could address how patterns are achieved while the cells proliferate and the tissue undergoes morphogenesis, which may affect cell positioning and exposure to signals, and therefore cell specification. We were inspired by the pioneer work of Julien Lewis, who was able to foresee how things would work through in principle simple observations (Haddon and Lewis, 1996).
After a while trying to understand how the different progenitor pools were spatially organized within the otic vesicle using classical technics, it became clear to us that only imaging the whole developing inner ear and going from developed structures backwards in time to founder cells would help us to unveil the answer.
As you can imagine, this turned out to be a bit more complex than expected. It could only be accomplished through high resolution in vivo imaging, and considering that the otic vesicle undergoes extensive morphogenesis at the same time cells specify and proliferate it constituted a major challenge. It required simultaneously tracing the lineages while resolving the kinetics of cell proliferation and fate behavior. Recent developments in 4D-microscopy and cell tracking tools were being developed, but that was something that a small group could not afford to dream about. However, we were lucky enough to learn about the efforts of the lab of a former colleague, Nadine Peyrieras, in developing original methodologies and tools for the in vivo multiscale and multimodal observation of biological processes (Olivier et al., 2010; BioEmergences). After discussing with Nadine, we were excited because we felt we could participate in filling the void between gene regulatory networks and tissue architecture.
Our goal was to address two important questions: i) how neurons and hair cells are specified within similar domains of the otic epithelium; and ii) how from a simple otic epithelium this complex 3D-structure (the embryonic inner ear) is generated with the precise allocation of the neurosensory elements. We undertook this task without being conscious of the enormous effort and all the troubles we had to face, and combined high-resolution imaging with genetic tools using zebrafish embryos. It started our long journey in learning how to image for long periods of time using the best fluorophors, how to use new platforms for image processing, how to write in different programming languages (we became fans of coursera), and most importantly we had to change our prefigured ideas of how things work. Finally, we could record cell lineage information and observe at every time the spatial context of the cells within the whole otic vesicle! This period was very fruitful, and we realized how powerful these tools are and that we can use imaging for discovery.
Video 2:Early organization of neuroblasts within the SAG. Tg[cldnb:lynGFP]Tg[Brn3c:GFP] embryos injected with H2B-mCherry mRNA were imaged, and reconstructed cell centers were color-coded according to their location/identity (see legend). The projection view video (large panel) simultaneously displays the topological organization of cell groups selection and tissue architecture as a projection of the GFP channel (plasma membranes in grey) in x,y,z-axes. The distinct visualization modes displayed on the right hand side allow for a detailed 3D-visualization of data during the analyses. Orthogonal views are used to validate cell tracking, the oblique slice view allows orienting the orthoplane along the embryonic axes, and the rendering view permits to display validated cell centers in the context of the whole image volume.
These experiments led to understand the remodeling of the neuronal progenitor domain upon neuroblast delamination (Figure 1), and revealed that the order and place of neuroblasts’ delamination from the otic epithelium prefigure their position within the statoacoustic ganglion (SAG) (Video 3). Up to date, we knew neuroblasts were exiting the otic epithelium by a process called delamination, and they formed the SAG beneath, but we could not imagine the impact that delamination and morphogenesis had in the spatiotemporal distribution of neurosensory cell progenitor pools. By following the individual progenitors we unveiled that approximately 25% of the cells of the otic vesicle delaminate within few hours, and that the homeostasis of the system was maintained, most probably due to the proliferative capacity of the non-sensory epithelium. The developmental strategy used by distinct progenitor populations differed: neuronal specification is concomitant with proliferation (before/after delamination), while hair cell specification and differentiation lead to postmitotic cells indicating that the final number of sensory cells relies on the control of the progenitor pool.
Figure 1:Dynamic map of neuronal progenitors (orange circles) and their epithelial neighboring cells (grey circles) in the context of the whole otic vesicle (grey dots) over time. Color intensity of cell centers depicts the position of cells along the dorsoventral axis of the otic vesicle. The map was built after following the lineages from 18hpf to 26hpf of all encircled cells. Note how neuroblast delamination impacts on the size and position of the progenitor domain (orange circles) over time.
Video 3: The order and place of delamination foreshadows their location within the SAG. A cohort of 144 delaminated neuroblasts was analyzed for time and place of delamination. Reconstructed cell centers were color-coded according to four delamination intervals: 18-20hpf white, 20-22hpf yellow, 22-24hpf orange, 24-30hpf red, or for position along the anteroposterior (AP) axis in the otic epithelium and followed from 18hpf to 30hpf. Note that neuroblasts exit randomly from the delamination domain; those delaminating earlier are located more medially in the SAG than the later delaminating ones prefiguring the gradient of differentiation (middle panel). The relative position of neuroblasts along the AP is maintained from the otic epithelium to the SAG (right hand side panel). Reconstructed cell centers were displayed as colored dots together with the corresponding volume rendering images (plasma membranes in grey).
Moreover, we provide the dynamic map of neurosensory progenitors based on in vivo cell lineage studies (Video 4) supplying a global and temporal perspective to previous otic neurosensory lineages analyses, which were mainly focused on the spatial dynamics of gene expression (Durruthy-Durruthy et al., 2014).
Video 4:Maps of neuroblasts and posterior macula hair cell progenitors in the whole otic vesicle. The neuronal progenitors (orange) and the posterior macula hair cell progenitors (blue) are plotted in the context of the whole otic vesicle (grey dots) at 24hpf. Tether cells are shown as black circles. The animation displays a rotation of otic vesicles around the anteroposterior axis. Note that the two progenitor domains are adjacent, and neuroblasts are located more ventrally while sensory progenitors are more medially.
We are very enthusiastic with this paper because the information about cellular/population dynamics and lineage relationships of neurosensory elements in the inner ear enables us to understand the proportions of the system and provide the cellular data to complement the well-described gene regulatory networks involved in neurosensory specification. We are convinced that for the understanding of how the developmental gene regulatory networks function during tissue degeneration and regeneration, as well as in normal patterning, we need to move forward into 4D-imaging and put previous knowledge in the context of the dynamics of the system.
References
Durruthy-Durruthy, R., Gottlieb, A., Hartman, B. H., Waldhaus, J., Laske, R. D., Altman, R. and Heller, S. (2014). Reconstruction of the mouse otocyst and early neuroblast lineage at single-cell resolution. Cell 157, 964–978.
Dyballa, S., Savy, T., Germann, P., Mikula, K., Remesikova, M., Špir, R., Zecca, A., Peyriéras, N. and Pujades, C. (2017). Distribution of neurosensory progenitor pools during inner ear morphogenesis unveiled by cell lineage reconstruction. Elife 6: e22268.
Haddon, C. and Lewis, J. (1996). Early ear development in the embryo of the zebrafish, Danio rerio. J. Comp. Neurol. 365, 113–128.
Olivier, N., Luengo-Oroz, M. A., Duloquin, L., Faure, E., Savy, T., Veilleux, I., Solinas, X., Debarre, D., Bourgine, P., Santos, A., et al. (2010). Cell Lineage Reconstruction of Early Zebrafish Embryos Using Label-Free Nonlinear Microscopy. Science 329, 967–971.
Raft, S. and Groves, A. K. (2015). Segregating neural and mechanosensory fates in the developing ear: patterning, signaling, and transcriptional control. Cell Tissue Res 359, 315–332.
Established by the British Society for Developmental Biology (BSDB) in 2014, The Gurdon/The Company of Biologists Summer Studentship scheme provides financial support to allow highly motivated undergraduate students an opportunity to engage in practical research during their summer vacation. Each year, ten successful applicants spend eight weeks in the research laboratories of their choices, and the feedback we receive is outstanding.
Our third report from the 2016 class comes from Iona Imrie, who undertook her studentship with Jamie Davies in Edinburgh.
Development of the vascular system in the mouse mesonephros
During the summer of 2016, I was fortunate enough to be awarded a BSDB Gurdon Studentship. The funding enabled me to undertake a research project in the lab of Jamie Davies at the University of Edinburgh. Under the supervision of a PhD student in the lab, David Munro, I studied the vascularization of a primitive and transient murine kidney- the mesonephros.
During mammalian embryogenesis, 3 paired renal organs develop sequentially in a cranio-caudal direction. The most primitive kidney is the pronephros, followed by the mesonephros and then the permanent kidney, the metanephros. Both the pronephros and mesonephros are temporary, with the pronephros being nonfunctional and the mesonephros being functional in some species. The mouse mesonephros comprises 18-26 pairs of tubules. The most cranial tubules connect to the nephric duct, while the caudal tubules do not. The mesonephros first appears at embryonic day 9 (E9), before regressing completely in females. In males, the cranial tubules do not degenerate, instead forming the epididymis. Given that, (a) the fate of the mesonephros is sexually dimorphic and (b) the mesonephric tubules are not uniformly connected to the nephric duct, there could be differences in vascular development that run in parallel with these differences in morphology. During my project, I observed vascular development in order to identify where blood vessels came from and if there were differences in vascular development between sexes and cranial/caudal tubules of the mesonephros.
My project began with dissecting embryos at E11.5. The mesonephroi obtained by dissection were stained with antibodies against CD31, Laminin and Pan-cytokeratin. These antibodies stained endothelia, basement membrane and nephric duct/tubules respectively.
I set out to create a timeline of mesonephric development starting at E10.5 through to E15.5 (Figures 1-4). Dissecting at E10.5 proved difficult due to the small size of the embryo. The gonad develops adjacent to the mesonephros, and I left itattached to the mesonephros in order to observe interactions of vasculature between the two.
Having dissected and stained mesonephroi from various ages, I made the following observations:
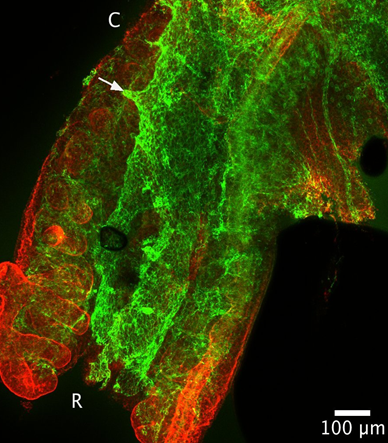
E10.5– Aorta branches into mesonephros (Figure 1).
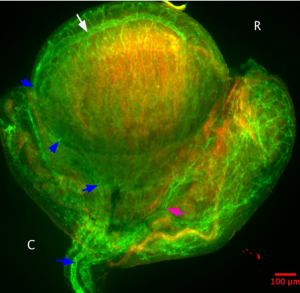
Figure 1: Green=CD31, Red=Laminin. The aorta is shown centrally, between the two mesonephroi. Branches of the aorta going toward the mesonephros can be seen (white arrow). C indicates the caudal end of the mesonephros, and R is the rostral end.
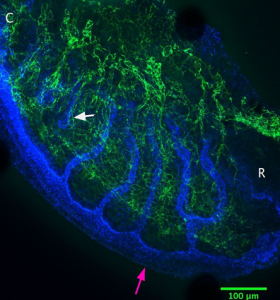
E11.5– Tubules elongate and the mesonephros vascular plexus (mvp) begins to form (Figure 2).
Figure 2: Blue= Pan Cytokeratin, Green= CD31. Compared to E10.5, the tubules look longer. The pink arrow shows the nephric duct, and the white arrow shows the first tubule that is disconnected from the nephric duct. There is a collection of vasculature at the distal end of the tubules where the aorta would be. There are branches into the mesonephros from here.
E13.5– In females, branching of mvp into mesonephros and gonad occurs. In males, branching into the mesonephros occurs but branching into the gonad is not as visible as in females.
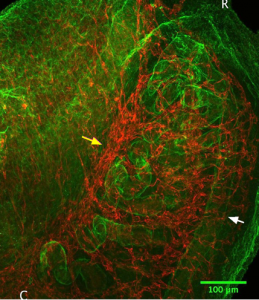
E14.5– Formation of the coelomic vessel on male gonad. Continued branching of mvp into mesonephros/gonad in females (Figure 3).
Figure 3: Immunostaining of an E14.5 female mesonephros (right) with gonad attached (left). Green= Laminin, Red= CD31. The white arrow points to the nephric duct and the yellow arrow points to the mvp. Blood vessels seem to branch from the mvp into the mesonephros and into the gonad
E15.5– Branching of coelomic vessel into gonad, and joining of the gonadal artery to this vessel (Figure 4).
Figure 4: Green=CD31, Red=Laminin. It seems that there is more CD31 staining in this E15.5 male than at earlier ages. The coelomic vessel (white arrow) which first appeared at E14.5 now joins to the gonadal artery (blue arrows). This vessel branches into the mesonephros and bifurcates (pink arrow).
Overall, the vasculature of the mesonephros seems to branch from the mvp and form a network of vessels around the nephric duct. The vessels around the tubules do not look particularly organized. However, confocal slices at 40x objective show vessels wrapping around the tubules. I did not observe glomeruli in the tubules at any age- these structures are rarely found in mouse mesonephroi.
As an adjunct to the main project of vascularization, I tested the functionality of the mesonephric tubules using an assay developed by one of Prof. Davies post doctoral researchers, Dr. Melanie Lawrence. The assay tested the functionality of the tubules to uptake fluorescent anions and cations. The ability of cells in these tubules to transport organic anions and cations would suggest the tubules are functioning as a primitive kidney. It has been assumed that the murine mesonephros has this role, but this has never been proven. Melanie had been using her assay to answer this question, using mesonephroi from different gestational ages. The unpublished data that Melanie has collected shows that these tubules do have the ability to transport organic anions and cations. I was able to help with assaying at some of the gestational time points.
I cultured mesonephroi using the Sebinger culture method, and then assayed the uptake of 6-carboxyfluorescin (anion) through organic anion transporters (OAT) in the basolateral membrane of tubule cells, and uptake of the DAPI (cation) through organic cation transporters (OCT), also in the basolateral membrane. As a control, inhibitors of OAT (Probenecid) and OCT (cimetidine and metformin) were added to mesonephric cultures to show that any fluorescence seen in the tubules was due to uptake via these transporters and not by another process.
The Gurdon Studentship has been an invaluable opportunity. I cannot stress enough how important it is for medical sciences students like myself to spend time in a research lab. As a second year, I didn’t really know what being a scientist would be like. From spending time in the Davies Lab, I have been able to understand and practice techniques that were briefly introduced in lectures, while developing a scientific mind, improving my time management skills and working alongside scientists who are extremely dedicated to their research. I would especially like to thank Jamie Davies for giving me this opportunity, and David Munro and Chris Mills for their guidance throughout my time in the lab.
A position (#121670) is available immediately for a Postdoc Associate to contribute to our studies in neural crest and placodes. The Postdoc Associate will conduct independent research and assist in the training of students in the laboratory of Dr. Lisa Taneyhill at the University of Maryland. Laboratory skills should include the ability to perform various molecular biology and biochemical assays, such as recombinant DNA/cloning; DNA, RNA, and/or protein blotting; immunohistochemistry; and/or in situ hybridization. Experience with microscopy and spectroscopy, chick embryology (including microdissections and electroporation), and tissue culture is desirable. For more information on the lab, please see http://www.ansc.umd.edu/people/lisa-taneyhill. Qualifications: An advanced degree (Ph.D.) in Developmental, Molecular and/or Cell Biology is required. Fluency in spoken and written English is required. Compensation: Salaries are highly competitive, negotiable and commensurate with qualifications. Fringe benefits offered. Applicants must apply through eTerp at https://ejobs.umd.edu. Applications will be accepted until a suitable candidate is identified.
How does a developing embryo coordinate DNA replication with gene transcription? This fundamental question is the focus of today’s paper, recently published in eLife. We caught up with lead author Ehsan Pourkarimi and his supervisor Iestyn Whitehouse, Assistant Professor in the Sloan Kettering Institute’s Molecular Biology programme, to hear the story behind the work.
The Whitehouse lab, with Iestyn front row middle, Ehsan standing second from right, and middle author James standing right. From the Whitehouse lab homepage.
Iestyn, can you tell us your scientific biography and the general focus of the Whitehouse lab?
IW My first project as a graduate student was using biochemistry to study how nucleosomes are repositioned along DNA by so-called chromatin remodelling complexes in the lab of Tom Owen-Hughes in Dundee, Scotland. After that, I wanted to learn more about chromatin in vivo so I joined Toshi Tsukiyama’s lab at the Fred Hutch in Seattle. There, I learned to use budding yeast and genomics and studied how genes are activated and repressed by nucleosomes.
I’ve always been interested in the question of whether chromatin states can be inherited through generations. When I started my own lab, I wanted to focus on understanding the basics of DNA replication in vivo with the hope that would allow a better understanding of how chromatin may be intertwined with DNA replication. More generally, my lab works on chromatin and how it relates to gene expression and DNA replication.
Sloan-Kettering would seem an ideal place for a molecular biologist dabbling in developmental biology?
IW Absolutely, although I must admit I was first attracted to come here because of the strength of the research in DNA replication and genomic integrity. With my background in chromatin and gene transcription I wanted to be around people who could help me with some project ideas I had about DNA replication. As the projects in my lab have developed it’s a tremendous benefit to be able to interact with developmental biologists here at Sloan Kettering.
And Ehsan, how did you come to work in New York with Iestyn?
EP During both my undergraduate and PhD research I mostly focused on how cells commit suicide or more precisely apoptosis, a genetically regulated process of programmed cell death. I did my PhD in Anton Gartner’s labin Scotland, for a few exciting years that turned out to be scientifically very productive. Toward the end of my PhD, I became fascinated about the field of chromatin biology and epigenetics. The last year of my research in Scotland mostly revolved around understanding how cells respond to acute stress conditions such as starvation. Further, I was interested in a possible role of chromatin remodelers and histone modifications in such stress responses. This research led to the discovery of the function of histone H3.3 variants in both starvation response and aging.
It was around the same time that Iestyn and Duncan Smith (a former postdoc in Iestyn’s lab) for the first time purified and sequenced Okazaki Fragments in budding yeast; the biased distribution of yeast Okazaki fragments perfectly matched the previously known replication origins. Right then I was sure that this approach could be utilized in metazoans. I was motivated to find out if we finally could get a complete picture of metazoan replication origins. So I decided to shift my research from how cells die to how they are born. Iestyn’s lab was the ideal place for my new interest. After meeting with Iestyn and discussing the idea of establishing C.elegans system in the lab, I moved to New York. Iestyn is among the top scientists in the field of chromatin biology and open to new ideas and approaches, which has made my transition to this field easy and exciting. Three years later, I can confidently tell you that moving to New York and switching my research field was one of my best decisions.
“I was motivated to find out if we finally could get a complete picture of metazoan replication origins. So I decided to shift my research from how cells die to how they are born”
Your previous work has dealt with chromatin and replication in S.cerevisiae. What made you decide to use C. elegans in this paper?
IW Although I enjoy working with yeast, I think I’d work with any organism if there were an interesting question we could address. In terms of this project, I was interested in testing whether our general findings about the relationship between chromatin and DNA replication from budding yeast would hold up in another organisms. Ehsan came to me with many years of experience working with C. elegans; he told me he wanted to map DNA replication in worms – I said go for it!
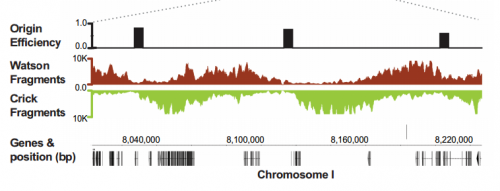
Replication origins and Okazaki fragments mapped onto the genome, from Figure 1, Pourkarimi, et al. 2016. eLife.
Was it difficult to apply techniques originally used in yeast to developing worms?
IW & EP If you would ask me this question 3 years ago I would bet against myself! Having said that, the beauty of having questions is that they force you to challenge yourself. I wouldn’t necessarily say it was difficult, but it required hard work. We needed tons of optimization and we went through many many trails and errors. The beauty of C. elegans is its ease of genetic manipulation. Once we knew how strongly we needed to knock down lig-1, we were able to radiolabel and visualize Okazaki fragments. The most challenging part was purifying Okazaki fragments without causing too much DNA damage to developing embryos.
“The beauty of having questions is that they force you to challenge yourself. I wouldn’t necessarily say it was difficult, but it required hard work”
Can you give us the key results of your paper?
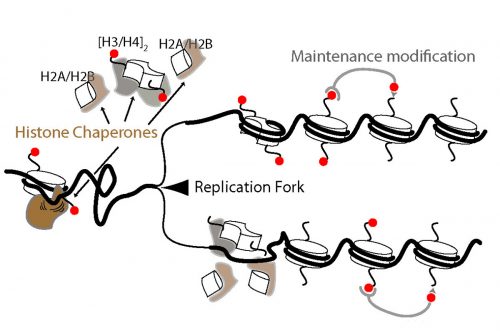
IW & EP We have studied DNA replication in multicellular Caenorhabditis elegans embryos and have identified a distinct set of replication origins. Interestingly, replication origins are marked by histone proteins containing specific modifications such as H3K27ac. These modifications are typically associated with sites of active gene transcription. This finding raises a key question: Does transcription define the sites of replication origins, or do replication origins define where transcription will occur?
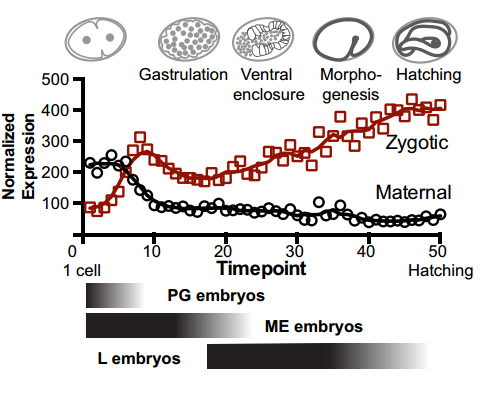
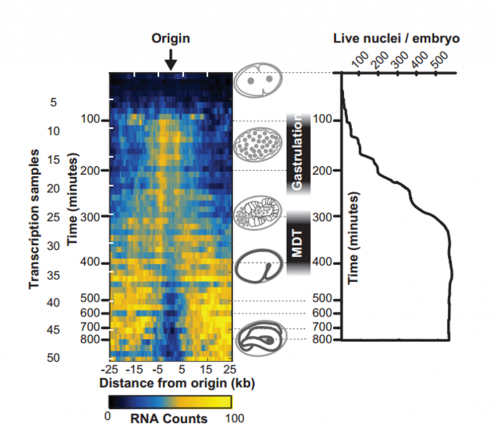
To answer this, we identified replication origins in very early embryos (pre-gastrula); at a stage in embryogenesis in which most genes are inactive. Surprisingly, we find that replication origins are present at this very early stage, which shows that replication origins precede active transcription. Nevertheless, we found that once transcription begins, it does so in close proximity to the pre-defined origins. This close association of gene transcription and DNA replication persists until the embryonic cells stop dividing and experience a profound change in their gene activity and morphology. At this stage of development, the association between DNA replication and gene transcription breaks down and genes located farther away from replication origins begin to be transcribed. Our work suggests that the genome has evolved to couple DNA replication with gene transcription in rapidly dividing cells.
Maternal and zygotic transcripts expression through development, from Figure 3, Pourkarimi, et al. 2016. eLife.
Does this work imply that the layout of genome is tied to the mode of development of the animal?
IW & EP Yes, it’s an attractive possibility. The rapidity of S phase in embryonic cells – which, in C. elegans is on the order of 15-20 minutes – is probably a strong determinant of genome organization. To replicate a 100mb genome in that timeframe you need a replication origin about every ~75kb. Like other organisms, replication origins in C. elegans are associated with transcriptionally active chromatin marks, so we think that the requirement for regularly spaced origins has influenced gene organization: genes associated with growth are clustered near replication origins. We’re very interested in the idea that replication origins may be intimately tied to nearby gene activity. One consequence of this is that activation and inactivation of origins will profoundly alter not only the transcription program but also the kinetics of S phase.
IW/EP One of the striking features of the MDT is the switch from the expression of “growth” to “differentiation” genes. Broadly speaking you want to express growth genes in cycling cells (when you’re replicating DNA) and differentiation genes when you’re not. If you couple the expression of growth genes with replication origin function then inactivation of replication origins may be part of the trigger to shift the transcriptional program from growth to differentiation. Of course, this is mostly speculation, nature is full of surprises and we’ll wait and see how it turns out.
Transcript abundance related to distance from replication origin and number of nuclei through development. From Figure 4, Pourkarimi, et al. 2016. eLife.
You reference and even include comparison data from quite a few papers published in the seventies and eighties. What was it like to revisit this old work?
IW & EP It’s always rewarding to go back and dig through the older literature – we cite some classic papers that have stood the test of time. It’s also very important understand what current dogma is based on: all too often what we think we know isn’t really supported by the facts and when you go back to the literature you realize that there are large gaps in our knowledge. In this paper we’ve found that replication origins are established very early in development, which goes against the dogma; the old data isn’t wrong, rather we’ve found that C. elegans behaves differently to Xenopus and Drosophila.
“All too often what we think we know isn’t really supported by the facts and when you go back to the literature you realize that there are large gaps in our knowledge”
When doing the research, did you have any particular result or eureka moment that has stuck with you?
EP The very first time I experienced such feeling was during my undergraduate studies in Budapest in the lab of Krisztina Takacs. At the time, I was working on the C. elegans counterpart of the anti-metastasis gene nm23. After long hours of staring into a microscope, I could clearly see nm23 is modulating the RAS/MAPK pathway; a very unexpected finding that made me lose sleep for many nights. Back then, in my innocence, I thought I would never forget the joy and the satisfaction I felt in that moment. Years later in a meeting one of my former professors congratulated me on passing the test of time on the same finding, and I realized that that was my eureka moment. As for this research, I will have my eureka moment once time puts its stamp of approval on our findings.
And what about your plans following this work?
EP We have opened up multiple interesting questions in this field. Currently we are interested in looking at replicating pattern in highly specialized somatic cells, which are transcriptionally dissimilar to embryonic cells. We know that histone modifications are altered in such cells, which indicate a possible variant replication profile. At the same time, we are using different genetic screens to pinpoint upstream component(s) of replication initiation.
Do you plan to continue this developmental strand in the Whitehouse lab?
IW Yes – although I wouldn’t classify myself as a developmental biologist just yet. Maybe the best way to describe my thinking is that I’ve come to realize (rather belatedly) that context is all-important. As molecular biologists we typically study processes and systems far removed form the context in which they typically function. Mostly, this is dictated by practicality of getting the experiments done. But the cost of the practicality is often paid by not being able to contextualise the results. In the case of chromatin, for example, I think we understand the basic principles of what chromatin is, but there are many aspects – not least the biological functions of histone modifications – that are still a real mystery. Generally speaking, I think we’ll figure out some of the mysteries if we can identify the biological context in which certain chromatin states and histone modifications come in to play. This is where developmental biology will be especially powerful – we’ll see how far we get!
“As molecular biologists we typically study processes and systems far removed form the context in which they typically function…the cost of the practicality is often paid by not being able to contextualise the results”
Established by the British Society for Developmental Biology (BSDB) in 2014, The Gurdon/The Company of Biologists Summer Studentship scheme provides financial support to allow highly motivated undergraduate students an opportunity to engage in practical research during their summer vacation. Each year, ten successful applicants spend eight weeks in the research laboratories of their choices, and the feedback we receive is outstanding.
Our second report from the 2016 class comes from Lauren Miller, who undertook her project with Richard Poole in UCL.
The C. elegans dilemma: should I stay or should I go?
After being awarded the BSDB Gurdon Summer Studentship, I entered the lab with the enthusiasm that only an undergrad who has never spent 12 hours looking down a microscope can possess, and the surety that I was going to discover something. I then spent the next month failing to get my assay to work, which I feel gave me a much healthier perspective of the kind of dedication this whole “discovering” thing was going to require. Happily though, after another couple of months, I did manage to produce some interesting data!
Nervous systems are incredibly interesting, complicated networks, and trying to elucidate what they are doing through behavioural studies is even more challenging. Animal nervous systems are largely left-right bilaterally symmetric, yet they are often functionally lateralised. This is well shown in the human brain where specific functions, such as language, are assigned specifically to only one of the two bilaterally symmetric hemispheres. Congenital defects that disrupt underlying asymmetry have been associated with a wide variety of human neurological disorders, yet the development of left-right asymmetry in the nervous system is poorly understood. The main aim of my project was to see if specific neuroanatomical asymmetry observed in the nervous system of C. elegans corresponded to a functional behavioural asymmetry.
And to do this I had to pick worms, lots and lots of worms. In fact, the first time I tried to pick enough to do a complete assay it took me over 8 hours, and I had barely collected half of them. I then had to beg one of the labs PhD students to help me pick the rest, and I watched in horror as he finished the other half in about 15 minutes. The secret, I learned, was to not pick them up one at a time, but to pick up about 15 or 20 before transferring them to the other agar plate. Fortunately, I can now collect them all in an hour, not that it wasn’t character building doing it the hard way, but I can get an awful lot more done now.
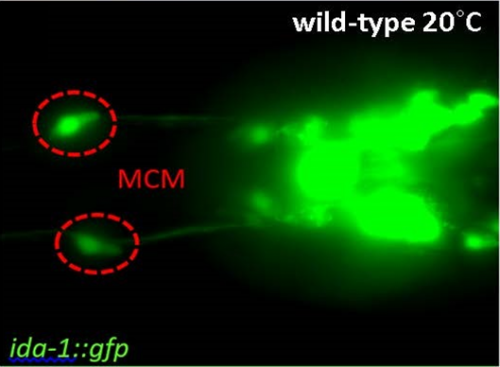
C. elegans are naturally attracted to several tastes and odorants, however they can learn to associate them with aversive stimuli and switch their behaviour to being repelled. The assay I was carrying out conditioned males to associate benzaldehyde, which they are attracted to, with different stimuli. When they are conditioned to associate the smell with starvation they are repelled, however this response in males can be trumped if they are also conditioned with mates at the same time. This is sexual conditioning, which has recently been attributed to two newly discovered male specific neurons called MCM’s (M. Sammut, 2015)
Figure 1: Micrograph with a dorsal view of male C. elegans oriented anterior to the left. MCM’s are green (ida-1::gfp) (L. Lin, 2015).
Normal males behave like this after being conditioned to associate benzaldehyde with different stimuli:
Figure 2: Behavioural responses to odorant after conditioning of unablated males.
Since this was a perfect example of a pair of left-right bilaterally symmetric neurons, the interesting question was now were they indeed functionally lateralised, as was suspected from their connectivity’s. If I ablated either the left or the right MCM what would happen? Would there be an asymmetry in the circuit? Would the worms behave differently if I ablated the right MCM versus the left MCM?
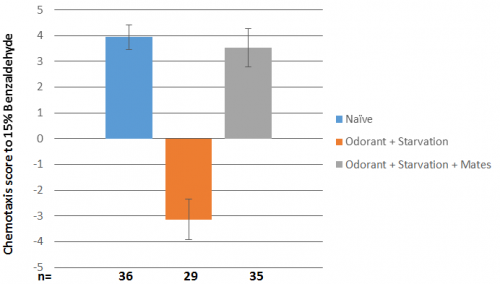
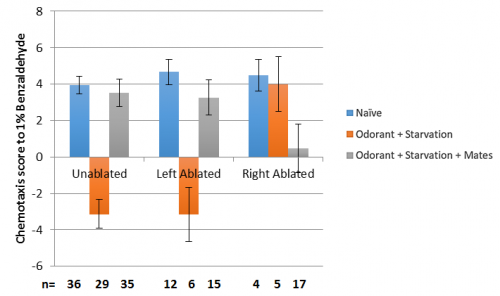
As it turns out they did, but not the way we expected; from the connectivity’s it was hypothesised that the right MCM may be more important for sexual conditioning than the left. This did turn out to be the case, but what also happened was that it disrupted the worms’ ability to aversively condition to benzaldehyde, which is usually very robust. The results that were obtained are as follows:
Figure 3: Results of unablated, left and right ablated C. elegans. The “n” for aversive conditioning with left and right ablated animals is low as in the first few assay’s most of the ablated worms were put in the sexual conditioning plate to try and generate data for this condition; aversive conditioning was not expected to be effected as in mutants with neither MCM, males behave the same as the wild type.
Firstly, this shows clearly that there is an asymmetry in the function of the MCM’s; ablating the left MCM does not affect behaviour at all, which is very exciting! Secondly the worms with the right MCM ablated on the sexual conditioning plate were either very strongly attracted to benzaldehyde or repulsed in an almost 50/50 ratio, hence the bar is very small, and the error is huge. Thirdly there are other interactions going on with the sensing of benzaldehyde, which is sensed by the AWC neurons, as right ablated worms do not appear to aversively condition.
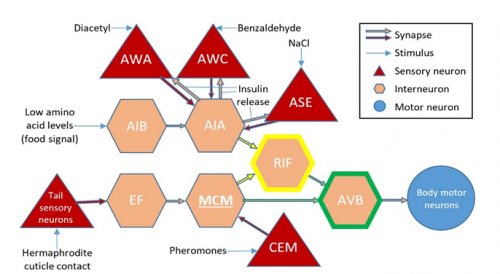
This is the connectivity of the AWC’s with the MCM’s:
Figure 4: Connectivity of the MCM’s with respect to certain sensory and interneurons (L. Lin, 2015).
Both AWC’s sense benzaldehyde, but they are asymmetric in the sense that during development STR-2, a G-protein coupled receptor, is expressed randomly in one of them, turning it on, whilst the other stays off (P. Wes, 2001). The CEM’s could also be involved, as they are responsible for sensing pheromones given off by the hermaphrodites. There are several theories on what might be happening so far, but further experiments, such as ablating both the MCM’s, will be needed to really explain the source of this behavioural asymmetry.
I loved my time in this lab, and if I didn’t have a degree to finish I’d probably still be there trying to figure out this very interesting behaviour!
References
Lin, A. B., 2015. Establishing a Behavioural Paradigm for Sexual Conditioning to the Odorant Benzaldehyde – a role for the MCM neurons?, London: UCL.
Sammut, 2015. Glia-derived neurons are required for sex-specific learning in C. elegans. Nature, Volume 526, p. 385–390.
Wes, C. B., 2001. C. elegans odour discrimination requires asymmetric diversity in olfactory neurons. Nature, Volume 410, pp. 698-701.
Certain scientific meetings have their place in the book of history as a visible shift — or a turning point, if you will — in a particular field. The Cuvier-Geoffroy debate at the French Academy of Sciences in 1830 set the tone for the subsequent two centuries of biology in search for determinants of organismal forms. Closer to our times, you might think of the Santa Fe workshop that kick-started the Human Genome Project or, even more recently, the creation of a new society (the Pan-American Society of Evolutionary Developmental Biology) following a NESCent workshop. Common threads connecting many such examples are small sizes that allow intellectual catalysis (typically no more than 50 people participating) and timing them just ahead of the curve. The First International Hemichordate Meeting held at Hopkins Marine Station (Stanford University) from December 9 to 11 2016 may be remembered as just such a shift in the history of study in chordate origins.
A group photo on the final day of the First International Hemichordate Meeting (provided by Chris Lowe)
A Prelude to the First Hemichordate Meeting
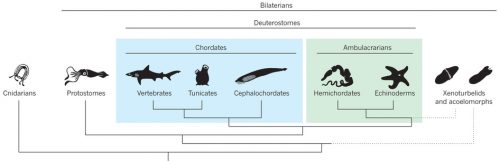
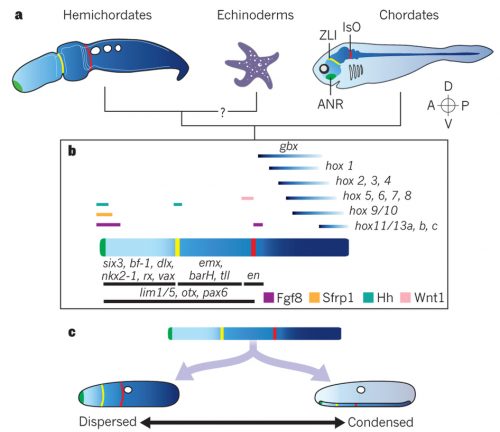
Hemichordates are a small phylum of deuterostomes. The most conspicuous members — enteropneusts — are commonly called acorn worms, and their preposterous appearances are unforgettable. Hemichordates contain another living lineage of deep-sea, tube-dwelling, filter-feeding forms called pterobranchs. As the name suggests, hemichordates have long been considered our close invertebrate relatives, nested just outside chordates. Unlike their sister lineage that is echinoderms, hemichordates sport pharyngeal slits, and since William Bateson, embryologists have been looking for a homologue of notochord or even a bona fide notochord itself in acorn worms. So, classically, this is the group of animals that supposedly root many characters of chordates. To the post-Bateson chordate researchers, however, hemichordates seemed too distant and too different to inform vertebrate origins directly. Cephalochordates and tunicates each have clearly vertebrate-like morphological traits, even though a closer look at their development does not support simple progression to vertebrates from either body plan.
A phylogenetic scheme of deuterostomes, including hemichordates (from Lowe et al. 2015, Nature 520, 456–465).
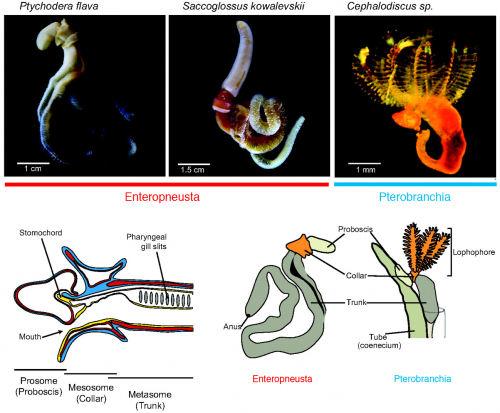
Hemichordate diversity and body plan (from Röttinger & Lowe, 2012, Development 139, 2463-2475).
But the tide is turning, and hemichordates are making a remarkable comeback onto the scene of the great chordate debate. I will touch on how and why in this report, and the change is palpable. In 2013, I attended a symposium “The Making of a Vertebrate” organized by RIKEN CDBin Japan, where even the conveners expressed their astonishment at how they ended up with so many (five!) hemichordate talks and how insights from hemichordates led the discussion throughout the symposium. I remembered this vividly. It did feel like that hemichordates are catching on.
Chris Lowe makes a case for hemichordates in the symposium “The Making of a Vertebrate” in 2013 (RIKEN CDB)
Hemichordate researchers at “The Making of a Vertebrate” symposium in 2013: Andreas Hejnol, Ariel Pani, Stephen Green, and Billie Swalla (L-R) at the table and Tom Humphreys with a wine glass (RIKEN CDB).
Three years since the RIKEN symposium, Chris Lowe (Stanford University) and Yi-Hsien Su (Academica Sinica) organized the First International Hemichordate Meeting. Hemichordate workers are sending a signal that the community is coming together, and that there is a plenty of exciting stuff to come. In my view, there are three different streaks among modern hemichordate workers: the Saccoglossus school that mainly uses the direct-developing acorn worm S. kowalevskii, geographically centered in North America; the Ptychodera school that mainly uses the indirect-developing P. flava, centered in Japan, Taiwan, and Hawaii; the pterobranch school that traces its origin to the study of ‘lophophorates’ (and perhaps most naturalist in its spirit — so this school also includes those who look at the strange family of deep-sea acorn worms, torquaratoriids). My report is organized along with this classification.
Hemichordates and the ‘Holy Grail’ of the Chordate Debate
Chris Lowe opened the discussion by providing an overview of the research that fueled this hemichordate revival. This includes the seminal papers describing expression profiles in the ectoderm of S. kowalevskii here and here. He outlined the prospect and challenges to identify shared features of hemichordate and vertebrate development. This introduction reflects a cautious approach to what some may consider as support for homology, especially around the holy grail of notochord.Nori Satoh (OIST) certainly resonates with it. About the hotly debated stomochord — an endodermally derived supporting structure of the proboscis, proposed as a notochord homologue by Bateson — he pointed out FoxE expressions shared not with notochord but rather with the endostyle. Coupled with the fact that hemichordate D-V axis is opposite to that of vertebrates, Satoh asked at what developmental stage hemichordates should be compared with chordates. The prospect is bright to recognize developmental traits of deuterostomes, but the window of comparison may be narrow for comparison of chordate traits. He highlighted a locomotory shift from ciliary bands to tail muscles as a significant event toward chordates and outlined his current efforts to characterize these evolutionary transitions through comparative genomics. He explores these thoughts further in the new book from the Academic Press, Chordate Origins and Evolution.
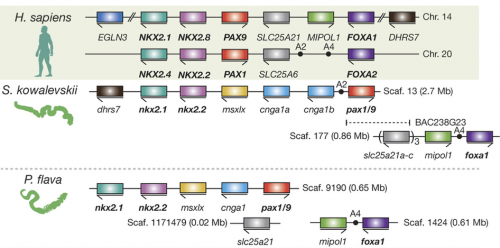
When it comes to genomics, Dan Rokhsar (University of California, Berkley) has acted as a hub of the Hemichordate Genome Project that published genomes on the direct-developing S. kowalevskii and the indirect-developing P. flava. Some of his comments that attracted interest from the audience are about conservation of synteny and about ‘pharyngeal’ gene clusters. The published genomes to which many other participants of the meeting contributed now form a foundation for some of the exciting ‘omics’ research going on. Jessica Gray (Harvard University) is certainly among the movers and shakers on this frontline. Her work on miRNA regulatory networks in S. kowalevskii, particularly those of neural and muscle development, captured attention from the audience (which also won a presentation award earlier in the inaugural meeting of the Pan-Am Evo-Devo Society). She and Marc Kirschner (Harvard University) are pioneering single-cell transcriptomics in S. kowalevskii. Kirschner’s talk and Gray’s poster caused quite an excitement — many participants can think of unique and interesting applications that could advance their research, especially by correlating the transcriptomic landscape with the observed expression profiles of marker genes.
These technical advances are indispensable to the ongoing efforts to compare S. kowalevskii with vertebrates. Stephen Green (California Institute of Technology) brought mesoderm to the discussion. Green is unique among the participants for using lamprey as a comparative model. He suggests that differential patterning of anterior and posterior mesoderm in S. kowalevskii perhaps shares an evolutionary root in the deuterostome stem with the cranial versus somitic differentiation in vertebrates. Rather than looking at region of a germ layer, Elena Silva (Georgetown University) focused on deriving neural progenitors. Here, too, parallels are observed with the pathways specifying neural progenitors in vertebrates. In the poster session, the Lowe lab displayed the current state of the art in research on S. kowalevskii that employs reporter expression, functional tools (CRISPR, RNAi), behavioral assays. These are Paul Minor, Nat Clarke, and José Lopez. But it was John Gerhart(University of California, Berkeley) who provided weight to this discussion. In the closing lecture of the meeting, he explored the question whether or not S. kowalevskii has a vertebrate organizer. This isn’t a run-of-the-mill comparison. Gerhart and his colleagues have been cloning almost every thinkable gene from S. kowalevskii that would be important in evolutionary context. Based on this massive dataset — a major portion of which remains to be published — he critically tested predictions arising from a hypothesis that the organizer evolved in the hemichordate-vertebrate common ancestor. This analysis intersects with such standing questions in hemichordate research as: Do hemichordates have a notochord homologue? How is the A-P patterning set in hemichrodates? How did the D-V axis evolve in deuterostomes?
Do hemichordates have a vertebrate-like organizer? (from De Robertis & Kuroda, 2004, Ann Rev Dev Cell Bio, 20: 285–308)
Regarding the polarity, the A-P patterning had multiple talks. Sébastien Darras (L’Observatoire Océanologique de Banyuls-sur-Mer) took advantage of functional tools available in S. kowalevskii to characterize the role of Wnt in establishing the A-P axis. Jens Fritzenwanker (Georgetown University) addressed axial elongation with a focus on the posterior growth zone. Yann Le Petillon (Academia Sinica) used the indirect-developing enteropneust, P. flava, to illustrate an interaction of Wnt and BMP signaling pathways to pattern and restrict anterior neural ectoderm. Ryan Range (Mississippi State University) gave an engaging talk by mapping complex GRNs for the anterior sensory organ of sea urchin larvae and dissecting multiple and complementing functions of Wnt pathways. This talk features his latest and very neat paper in Development.
Perspectives from Indirect-developing Enteropneusts
The indirect developing P. flava constitutes another stream of research on hemichordate development with the Asia-Pacific distribution. Kuni Tagawa (University of Hiroshima) provided this historical context by looking back on Tom Humphreys’s career. Tagawa was called on to Hawaii by Nori Satoh to work with Humphrey and collect P. flava. Early results from this collaboration include the paper describing Brachyury expression in the context of testing for a notochord homologue. Hiroshi Wada (University of Tsukuba) is another student of Satoh’s. He discussed a pygochord in Balanoglossus simodensis (a Japanese enteropneust). Whether this structure can be considered potential homologue of notochord will remain controversial, to say the least.
Over the last several years, Yi-Hsien Su and Jr-Kai “Sky” Yu (Academica Sinica) have built a team now leading the P. flava research (as well as echinoderms and cephalochordates). Su has a stable supply of embryos from a Taiwanese locality, and some of the research that is taking off from the lab was featured in the meeting. Her own work concerns the regulation of BMP signaling to establish D-V axis among ambulacularians (the lineage containing hemichordates and echinoderms). In addition to Le Petillon’s work on the A-P patterning, Tzu-Pei Fan from the Su lab reported on FGF signaling in the mesoderm development of P. flava.
Development of P. flava through a larval stage (G, H) (from Röttinger & Martindale, 2011, Developmental Biology, 354, 173-190)
Billie Swalla (Friday Harbor Laboratories) is another long-time P. flava expert and a chief architect of the current chordate phylogenetic framework. Her team contributed some unique perspectives to the meeting bylooking at regeneration by regions — proboscis, collar, and trunk. Her student Shawn Luttrell explored this work further in the context of stem cells. Formerly at the Swalla lab and now at the Halanych lab, Michael Tassia (Auburn University) called our attention to the immune system with phylogenetics and bioinformatics of Toll-like receptor pathways in bilaterians and showed that hemichrodates are the key taxon to consider the evolution of immunological memory toward vertebrates.
Paul Gonzalez (Stanford University) delivered a talk literally hot off the press —published in Current Biology just days ahead of the meeting. He pioneered another indirect-developing enteropneust, Schizocardium sp. in the Lowe lab. Surprisingly, the larvae lack expression profiles characteristic of the Hox-patterned trunk — in Richard Strathmann’s words, they are ‘swimming heads’. The trunk expression profiles appear across the metamorphosis. His work hints great potentials in comparison between direct- and indirect-developing enteropneusts in the context of deuterostome evolution.
Speaking of unconventional, you must have realized by now that I saved the most curious topic in hemichordate research to the last. Although a major class of hemichordates called enteropneusts receives much of the attention — and although this hemichordate conference so far could have been an enteropneust conference — a minor class called pterobranchs has always generated interesting questions. The living pterobranchs are represented by only two, mostly deep-sea, tube-dwelling genera: Cephalodiscus and Rhabdopleura. They used to be a thriving group, says Chris Cameron(Université de Montréal). He and his collaborators in paleontology are reaching consensus that graptolites, common fossils of the Paleozoic rocks, are nested within the Pterobranchia. So the two genera are quite literally living fossils. Excitingly, now he has a Mediterranean locality that provides a stable supply of the living pterobranchs, although the initial work is still ongoing. Cameron’s interest is to identify root conditions for hemichordates and ambulacularians. In that vein, he also reviewed stem enteropneusts from the Cambrian and Carboniferous localities (Cambrian stem enteropneusts were tube-dwelling! But, as Ken Halanych points out, so are the living Antarctic torquaratoriids) and described echinoderm-like mineralized ossicles in the living enteropneusts.
The pterobranch Cephalodiscus sp. (source = Halanych lab)
It probably becomes apparent by this point that no good work in biology is possible without systematics, whether talking about enteropneusts or pterobranchs. Gonzalez’s species of Schizocardium does not even have a name yet. The evolutionary narratives based on developmental traits — a major justification for why looking into hemichordates — entirely depends on the current phylogenetic scheme. Even the major ambulacularian-chordate schemes accepted by most workers today can be shaken up easily. With xenoturbellids falling out from the deuterostome stem, for example. Ken Halanych (Auburn University) has been instrumental toreorganize and clean up this areaover the last two decades, and his eloquent talk on hemichordate systematics reflected his naturalist spirit well. The audience was captivated by the giant torquaratoriid from Antarctica, for example. His pep talk for pterobranchs certainly upped the enthusiasm for this minor class by a notch for Cameron’s talk. Joie Cannon (University of California, Santa Barbara) extended Halanych’s theme by going through her PhD work that took her around the world several circles to collect specimens for molecular phylogeny (see her papers here, here, and here). In hemichordates, so many taxa remained undescribed or even yet to be discovered. There is not a single tome or even a global guide that the workers can refer to for species identification. Cannon’s work laid the track, but no train has rolled on it yet. Like many workers who work with rather uncharismatic invertebrates, there is an acute need for systematics. The field is waiting.
In conclusion of the meeting, Lowe led a roundtable discussion. Some points that stood out are: the Marine Biological Laboratory, Woods Hole, has been a center of research on S. kowalevskii. Lowe is consulting with the Lab for increasing accessibilities and improved facities that could accommodate more researchers. Su followed up on this by extending an open invitation to collaborate on P. flava in Taiwan as well. Cameron and Halanych proposed creating some kind of portal — a listserver would be a start. Green described the current challenges to maintaining a lab culture of S. kowalevskii in his home lab at Caltech.
Prospectus of Hemichordate Research
As a graduate student, I was thrilled to witness a community forming. The modern hemichordate research has clearly hit the critical mass that now some community efforts are required to carry it to the next stage. The Hemichordate Genome Project got the train moving, and more and more people are jumping on it to take a ride. The ever-increasing developmental information, coupled with evolutionary insights based on sound phylogenetic framework, should transform our understanding of this small phylum over the next decade or two. The meeting showcased enough of the momentum.
The very picturesque Monterey Bay, viewed from the Hopkins Marine Station (photo by author)
Simultaneously, some challenges still remain. In my view, the poor understanding of hemichordate natural history (taxonomy, anatomy, behavior, etc.) is holding back further progress. This gap in our knowledge is even more glaring today than ever, not least because favourite habitats of enteropneusts used for lab research appear to be shrinking rapidly. It echoes the outcry from biological communities in general: “We need more taxonomists!” On other counts, pterobranch development remains as a bottleneck. The debate on the deuterostome stem has not settled. None of the currently used hemichordate models has undergone a full lifecycle in lab.
Historically, the hemichordate research has relied on no more than several active labs working concurrently. So the meeting had the feel of a party in the neighbourhood: polite and friendly. No heated debate ensued even when philosophical differences were apparent. This civil attitude is welcome, but for the community to be vibrant, a controversy, too, may help. Wild speculations should also have a place in the field that is still in its infancy. One curious observation from the meeting is that all but one of those who compared hemichordates and vertebrates explicitly referred to Xenopus. Surely, Xenopus is a useful model, and has been the one of choice at the Gerhart and Kirschner labs. But would different predictions and interpretations be possible if compared with other vertebrate models? How should one identify deep homology and cooption event in that comparative framework? To create this kind of discourse, perhaps there were few opportunities for all of the participants to get together except for reception on the eve and banquet in the finale. All in all, though, the prospect is wide open, and the next meeting is being planned a couple of years from now. The second meeting will mark another memorable point in the history of hemichordate research. Until then, my new worm-loving friends!
Established by the British Society for Developmental Biology (BSDB) in 2014, the Gurdon/The Company of Biologists Summer Studentship scheme provides financial support to allow highly motivated undergraduate students an opportunity to engage in practical research during their summer vacation. Each year, ten successful applicants spend eight weeks in the research laboratories of their choices, and the feedback we receive is outstanding.
We will hear four stories from the 2016 class over the next four days, starting with Ji Hye Moon, who undertook a project in Richard Wingate’s lab in King’s College, London. You can read all the previous reports here.
The effects of transient gestational hypothyroidism on the development of foetal cerebellar nuclei
Me (Ji Hye Moon) at the far back with my colleagues from the Wingate Lab
As a graduate medical student with a previous degree in Middle Eastern Studies and Politics, I had not previously been exposed to scientific laboratory work. Progressing through the two years of pre-clinical studies, I surprised myself on just how much I enjoyed learning the science that underpins medicine and I became very aware of my lack of experience in research. I felt that I would benefit enormously from gaining exposure to this field, both in terms of my future competency as a clinician, as well as on my ability to pursue research further down in my career. As such, I felt very privileged to be able to start building up my skills and experience in lab-based research through the summer studentship this year.
The BSDB Gurdon Summer Studentship gave me the opportunity to spend 8 weeks this summer in the lab of Dr Richard Wingate in the Department of Developmental Neurobiology at King’s College London. I joined a project which aimed to examine the effects of transient gestational hypothyroidism on the development of the deep cerebellar nuclei in the foetal brain using mouse and chick models. I found this project particularly interesting as the effects of gestational hypothyroidism in the earlier stages of pregnancies are currently quite poorly understood, yet have been linked to postnatal cognitive deficits in the absence of gross malformations of the foetal brain.
The project used the drug methimazole, which is used to treat hyperthyroidism, in order to induce a transient hypothyroidism in pregnant mouse mothers to study its effect on the development of the foetal cerebellum at various embryonic and postnatal ages. All the brain samples were harvested before I started my studentship, so there were a lot of brain samples ready to be studied using immunohistochemistry (IHC) and in-situ hybridization (ISH) methods.
For the first part of the studentship I helped produce the wax and cryosections of the harvested brains necessary for IHC and ISH experiments. For this part of the work, I learned and honed the basic histology skills hands-on, through wax embedding, wax sectioning and cryosectioning of the mouse brain samples. Although this was quite repetitive work, I actually I found it really enjoyable and experienced a lot of satisfaction when I produced my first slide of two perfectly aligned ribbons each containing five slices of tiny, miniscule embryonic mouse brains.
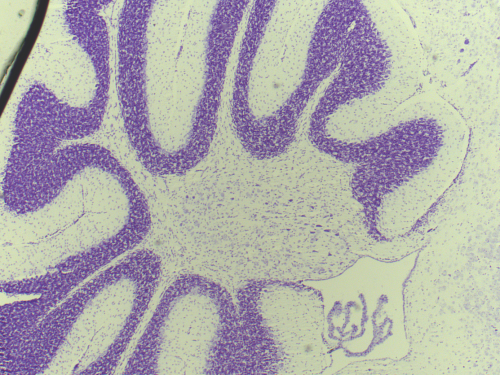
A section through the mouse cerebellum (Nissl stained) at postnatal age P23. The cerebellum is an exquisite structure with its distinct layers folded into folia.
Finally, the last few weeks of my studentship were spent producing nissl stains of the control and the experimental juvenile mice brains, which I then carefully studied to look for any subtle structural differences. I noted a potential decrease in the density of the Purkinje Cell Layer in the cerebella of the experimental group, but unfortunately was unable to complete a systematic comparison as my studentship came to an end.
I enjoyed the 8 weeks at the Wingate Lab and have benefited enormously from it. From an academic point of view, my science literacy level has improved immensely. Being exposed for the first time to the fields of mice and chick embryology and neuroanatomy as well as learning and practicing a wide range of experimental methods such as IHC and in-ovo electroporation, provided a steep learning curve that kept me focused and busy for the entire length of the studentship.
After spending two years almost exclusively in the lecture theatre, I really appreciated the hands-on learning experience and the stimulating environment. I learned a huge amount from attending the weekly lab meetings, talks and seminars from leading experts as well as interacting with my colleagues at the lab. Most of all, nothing could beat the depth of learning I gained from carrying out the experiments myself. It was fantastic to see the science come alive in front of my eyes and I found it a powerful way to learn and understand complex concepts. I was very lucky in that Dr Wingate encouraged me to tryout a variety of experimental techniques and always made time in his busy schedule for my questions.
The summer studentship was a rewarding experience on a personal level as well. It gave me a taste of what a career in research is like and an insight into the delights and the difficulties of research work. Ultimately, I’ve come to have a greater appreciation and admiration for research scientists and the enormous contribution they make to modern medicine and to the innovations that I will put into practice throughout my medical career.
I am very grateful to Dr Wingate, Dr Wilson, Margarita, Deviana, Tristan & Flo for being great hosts and teachers to me. A huge thank you in particular to Richard and the BSDB for making this opportunity possible for me!
The group of Chen Luxenburg at the Faculty of Medicine, Tel Aviv University invites applications for PhD student position.
Our laboratory is looking for excellent and highly motivated PhD students to study the role of the actomyosin cytoskeleton in skin development. Our goal is to understand how cytoskeleton derived signals regulate stem cell ability to create the skin epidermis during development, maintain it in the adult and repair it upon wounding. On top of molecular biology, tissue culture, advanced microscopy and mouse work we utilize state of the art technology that allows us to rapidly manipulate the function of any gene of interest in epidermal stem cells in utero. Several exciting projects are available for the successful candidates.
We offer state of the art laboratory with dynamic and international atmosphere and full financial support (tuition and stipend)
Candidates should hold Master’s degree in Biology/Life-sciences or related field. Preference will be given to candidates familiar with tissue culture techniques, microscopy and mouse work.
Interested candidates should email their CV and a brief paragraph describing their research experience and career plans to Dr. Luxenburg (lux@post.tau.ac.il)
 (No Ratings Yet)
(No Ratings Yet)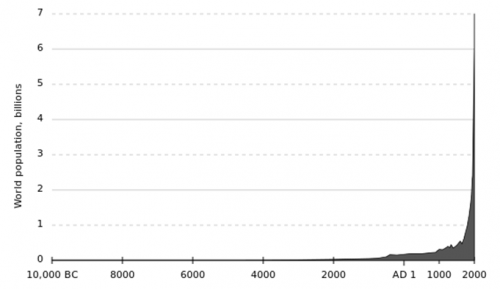

 (1 votes)
(1 votes)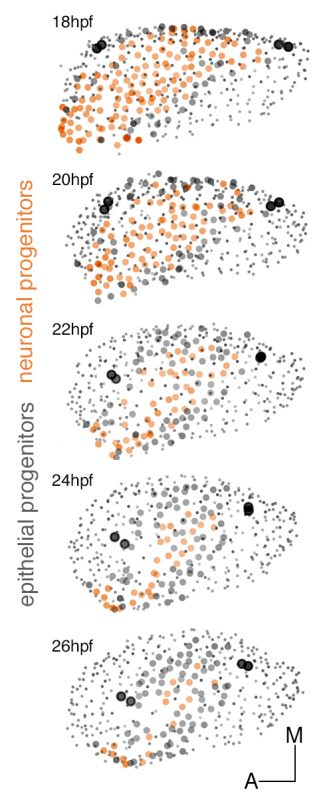
 (3 votes)
(3 votes)










{kind=link}
{kind=link}
{kind=link}
{kind=link}