It was spring in 2015 when I first met Dr. Stefan Schulte-Merker in Osaka, Japan. There, it became apparent that both of our laboratories had been interested in same gene and analyzed mutant mice and fish, which develop lymphedema as a consequence of lymphatic defects. Our group have long been interested in extra cellular matrix (ECM) biochemistry. We recently tried to screen functionally unknown ECM proteins and investigated their function using mutant mice. On the other hand, Stefan’s group has studied lymphatic vascular development, and screened novel lymphangiogenic factor using fish. We both took totally different approach at first, but finally reached same gene. In the meeting, we exchanged data and had a great discussion. However, the time was so limited that we could not have enough time for discussion. From then, I hoped to collaborate with Stefan, and fortunately I got a chance to visit to Stefan’s lab by successfully applying for a Company of Biologists Travelling Fellowship.
Münster from the air. Credit: Bernhard Kills, Wikipedia.
In 2016 April, I went to Münster in Germany. Münster is a beautiful city that has a lot of historical place and nature side by side. If you walk a little from city center, you can go to a lake Aasee and enjoy winds of nature. Every Wednesday and Saturday, an open-air market is held in front of Dome, and it soon became my favorite thing. Everyday, after having delicious bread and cheese for breakfast, I went to Stefan’s lab.
The main purpose of my stay was learning how to observe whole mouse embryo staining using ultramicroscopy technique (thanks to cooperation with Dr. Kiefer ). I also learned zebrafish experiments so I could learn merits using fish. During these experiments, I could share detailed practical information, which I could not get from research articles. Furthermore, we had several meetings. It was really important for me, because I could share ideas with vascular biologists. They provided me a lot of advice that I never thought about it. At the same time, I could return my opinions as an ECM biologist, which brings me to realize what is my strength point and what I have to study more. Now I think I can understand my research better than before.
“During these experiments, I could share detailed practical information, which I could not get from research articles. Furthermore, we had several meetings. It was really important for me, because I could share ideas”
I would like to thank Stefan and lab members for their hospitality. They always talked to me friendly and asked me to lunch. Many of them gave their time for me to teach experiments and to have meetings. Stefan kindly showed me around town. He took me to café several times so that we could talk not only about science itself but also about scientific things, like career or the system of scientific research in Germany. It helps me to think about my future as a scientist. After visiting to Stefan’s lab, I re-boost up my motivation in research, and I came up with new ideas. This collaboration was really profitable. I do not doubt that it will improve lymphatic vascular research.
Ectopic wings upon ectopic activation of JAK/STAT (Carles Recasens-Alvarez and Ana Ferreira, IRB Barcelona)
Researchers at IRB Barcelona identify a fundamental role of the JAK/STAT signalling pathway in the development and growth regulation of limbs in Drosophila.
Published in Nature Communications, the study paves the way to research into the function of this pathway in vertebrate development and its possible involvement in human congenital diseases.
Many of the secrets of life, such as how we become a certain size and shape, have been uncovered in studies performed over more than 100 years and involving animal models such as the fruit fly Drosophila melanogaster. Now, IRB Barcelona researchers headed by ICREA Professor Marco Milándisclose a new signal that participates in the specification and growth of fly wings.
In Nature Communications, the scientists conclude that the JAK/STAT signalling pathway, known to be tightly linked to inflammatory processes and tumour growth, determines where, when and how a wing develops in Drosophila. PhD student Carles Recasens, who will defend his thesis with the results of this study in January, has discovered that “JAK/STAT appears at key time points in the development of the appendage and that it collaborates with Wingless/Wnt, Dpp/BMP and Hedgehog in wing specification and growth”. These findings pave the way to studying the participation of JAK/STAT in human development and its possible implications in congenital diseases that involve limb malformation.
“Given the similarities in the molecules and the mechanisms involved in limb development in vertebrates and invertebrates, the fly is a very useful genetic model in which to identify new genes that potentially participate in limb development in vertebrates and their possible association with congenital diseases,” says Ana Ferreira, who has participated in the study. Marco Milán, head of the Development and Growth Control Lab, adds that “the patterns that determine how flies and humans are built are very similar and the basic molecular mechanisms have been conserved throughout evolution. We share a lot of basic biology and we increasingly find that what happens in flies also happens in humans”.
This work has identified three defined functions of JAK/STAT in fly development. First, it cooperates with Wingless (Wnt in humans) to specify where the wing will develop. Second, it helps cells that produce Hedgehog (Sonic hedgehog in humans) to survive and proliferate in order to induce the expression of Dpp (BMP in humans), a molecule that organises the patterning and growth of the whole wing. And third, it delimits the action of Dpp so that the wing grows in the right place. In summary, JAK/STAT controls the three main cell signals responsible for the specification and growth of limbs, both in vertebrates and invertebrates.
Carles Recasens and Ana Ferreira will be joining the recently opened Francis Crick Institute in London to undertake postdoc training, during which they will continue to address the fruit fly.
Reference article:
JAK/STAT controls organ size and fate specification by regulating morphogen production and signalling
Carles Recasens-Alvarez, Ana Ferreira and Marco Milán
Our latest monthly trawl for developmental biology (and other cool) preprints. See June’s introductory post for background, and let us know if we missed anything
2016 saw preprints in the life sciences really taking off…
…lots of preprints were deposited in December, predominantly on bioRxiv, though we also found a handful on arXiv, PeerJ, and our first entry from the recently launched Wellcome Open Research.
Tissue-wide changes in cell orientation pattern during the expansion of the histoblasts of the Drosophila pupal abdomen. Video S1 from Mangione & Martin-Blanco, 2016.
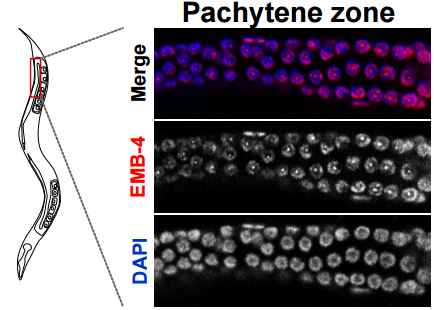
Localisation of EMB-4 protein in the germline of adult c. elegans, From Figure 2, Alkay, et al. 2016.
The Aquarius/EMB-4 helicase licenses co-transcriptional gene silencing. Alper Akay, Tomas Di Domenico, Kin Man Suen, Amena Nabih, Guillermo Eduardo Parada, Mark Larance, Ragini Medhi, Ahmet Can Berkyurek, Christopher J. Wedeles, Xinlian Zhang, Ping Ma, Angus I. Lamond, Martin Hemberg, Julie M. Claycomb, Eric Alexander Miska
Human pancreatic β cell lncRNAs control cell-specific regulatory networks. Ildem Akerman, Zhidong Tu, Anthony Beucher, Delphine Rolando, Claire Sauty-Colace, Marion Benazra, Nikolina Nakic, Jialiang Yang, Huan Wang, Lorenzo Pasquali, Ignasi Moran, Javier Garcia-Hurtado, Natalia Castro, Roser Gonzalez-Franco, Andrew F. Stewart, Caroline Bonner, Lorenzo Piemonti, Thierry Berney, Leif Groop, Julie Kerr-Conte, Francois Pattou, Carmen Argmann, Eric Schadt, Philippe Ravassard, Jorge Ferrer
Impact of regulatory variation across human iPSCs and differentiated cells. Nicholas E Banovich, Yang I Li, Anil Raj, Michelle C Ward, Peyton Greenside, Diego Calderon, Po Yuan Tung, Jonathan E Burnett, Marsha Myrthil, Samantha M Thomas, Courtney K Burrows, Irene Gallego Romero, Bryan J Pavlovic, Anshul Kundaje, Jonathan K Pritchard, Yoav Gilad
Reversible metamorphosis in a bacterium. Karina Ramijan, Joost Willemse, Eveline Ultee, Joeri Wondergem, Anne van der Meij, Ariane Briegel, Doris Heinrich, Gilles van Wezel, Dennis Claessen
Genetic variation and gene expression across multiple tissues and developmental stages in a non-human primate. Anna J Jasinska, Ivette Zelaya, Susan K Service, Christine Peterson, Rita M Cantor, Oi-Wa Choi, Joseph DeYoung, Eleazar Eskin, Lynn A Fairbanks, Scott Fears, Allison Furterer, Yu S Huang, Vasily Ramensky, Christopher A Schmitt, Hannes Svardal, Matthew J Jorgensen, Jay R Kaplan, Diego Villar, Bronwen L Aken, Paul Flicek, Rishi Nag, Emily S Wong, John Blangero, Thomas D Dyer, Marina Bogomolov, Yoav Benjamini, George M Weinstock, Ken Dewar, Chiara Sabatti, Richard K Wilson, J David Jentsch, Wesley Warren, Giovanni Coppola, Roger P Woods, Nelson B Freimer
Mindboggling morphometry of human brains. Arno Klein, Satrajit S. Ghosh, Forrest S. Bao, Joachim Giard, Yrjo Hame, Eliezer Stavsky, Noah Lee, Brian Rossa, Martin Reuter, Elias Chaibub Neto, Anisha Keshavan
The Fruit Fly Brain Observatory: from structure to function. Nikul H Ukani, Chung-Heng Yeh, Adam Tomkins, Yiyin Zhou, Dorian Florescu, Carlos Luna Ortiz, Yu-Chi Huang, Cheng-Te Wang, Paul Richmond, Chung-Chuan Lo, Daniel Coca, Ann-Shyn Chiang, Aurel A Lazar
NeuroNLP: a natural language portal for aggregated fruit fly brain data. Nikul H Ukani, Adam Tomkins, Chung-Heng Yeh, Wesley Bruning, Allison L Fenichel, Yiyin Zhou, Yu-Chi Huang, Dorian Florescu, Carlos Luna Ortiz, Paul Richmond, Chung-Chuan Lo, Daniel Coca, Ann-Shyn Chiang, Aurel A Lazar
MultiCellDS: a standard and a community for sharing multicellular data. Samuel H. Friedman, Alexander R.A. Anderson, David M. Bortz, Alexander G. Fletcher, Hermann B. Frieboes, Ahmadreza Ghaffarizadeh, David Robert Grimes, Andrea Hawkins-Daarud, Stefan Hoehme, Edwin F. Juarez, Carl Kesselman, Roeland M.H. Merks, Shannon M. Mumenthaler, Paul K. Newton, Kerri-Ann Norton, Rishi Rawat, Russell C. Rockne, Daniel Ruderman, Jacob Scott, Suzanne S. Sindi, Jessica L. Sparks, Kristin Swanson, David B. Agus, Paul Macklin
Figure 1. C.dipterum male (A, B) and female (C, D) individuals. (CE) Compound eye, (TE) Turbanate eye, (oc) ocelli.
The lab uses the development of insect eyes to investigate the mechanisms and gene networks that regulate tissue growth and final size. Most of our projects use Drosophila melanogaster as a model, but we have recently established two other systems, Episyrphus balteatus (the marmalade hoverfly) and Cloeon dipterum (a mayfly) to address how these mechanisms have changed during Evolution to give rise to the huge diversity found in insect visual systems.
Cloeon dipterum belongs to the Baetidae family of Ephemeroptera, which together with dragonflies and damselflies (Odonata) form the ancient group Paleoptera. Fossil records date ephemeropterans to the Carboniferous, being these insects the first ones that developed wings. This order has a hemimetabolous development, comprising three different phases: nymph, sub-imago and imago. The Baetidae family of mayflies, including C. dipterum, exhibits a striking sexual dimorphism, consisting in the presence of an extra pair of dorsal eyes in males. Females of this species harbour two different types of eyes: the lateral compound eyes and the three frontal ocelli, typical of insects whereas males also develop a pair of extraordinary enlarged compound eyes called “turbanate” due to their shape (Figure 1).
The mayflies have a life cycle that is divided in two main phases, an aquatic phase and a terrestrial phase. The nymphs are aquatic and they can live for several months in freshwater streams, undergoing several moults until they reach their final size. Then, they emerge from the water as sexually immature subimagos that have to moult once more to acquire their sexual maturity. Thousands of adult individuals form swarms and mate while flying some metres above the water. Finally, the females lay the fertilised eggs onto the surface of the water, where they will develop and later hatch as nymphs. One of the particularities of C. dipterum is that it is one of the few ovoviviparous Ephemeroptera species. Thus, once the adult female lays the eggs, they immediately hatch as tinny swimming nymphs (Figure 2).
Figure 2. C. dipterum nymphs. (A) Nymphs hatching from the eggs right after being delivered. (B) One hatchling few minutes after hatched. (C) Three-weeks-old male nymph.
Closing the life cycle in the lab was one of the first challenges we encountered. Due to space limitation and lack of infrastructure (we do not have a pond inside the institute to leave a swarm of mayflies mate freely), we have to perform forced copulation with them [1]. We grasp the female by the wings with a pair of forceps and place it in an inverted position. The male is placed against the female allowing direct contact between the external genitalia of both. The male genitalia forceps clasp around the female abdomen and the copula starts. The duration of the matings can vary from few seconds to several minutes (Figure 3). After the mating, we keep the females in a petri dish with some wet paper and wait for 15 days, until the embryos have fully developed inside the female abdomen. It is from this moment that we put the females onto the surface of water to allow them to deliver their offspring. We keep the nymphs in beaker glasses with water and air bubbling. These beakers are inside PET bottles to prevent the subimagos to fly away when they emerge from the water (Figure 4).
Figure 3. Couple of mayflies mating after forced copula
A typical day in a mayfly lab starts collecting the subimagos and imagos that have emerged and moulted during the night. These will be the specimens that we will use two days later for our crosses, so we put them in a falcon tube with some wet paper to maintain the humidity. As adults, mayflies do not feed, they lack mouth parts, so we do not need to worry about providing them with some food, we just need to avoid their desiccation. Nymphs, on the contrary, are very voracious. We need to feed them every morning with algae or vegetarian fish food flakes. It is very convenient to be in an institute with several zebrafish and medaka labs, as they are our main algae providers, just by scratching the algae that grow on the surface of the fish tanks. After taking care of the nymphs we can start performing some experiments. Having the culture established in the laboratory permits us to select the nymphs at the appropriate developmental stage to study the ontogeny of the turbanate eye and the genes that are involved in the process.
Figure 4. The culture in the lab. Several nymphs (B, purple arrows) pace on the glass walls. One subimago just emerged from the water (C, red arrow). Adults are getting ready to mate (D).
The techniques we use are not very different from the ones applied in other Evo-Devo and Developmental Biology labs working with well-established model organisms. Therefore, widespread procedures like imaging and gene expression studies are performed on a daily basis with the mayflies. Confocal microscopy and immunostainings are used to study the morphology of the turbanate retina and its associate brain centres along its development. Markers, such as antibodies for proliferation or specific cell types, serve us to investigate growth rates and patterning dynamics of the new organ (Figure 5). We have also generated some tools to investigate gene expression. We are able to search for genes in a transcriptome that we have produced and to look for their spatial expression using in situ hybridization at the desired developmental points. Our next goal is to establish functional tools in order to test the candidate genes we are identifying from our gene expression experiments.
Figure 5. We can use imaging techniques to visualise the development of the turbanate eye.
Setting up a new model in the lab presents some challenges, but it will help us to answer many questions regarding the origin and evolution of traits that appeared for the first time in insects, thus, every goal we accomplish is really rewarding and exciting.
References and Notes
McCafferty, W.P., and Huff, B.L., Jr. (1974). Parthenogenesis in the mayfly Stenonema fermoratum (Say) Ephemeroptera: Heptageniidae). Entomological news 85, 76-80.
Isabel Almudi holds a MSCA IEF 657732 fellowship funded by the H2020 program of the European Commission.
The Mayfly team. Carlos Martin-Blanco (master student, left) and myself.
More than 2000 years ago, Hippocrates (460-377BC) and Aristotle (384-322BC) described the human uterus as a series of chambers with a lining of tentacles or suckers. They believed that blood vessels connected the breast to the uterus allowing the pumping of breast milk into the uterine cavity. In this way, the embryo would be nourished by suckling on milk through the uterine tentacles whilst simultaneously being prepared for breast-feeding after birth. These early anatomical descriptions were the product of animal studies and philosophical theory, as laws and religion prohibited dissection of cadavers. Much of our present day understanding of pregnancy and the nature of embryo-uterine interactions is founded upon the documentation of human anatomy by artists such as Johannes de Ketham (1470-1491) and Leonardo da Vinci (1452-1519) and photographic evidence due to advent of microscopy and histology in the 16th century.
Left: drawing by Ketham, Right: drawing by Leonardo da Vinci, showing the milk vein connecting the breast and the uterus.
Widespread use of anatomical drawings and photographs is proof that visualization is key to the understanding of mechanisms governing basic biology. Oddly enough, though the embryo itself is much smaller (and less visible) than the uterus, we have a deeper understanding of how the fertilized egg develops into the blastocyst stage embryo. This is the result of laboratory techniques in blastocyst isolation and embryo culture combined with live imaging of fluorescent labeled proteins in the transparent early embryo. Missing is an understanding of the interaction between the embryo and its niche, the uterine lining, and how the embryo is guided to its site of implantation for attachment and future growth.
While working in the laboratory of Diana Laird on a project involving 3D imaging of the intact ovary (Faire et al., 2015), she and I contemplated how our imaging method could be applied to studying other organs. Meanwhile, my studies of the non-canonical Wnt receptor Ror2 identified a high level of expression in the mouse and human endometrium (Arora et al., 2014), and I wondered about the spatial distribution of this expression. Putting the two together, we tested the ovarian method of whole mount immunofluorescence and confocal imaging on the intact uterus. This worked well for neonatal uteri (which are not as thick and don’t have much muscle), but poorly in adult uteri. I made some key changes to the way I fixed the uteri by preserving their in situ length and incubating with antibodies for multiple nights, which led to better penetration of the antibodies.
The confocal imaging for three different antigens in a full-length mouse uterus requires ~5 hours of imaging time (about 18X2 tiles at a 10X magnification) and generates a data file size of 10-12GB. Using Imaris, an image analysis software, I reconstructed 3D visualizations of non-pregnant and pregnant adult mouse uterine epithelium.
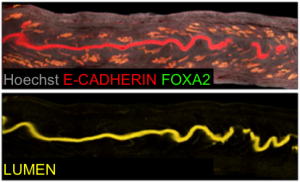
Generation of lumen only signal.
Our next goal was to isolate the uterine luminal epithelium independent of the glands. To this end, we computationally subtracted FOXA2+ glandular signal from the total E-CADHERIN+ epithelial signal. This gave rise to a lumen-only signal helping us generate the first 3D renderings of the mouse uterine lumen.
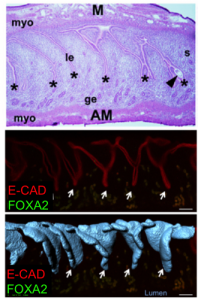
Top: H & E sections (Cha et al., 2014), Middle: optical Z slice, Bottom: overlap of 3D rendered folds and optical Z slice. Asterisks and arrrows indicate uterine crypts.
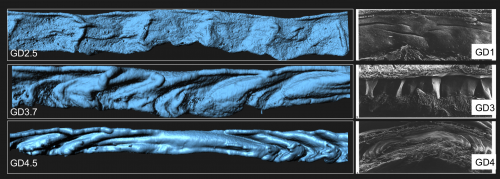
To our surprise, the 3D architecture of the uterine luminal epithelium was not static during preimplantation stages with many structural changes that would be impossible to discern using histology. Prior to implantation, the lumen undulated in stereotypical folds that were oriented perpendicular to the ovarian cervical axis. When overlapped with the optical Z-slices and compared with published 2D histological sections we determined that these folds are indeed structures known as uterine crypts.
We established a daily time course of the pattern of luminal folds during early pregnancy and ultimately their resolution around the site of implantation. Combing through literature, I also identified structural similarities between the luminal folding dynamics in my imaging and SEM images from the luminal side of rat uteri published in the 1990s (Winkelmann and Spornitz, 1997).
Left: Folds observed in mouse uterine lumen, Right: Uterine architecture observed with SEM images in pregnant rat uteri at different stages of pregnancy (Winkelmann and Spornitz, 1997).
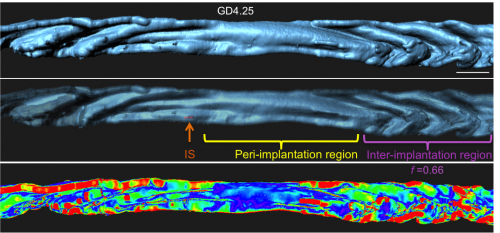
Next I faced the challenge of quantifying something that was quite obvious to the eye, but still needed measurement to confirm. We collaborated with the Biological Imaging Development Center at UCSF to work out a computational algorithm for quantifying the degree of luminal folding. With the surface renderings generated in Imaris, the algorithm calculated surface curvature and derived a simple expression for folding factor (f). The Matlab script for surface curvature interfaced with Imaris to depict the folds as a heat map where deep folds were painted red and flatter regions blue. This heat map helped us divide the entire uterine lumen into three segments – the implantation site (where the embryo is present), the peri-implantation region (flattened lumen) and the inter-implantation region (folded lumen).
Folds resolve at implantation site (IS).
We next used 3D imaging methods to elaborate on the function of uterine glands. Although ancient philosophers believed the uterus to be connected to the mammary gland, the breast and uterus were eventually shown to be anatomically distinct.
Depiction of uterine glands as floating structures in the stroma (Teixeira et al., 2008).
Additionally, the composition of the uterine milk was not composed of albumins, but rather, uterine secretions termed “the embryotrophe” secreted by the uterus’s own glands. In 2D histological sections, these glands appear as discrete epithelial structures floating in the stroma. We were keen to evaluate the clustering of glandular structures using previously described algorithms in the lab. Using FOXA2 as a marker for glands, our 3D imaging technique allowed us to show that that glands are actually branched structures connected to the uterine lumen by a small duct, giving the appearance of grapes on a vine. Surprisingly, the floating structures observed in 2D sections were connected to each other and to the uterine lining, when viewed in 3D. These glands are also present only at the antimesometrial side of the uterine lumen, where the embryo eventually implants in the pregnant uterus.
Glands are present on the antimesometrial side of the uterine lumen.
The ancient Greeks were right – uterine glands are still considered to be important for secreting substances that will nourish the embryo until the placenta is formed. Studies of farm animals and of mice have shown that glandular secretions are key to embryo development, as animals with defective glands are sub-fertile or infertile. However, no 3D imaging studies have been done to observe the organization of these glands or chart any changes in glands associated with implantation.
Top: Glands in non-pregnant uterus, Bottom: Bent glandular ducts in implantation stage uterus.
I noticed that the glands of the implantation stage uterus had a very characteristic organization. At this time, glandular ducts elongate and bend towards the site of implantation. This was a curious observation, considering that uterine glands are classically considered to be exocrine glands that secrete molecules into the uterine lumen (or down the trunk into the roots of the vine in my analogy). This ductal elongation would paradoxically increase the length of the path that glandular secretions have to travel before reaching the embryo. However, the glandular duct elongation also effectively brings the uterine glands in close spatial proximity to the stroma that will decidualize. These observations led us to hypothesize that uterine glands might secrete their factors into the stroma or into the closely associated vasculature and not just into the lumen, assisting in implantation and embryo growth.
All of this mouse work made us curious about the human uterine architecture. We were fortunate enough to obtain proliferative phase human endometrial hysterectomy samples through Dr. Linda Giudice in our department at UCSF. The glandular organization in humans is much more complex than in mice. Our method opens up possibilities to elaborate on the intricacies of epithelial organization in the human endometrium.
My vision is that this methodology will transform the way we view implantation. We will be able to elucidate the mechanisms by which implantation is disrupted in genetic mutants. Using these techniques, we are now beginning to ask questions about embryo spacing, crypt formation, and the interactions between the embryo and uterine epithelium as it travels through the lumen to find its site of attachment. This will lead to a better understanding of the conversation between the fetus and the maternal lining ultimately improving our approach towards – artificial reproductive technologies in the clinic, treatment of infertility and identifying novel targets for contraception.
Teixeira, J., Rueda, B. R. and Pru, J. K. (2008) Uterine stem cells. In Stembook(ed: L. Girard), (Internet). Cambridge (MA): Harvard Stem Cell Institute.
The tenth paper featured in this series comes from the first issue of Development for 2017, and uses computational modelling to investigate the importance of different cellular processes in spiral cleavage of the early embryo.
We caught up with the paper’s first author Miguel Brun-Usan and Isaac Salazar-Ciudad, group leader at the University of Helsinki in Finland, to find out more about the history of the work and how computational modelling can help developmental biologists.
The Salazar-Ciudad lab, with Miguel (first left) and Isaac (third left)
So Isaac, can you give me your scientific biography and the main questions your lab is trying to address?
ISC I studied Biology in the Autonomous University of Barcelona. Some months before finishing, I started to look for research group in evolution in Spain. I found Pere Alberch‘s research and I immediately got hookeded by his approach. He approached evolution not from the reductionistic population genetics approach (that I had studied in detail), but from the complementary approach of evolution and development. I met him and he suggested me to contact Ricard V. Solé (at the time at the Polytechnic University of Catalonia) and Jordi Garcia Fernandez (at the University of Barcelona) since he was at the time already quite ill. I started my PhD with Ricard and Jordi on mathematical models of gene networks in evolution and development. During that time I also started to collaborate with Stuart A. Newman and Jukka Jernvall.
After my PhD, I went to Helsinki University to work as a Marie-Curie postdoctoral fellow with Jukka Jernvall in tooth morphogenesis and evolution. In 2008 I moved to the Autonomous University of Barcelona as a Ramón y Cajal fellow. This position was supposed to be tenure-track but with the financial crisis in Spain it soon became clear that it would not be tenured. In 2011 I moved back to the University of Helsinki as a junior group leader and Finnish Academy Fellow.
“The questions we aim to address in my lab are how the way development works affects the direction of evolutionary change and how development itself evolves”
The questions we aim to address in my lab are how the way development works affects the direction of evolutionary change and how development itself evolves. Development determines which kind of morphological variation is possible due to genetic mutation, and then it has a strong influence, together with natural selection, on how the morphology is going to change in evolution. To study that we build computational models of how genes and cells interact in networks to construct the body. We do that for specific organs in close collaboration with experimental groups (we have been working with mammalian teeth, fly wings, fly segmentation and turtle carapace) but also in general. We also set those models to evolve in the context of populations under natural selection to acquire general principles about how gene networks and development as a whole evolves.
What is science and life like in Helsinki?
ISC It is great. The funding situation has been, until the last elections, rather good (especially compared to Spain). Helsinki has in proportion to its small size a rather large and active scientific life with many high quality researchers. There is a relatively high concentration of researchers working in evo-devo and development (again in proportion to its size) and a long tradition in developmental biology and in evolutionary biology.
Helsinki has a very high quality of life according to many non-Finnish magazines (Newsweek, Daily Telegraph, etc…). I am originally from Barcelona that, like many South and Central European cities, is rather dense and devoid of green areas. Barcelona is good for visiting but not so good for living. Helsinki is the contrary. Living in Helsinki is like living in a large nice park. Even in the poorest neighbourhoods, one is always a few steps away from a forest, a park or the sea. In spite of its parkland nature, public transportation is quite good. In summer the city centre has a very active cultural live with many free outdoor concerts and large crowds of cheerful people relaxing in the parks. Winter is great if you like winter sports and snow, as I do. In addition, almost everybody speaks English so it is quite easy to have a normal life in spite of being a foreigner.
And Miguel, how did you come to work with Isaac?
MBU Five years ago, I finished a Master’s Degree in Evolutionary Biology in the Complutense University of Madrid, which gave me a grasp of why evo-devo is relevant to understand evolution. As a person who has been fascinated by biological evolution since childhood, I quickly started to look for a PhD position in a research group working on morphological evolution. Unfortunately, these groups are very scarce in Spain.
However, through an active search, I found in a Spanish webpage for graduated students (RedIris) an advert from Isaac which seemed to fulfil perfectly my academic interests. I had read some of his papers, but I did not know him personally, so I immediately contacted him and we had several interviews. After these interviews, Isaac agreed I had the right motivation and background to work in his group. I had no previous experience in programming.
Developmental rules and cell processes considered in the work, from Figure 1, Brun-Usan et al. 2017
Your paper describes computational models of cleavage. How can this sort of modelling complement descriptive and experimental embryology?
ISC & MBU The main way in which models can help descriptive and experimental embryology is by discarding existing hypotheses. In our case for example, the model allows to prove that some existing hypotheses can not lead to the cleavage patterns we observe, it is logically impossible. On the other hand models allow to suggest which hypotheses, alone or in combination, could in fact produce the observed patterns. Models, however, do not show that these suggested hypotheses are right. This latter thing always requires, ultimately, experiments but the model suggests which hypotheses, and then which experiments, are the most promising.
“The main way in which models can help descriptive and experimental embryology is by discarding existing hypotheses”
Do you think modelling is currently well-used and appreciated in the field?
ISC & MBU No. For the last ten years there has been a steady increase in the interest, and in the number of articles, in modelling in developmental biology. This is certainly good news but the advantages of modelling are not yet widely recognized and there is a widespread misunderstanding and misuse of modelling. In our view there are several reasons for it.
First, a large proportion of developmental biology is still mostly concerned with genes and not so much with processes. Genes are crucially involved in most developmental processes but are not, on their own, enough to explain any of them. Hypotheses about how a system develops are only possible when considering how genes interact, how cells interact or both. Then for most systems we do not know enough to even start to build a reasonable hypothesis, and so we can not build any models either. In many developmental systems we know a lot, typically a long list of genes important for it, but we are missing some crucial data, like which cell behaviours are used (cell division, cell adhesion, apoptosis, etc…), that is necessary to make predictive models.
Second, the difficulty understanding what models are for is usually not due to difficulties understanding models or mathematics as such, but to difficulties understanding hypothesis-driven research. Most research in developmental biology is, in practice, not hypotheses-driven. A large proportion of the research is devoted to identify interactions (usually genetic) that are necessary for the development of one or several organs. This exploratory work is certainly necessary but there should also be work (and there is in fact some) proposing hypotheses about how the identified interactions get coordinated to explain how a system develops, and proposing how to test these hypotheses. Models are helpful in these latter two things. Each model is simply a mathematical implementation of an hypothesis but, because of being mathematical, such implementation provides accurate predictions of how developmental variables of interest (for example organ shape) change over developmental time. These accurate predictions can then be compared with the observed development to falsify, or not, the hypotheses on which the model are built.
“For the last ten years there has been a steady increase in the interest in modelling in developmental biology. This is certainly good news but the advantages of modelling are not yet widely recognized and there is a widespread misunderstanding and misuse of modelling”
By exploring which hypotheses are consistent with experiments and which ones are not one learns. Models are not absolutely necessary for this hypothesis-driven research but they are quite useful for it and if one is not used to this hypothesis-driven research it is not possible to appreciate and use models. This relates to a common criticism to models: that models usually do not include all available data, or that their results are not totally realistic. This may be more the case for some models than for others, but the fact is that no model is perfect simply because no hypothesis is. It is only by making hypotheses and falsifying them that one can learn.
Third, many developmental biologists feel uneasy with programming or mathematics. Although the actual mathematics involved are often relatively simple (I for example taught myself the necessary maths for the models I do) this has created a niche for non-biologists to work in modelling in developmental biology or biology in general. From my experience as a reviewer that is often quite sub-optimal. This is specially the case for computer scientists and engineers since they are usually even less familiar with hypotheses-driven research and, in addition, their lack of a deep understanding of biology precludes them from evaluating which questions are relevant for biological theory and which ones are not. This adds to the clichés about the difficulty in communication between the people making the models and the experiments. I have avoided those by being myself a biologist and working only with experimentalists doing hypothesis-driven research.
What led you to focus on spiral cleavage in particular?
ISC & MBU Miguel was quite fascinated by the study of theoretical morphospaces as are often done in palaeontology. In these studies one can reproduce all the morphologies in a clade, for example the shape of the shell as in Raup’s classic article, by giving specific values to an equation. This equation is, however, merely phenomenological; it does not include any understanding of the processes by which those morphologies are built. Miguel wanted to do something similar but based on the rules of development, as Isaac had done in the past on a more restricted scale. The aim was then to be able to pinpoint which aspects of development have to change to reproduce the variation between many species. Miguel had a long lasting interest in very early development and we realized that, in fact, the developmental stage for which there is information about more species is cleavage. Originally Miguel wanted to do it for all kinds of cleavage but we set first for spiral cleavage because it is the most common cleavage type at the phylum level.
The combination of rules and their parameters that can reproduce spiral cleavage, from Fig 2, Brun-Usan, et al. 2017.
Can you give us the key results of your paper in a paragraph?
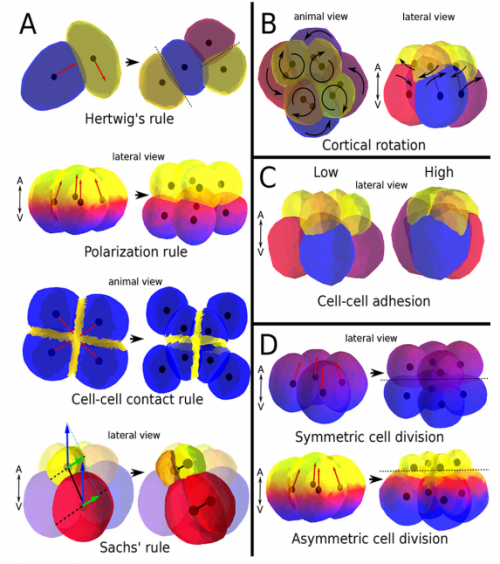
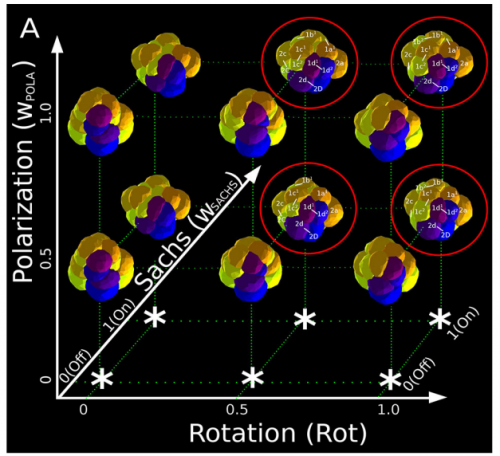
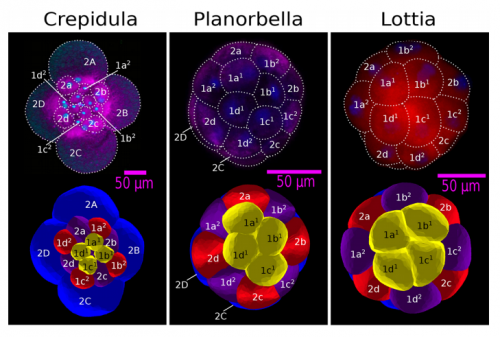
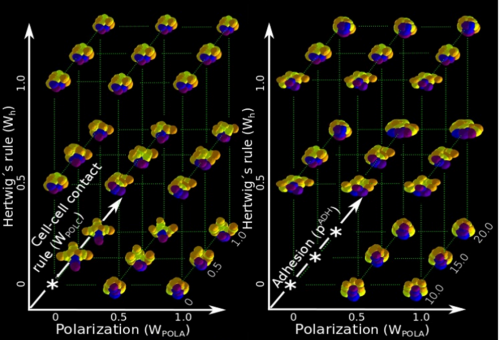
ISC & MBU Different cellular processes have been hypothesized to be responsible for the development of the specific spatial arrangement of blastomeres in the spiral blastula. These include the orientation of cell division according to an animal-vegetal gradient, according to cell’s main axis (Hertwig’s rule), according to the contact areas between cells or orthogonally to previous divisions (Sach’s rule). We use a computational model of cell and tissue bio-mechanics to implement the different existing hypotheses about how the specific spatial arrangement of cells in spiral cleavage arises during development. We found that cell polarization by an animal-vegetal gradient, a bias to perpendicularity between consecutive cell divisions (Sachs’ rule), cortical rotation and cell adhesion, when combined, reproduce the spiral cleavage while other combinations of processes can not. Specifically, cortical rotation is necessary in the 8-cell stage to displace all micromeres into the same direction, being this displacement random in direction if only cell adhesion is included. By varying the relative strength of these processes we reproduce the spatial arrangement of cells in the blastulae of seven different species (four snails, two polychaetes and a nemertean).
Comparing real embryos (top row) with simulated ones (bottom row), from Fig 3, Brun-Usan, et al. 2017.
And what does your model suggest about the relationship between intercellular communication and spatial arrangement?
ISC & MBU One of the article’s results is that, in fact, the spiral cleavage, at least in its most generic form and also for the species we consider, does not require of any inductive events between cells (no diffusion of molecular factors between cells required). In other words, the relative spatial arrangement of cells does not require from cell communication. This does not mean that this communication does occur, we know it occurs in many species and this communication may be required to specify cell fate and, perhaps, for spiralian cleavages that depart significantly from the generic one.
Can your work tell us something about the evolution of cleavage patterns in animals?
ISC & MBU Bibliography shows us that most animal clades use the same set of basic (and evolutionarily old) cell processes for building up their cleavage patterns. Thus, even though our work is only concerned with spiral cleavage, our results can shed light in other, non-spiralian cleavage patterns. First, non-spiralian patterns have to be generated by other spatio-temporal combinations of similar cell processes, and variation within these non-spiralian patterns are very likely produced by variation in the strength of these other rules. Second, as is the case for the spiral pattern, some patterns may be more likely to arise from development than others (they require less cell processes, and/or less coordination between them). This may account for the uneven distribution of cleavage patterns among animals.
“Most animal clades use the same set of basic (and evolutionarily old) cell processes for building up their cleavage patterns. Thus, even though our work is only concerned with spiral cleavage, our results can shed light in other, non-spiralian cleavage patterns”
As someone who used to work in a ‘wet’ lab, I’m wondering about the process of modelling – how long does it take to run, and when do you know that you have an interesting result? Do you get ‘eureka!’ moments?
MBU I would like to make here a clear distinction between the model (SpiralMaker) itself and the in silico experiments we have performed with this model.
The model (or more precisely, its more general version EmbryoMaker), comprises more than ten thousand lines of programming language, organized in different inter-dependent modules. Most of it is not used in this article however. Such a big code has been built by different people working in Barcelona and in Helsinki during four years (specifically, I programmed many parts of the code relevant for non-epithelial cells). The most challenging aspect of this stage was to implement the different biological phenomena in a mathematical model and to make the model work as a coherent whole. Finally, we had to check that each implemented biological phenomenon worked realistically, but there was little room for Eureka! moments at this stage.
Some of the combinations of rules that failed to give spiral cleavage patterns, from Fig S2, Brun-Usan, et al. 2017.
Once we had this computational model ready to use, we performed the two in-silico experiments described in the paper. First, we had to design such experiments at conceptual level (what to simulate, which phenomena to include, until which stage should one run the simulation, which data to collect, how to compare results …) because normally there is no previous experimental protocols as in wet-lab procedures. This is a rather creative stage of the project. Then, the bulk of simulations (each one taking several hours of computation time) was automatically run in order to create the theoretical morphospace. It was really exciting to look for first time the resulting simulated embryos, and to realize that they were very similar to real ones in visual appearance. This early subjective appreciation was later confirmed by quantitative measures of the morphological similarity between the real and the simulated embryos (for some species, the similarity was 100%). This was the real Eureka moment of our research !
“It was really exciting to look for first time the resulting simulated embryos, and to realize that they were very similar to real ones in visual appearance”
What next for you after this work, Miguel?
MBU I have recently presented my Doctoral Thesis (November, 2016) in the Autonomous University of Barcelona, whose main chapters were devoted to the modelling of spiral cleavage. Currently, I am looking for future (Post-Doc) projects in which I could take advantage of the abilities acquired during my PhD and that would be in consonance with my intellectual interests. In that sense, I would like to continue in Evo-Devo modeling, specially in its more theoretical side. This is because I think that this theoretical/modelling approach is the fastest, cheapest and simplest way to address many questions of capital interest in biology (e.g. how biological complexity evolves, or how development itself evolves …).
And where next for the Salazar-Ciudad lab?
ISC At the moment I have several lines of research in the lab. One is taking a similar approach that in here but in mammalian tooth development (system in which I have been working for many years). One member of my group is using EmbryoMaker (a general modelling framework developed by us, described in a previous Node post) to implement a more realistic tooth model. We have built several tooth models in the past but their bio-mechanics were not precise enough to explore in enough detail the early tooth morphogenesis. This person is also making bio-mechanical experiments on tooth germs to contrast the model predictions.
In another line of research we are using a fly wing model we published in 2015 to explore how fly morphology changes with temperature. In yet another line of research we are using an existing model of development and evolution in tooth morphology to measure how accurately the statistical approach of quantitative genetics (mostly through Lande’s equations) holds for complex multivariate morphologies that are the result of a complex developmental process.
Finally we are using the EmbryoMaker in a ensemble approach in which we build a huge number of random networks and check which embryo morphologies they produce. This apparently crazy approach has the advantage that it may allow us to identify in an unbiased way general structural principles that gene networks need to fulfil to be able to produce complex morphologies. I am also starting to look for a new place to work.
University of St Andrews, School of Biology, Scottish Oceans Institute, Salary: £32,004 per annum, Fixed Term: 3 years, Start: 1 March 2017 or as soon as possible thereafter
We are seeking a post-doctoral researcher to work in the laboratory of Dr David Ferrier at the University of St Andrews for three years. The aim is to understand the regulatory mechanisms controlling the homeobox genes of the ParaHox cluster in invertebrate chordates (amphioxus and sea squirts), largely via means of reporter gene transgenics in the ascidian Ciona intestinalis (see Garstang et al (2016) BMC Evolutionary Biology 16:57). The focus will be on general regulatory mechanisms operating across the ParaHox cluster (with an initial focus on TCF/Lef and CTCF transcription factor binding sites) in order to better understanding how these major developmental control genes build animal nervous systems and guts as well as revealing general principles of gene cluster control. The ideal candidate will be an enthusiastic developmental biologist with a strong interest in evolution, with experience in molecular biology and microscopy. A PhD is essential.
The work will be based at the Scottish Oceans Institute, Gatty Marine Laboratory at the University of St Andrews, Scotland.
The University of St Andrews is committed to promoting equality of opportunity for all, which is further demonstrated through its working on the Gender and Race Equality Charters and being awarded the Athena SWAN award for women in science, HR Excellence in Research Award and the LGBT Charter; http://www.st-andrews.ac.uk/hr/edi/diversityawards/.
Interviews will be held during the last 2 weeks of January 2017
We are seeking a highly motivated and ambitious candidate to join our lab facilities funded jointly by the CPR and DanStem
The Danish Stem Cell Center (DanStem) and the Novo Nordisk Foundation Center for Protein Research (CPR) at Faculty of Health & Medical Sciences at the University of Copenhagen are looking for a sequencing specialist to join the DanStem and CPR lab facilities starting February 2017 or upon agreement.
DanStem comprises of two sections: The Novo Nordisk Foundation Section for Basic Stem Cell Biology, where we address basic research questions in stem cell and developmental biology (BasicStem). The second Section for Strategic Translational Stem Cell Research and Therapy (TransStem) is focused on the translation of promising basic research results into new strategies and targets for the development of new therapies for cancer and diabetes. Find more information about the Center athttp://danstem.ku.dk
CPR promotes basic and applied discovery research on human proteins of medical relevance. The vision of CPR is to be the world leading center in combining the power of integrative protein technologies and their application to accelerate understanding of the biological processes underlying health and disease. For more information about CPR, please visit http://www.cpr.ku.dk
We are seeking a highly motivated and ambitious candidate to join our lab facilities funded jointly by the CPR and DanStem.
Job description
The next generating sequencing (NGS) specialist will focus on running the new DanStem/CPR sequencing facility and providing assistance to users with processing their data. The work involves programming and maintenance of a liquid handling robot and sequencer. The major responsibilities of the position will be to optimize robotic applications to NGS and to map user’s data. In addition, stocking and supplying consumables for these and any additional equipment purchased for the facility. The applicant will also be assisting users in the processing and storage of their data. The applicant will be expected to push forward new approaches to single cell sequence and other related techniques.
Your profile
– A degree in biotechnology, molecular biology, bioinformatics, or appropriate biological or biomedical science with either PhD or MsC.
– Candidate must have experience with both dry (computer based) and wet (laboratory) work.
– Previous experience in Java programming and NGS bioinformatics (mapping of NGS data) would be a big advantage
– Theoretical and practical knowledge of Next Generation Sequencing sample preparation workflows and applications would be a plus.
– Proven ability in single cell analysis or PCR would be a plus.
– Good English communication skills, both oral and written, are prerequisite for the successful candidate
We offer
– Stimulating, challenging and multifaceted research environment
– Possibility for continued education and training
– Attractive employment conditions
– Centrally located work place
Employment conditions
The employment has an initial duration until the end of 2020 with a possibility of extension. The employment is scheduled to start 2017 or upon agreement with the chosen candidate. The place of work is at DanStem, University of Copenhagen, Blegdamsvej 3B, Copenhagen.
Salary, pension and terms of employment are in accordance with the provisions of the collective agreement between the Danish Government and AC (the Danish Confederation of Professional Associations). In addition to the basic salary a monthly contribution to a pension fund is added (17.1% of the salary).
The application must include
1. Motivation letter
2. Curriculum vitae incl. education, experience, previous employments, language skills and other relevant skills
3. Copy of diplomas/degree certificate(s)
Questions
For further information about the position please contact Professor Joshua Brickman,joshua.brickman@sund.ku.dk.
Application
Your application must be submitted in English by clicking “Apply Now” below or via this advertisement found on http://employment.ku.dk.The University of Copenhagen wishes to reflect the diversity of society and welcomes applications from all qualified candidates regardless of personal background.
Only applications received in time and consisting of the above listed documents will be considered.
Applications and/or any material received after deadline will not be taken into consideration.
Application deadline: January 29, 2017
Founded in 1479, the University of Copenhagen is the oldest university in Denmark. It is among the largest universities in Scandinavia and is one of the highest ranking in Europe. The University´s eight faculties include Health Sciences, Humanities, Law, Life Sciences, Pharmaceutical Sciences, Science, Social Sciences and Theology. www.ku.dk
This Editorial originallyappeared in Development, Volume 144, Issue 1
Olivier Pourquie
The past 15 years have witnessed major evolutions in the field of developmental biology – in imaging technology, genome editing, development of new ‘model’ systems and many other areas. Much of the action in our domain is now taking place in the stem cell field. Since I took over as Editor in Chief of Development, I have encouraged submission and publication of the best possible stem cell papers in the journal. We created a dedicated ‘Stem Cells and Regeneration’ section and have tried to promote the area in our front section (review-type content) with the recruitment of Caroline Hendry, our Reviews Editor dedicated to the stem cell field. More recently, we have extended this initiative to try to capture papers relating to human developmental biology, a field that has grown beyond its descriptive anatomical roots largely thanks to the development of stem cell-based technologies such as organoids; from 2017 onwards, these papers will also be collected together in a special ‘Human Development’ section of the journal. To demonstrate our commitment in these fields, we have published special issues on stem cells and regeneration in 2013 (http://dev.biologists.org/content/140/12) and on human development in 2015 (http://dev.biologists.org/content/142/18), which proved to be great successes.
We have also built a team of editors that include world leaders in stem cell biology. To further promote Development in this area, we recruited Melissa Little, who pioneered kidney organoids, to act as a guest editor for one year (2016), focusing on putting together a Special Issue on organoids to be published in early 2017. Finally, in 2014 and 2016, we organized very successful dedicated meetings on the theme ‘From Stem Cells to Human Development’. The 2016 meeting showcased the newest developments in the field of human developmental biology, including progress in emerging technologies such as organoids, bioengineering and single cell analyses (Craft and Johnson, 2017). We hope to continue this series of meetings in 2018, so look out for further announcements.
This editorial strategy – to promote the stem cell field – was initiated in 2010 and it is now time to look back and examine whether our efforts have been successful. But how do we measure success? Like large sectors of the community, I and Development’s editors feel that inappropriate prominence has been given to the Impact Factor (IF) (Pourquié, 2013). Development’s IF is essentially stable, and the journal performs well in other metrics, particularly the cited half-life, reflecting the longevity of our papers (see http://dev.biologists.org/content/about#metrics). Looking at a more granular level, the Stem Cells and Regeneration section of the journal (which now represents roughly one quarter of the research papers published) outperforms other research content in terms of citation levels, as do Techniques and Resources papers. Looking at other measures of journal performance, download and altmetrics data also speak to the success of our stem cell strategy, with stem cell papers typically among the most accessed research content each year. Along with the support we have received from the community (Pourquié and Brown, 2016a), these metrics give us confidence that our strategic focus on the stem cell field has been justified.
However, this is not to say that we have forgotten other aspects of developmental biology, or that we do not see and value the significant advances being made in many areas of the field. The plant field continues to fare very well in the journal – plant papers rank alongside animal stem cell papers in terms of citation rates – and we recently published a Special Issue dedicated to plant development (http://dev.biologists.org/content/143/18) in honour of the pioneering plant developmental biologist Ian Sussex.
Looking forwards, we see a growth in the areas of biophysical, mechanical, quantitative and computational developmental biology. 2017 sees the 100 year anniversary of the publication of D’Arcy Thompson’s landmark book On Growth and Form (Thompson, 1917) and we are planning to mark this occasion with a Special Issue of the journal entitled ‘On Growth and Form – 100 years on’ – which will celebrate the advances we have made in the past century, and look forwards to the future challenges of the field. We’re delighted to announce that this issue will be guest edited by L. Mahadevan (Harvard University), working alongside our longstanding editor Thomas Lecuit (see http://dev.biologists.org/content/special-issue-growth-and-form for more information). More generally, we hope that Development’s pages reflect the broad (and growing) scope of our field, and showcase some of the most exciting advances being made in labs across the world.
To continue to attract such papers, we need to respond to community concerns around the publishing process. To this end, we have recently introduced several important changes to the reviewing process. We now provide more explicit reviewing guidelines and forms, aiming to streamline the reviewing process as much as possible. Importantly, we have now introduced cross-referee commenting in the review process (Pourquie and Brown, 2016b). This means that once the reviews are completed, the reviewers are able to see each other’s reviews and have a 48 h window in which to provide extra feedback to help the editor make a decision. Although it is still early days for this new system, we feel it will help generate more consensual reviews and shorten the time to process papers. In addition, we are looking at ways of making it easier for authors to submit their work to Development. We recognise that you have a wide choice of journals to which you can submit, and we hope to minimise the effort required to send your paper to us. Although our Guide to Authors details specific formatting guidelines, we aim to be as flexible as possible in terms of article format at initial submission, and are working on streamlining this process further; we welcome your feedback on what changes we could make to improve the author experience.
Finally, preprints are also becoming a hot topic in life-science publishing, and Development has adopted a preprint-friendly policy. Preprints are manuscripts that are deposited in a public repository such as bioRxiv (http://biorxiv.org/) prior to publication. These manuscripts, which usually have not yet been peer-reviewed, are freely accessible to the entire scientific community. In 2015, we made it possible for authors submitting to Development to have their paper automatically deposited in bioRxiv, making it public in a matter of days. Now, you can also submit to Development through the bioRxiv submission system. This preprint model has been in place for more than 20 years in the maths and physics communities and it offers many advantages. Most importantly, research results are not held up from the scientific community for months or years due to the ever-increasing length of the review process. Thus, the data are immediately available and not hidden behind a paywall. This also offers the scientific community an opportunity to comment and help the authors consolidate the work prior to formal journal publication. In the past year or so, the number of preprints submitted to bioRxiv and other preprint servers has exploded and this trend is unlikely to slow down. The Node now posts a monthly list of new developmental biology-related preprints, making it easier for you to find the latest research in your field. Preprints have a digital object identifier (DOI) and are thus citable, and there is increasing support for allowing (and encouraging) preprints to be listed and considered in evaluation of grants and CVs. Although there is currently no direct conflict between preprint servers and traditional journals, the increasing use of preprint repositories will clearly impact the publishing world and as a community we need to start thinking about the future of scientific publications. Through what feels like a period of change, Development will strive to make publishing a less painful process for authors, and a richer experience for readers. Although Bob Dylan clearly wasn’t talking about either developmental biology or scientific publishing, the times certainly are a-changin’ for all of us.
I would like to conclude by thanking the team of academic editors who deserve great credit for their dedication and enthusiasm for the job, and our editorial board for their engagement and support. I particularly want to acknowledge Melissa Little for her great contributions as a guest editor. I also thank the Development staff: Editorial Administrators Hannah Morton and Debbie Thorpe; Production Editors Andrea Bowden, Colin Davey, Jane Gunthorpe and Lindsay Roberts; the Executive Editor Katherine Brown and Reviews Editors Seema Grewal and Caroline Hendry; the Node’s Community Managers Catarina Vicente, who left the Company in June, and her replacement Aidan Maartens; as well as the company’s production department and Publisher Claire Moulton. I also thank everyone who has reviewed for the journal this year (see supplementary information for a full list of our referees from the past 12 months) – without your assistance, the journal would cease to function. We look forward to a successful 2017 and wish all our authors, reviewers and readers the same.
Here are the highlights from the new issue of Development – the first one of the year. Happy reading…and Happy New Year!
Embryos rewired: the changing metabolome of early embryogenesis
During early mammalian embryogenesis, the developing embryo must adapt to changing metabolic demands and substrate availability. It has long been thought that a metabolic shift from glycolysis to OXPHOS takes place during this period and may be linked to the onset of choroallantoic branching, a major milestone in placental development that increases nutrient availability via the maternal circulation. But a simple shift from glycolysis to OXPHOS cannot explain how the array of macromolecules that are required to fuel cell proliferation are made, since OXPHOS produces mainly cellular energy and little else. Now, on p. 63, Yoshifumi Yamaguchi, Masayuki Miura and colleagues revisit this theory using state-of-the-art mass spectrometry techniques to survey the carbon flow of intracellular metabolites in the whole mouse embryo from embryonic day (E) 8.5 to E10.5, the period in which extensive choroallantoic branching occurs. The authors first establish the metabolic profile of the embryo during this period and show that, while metabolites indicative of OXPHOS do increase over this period, this is not accompanied by a decrease in glycolysis. Rather, the end product of glycolysis, lactate, also increases markedly from E8.5 to E10.5. The authors observe a decrease in the activity of phosphofructokinase-1 and go on to show how this results in the redirection of glucose into the pentose phosphate pathway, which is key for biomass production. This study provides insight into the dynamic metabolic profile of the developing embryo and sheds light on the long-standing question of whether and how a metabolic shift occurs during choroallantoic branching.
A new rule for spiral cleavage school
Spiral cleavage – the process by which cells of the early embryo divide and spiral around the pole-to-pole axis of the embryo – is the most common mode of animal development. Many hypotheses exist to explain how the precise spatial arrangement of cells is coordinated during this process, but to date there has been no systematic approach to verify which, if any, of these hypotheses are true. Now, on p. 54, Isaac Salazar-Ciudad and colleagues construct a computational framework in order to simulate early spiral cleavage behaviour. Using this model, they are able to constrain the behaviour of cells with existing hypotheses so as to determine which are important for the emergence of spiral cleavage and which are not. The authors find that none of the hypotheses proposed over time can produce the precise arrangement of cells observed during spiral cleavage, but that a small subset of them can do so if combined. Specifically, animal-vegetal polarization of cell division, Sachs’ rule in which cell division is oriented perpendicularly to the previous cell division, cortical rotation and adhesion are the main contributing variables to spiral cleavage. Finally, the authors show that their model can be used to generate a range of different embryo geometries corresponding to what is seen in seven different spiralian species. This elegant study highlights the power of computational approaches in understanding developmental processes, and brings insight into the specific parameters that govern spiral cleavage.
A supporting (cell) role for Wt1 in gonad development
The developing mammalian gonad comprises multiple different cell types, each with unique and important roles. Among these are the germ cells, which eventually become gametes and are crucial for generational inheritance, as well as supporting cells and steroidogenic cells, which support and nourish the gametes and provide important hormonal regulation, respectively. Despite the importance of supporting and steroidogenic cell types, the molecular mechanism that leads to their specification remains unclear. In this issue, on p. 44, Fei Gao and colleagues uncover a mechanism by which the expression of Wilms’ tumor 1 (Wt1) directs the lineage specification of supporting cells via the suppression of steroidogenic factor 1 (Sf1). Sf1 usually directs the specification of steroidogenic cells; however, the authors show that Wt1 binds directly to the promoter region of Sf1 in both sexes, supressing its expression. Deletion of Wt1 in the mouse undifferentiated genital ridge somatic cells before sex determination completely blocked the differentiation of the supporting Sertoli (in male) and granulosa (in female) cells, and resulted in the differentiation of steroidogenic cells instead. This study provides novel insight into somatic cell differentiation during gonadal development and provides a molecular mechanism for the specification of the supporting cells in both sexes.
PLUS:
The times they are a-changin’
The past 15 years have witnessed major evolutions in the field of developmental biology – in imaging technology, genome editing, development of new ‘model’ systems and many other areas. In his Editorial, Development’s Editor in Chief Olivier Pourquie highlights how Development has evolved with these changing times and discusses the journal’s plans for the future.
Towards a CRISPR view of early human development: applications, limitations and ethical concerns of genome editing in human embryos
The application of CRISPR-Cas technology to human cells has evolved in parallel with increasingly powerful methods of cell culture and analysis, and it is now possible to modify the genome of a human embryo in a highly efficient and specific way. In their Spotlight article, Alvaro Plaza Reyes andFredrik Lanner summarize the CRISPR-Cas genome editing system and discuss its potential applications and limitations in human pre-implantation embryos, and the ethical considerations thereof.
From stem cells to human development: a distinctly human perspective on early embryology, cellular differentiation and translational research
In late September 2016, over 100 scientists with common interests in human development, disease and regeneration gathered for The Company of Biologists’ second ‘From Stem Cells to Human Development’ meeting, which was held in historic Southbridge, USA. In their Meeting Review, April Craft andMatthew Johnson highlight some of the exciting new findings that were presented, and discuss emerging themes and convergences in human development and disease that arose during these discussions.
Understanding development and stem cells using single cell-based analyses of gene expression
In recent years, genome-wide profiling approaches have begun to uncover the molecular programs that drive developmental processes. In their Review, Patrcik Cahan and colleagues discuss how single-cell RNA sequencing has provided key insights into mammalian developmental and stem cell biology, emphasizing the analytical approaches that are specific to studying gene expression in single cells.


 (2 votes)
(2 votes)
 (No Ratings Yet)
(No Ratings Yet)




 (18 votes)
(18 votes)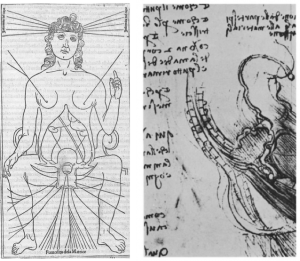







 In late September 2016, over 100 scientists with common interests in human development, disease and regeneration gathered for The Company of Biologists’ second ‘From Stem Cells to Human Development’ meeting, which was held in historic Southbridge, USA. In their
In late September 2016, over 100 scientists with common interests in human development, disease and regeneration gathered for The Company of Biologists’ second ‘From Stem Cells to Human Development’ meeting, which was held in historic Southbridge, USA. In their  In recent years, genome-wide profiling approaches have begun to uncover the molecular programs that drive developmental processes. In their
In recent years, genome-wide profiling approaches have begun to uncover the molecular programs that drive developmental processes. In their {kind=link}
{kind=link}