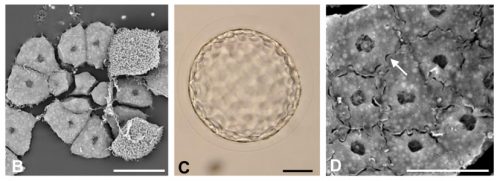
Here are the highlights from the new issue of Development…the last one of the year!
SETting chromatin state through transcription
Setd5 is a poorly characterised murine member of the SET domain family, generally associated with histone methyltransferase activity. However, the closest homologues of Setd5 are thought to be catalytically inactive, and have instead been associated with the regulation of histone acetylation levels at genes. On p. 4595, Anna Osipovich and colleagues generate Setd5 mutant mice and embryonic stem cells (mESCs). Setd5 homozygosity is lethal, with mutant embryos failing to survive beyond E10.5. Phenotypically, mutants display multiple defects, most notably in the cardiovascular system. Globally, cell proliferation is impaired and apoptosis increased. The mESC system reveals phenotypes consistent with the in vivo observations, including impaired differentiation down the cardiac lineage, while RNA-seq analysis shows that over 10% of coding genes are dysregulated in mutant cells – including key genes involved in cardiovascular development. Setd5 interacts with members of the polymerase-associated factor 1 complex (PAF1C) and NCoR co-repressor complex, the latter of which mediates gene silencing through histone deacetylation. Although the precise developmental consequences of Setd5 ablation have yet to be fully understood, this work suggests that this protein might cooperate with PAF1C and NCoR to mediate co-transcriptional regulation of histone acetylation and gene activity.
A new view on implantation
Implantation of the blastocyst into the uterus is obviously a critical step in mammalian development, yet we understand very little about the three-dimensional environment into which the embryo implants. It is known that, in mouse at least, blastocysts attach in uterine crypts, but how these form and whether such structures are also found in human is unclear. Here (p. 4749), Diana Laird and colleagues seek to provide new insights into uterine architecture before, during and after implantation. The authors develop sophisticated imaging and computational tools to characterise the 3D structure of the mouse uterine luminal and glandular epithelium, showing that the pattern of folding alters dramatically prior to implantation, giving rise to folds that overlap with structures described as crypts. Moreover, uterine glands reorient towards the site of implantation and show structural changes. This technology is able to detect architectural defects in mutant animals (such as aberrant luminal folding in Wnt5a mutants) and can also be applied to human uterus samples – as well as, potentially, other organs. This work provides an unprecedented view of the environment into which the embryo implants, and opens up avenues for further analysis of the mechanisms underlying uterine restructuring during early pregnancy.
Sparking regeneration with ROS
During regeneration, multiple signalling pathways act to coordinate the various processes required to regenerate an injured organ or body part. Both reactive oxygen species (ROS) and electric currents have been shown to modulate regeneration, but how they exert their effects, and whether their activities might intersect, is poorly understood. Here (p. 4582), Fernando Ferreira, Min Zhao and colleagues set out to address the potential interplay between ROS and bioelectric phenomena using the Xenopustadpole tail regeneration model. They uncover a dual role for NADPH oxidases in regulating bioelectric activities: NADPH oxidase-driven electron flow induces membrane depolarisation, while the hydrogen peroxide produced leads to activation of sodium channels in cell membranes of the regeneration bud, with consequent effects on transepithelial potential and electric currents that mediate regeneration. Moreover, external application of hydrogen peroxide can induce tail regeneration during the refractory period in the tadpole’s life – when regeneration is normally blocked – as well as the formation of ectopic tails at injury sites during the regenerative period. Although the mechanisms by which bioelectric activities might modulate the cellular processes required for regeneration still require further investigation, this work links two previously unconnected regulators of regeneration and provides convincing evidence for redox-bioelectric integration in this context.
Robust transcriptional control of multiciliogenesis
Multiciliated cells (MCCs) are found on various epithelia where they drive fluid flow – such as in the airways, the brain ventricles, and the skin of Xenopus embryos. Their differentiation is known to be coordinated by transcriptional regulators such as Multicilin and Gemc1, as well as by the key transcription factor Foxj1, which is also required for cilium formation in cells that produce just a single motile cilium. On p. 4654, Chris Kintner and colleagues identify another transcription factor required for proper differentiation of MCCs – Foxn4. Through an elegant combination of morpholino and CRISPR-based loss-of-function technologies, they show that loss of foxn4 disrupts docking of basal bodies to the cell surface – an essential prerequisite for cilium extension. This phenotype is reminiscent of the foxj1 phenotype, except that it largely recovers over time and that foxn4 has no apparent effect on cells with a single cilium. Through RNAseq and ChIPseq analyses, the authors find that Foxn4 promotes expression of a subset of Foxj1 targets. They propose that Foxn4, acting downstream of Multicilin, might be required to promote high-level expression of Foxj1 target genes that may be necessary for efficient generation of multiple cilia.
PLUS
An interview with Doug Melton
Doug Melton is Xander University Professor at Harvard University, co-director of the Harvard Stem Cell Institute and a Howard Hughes Medical Institute Investigator. His lab investigates the development of the pancreas, and uses insights from this process to direct the production of insulin-producing beta cells from stem cells. We met Doug at the 2016 Society for Developmental Biology-International Society of Differentiation (SDB-ISD) joint meeting in Boston, USA, where he gave the Jean Brachet Lecture. See the Spotlight article.
Fox transcription factors: from development to disease
Forkhead box (Fox) transcription factors regulate diverse biological processes both during development and throughout adult life. Mutations in many Fox genes are associated with human disease and, as such, various animal models have been generated to study the function of these transcription factors in mechanistic detail. In their Primer, Maria Golson andKlaus Kaestner review these studies and provide an overview of the Fox family, highlighting several key Fox transcription factor families that are important for mammalian development.
The many faces of hematopoietic stem cell heterogeneity
Not all hematopoietic stem cells (HSCs) are alike: they differ in their physical characteristics, they respond to different extrinsic signals, and they have different lineage outputs following transplantation. This raises questions as to why HSC subtypes exist, how they are generated, and whether HSC heterogeneity affects leukemogenesis or treatment options. In their Review, Mihaela Crisan andElaine Dzierzak provide a developmental overview of HSC subtypes during embryonic, fetal and adult stages of hematopoiesis and discusses the possible origins and consequences of HSC heterogeneity.
Department/Location: Wellcome Trust – Medical Research Council Cambridge Stem Cell Institute, University of Cambridge
Salary: £29,301-£38,183
Reference: PS10876
Closing date: 08 January 2017
Fixed-term: The funds for this post are available until 30 June 2022 in the first instance.
The Cambridge Stem Cell Institute is a world-leading centre of excellence in stem cell biology and regenerative medicine, supported by the Wellcome Trust and the Medical Research Council (www.stemcells.cam.ac.uk). The Institute comprises over 300 scientists whose research spans fundamental science through to clinical applications. Our vision is to develop a deep understanding of stem cell biology for the prevention and treatment of human disease.
Public Engagement (PE) is an essential part of our work. We seek to provide opportunities for the public to explore and question research developments and for researchers to improve their understanding of public views. The purpose of the PE Manager role is to create and support a community of scientists who recognise the importance of dialogue with the public and who have the skills and opportunities to undertake PE activities. Overall, the CSCI aims to have highly visible public engagement that is woven into all aspects of scientific research and the PE Manager is envisioned to develop this ethos across the institute.
The PE Manager is responsible for the implementation and development of the PE Programme, which includes a wide variety of public events for different target communities, as well as digital engagement activities and professional development training for researchers. The post holder will also be responsible for developing a reward and recognition framework for researcher-led public engagement that aligns with institutional public engagement priorities. The role involves both long-term, strategic planning and detailed event management (see further particulars). The PE Manager will manage one part-time (50%) Events Administrator and there may be opportunities to expand the team from 2018 onwards.
We are seeking an innovative and self-motivated public engagement professional to lead the programme through an exciting phase of growth. As we plan for our move to custom-built premises on the Cambridge Biomedical Campus in 2018, you will need to take the lead on multiple large-scale, innovative projects in order to cement PE in the working culture of our institute.
You will be educated to degree level (or equivalent), ideally in a scientific subject. You will have had experience in a similar role, preferably having managed a small team. You must demonstrate a proven track record in relationship building, event organisation, report writing, and data management. You will have outstanding organisational and administrative experience and be comfortable working to tight deadlines with minimal supervision. You should have demonstrable experience in web-based/social media communication and you should have excellent written and verbal communication and negotiation skills.
The post will work in close collaboration with senior roles in the Admin team, with supervision from the PE steering committee and SCI Administrator and will report to the Institute Director.
To apply online for this vacancy and to view further information about the role, please visit: http://www.jobs.cam.ac.uk/job/12282. This will take you to the role on the University’s Job Opportunities pages. There you will need to click on the ‘Apply online’ button and register an account with the University’s Web Recruitment System (if you have not already) and log in before completing the online application form.
Please upload your current CV and cover letter with your application by Sunday 08 January 2017.
Interviews will be held on the morning of Monday 23 January 2017.
Informal enquiries are also welcome via e-mail to David Kent at dgk23@cam.ac.uk.
Please quote reference PS10876 on your application and in any correspondence about this vacancy.
The University values diversity and is committed to equality of opportunity.
The University has a responsibility to ensure that all employees are eligible to live and work in the UK.
Craniofacial Biology – University of Southern California Health Science Campus
We are seeking a promising postdoctoral research associate, with expertise in molecular biology and bioinformatics. The program’s goal is to create the next generation of cutting-edge dental and oral health researchers in the U.S. and to shape independent scientists who are able to initiate research programs that will ultimately improve world health. This appointment provides a broad, interdisciplinary experience preparing postdoctoral researchers to generate new discoveries that identify, prevent, treat and cure diseases of the craniofacial complex. The ideal candidate seeks advanced training in all aspects of molecular and oral biology, bioinformatics and oral pathology.
Postdoctoral associates will thrive in an integrated curriculum that includes mentoring, scientific advancement, career development, publication and grantsmanship. The curriculum is taught through symposiums, seminars, clinical research centers and collaborate research. Mentors are committed to helping students transition from scholar to independent investigator in an academic or industry environment.
Position Accountabilities:
serves as a research associate for the purpose of enhancing and developing research competencies. Participates in planning, designing and conducting highly technical and complex research projects under the direction of a supervising mentor/ personal investigator (PI). May or may not work independently.
Identifies, researches, compiles and evaluates data sources, background information and/or technology related to area of specialization.
Analyzes and evaluates research data utilizing computers and provides interpretations requiring significant knowledge of a specialized area of research. Searches literature, utilizing all available resources including electronic, regarding new methodology and designs experiments accordingly.
Contributes to the development of research documentation for publication and/or prepares technical reports, papers and/or records.
Operates and maintains sophisticated laboratory/scientific equipment.
Minimum Education: Ph.D. or equivalent doctorate within previous five years
Minimum Experience: 0-1 year
Minimum Field of Expertise:Education in Molecular Biology, Bioinformatics and/or Biostatistics research with advanced knowledge of equipment, procedures and analysis methods. US citizenship required.
Preferred Education: Ph.D. in Molecular Biology with experience in Bioinformatics
Preferred Experience: Directly Related Research. Publications in peer-reviewed journals in the same or related field.
Skills: Analysis
Assessment/evaluation
Communication — Written and Oral Skills
Conceptualization and Design
Organization
Planning
Problems identification and Resolution
Project Management
Research
Statistical analysis
Special Instructions to Applicants: A copy of the doctoral diploma or other certification that indicated that the terminal degree has been completed satisfactory is required. If the doctoral candidate has not yet obtained a degree, he/she should provide evidence that a thesis has been approved together with a documented indication of the expected date of formal graduation. It is the responsibility of the faculty mentor to verify documentation. The documentation is to be filed with the Office of Postdoctoral Affairs.
For immediate consideration, please email CV to: Janice Bea (jbea@usc.edu)
November turned out to be a bumper month on the Node with posts on research (current and historical), meetings and new resources, as well as interviews and a meeting report. Plus some beautiful science-inspired art. Here are some of our highlights, as well as our pick of the best of the web this month.
Happy Thanksgiving! Yes, we CT scanned our turkey, and like any good dinosaur biologist, prepared and accessioned the skeleton (OUVC 10789). pic.twitter.com/V2YbF6mF22
The first issue of @Nature was printed today in 1869. In opening article Huxley writes on obsolescence of theories but permanence of poetry. pic.twitter.com/rNXy5mbyAD
Staying underwater, Uwe Irion and colleagueslink heterotytpic, gap junction-mediated cell interactions with cell morphology during zebrafish skin patterning.
The JCS team featured Celeste Nelson as a Cell Scientist to Watch, whose lab is “focused on studying how groups of cells physically position or turn themselves into tissues.”
Nicholas Pilon and colleagues describe how a mouse line found in a screen for genes involved in neural crest development provides a model for Waardenburg syndrome type 4.
Colin Bingle and colleagues develop an in vitro model of the murine middle ear epithelium, recapitulating cell populations and protein production.
Our latest monthly trawl for developmental biology (and other cool) preprints. See June’s introductory post for background, and let us know if we missed anything
This month, we found preprints covering various aspects of plant growth and patterning, a lot of cell biology – including insights into microtubules organisation, RNA localisation and yeast size control – as well as a bunch of tools. One of the most talked about preprints of the month comes from our ‘Away from the bench‘ section: a guide for how to structure scientific papers. Happy reading!
MultiCellDS: a community-developed standard for curating microenvironment-dependent multicellular data. Samuel H. Friedman, Alexander R.A. Anderson, David M. Bortz, Alexander G. Fletcher, Hermann B. Frieboes, Ahmadreza Ghaffarizadeh, David Robert Grimes, Andrea Hawkins-Daarud, VStefan Hoehme, Edwin F. Juarez, Carl Kesselman, Roeland Merks, Shannon M. Mumenthaler, Paul K. Newton, Kerri-Ann Norton, Rishi Rawat, Russell C. Rockne, Daniel Ruderman, Jacob Scott, Suzanne S. Sindi, Jessica L. Sparks, Kristin Swanson, David B. Agus, Paul Macklin
The Monarch Initiative: Insights across species reveal human disease mechanisms. Christopher Mungall,Julie McMurry, Sebastian Koehler, James Balhoff, Charles Borromeo, Matthew Brush, Seth Carbon, TOM CONLIN, Nathan Dunn, Mark Engelstad, Erin Foster, Jean-Philippe Gourdine, Julius Jacobsen, Daniel Keith, Bryan Laraway, Suzanna Lewis, Jeremy Nguyen Xuan, eKent Shefchek, Nicole Vasilevsky, Zhou Yuan, Nicole Washington, Harry Hochheiser,Tudor Groza, Damian Smedley, Peter Robinson, Melissa Haendel
The Wellcome Trust – Medical Research Council Cambridge Stem Cell Institute is founded on the concept that deep understanding of stem cell biology will contribute to transforming future healthcare (http://www.stemcells.cam.ac.uk). In 2018 we will move into a new purpose built building adjacent to Addenbrooke’s Hospital and multiple research institutes – http://cambridge-biomedical.com/.
The Institute has openings for Group Leaders who will complement and synergise with our existing programmes. Areas of particular interest include:
i. The interface between physical, materials or engineering sciences and stem cell biology
ii. Cell and gene therapy
iii. Ageing of stem cells
Junior group leader candidates will have a minimum of 3 years post-doctoral experience, distinctive research achievements, and an original project proposal. Senior group leader candidates will be internationally recognised for independent high quality science and have an exceptional and well-founded research proposal.
The Institute offers a collegiate environment with excellent core facilities plus extensive opportunities to pursue basic and disease focussed studies. Successful candidates will be supported to obtain external personal fellowship and grant support within 1-2 years. Interim start-up packages may be available. Depending on experience, non-Clinicians can expect remuneration between £39,324 and £66,835.
To apply online for this vacancy and to view further information about the role, please visit: http://www.jobs.cam.ac.uk/job/12123. This will take you to the role on the University’s Job Opportunities pages. There you will need to click on the ‘Apply online’ button and register an account with the University’s Web Recruitment System (if you have not already) and log in before completing the online application form.
Applicants should upload a curriculum vitae (max 3 pages, to include date of PhD and details of any career gaps if applicable) with contact details of 3 referees, and a 1-2 page outline of your research proposal, by Sunday 29th January 2017.
Interviews will be held in April 2017. Please quote reference PS10734 on your application and in any correspondence about this vacancy.
The University values diversity and is committed to equality of opportunity. The University has a responsibility to ensure that all employees are eligible to live and work in the UK. Benefits include generous maternity/ paternity leave, flexible working and funds for returning carers and other family-friendly schemes.
Since the first reported results from Yamanaka et al. in 2006, pluripotent stem cell culture has become an advantageous approach for modeling human disorders and diseases. The directed differentiation of stem cells into particular cell types can also be the basis for powerful in vitro models of early developmental defects in humans. Our lab is interested in neural tube closure as well as neural crest cell development, and to investigate our questions we use the mouse as our model system. However, we also aim to translate our findings to human development. Thus, my mentor, Dr. Lee Niswander at the University of Colorado Denver School of Medicine, and I decided to implement an in vitro model using human stem cells and test our findings from mouse in human neural crest development.
My thesis project in the Niswander Lab aims to investigate the epigenetic regulation of neural crest cell development, a migratory cell population that can differentiate into disparate cell types. My interests in both neural crest development and the epigenetic mechanisms that regulate transcription, is what sparked the idea of a collaborative visit with Dr. Ruchi Bajpai and her lab at the University of Southern California, in Los Angeles, California. Dr. Bajpai’s research interests are very similar to my own, and we identified her as a great potential collaborator as she has pioneered the directed differentiation of stem cells into neural crest, the use of fluorescently tagged enhancers in stem cells, and also defined the epigenetic signatures of neural crest enhancers. Therefore, we sought to establish a collaboration with her lab so that I could learn the directed differentiation of iPSCs into neural crest fates and subsequently test our own hypotheses of the epigenetic regulation of human neural crest development.
With the gracious assistance from the Company of Biologists, the Traveling Fellowship that I was awarded enabled me to travel from Denver, Colorado to Dr. Bajpai’s lab in the City of Angels, also known as Los Angeles. This was a tremendously rewarding experience for me. It was wonderful being able visit a new university and with the help of Dr. Bajpai and her lab, I was not only able to learn how to successfully culture and differentiate iPSCs into neural crest, but I was also able to use this method to begin testing our own hypotheses. One important step was learning how to perform chromatin immunoprecipitation (ChIP) with our differentiated neural crest cells. I also learned the technique of lentiviral infection of stem cells, which we used in combination with fluorescently tagged enhancer sequences. Back in Denver I will continue with these methods of lentiviral based modification and the directed differentiation of iPSCs into neural crest during my thesis work investigating the epigenetic regulation of neural crest development. In the future, we hope to compare datasets that we will obtain from our own ChIP-seq experiments with those generated by Dr. Bajpai with other epigenetic regulators.
Overall the experience was fantastic. It highlights the importance of collaborative science and the impacts of a strong scientific community. I have not only improved myself as a scientist, but I have also expanded my scientific network, which will benefit me for the rest of my career. I would like to thank the Company of Biologists, and also Dr. Bajpai and her lab, for the opportunity and support.
This post originally appeared on Annotations, the DMDD blog.
New image and phenotype data for embryos and placentas from embryonic lethal knockout mouse lines has been made available on the DMDD website today. The knockout data includes the ciliary gene Rpgrip1l as well as Atg16l1, a gene encoding a protein that forms part of a larger complex needed for autophagy. In total we have added HREM image data for 10 new lines, embryo phenotypes for 11 lines and placenta image and phenotype data for 6 lines.
The new data was released at the same time as enhancements to our website, which were described on the DMDD blog. Keep reading to see some highlights from the phenotype data.
DETAILED EMBRYO PHENOTYPES REVEALED
The comprehensive and detailed nature of DMDD embryo phenotyping means that we are able to identify a wide range of abnormalities. In the data released today, a total of 423 phenotypes were scored across 78 embryos. These included gross morphological defects such as exencephaly and edema, but also abormalities on a much smaller scale such as an unusually small dorsal root ganglion, absent hypoglossal nerve and narrowing of the semicircular ear canal.
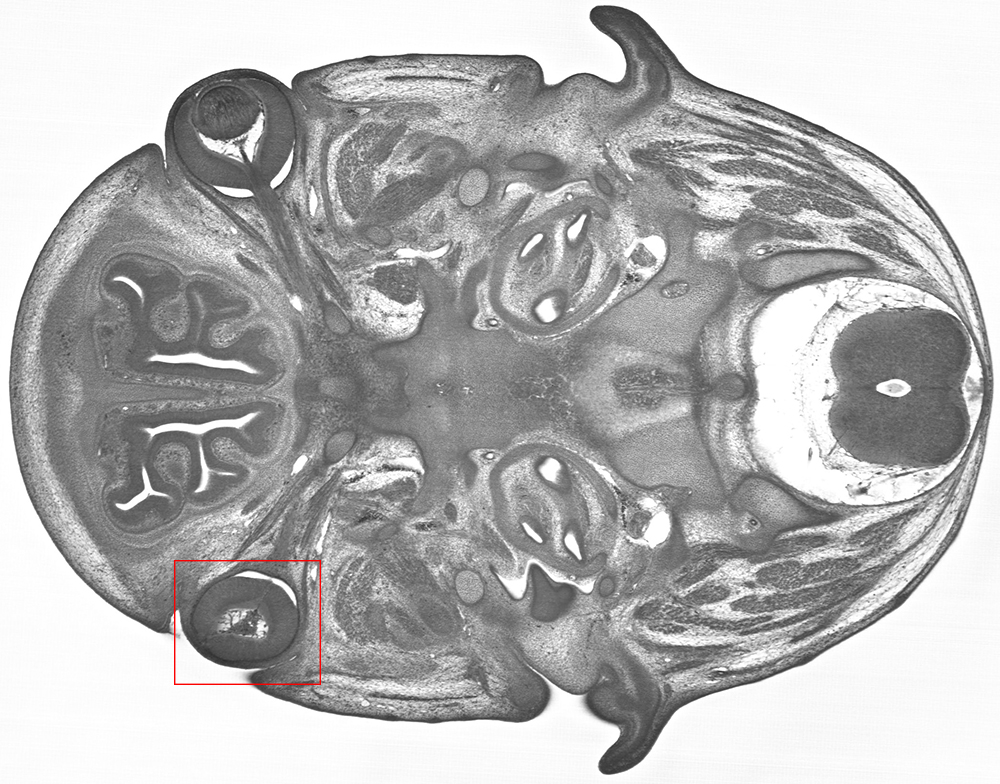
In the image below, a Trim45 embryo at E14.5, was found to have abnormal optic cup morphology and aphakia (a missing lens).
HREM imaging of a Trim45 knockout embryo reveals abnormal optic cup morphology and aphakia on the left side.
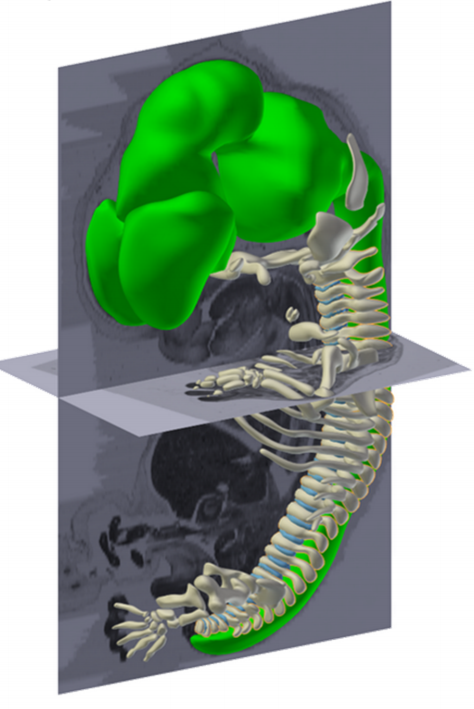
3D modelling of the exterior of an Rpgrip1l knockout embryo at E14.5 revealed a cleft upper lip, as well as polydactyly.
A 3D HREM model of an Rpgrip1l embryo shows a cleft upper lip.
All phenotypes are searchable on the DMDD website, highlighted on relevant images, and the full-resolution image data is available to explore online.
SYSTEMATIC PLACENTAL ANALYSIS
DMDD also carries out systematic phenotyping of the placentas from knockout lines. The image below shows a Cfap53 knockout placenta at E14.5, which was found to have an aberrant fibrotic lesion. The density of fetal blood vessels was also considerably reduced, the overall effect being to reduce the nutrient flow from mother to embryo.
Placental histology for the line Cfap53 shows a fibrotic lesion (large arrow) and several regions of reduced blood vessel density (small arrows).
GENE EXPRESSION PROFILES
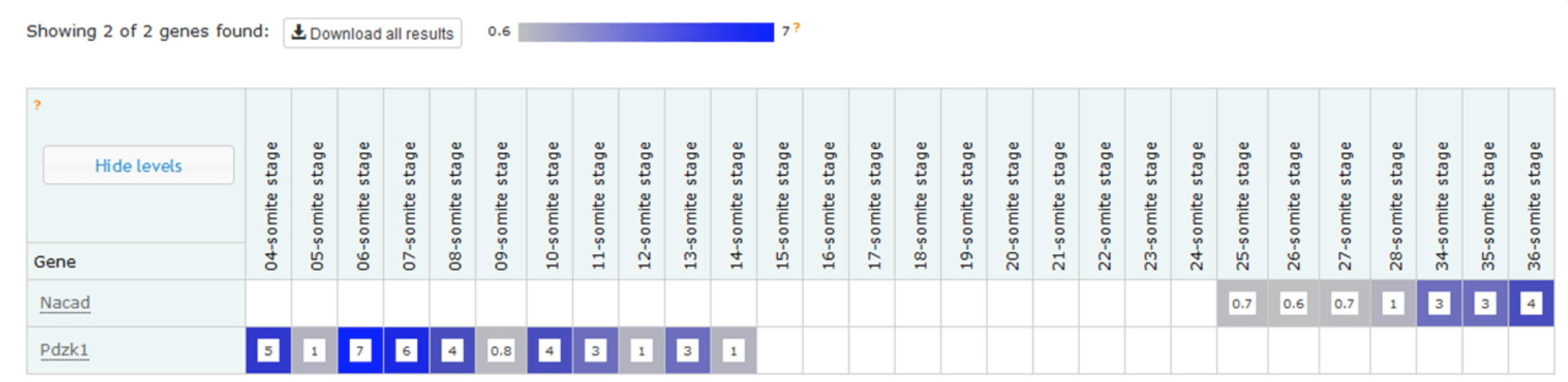
Work is underway to measure the gene expression profiles for embryos from embryonic lethal knockout lines, a study that complements the morphological phenotype data we are gathering. One of our ultimate goals is to allow data users to explore correlations between gene, morphological phenotype and gene expression profile. The first part of this dataset was released recently – a temporal baseline gene expression profile for wild type embryos.
Example expression profiles of Nacad and Pdzk1 with increasing somite number. The data shows that, at this depth of sequencing, Nacad is switched on during somitogenesis and Pdzk1 is switched off.
The expression data is now accessible via a dedicated wild type gene expression profiling page on the DMDD website, which also gives background information about the analysis. Mutant expression data will follow in the new year.
LINKS BETWEEN DMDD GENES AND HUMAN DISEASE
Many of the genes studied by the DMDD programme are known to have links to human disease, including several new lines that have been made available in this release.
Cfap53: the human ortholog of this gene is known to be associated with visceral heterotaxy-6, in which organs have an abnormal placement and/or orientation.
Embryonic lethal lines with no known links to human disease may also be novel candidate genes for undiagnosed genetic disorders. Visit the DMDD website to explore the phenotype data.
Sarah, Kristian and Joy in the lab, plus some toys
So Kristian, how did you come to establish your lab, and what are the main questions you are interested in?
KF I originally started off as a vet. However, after a short time in practice I realized that all pets have owners and that I had to spend a significant amount of time with them rather than with the animals. I quickly figured out that science is much more fun and started my PhD in neuroscience. Soon after I moved towards biophysics, and started my lab 5 years ago in the Department of Physiology, Development and Neuroscience at the University of Cambridge, after finishing my postdoc at Cambridge’s Physics Department. Our lab works at the interface of life and physical sciences; we are mainly interested in how mechanical signals control cell function in the nervous system. Particularly, we want to understand how local tissue stiffness, cellular forces, and cellular mechanosensitivity contribute to CNS development, and how changes of cells’ mechanical environment impede, for example, neuronal regeneration after spinal cord injuries.
Do you think cell mechanics are adequately appreciated in current developmental biology?
KF Definitely much more than in many other branches of biology. Forces are an old concept in developmental biology. During development, cells move all the time, and there is no motion without forces. Pretty much every developmental biologist I know appreciates cell mechanics, which makes me happy.
“During development, cells move all the time, and there is no motion without forces. Pretty much every developmental biologist I know appreciates cell mechanics”
On to the current paper: I understand it has had a long gestation?
KF That’s correct. It started some 8 years ago, when I applied for a postdoctoral fellowship with Christine Holt. That’s when I came in contact with Xenopus for the first time. Since then, we have been working on the story, had to develop a number of new experimental techniques and approaches, and to learn a lot about the mechanobiology of developing retinal ganglion cells. It went through 2 generations of highly talented postdocs and graduate students in my lab, and now I’m very happy that we finally managed to publish the paper.
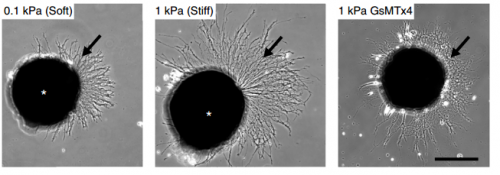
Cultures of Xenopus eye primordia on soft or stiff substrates, and with spider venom peptide added. From Figure 1, Koser, et al. 2016. Nature Neuroscience.
And Joy and Sarah: how did you come to be involved with this story? Were you working fairly separately, or was there a lot of collaboration?
AJT & SKF This story grew up over a long time! Joy started working on it before Sarah joined the lab, focusing on brain mechanics during development in vivo. Sarah started about a year later, working on the in vitro side. The experimental work was fairly independent, but there was a lot of scientific discussion behind the scenes among all the authors and other members of the lab. We have since started working on a collaborative project that uses Sarah’s expertise in in vitro cultures and Joy’s experience in atomic force microscopy.
Why choose the Xenopus retinal ganglion cell (RGC) as a model?
AJT & SKF The RGC model is great because it provides both a simple in vitro system and an elegant in vivo system. The structure of the optic tract is stereotypic and easily visualized, making it a brilliant system for seeing how different perturbations affect axon growth and guidance. To study in vivo brain mechanics, we needed an organism where the brain was both accessible in the embryo and developed at a rate that was practical for our measurement timeline. That’s quite a challenging set of requirements and Xenopus fit the bill nicely. Furthermore, and fortunately for us, the rate of Xenopus development can be sped up or slowed down by changing the temperature, which allows a lot of flexibility when planning experiments.
“The Xenopus retinal ganglion cell model model is great because it provides both a simple in vitro system and an elegant in vivo system”
KF Also, with Christine Holt we had a great collaborator who is a leading expert in Xenopus CNS development. This made the choice pretty easy.
AJT & SKF – On the downside, frogs are a less widely used model system and also have a complicated (allotetraploid) genome. They’re therefore less developed as a genetic model than other organisms like zebrafish, Drosophila, and even mice. Sometimes that can be quite frustrating—knockout frogs simply don’t exist, so other techniques, like morpholinos, drug treatments, or RNA/DNA injections must be used instead. Joy worked with Drosophila during her MPhil and undergraduate dissertation, so swapping to Xenopus embryos for the PhD felt a bit constraining at first! However, the Xenopus genome is known now, and CRISPR/Cas9 is showing great promise.
KF: In the end, the pros outweighed the cons by far.
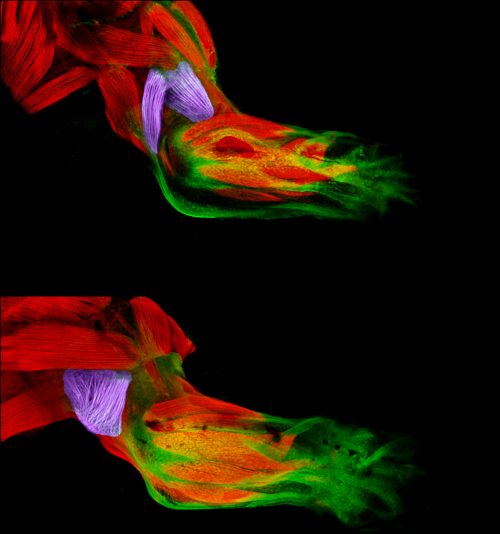
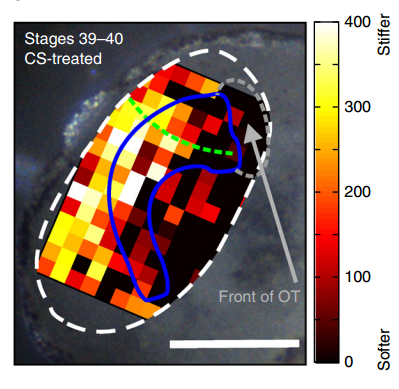
Xenopus brain overlaid with AFM-based stiffness map, from Figure 2, Koser, et al. 2016. Nature Neuroscience.
How did you go about testing the mechanics of the brain, and the responses of neurons to their mechanical environment?
AJT To measure in vivo brain mechanics, I borrowed a technique from materials science called atomic force microscopy (AFM). This is an extension of the sense of touch, just as the optical microscope is an extension of our sight. AFM uses a flexible probe (a leaf spring) to apply a set force to the brain surface; the amount by which the probe deflects (bends) is proportional to the stiffness of the tissue. It’s a bit like tapping a piece of concrete or prodding a piece of jelly. The method we used was developed in our lab over years to enable the measurement of local brain stiffness in the living embryo at cellular resolution. The AFM probe can also be used to apply a mechanical stimulus to the brain (e.g., to push on it for hours and hours) and track neuronal responses, which I did for the compression stiffening experiments.
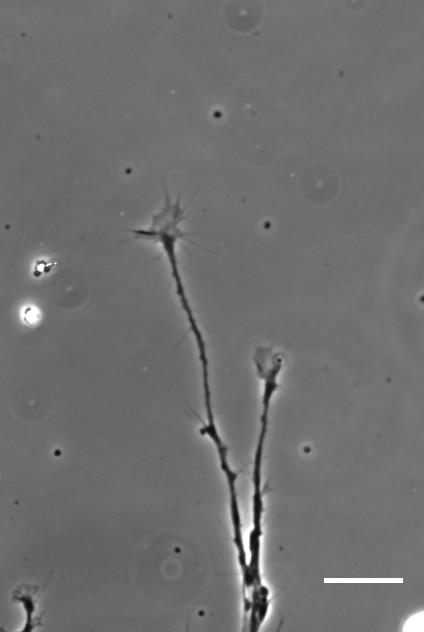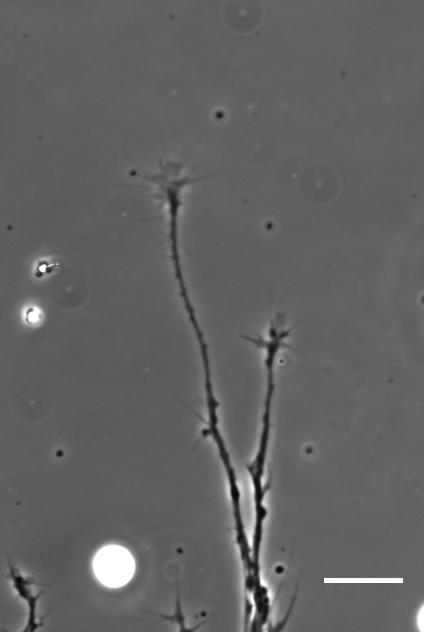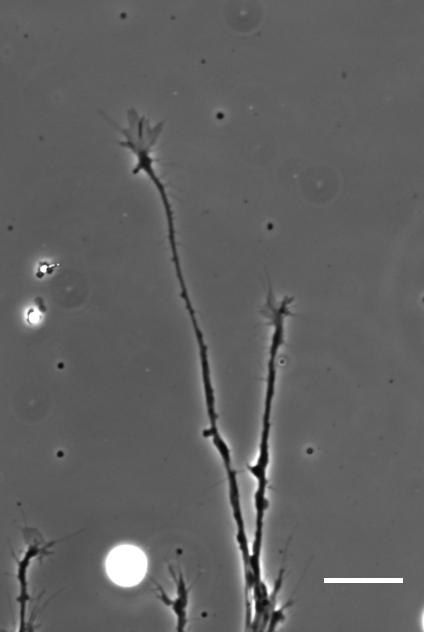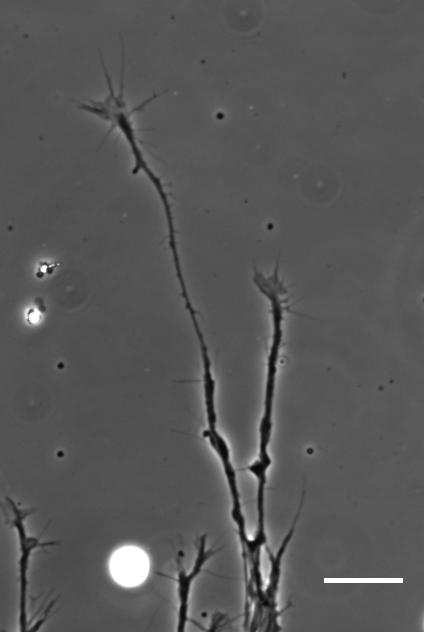
SKF For the in vitro studies, we made compliant gels out of a polymer called polyacrylamide. Varying the ratio of two of the gel’s components allowed us precisely control the stiffness of these substrates, and we selected compositions that gave us stiffnesses near the upper and lower bounds of in vivo Xenopus brain stiffness. As brain tissue is extremely soft (comparable to cream cheese), finding the right composition was a challenge. Once we grew neuronal cultures on these substrates we were able to analyse their morphology and growth dynamics as a function of substrate stiffness using simple phase contrast microscopy.
“For the in vitro studies, we made compliant gels out of a polymer called polyacrylamide…As brain tissue is extremely soft (comparable to cream cheese), finding the right composition was a challenge.”
To further explore neuronal responses to their mechanical environment, we developed substrates containing a linear gradient in stiffness. These gradient substrates allowed us to mimic the stiffness gradients that the AFM measurements revealed in the developing brain in vivo; we could then isolate this aspect of the neuron’s environment to get a clearer idea of their mechanical response in the absence of other stimuli like chemical cues.
KF To test the importance of neuronal mechanosensitivity in vivo, we perturbed both brain stiffness and neuronal mechanotransduction in the developing embryo. While mechanically perturbing the local stiffness gradient found in the mid-diencephalon redirected axon growth, both chemically softening brain tissue and preventing neurons from detecting ‘stiff’ (either by pharmacologically blocking mechanosensitive ion channels including Piezo1 or by downregulating the expression of Piezo1) led to unusual neuronal growth patterns, and the axons never arrived at their target, the optic tectum.
Could you sum up the key results of your paper in a paragraph?
KF, AJT & SKF The key result of our paper is that neuronal growth is not only controlled by chemical signals, as it is often assumed, but also by mechanical signals. We have shown that neurons grow faster and straighter on stiffer substrates, while they slow down and grow in a more random fashion on softer ones. Piezo1, a mechanosensitive ion channel, is crucially involved in transducing the mechanical signal (here: tissue stiffness) into a response of the neuron. Developing brain tissue in vivo is mechanically highly heterogeneous and exhibits very reproducible stiffness gradients, which are likely established by changes in local cell densities in the tissue. These stiffness gradients guide neurons towards the softer side, making them turn towards their target – of course in combination with chemical guidance cues. Interfering with either tissue stiffness or neuronal mechanosensitivity was sufficient to mess up neuronal growth and pathfinding.
Time-lapse movie of cultured Xenopus retinal ganglion cell axons. Supplementary Video 2 from Koser, et al. 2016. Nature Neuroscience
So how does a cell integrate mechanical information with the classical chemical cues? Do mechanical signals carry more weight?
SKF That’s an area we’re increasingly interested in now, and in fact is the topic of my Ph.D! Growing neurons encounter a massively complex environment. They are barraged with myriad chemical and mechanical cues and must integrate all this information to arrive at a coherent response. The growth cone – a sensory-motor apparatus at the tip of the growing neuron – is amazingly complex and how it performs this ‘analysis’ is not yet clear.
We think that chemical and mechanical signals likely work together in development. We require multiple senses in order to understand our surrounding environment completely, and likewise neurons need both chemical and mechanical information to inform their growth and development.
Additionally, once mechanical cues get past the membrane they are likely carried through similar or overlapping signal transduction cascades as chemical cues. For example both chemical and mechanical guidance signals likely affect Rho/Rac signalling to control cytoskeletal dynamics in the growth cone, which eventually determines neuronal growth. That chemical and mechanical signals converge on common pathways and signalling cascades indicates that these two types of signals likely interact in many elegant and subtle ways.
“Growing neurons encounter a massively complex environment. They are barraged with myriad chemical and mechanical cues and must integrate all this information to arrive at a coherent response”
KF I’m not sure if mechanical signals carry more weight than chemical ones. It’s probably more likely that tissue stiffness is used to modify responses to chemical cues and make the responses more robust. Topological cues (such as scratches in the surface of a culture substrate), on the other hand, which I’d also classify as mechanical signals, have long been known to be able to override chemical signals and guide growing neurons or migrating cells along them. I think this is a difficult question and we are only beginning to understand the role of mechanical signals in controlling development.
Unlike in the durotaxis of cell collectives in vitro, the neurons prefer to grow towards softer tissue. Is this just an in vivo/in vitro distinction, or do you expect this preference to vary depending on developmental context?
AJT We see this in vitro as well as in vivo, at least with Xenopus retinal ganglion cells. When bundles of retinal ganglion cell axons grow over a culture substrate with a stiffness gradient, they still tend to turn towards the softer region. Since RGC axons grow faster on stiffer substrates than they do on soft, we think this ‘mollotaxis’ is at least partly a collective effect. When a bundle of axons is exposed to a stiffness gradient, the ones on the stiffer side will grow faster than the ones on the softer side. However, as the axons are tightly coupled to each other along their length by cell adhesion molecules, the faster growing ones will be held back by the slower ones and bend towards them, i.e. towards soft. It’s analogous to plants growing in the sun: the cells in the stem pointing away from the sun grow faster than the cells on the brighter side and thus push the whole plant towards the light.
Whether this is a general property of developing neurons is still an open question. Certainly previous in vitro work has shown that different neuronal populations vary in their response to soft versus stiff substrates, so there may be an element of cell type specificity. Different neuronal populations may have different preferences. Growing axons can also change their response to the same chemical cue over time – so why not to mechanical ones too?
“Growing axons can also change their response to the same chemical cue over time – so why not to mechanical ones too?”
When doing the research, was there a particularly exciting result or eureka moment that has stayed with you?
AJT The compression stiffening experiments really stood out to me. Compression stiffening is a phenomenon where brain tissue increases in stiffness when a force is applied to it. I took advantage of this to increase the local tissue stiffness in different areas of the embryo brain in vivo and see if the RGC axons changed their growth behaviour. It turned out that they grew away from the stiffened region regardless of where the force was applied. It was amazing to see such a dramatic neuronal response to a stimulus that was purely mechanical (while all chemical guidance cues were very likely present as normal)! This was an exciting experiment, but at the same time the principle was refreshingly straightforward: what would this do if I poked it?
SKF Once we got the in vitro GsMTx-4 experiments working – where we inhibited mechanosensitive ion channels and then examined neuron morphology- the phenotype of treated axons lined up really precisely with that of axons grown on soft substrates. There was a beautifully clear correspondence between those two conditions—much better than we could have hoped for!
And what about the flipside: any particular moments or frustration and despair?
AJT On my first try of the compression stiffening experiment – which took a whole weekend, and an additional two days and nights to prepare the brains for imaging – I saw to my horror that the control specimens looked very, very wrong. It turned out to be a problem with one of our stock solutions, and thus easily fixed, but it was still extremely frustrating to repeat such a time-consuming experiment for such a simple reason. (Not to mention losing my Sundays off!)
SKF Working with the GsMTx-4 – the tarantula-venom derived peptide that blocks mechanosensitive ion channels – was quite frustrating. Batches seemed to vary in strength quite significantly and also degrade and lose strength over time, so you never quite knew what was going to happen when you put GsMTx-4 on your neurons. I killed quite a few sets of cultures by overdosing them, before I figured out how to handle it properly.
Finally, Joy and Sarah: what next for you two after this project?
SKF I’m still at the beginning of my PhD, so I’ll spend the next few years trying to gain some insight into how cells integrate chemical and mechanical cues.
AJT I am working on a follow-up to the in vivo experiments in the paper, looking in more detail at the changes in brain mechanics at different stages in development. The overarching goal is to finish writing up my PhD, as I only have a year or so left now!
“If tissue stiffness changes during pathological processes, what are the consequences for cells in the tissue?”
And Kristian, where will this work take you?
KF That’s an excellent question. This work has raised many more questions than it actually answered. We are now in the great position to choose from many fascinating questions and to work on what excites us most. We will certainly continue to work on the importance of mechanical signals for neuronal and CNS development, but will also look at regenerative processes. If tissue stiffness changes during pathological processes, what are the consequences for cells in the tissue? Also, the question you asked earlier about the integration of mechanical and chemical signals is key to understanding what is really going on during development and pathology. I guess we have enough open questions to keep us busy for quite a while.
David E Koser, Amelia J Thompson, Sarah K Foster, Asha Dwivedy, Eva K Pillai, Graham K Sheridan, Hanno Svoboda, Matheus Viana, Luciano da F Costa, Jochen Guck, Christine E Holt & Kristian Franze. 2016. Mechanosensing is critical for axon growth in the developing brain. Nature Neuroscience 19, 1592–1598.




 Doug Melton is Xander University Professor at Harvard University, co-director of the Harvard Stem Cell Institute and a Howard Hughes Medical Institute Investigator. His lab investigates the development of the pancreas, and uses insights from this process to direct the production of insulin-producing beta cells from stem cells. We met Doug at the 2016 Society for Developmental Biology-International Society of Differentiation (SDB-ISD) joint meeting in Boston, USA, where he gave the Jean Brachet Lecture. See the Spotlight article.
Doug Melton is Xander University Professor at Harvard University, co-director of the Harvard Stem Cell Institute and a Howard Hughes Medical Institute Investigator. His lab investigates the development of the pancreas, and uses insights from this process to direct the production of insulin-producing beta cells from stem cells. We met Doug at the 2016 Society for Developmental Biology-International Society of Differentiation (SDB-ISD) joint meeting in Boston, USA, where he gave the Jean Brachet Lecture. See the Spotlight article. Forkhead box (Fox) transcription factors regulate diverse biological processes both during development and throughout adult life. Mutations in many Fox genes are associated with human disease and, as such, various animal models have been generated to study the function of these transcription factors in mechanistic detail. In their Primer, provide an overview of the Fox family, highlighting several key Fox transcription factor families that are important for mammalian development.
Forkhead box (Fox) transcription factors regulate diverse biological processes both during development and throughout adult life. Mutations in many Fox genes are associated with human disease and, as such, various animal models have been generated to study the function of these transcription factors in mechanistic detail. In their Primer, provide an overview of the Fox family, highlighting several key Fox transcription factor families that are important for mammalian development. Not all hematopoietic stem cells (HSCs) are alike: they differ in their physical characteristics, they respond to different extrinsic signals, and they have different lineage outputs following transplantation. This raises questions as to why HSC subtypes exist, how they are generated, and whether HSC heterogeneity affects leukemogenesis or treatment options. In their Review, provide a developmental overview of HSC subtypes during embryonic, fetal and adult stages of hematopoiesis and discusses the possible origins and consequences of HSC heterogeneity.
Not all hematopoietic stem cells (HSCs) are alike: they differ in their physical characteristics, they respond to different extrinsic signals, and they have different lineage outputs following transplantation. This raises questions as to why HSC subtypes exist, how they are generated, and whether HSC heterogeneity affects leukemogenesis or treatment options. In their Review, provide a developmental overview of HSC subtypes during embryonic, fetal and adult stages of hematopoiesis and discusses the possible origins and consequences of HSC heterogeneity. (No Ratings Yet)
(No Ratings Yet)















 (2 votes)
(2 votes)