We are looking for highly motivated and talented candidates for a 4yr-PhD programme in the laboratory of Dr Toki Takahashi at the University of Manchester. This studentship is funded by the BBSRC DTP and will start in September 2017. The overall aim of the project is to understand how our head is formed by combining a laboratory experimental approach with high-throughput bioinformatics analysis. This project is in collaboration with Dr Shane Herbert and Professor Magnus Rattray.
The vertebrate craniofacial development is a remarkably dynamic and intricate process, requiring the tightly regulated growth, migration and patterning of a particular cell population, cranial neural crest cells (CNC). For this reason, craniofacial malformations are among the most common, accounting for one third of human congenital defects. Whilst the causes of most cases are unknown, several clinical studies reported the mutations within Alx homeobox genes in recessive hereditary cases (ALX-related frontonasal dysplasia; ALX-FND). Our lab reported a new function of alx1 gene in frontonasal CNC migration, and demonstrated that alx1 is critical to zebrafish craniofacial development.
In this project, we will first establish a zebrafish model of ALX-FND by generating alx1 null mutant line with the CRISPR/Cas9 system. Using the real-time image analysis of this mutant, we will examine in details how alx1 controls the CNC migration. Furthermore, we will purify the migrating CNC from the mutant by FACS for deep sequencing of transcriptomes. Bioinformatics analysis from this targeted cell population will identify the downstream targets of alx1. By integrating the high-throughput data and the live image profiling of CNC migration into computer modelling, the project will elucidate the new alx1 gene regulatory network controlling vertebrate head development.
The Laboratory of Zebrafish Developmental Genomics headed by Cecilia L. Winata was established in partnership with the Max Planck Institute for Heart and Lung Research (MPI-HLR). The focus of our research is to elucidate the gene regulatory networks and epigenetic mechanisms regulating various aspects of developmental biology as the initial step towards understanding human genetic diseases. Our research employs state of the art methodology based on Next-Generation Sequencing as well as CRISPR/Cas9 genome editing technology.
We invite application from motivated and talented students to perform the research part of their Master thesis in the study of heart pacemaker development using zebrafish as a model. The project titled “Genomics dissection of the heart pacemaker in Zebrafish” will be funded by the First-TEAM grant from the Foundation for Polish Science (FNP) starting from January 2017 for 18 months. The First Team program of the Foundation for Polish Science co-financed from the European Union under the European Regional Development Fund.
The ideal candidate should be able to communicate in English language and possess good communication skills. A good understanding of any of the following subjects is essential: molecular biology, biochemistry, and/or genetics principles. Prior experience in lab work is highly desired.
We offer stipend in the range of 1500 – 2500 PLN/month depending on the candidates’ qualifications and time commitment, and the chance to work in a team of young and friendly scientists with interests in developmental biology, regulatory genomics, epigenetics, and biochemistry. The successful candidate will have the opportunity to work with the zebrafish model animal and learn and apply state of the art microscopy techniques on in vivo imaging of the heart, as well as other methods in zebrafish developmental biology.
Please forward your complete application including a CV, a motivation letter, university scores, and the contact of at least one referee to rekrutacjazdg[at]iimcb.gov.pl with the subject line “First-TEAM” before the 30th of April 2017. Thanking all applicants for their interest, only selected candidates will be invited for an interview.
Please include in your application the following statement: “In accordance with the personal data protection act from the 29th of August 1997, I hereby agree to process and to store my personal data by the Institution for recruitment purposes”.
Sanford Burnham Prebys Medical Discovery Institute, La Jolla, California
An NIH-funded postdoctoral position is available to investigate the signaling functions of retinoic acid (RA) during mouse embryo development. Our laboratory has reviewed recent advances in this field: Cunningham, T.J. and Duester, G. Mechanisms of retinoic acid signalling and its roles in organ and limb development. Nature Reviews Molecular Cell Biology 16: 110-123 (2015).
We are searching for a Postdoctoral Associate to explore the mechanisms of RA signaling during the early stages of organogenesis and limb formation using CRISPR/Cas9 gene editing, RNA-seq transcriptome analysis, and embryo chromatin immunoprecipitation.
Candidates should have a strong background in developmental biology. Salary $47,476 with full benefits.
Interested applicants should email their CV and names of three references to:
Gregg Duester, Ph.D.
Professor
Development, Aging, and Regeneration Program
Sanford Burnham Prebys Medical Discovery Institute
So far in this series, we’ve featured fly nuclear pores,lizard tails, squid eyes and mouse digits, and heard from researchers working in Germany, the US and Canada. Today, we switch model system and geographical location once again. The work was published recently in Development, and uses timed inhibition of sonic hedgehog signalling during chick wing development to test models of digit patterning. The people behind the paper are its two authors: postdoc Joseph Pickering and PI Matthew Towers, MRC Career Development Fellow at The University of Sheffield in the UK.
Matthew Towers (L) and Joseph Pickering (R, representing avian developmental biology in fancy dress)
So Matt, can you tell us how you came to form your lab, and what questions your team is aiming to answer?
MT From my days as a PhD student working on the functions of cell cycle genes during snapdragon leaf development, I have been interested in how growth and pattern specification are integrated. As a postdoctoral researcher I was fortunate enough to continue working on this problem – but on chick limb development – under the supervision of Cheryll Tickle, who has contributed greatly to this field. I set up my own lab studying limb development at Sheffield, first with the support of the university and my sponsor Marysia Placzek, and then with an MRC Career Development Fellowship. I recently obtained a Wellcome Trust Senior Research Fellowship.
“The key question my lab is addressing is how cells integrate their intrinsic developmental programmes with extrinsic signals during limb patterning.”
The key question my lab is addressing is how cells integrate their intrinsic developmental programmes with extrinsic signals during limb patterning. This work has shed light on some of the classic, but controversial models of limb development – in particular, how pattern is specified along the antero-posterior (thumb-little finger) axis, and, in collaboration with Marian Ros’ lab in Santander, the proximo-distal (shoulder-digit) axis. I also co-supervise two PhD students with Marysia who are studying how growth and pattern specification are integrated during the early development of the chick hypothalamus.
Sheffield seems like an exciting place for life sciences and developmental biology at the moment?
MT The Biomedical Science department is an exciting place to study the life sciences. We also have strong links with a number of excellent research centres that focus on a range of subjects including stem cell biology, sensory neuroscience and membrane biology. I am part of a very dynamic group of researchers in the Bateson Centre who use a variety of model organisms, including zebrafish, chickens, fruit flies, slime moulds and mice to investigate a range of questions in developmental biology. These include trying to understand the basic patterning process, to elucidating how pathogens subvert the normal development of an individual.
And Joseph, how did you come to join Matthew’s lab?
JP I completed my PhD with Anne-Gaelle Borycki in Sheffield studying the regulation of an extracellular matrix protein, Laminin α1, by Sonic hedgehog (Shh) signalling in zebrafish. In fact, my paper on this has just been published.
Matt started at the university just as I was writing up my thesis. I found his work very interesting, but he was not yet ready to hire a postdoc! I left the university for a role as a scientific sales rep, which gave me a good grasp of the kinds of developmental biology research going on around the country. I kept up to date with his research activities through discussions in the pub, and in 2014, I decided to take up a postdoc position in Matt’s lab. I had developed an interest in Shh signalling and patterning mechanisms since my days as an undergraduate student, and the chick limb seemed like the ideal model to study this further.
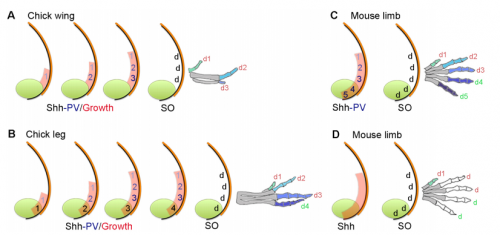
Models for digit patterning in different limbs of chick and mouse, from Figure 1 in the paper.
Your paper uses chicken wing development to test models of vertebrate digit patterning. What was the state of play before you started this work, particularly with regard to ideas of positional information and self-organisation?
MT&JP Positional information can explain how different digit identities (thumb vs. little finger) are specified in the chick wing. However, it has been apparent for a long time that digit number and spacing is determined by a self-organising mechanism that can be modelled by a Turing-like process. Before we started this work it was unclear how these two processes could act together to generate some of the diverse digit patterns found in nature. Thus positional information and self-organisation were often viewed as competing models of digit patterning.
“Positional information and self-organisation were often viewed as competing models of digit patterning.”
Could you talk us through the basic experimental rationale behind the paper?
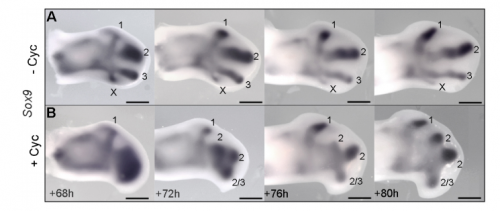
JP: I was investigating another problem using the drug cyclopamine to inhibit the Shh pathway in the chick wing bud. Unexpectedly, I noticed that at a specific stage of development, the cyclopamine-treated wings produced an extra digit. Matt had occasionally seen this phenotype when he was a Postdoc in Cheryll Tickle’s lab (MT – it gave me a headache because it did not seem to make any sense) and we talked about how similar it looked to patterns of digits produced by self-organisation in the absence of positional information. The study of self-organisation in the limb has had a renewed interest, especially in recent papers from James Sharpe and Marian Ros. Therefore, since it looked like we had transformed a chick wing pattern requiring positional information, to one not requiring it, I decided to study it further. I was able to routinely obtain the phenotype and this allowed us to characterise how the temporal interplay between positional information and self-organisation can produce diverse digit patterns.
Sox9 expression in untreated and treated wings, from Figure 4 in the paper
Could you sum up the key results of your paper in a paragraph?
MT&JP Previously, as a postdoc I had shown that Shh signalling integrates antero-posterior growth with the specification of cells with three antero-posterior positional values that give rise to the three chick wing digits (1-2-3). In our current Development paper we showed that Shh inhibition during a very precise point during this process could unexpectedly result in wings forming with four digits (1-2-2-2, based on the pattern of phalanges). We showed that this occurred because the specification of antero-posterior positional information is truncated. However, a switch to antero-posterior growth mediated by the overlying epithelium then occurs. This expansion allows for cells specified with a digit 2 positional value to give rise to up to three digit 2s by self-organisation. Interestingly, one of these digits 2s unexpectedly arises from the Shh-producing cells of the polarizing region – an ability lost in the dinosaurian ancestors of birds.
And to expand on this theme, what does this work suggest about the evolutionary history of digit patterning?
MT&JP Previously, we had presented a model for how positional information specifies the four different positional values of the chick leg digits (1-2-3-4). As the chick leg has remained unchanged during the evolution of amniotes in terms of phalangeal pattern, we suggest that it represents the patterning mechanism for four of the five digits of the common ancestor of birds and mammals. We speculate that the ancestral mechanism has been maintained in the bird wing, apart from the loss of the most-posterior digits that arise from the polarizing region (digits 4 and 5). However, in light of our new findings, we propose that the specification of positional information has been curtailed in the mammalian limb, but that, based on phalangeal pattern, self-organisation then results in the formation of several digit 2s (1-2-2-2-2).
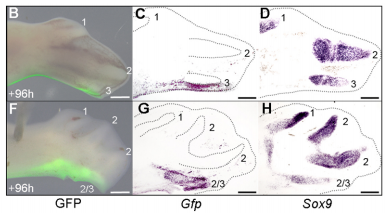
The GFP grafting experiment, from Figure 5 in the paper
When doing the research, was there a particularly exciting result or eureka moment that stayed with you?
JP Successfully grafting a polarising region from a GFP-expressing chicken into a wild-type chicken felt like quite an achievement by itself, but did not come close to the excitement of discovering that the graft contributes to the fourth digit of cyclopamine-treated wings! This is the first piece of work to clearly show that a digit can arise from the polarizing region of the chicken wing bud, and gave important insights into how mammalian digits may be patterned. We were also surprised because we had expected an additional central digit to arise in the cyclopamine-treated wings, not an extra posterior digit. I also once managed to juggle 3 eggs for at least 10 seconds.
And what about the flipside: any particular moments or frustration and despair?
JP Performing the experimental work in this paper was very repetitive and laborious due to the sheer number of embryos that would die. This was particularly frustrating when it involved difficult experiments such as tissue grafting.
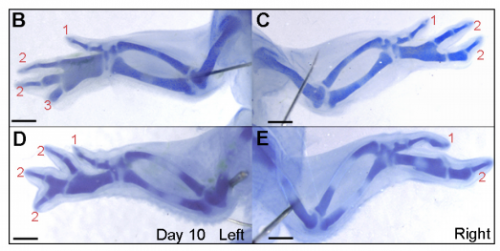
Removal of the apical ectodermal ridge after cyclopamine treatment, from Figure 7 in the paper
So what next for you, Joseph?
JP More chicks and drugs and rock ‘n’ roll! I’m currently trying to understand how the four-digit phenotype that I have characterised occurs at the molecular level. I am testing various candidate genes for their role in digit formation from the polarising region using chicken and mouse embryos, and also validating an RNA-sequencing screen that I have performed on cells of the developing chick limb.
More chicks and drugs and rock ‘n’ roll!
And what is the Towers lab currently working on?
MT Our recent research has highlighted the importance of cells measuring time to intrinsically execute their developmental programmes (Chinnaiya et al, Nat Comm 2014, Saiz-Lopez et al 2015). Now we are particularly interested in the molecular nature of intrinsic timing mechanisms, and how they could be used to scale pattern formation both within, and between different species.
29 March – 11 April 2017 Cold Spring Harbor Lab Xenopus course:
Xenopus is remarkable for modeling human diseases including birth defects, cancer, and stem cell biology. Xenopus has and continues to make a major impact in our understanding of cell and developmental biology.
Students are encouraged to target their own genes of interest using CRISPR technology and then analyze phenotypes using the diverse array of assays available in Xenopus. Specifically, techniques covered include microinjection, and various molecular manipulations including, CRISPR knockouts, morpholino based depletions, transgenics, and mRNA overexpression. In addition, students can combine these techniques with explant and transplant methods to simplify or test tissue level interactions. To visualize subcellular and intercellular activities, we will introduce a variety of imaging methods including time-lapse, fluorescent and confocal microscopy. Additional methods include mRNA in situ hybridization and protein immunohistochemistry as well as basic bioinformatic techniques for gene comparison and functional analysis. Biochemical approaches such as proteomics and mass spectrometry will also be discussed. This course runs in close association with two other courses: Quantitative Imaging and Protein Complexes.
This course is designed for those new to the Xenopus field, as well as for more advanced students who are interested in emerging technologies. We encourage students to bring their own genes of interest and will tailor aspects of the course to enable them to initiate studies on their specific projects. GENEROUS SCHOLARSHIPS AVAILABLE.
PhD studentship opportunity in the laboratory of Prof. Nicoletta Bobola at the University of Manchester, UK.
This project is to be funded under the MRC Doctoral Training Partnership.
Project description:
Cardiovascular disease is the leading cause of death worldwide. The majority of disease-associated loci identified by genome-wide association studies (GWAS) lie in non-coding regions, but without a functional delineation of the genome, it is currently impossible to understand the importance of these variations and their contribution to biological mechanisms and disease. Epigenomic profiling of chromatin features allows the identification of active, functional regions in the genome, located outside the coding sequence of genes. We will use ChIP-seq (chromatin immunoprecipitation with massively parallel DNA sequencing) on a defined developmental time course of mouse and human embryogenesis, to identify segments of the non-coding genome active in instructing formation of the cardiovascular system. We will associate the non-coding sequences identified with human genetic variation (GWAS) to uncover genetic variants associated to heart disease and study their function in vitro and in vivo (using transfection assays in cell lines and transgenic assays in zebrafish). Abnormal development of the cardiovascular system can lead to congenital heart disease, and increased risk of cardiovascular disease in adulthood. We expect that the results of this project will clarify how the human cardiovascular system develops and eventually expand diagnostic and therapeutic capacities.
The successful candidate will benefit from training in several cross-cutting skills, combining next generation sequencing (ChIP-seq, RNA-seq) and bioinformatics with traditional molecular biology and developmental biology techniques.
Candidates are expected to hold a minimum upper-second (or equivalent) undergraduate degree in a related biomedical/biological science such as Molecular Biology, Developmental Biology, Genetics or a closely related field. A Masters qualification in a similar area would be an advantage as would experience of human genetics, epigenetics and/or molecular biology techniques.
If you are interested in this project, please make direct contact with nicoletta.bobola@manchester.ac.uk
The 2016 BSDB Autumn meeting under the topic of ‘chimaeras and their use in studying developmental processes and disease models’ was organised by Jenny Nichols and Tristan Rodriguez. It took place on 28-30 August in The Pollock Halls at the University of Edinburgh and was sponsored by the Anne McLaren Memorial Trust Fund and The Company of Biologists.
The five sessions of the meeting were on (1) Lineage tracing and Potency, (2) Signalling Mechanisms, (3) Regenerative Medicine and (4) Human Development, (5) Disease Models and Gene Function, and saw a wide range of applications of chimaeras discussed in various model systems. Lectures presented a mixture of traditional uses of chimaeras, recent innovations within this historical context and a broad range of other ideas and approaches – incorporating both the weird and the wonderful! A detailed meeting report will follow, and the speaker list can be viewed here.
Embroidery by Mia Buehr
Chimaeras are made of cells from two or more different organisms of the same or different species. Since their first conception, chimaeras have been an essential tool to dissect cellular potential and are used to address a large number of questions in developmental biology using a variety of different model organisms, from plants to vertebrates. But they have also always been an inspiration for story tellers and other artists. It was therefore delightful to see artists exhibit on this topic on the conference. These artists were Mia Buehr, who showed computer-generated embroidery with developmental biology themes, such as chick embryos, early mouse embryos and chimaeras (LINK1, LINK2), and the jeweller Aurora Lombardo (LINK).
Finally, the Autum Meeting hosted the inaugural Dennis Summerbell Lecture which was given by Iwo Kucinski who carried out his work in Eugenia Piddin’s laboratory at the Gurdon Institute. Iwo’s lecture was entitled “The molecular signature of the loser cell status reveals key pathways regulating cell competition” (for more detail see here).
Overall, the meeting was a great success, paving the way towards the big BSCB/BSDB/Genetics Society Joint Spring Meeting (2-5 April 2017), for which registration is now open (LINK).
Dementia causes enormous personal hardship and costs the UK ~£23 billion every year. The second most common form is Frontotemporal lobar degeneration (FTLD). About 40% of FTLD cases have genetic causes, with >8% involving abnormal aggregate-forming GA, GR, PR, GP and AP dipeptide repeat proteins (DPRs).
This project will gain new understanding of this type of FTLD by unravelling neurodegenerative pathomechanisms of DPRs through using interdisciplinary approaches. We will focus on the hypothesis that toxicity is caused by DPR structure, comparable to amyloid plaques in Alzheimer’s disease. The project will capitalise on the complementary expertises of the three supervisors. The detailed aims and outcomes are:
(1) To generate purified DPRs and perform biochemical and biophysical analyses, in order to understand the reasons for their toxicity and identify useful therapeutic strategies which will benefit patients and their families.
(2) To generate transgenic Drosophila fly stocks to obtain primary neurons expressing the four DPRs. We will use powerful fly genetics and well established cell biological approaches to identify the neuronal death pathway (apoptosis, necrosis, autophagy), to then block cell death and carry out a detailed analysis of the DRP pathomechanisms upstream.
Normal and mutant primary Drosophila neurons in culture.
(3) There is substantial proof-of-principle for the use and translational potential of Drosophila neurons. To validate identified DPR pathomechanisms in mammalian contexts, we will carry out complementary experiments using well established DPR models in SH-SY5Y cells and inducible neuronal cell lines.
The training will therefore provide plenty of opportunities to acquire skills in a wide range of techniques within the areas of genetics, cell biology, and biochemistry, supervised by three specialists in these areas: Stuart Pickering Brown, Andreas Prokop and Andrew Doig. For more details, please contact Stuart via email: SPB@manchester.ac.uk.
At the Dept of Clinical Sciences, Umeå University, Sweden
Project description:
The selected candidate will work within the research project ”The molecular portfolio of the extraocular muscles”, led by Professor Fatima Pedrosa Domellöf at the Departments of Clinical Sciences at Umeå University, Sweden. The actual research project explores i) how the extraocular muscles adapt to strabismus surgery and ii) the importance of the unique properties of these muscles for their resistance against neuromuscular diseases. The candidate will mostly work with the development and evaluation of zebra fish models, microarrays and different types of imaging techniques such as immunofluorescence and confocal microscopy.
Qualifications:
The candidate must have a doctoral degree / have defended a PhD thesis. The candidate must have solid knowledge of muscle biology and experience of work with zebra fish and/ or microarrays. The applicant must be fluent in English both orally and in writings.
The application must contain:
A short cover letter (about one A4 page) with description of the applicant’s research experience, research interests and motivation for the application.
Curriculum Vitae including all relevant degrees with certifications, technical expertise, previous employments, publication list, as well as names and contact information of two references.
For more information, contact Professor Fatima Pedrosa Domellöf, Department of Clinical Sciences, Ophthalmology, Umeå University, e-mail: fatima.pedrosa-domellof@umu.se; 901 85 Umeå, Sweden.
The position is for two years. Start date December 2016.
Applicants have to apply using our E-recruitment system MyNetwork Pro and must be received by 2016-11-08 at latest. Ref code AN 2.2.1-1319-16
Disease Models & Mechanisms is pleased to announce the launch of a new Special Collection named Spotlight on Rat: Translational Impact. The rat is a key model for basic and preclinical studies of physiology, pharmacology, toxicology and neuroscience, underlining its importance in studies of human disease. There are many reasons for its suitability as a model system – the close evolutionary and genomic relationship to humans, the sophistication and sociability of the animal, the ease of physiological and behavioural measurements, and the recent proliferation of transgenic and knockout rats, enabled by new and improved technologies for genetic manipulation.
In an introductory Editorial, guest editors of the launch issue Tim Aitman and Aron Geurts discuss why it is a timely moment to review progress and prospects for rat-based translational research. The launch issue includes an exclusive interview with Howard Jacob, who provides his perspectives on the past, present and future or rat research, an update on the Rat Genome Database, and Reviews on the key advances made using this model in the fields of system genetics, rheumatoid arthritis and spinal cord injury. We also present an At a Glance poster article that describes important differences between rats and mice that impact on their use as model organisms for brain disorders. The original research articles published in the issue highlight the utility of the rat model across diverse areas, including neuroscience and neurobehaviour, musculoskeletal disease, oncology, metabolism, and infection and immunity.
Coming up soon are Review articles on rat models of obesity and renal disease, plus more original research.
 (3 votes)
(3 votes)
 (7 votes)
(7 votes)
 Finally, the Autum Meeting hosted the inaugural Dennis Summerbell Lecture which was given by
Finally, the Autum Meeting hosted the inaugural Dennis Summerbell Lecture which was given by  Dementia causes enormous personal hardship and costs the UK ~£23 billion every year. The second most common form is Frontotemporal lobar degeneration (FTLD). About 40% of FTLD cases have genetic causes, with >8% involving abnormal aggregate-forming GA, GR, PR, GP and AP dipeptide repeat proteins (DPRs).
Dementia causes enormous personal hardship and costs the UK ~£23 billion every year. The second most common form is Frontotemporal lobar degeneration (FTLD). About 40% of FTLD cases have genetic causes, with >8% involving abnormal aggregate-forming GA, GR, PR, GP and AP dipeptide repeat proteins (DPRs).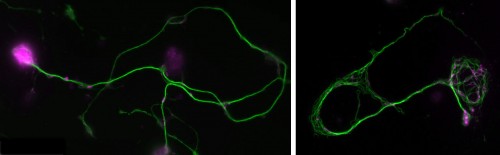
 ch of a new Special Collection named Spotlight on Rat: Translational Impact. The rat is a key model for basic and preclinical studies of physiology, pharmacology, toxicology and neuroscience, underlining its importance in studies of human disease. There are many reasons for its suitability as a model system – the close evolutionary and genomic relationship to humans, the sophistication and sociability of the animal, the ease of physiological and behavioural measurements, and the recent proliferation of transgenic and knockout rats, enabled by new and improved technologies for genetic manipulation.
ch of a new Special Collection named Spotlight on Rat: Translational Impact. The rat is a key model for basic and preclinical studies of physiology, pharmacology, toxicology and neuroscience, underlining its importance in studies of human disease. There are many reasons for its suitability as a model system – the close evolutionary and genomic relationship to humans, the sophistication and sociability of the animal, the ease of physiological and behavioural measurements, and the recent proliferation of transgenic and knockout rats, enabled by new and improved technologies for genetic manipulation.