EMBO celebrates 50 years
Posted by the Node, on 12 July 2014
![]()
The European Molecular Biology Organization (EMBO) is an organization that ‘promotes excellence in the life sciences’ in Europe and beyond. With more than 1500 members (leaders in their respective fields), it is a driving force in European science. Since its inception in 1964, EMBO has played an active role in promoting research in the life sciences, organising meetings and workshops, publishing several important journals in the molecular biology arena, supporting young researchers and influencing science policy in Europe. Many European (and non-European) scientists have benefited hugely from EMBO’s activities over the years – including several of us here at The Company of Biologists!
To mark the occasion of EMBO’s 50th anniversary, we have highlighted below some of the past Node posts that have a connection with EMBO- either highlighting the annual EMBO meetings, reporting form a workshop, or interviewing an award winner or EMBO director. For a great overview of why EMBO was first formed and its history in the last 50 years, do read this great article in Nature. Happy birthday EMBO!
EMBO on the Node:
 – Last year we interviewed EMBO’s current director, the developmental biologist Maria Leptin. In this interview she discussed her career and her vision for EMBO.
– Last year we interviewed EMBO’s current director, the developmental biologist Maria Leptin. In this interview she discussed her career and her vision for EMBO.
– Alexis Webb attended an EMBO course on lab management and considered the importance of postdocs acquiring the soft skills necessary to successfully run a lab.
– Alfonso Martinez-Arias shared his thoughts on the EMBO workshop ‘Morphogen Gradients’, and on the brave new world of quantitative developmental biology.
– Eva Amsen, the previous Node community manager, attended the EMBO meeting in 2012. She posted a day by day report (read the posts from day 1, 2, 3 &4). Eva also interviewed the winner of the EMBO Gold Medal 2012, the plant developmental biologist Jiří Friml and keynote speaker Linda Partridge.
– Natascha Bushati attended the EMBO meeting in 2011. There she had the opportunity to discuss the field with Eric Wieschaus and Marcos González-Gaitán, attend the career day and report from the talk sessions (here and here).
– In 2011 Ana Tavares was part of the committee organising the EMBO workshop on Biophysical Mechanisms of Development. They organised a science art competition entitled ‘The Physics of Life’ , and Ana’s post gave an overview of the competition (including a link to the competition’s beautiful image gallery!).
 (1 votes)
(1 votes) (No Ratings Yet)
(No Ratings Yet)






 Embryonic stem cell (ESC) cultures display a marked heterogeneity in the expression of Nanog, one of several core pluripotency factors required for proper development in vivo. In addition, Nanog levels have also been shown to fluctuate in individual ESCs in vitro; however, the extent and functional consequences of these fluctuations in different pluripotency states has not been fully established. Now, on p.
Embryonic stem cell (ESC) cultures display a marked heterogeneity in the expression of Nanog, one of several core pluripotency factors required for proper development in vivo. In addition, Nanog levels have also been shown to fluctuate in individual ESCs in vitro; however, the extent and functional consequences of these fluctuations in different pluripotency states has not been fully established. Now, on p.  The balance between excitatory versus inhibitory neuron specification during development is crucial for sensory information processing in later life. The basic helix-loop-helix (bHLH) transcription factors Ascl1 and Ptf1a are crucial for establishing this specificity in the dorsal spinal cord, but how these two factors, which recognise a similiar DNA motif, can have opposite downstream effects is unclear. Now, on p.
The balance between excitatory versus inhibitory neuron specification during development is crucial for sensory information processing in later life. The basic helix-loop-helix (bHLH) transcription factors Ascl1 and Ptf1a are crucial for establishing this specificity in the dorsal spinal cord, but how these two factors, which recognise a similiar DNA motif, can have opposite downstream effects is unclear. Now, on p.  During blastocyst development, asymmetric cell divisions generate polar and apolar daughter cells, which organise into outer and inner positions, respectively, to form the trophectoderm (TE) and inner cell mass (ICM) lineages. The Hippo signaling pathway is crucial for setting up this early lineage specification, but how Hippo signaling relates to cell position and polarity remains unclear. In this issue (p.
During blastocyst development, asymmetric cell divisions generate polar and apolar daughter cells, which organise into outer and inner positions, respectively, to form the trophectoderm (TE) and inner cell mass (ICM) lineages. The Hippo signaling pathway is crucial for setting up this early lineage specification, but how Hippo signaling relates to cell position and polarity remains unclear. In this issue (p.  Cell polarity is fundamental for biological activity across many varied cell types within different animal species. Intracellular trafficking regulates the differential distribution of proteins that is fundamental to establishing cell polarity, but how cell polarity regulators exert their effects on trafficking machinery is largely unknown. Now, on p.
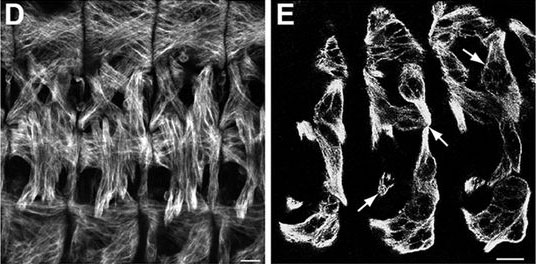
Cell polarity is fundamental for biological activity across many varied cell types within different animal species. Intracellular trafficking regulates the differential distribution of proteins that is fundamental to establishing cell polarity, but how cell polarity regulators exert their effects on trafficking machinery is largely unknown. Now, on p.  In March 2014, the RIKEN Center for Developmental Biology in Kobe, Japan, hosted a meeting entitled ‘Regeneration of Organs: Programming and Self-Organization’. Scientists from across the globe met to discuss current research on regeneration, organ morphogenesis and self-organization – and the links between these fields. As discussed by Daniel Goldman, a diverse range of experimental models and organ systems was presented at the meeting, and the speakers aptly illustrated the unique power of each. See the Meeting Review on p
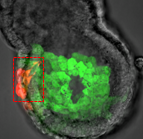
In March 2014, the RIKEN Center for Developmental Biology in Kobe, Japan, hosted a meeting entitled ‘Regeneration of Organs: Programming and Self-Organization’. Scientists from across the globe met to discuss current research on regeneration, organ morphogenesis and self-organization – and the links between these fields. As discussed by Daniel Goldman, a diverse range of experimental models and organ systems was presented at the meeting, and the speakers aptly illustrated the unique power of each. See the Meeting Review on p  Branching morphogenesis is the developmental program that builds the epithelial trees of various organs, including the airways of the lung, the collecting ducts of the kidney, and the ducts of the mammary and salivary glands. R
Branching morphogenesis is the developmental program that builds the epithelial trees of various organs, including the airways of the lung, the collecting ducts of the kidney, and the ducts of the mammary and salivary glands. R The formation of the vasculature is essential for tissue maintenance and regeneration, and understanding how vascular formation is coordinated in vivo can offer valuable insights into engineering approaches for therapeutic vascularization and angiogenesis. Here, Kyung Min Park and Sharon Gerecht discuss how the process of vascular development can be used to guide approaches to engineering vasculature. See the Review on p.
The formation of the vasculature is essential for tissue maintenance and regeneration, and understanding how vascular formation is coordinated in vivo can offer valuable insights into engineering approaches for therapeutic vascularization and angiogenesis. Here, Kyung Min Park and Sharon Gerecht discuss how the process of vascular development can be used to guide approaches to engineering vasculature. See the Review on p.