This interview first appeared in Development.
Philip Ingham is a geneticist and developmental biologist, based at the Imperial College, London – Nanyang Technological University, Lee Kong Chian School of Medicine in Singapore. From identifying the Drosophila trithorax mutant to the discovery of the vertebrate hedgehog genes, from perfecting in situ techniques in the fly to helping make zebrafish a successful model system, Phil has made significant contributions to the developmental biology field over three decades. In recognition for his career, Phil was awarded the Waddington Medal at the 2014 BSDB Spring meeting, where we had the opportunity to interview him. Phil is also the current president of the International Society of Developmental Biologists (ISDB).
 How are you finding this BSDB spring meeting?
How are you finding this BSDB spring meeting?
Very much as I remember it from the last time I came, 9 years ago. It is great that it is attended by many students and postdocs, with lots of questions after the talks. The programme is quite varied, the quality is very high and there is good international participation.
One of the reasons why you are attending this meeting is to receive the Waddington Medal. What does it mean to you to receive this prize?
The Waddington Medal is the best thing that I could imagine receiving! It is always nice to get recognition for what you do, but to receive an accolade from this society means everything to me. I have been associated with the BSDB since 1987, and I have been attending their meetings since I was a student. In fact, one of the earliest talks I ever gave was at the BSDB meeting in Sussex in 1982. So getting this award is really thrilling.
There is a rumour that before being a scientist you were a priest. This isn’t the case, but is it true that you didn’t always intend to be a geneticist?
I never intended to be a geneticist. When I went to university I don’t think I would have been able to tell you what genetics was! I had no knowledge of, and no interest in, biological sciences. I went up to Cambridge with the intention of reading social psychology, but as you couldn’t study that in your first year, I did religious studies and theology, as they were the closest related subjects. An important component of the course was looking at the effects of scientific thought on religious belief, especially Darwinism, and that is when I started thinking and reading about evolution. Then I wandered into a Biology of Cells lecture one day and saw a movie of mitosis, which I found spell-binding. That is when I decided I wanted to do biological sciences. Because of my background, the only subject I could study was genetics. You didn’t really need to know any biology to study genetics!
You were very fortunate to be invited to the exclusive EMBO Drosophila workshop in Crete when you were still a PhD student. How did this happen and what opportunities did it give you?
It was a fantastic opportunity. It happened because I was naïve enough to apply to it, as I didn’t realise what the rules and restrictions were. The fact that I had just submitted my first paper on the discovery of trithorax (Ingham and Whittle, 1980) to Molecular and General Genetics (now Molecular Genetics and Genomics) and Walter Gehring, who was also chairing the workshop selection committee, was handling it, probably had some impact. It was a chastening experience actually, as I had to give a short talk without preparation or slides. Most importantly, I got to meet several people who I still know to this day and with whom I have interacted very closely, such as Janni Nüsslein-Volhard, Eric Wieschaus and David Ish-Horowicz. David in particular was very kind and friendly towards me, and this played a part in my later decision to apply to his lab. Going to the Crete meetings is a fantastic way to get a clear overview of the cutting edge research that is going on, and all the leading people were there.
You mentioned that it was at this meeting that you first met Janni Nüsslein-Volhard, who later persuaded you to change from Drosophila to zebrafish. Why did you decide to focus on vertebrate development and was this a difficult transition?
Janni is a very powerful influence! She became interested in my work when I moved to David’s lab and started working on segmentation. In 1986 I was working at the LMB (MRC Laboratory of Molecular Biology, Cambridge, UK) and collaborating with Gary Struhl on the molecular analysis of one of Janni’s favourite mutants, even skipped. But when I visited her in Tübingen with the intention of showing her my data, she was much less excited than I expected, because she had already decided that fish was the next big thing.
She persuaded me that Drosophila was pretty much solved and that it was time to move on to a genetically tractable vertebrate model organism. I really am a geneticist, so the idea of having a new organism, starting afresh with a blank sheet of paper as I had done with flies nearly 10 years earlier, was very appealing. In addition, I knew I had the option of moving back to the newly established (but now defunct) Developmental Biology Unit of the Imperial Cancer Research Fund in Oxford, which was very vertebrate orientated, and I could see how working on fish would fit well there.
In retrospect, I think it probably wasn’t a good idea for me to start working on fish then, because it diverted my pretty limited resources away from flies. Compared with flies, fish can be incredibly frustrating: you can see the potential for doing experiments due to their beautiful embryology, but it just takes forever to do them. Although the fish community is generally very generous at exchanging material, we just don’t have the huge resources that exist in flies, and the genetics of fish is relatively primitive even now. So you can spend a lot of time creating mutants, transgenics and antibodies, and often, by the time you are ready to do the experiment, it is no longer relevant. That said, I think we are at the dawn of a new era when zebrafish will come of age as a powerful system for disease modelling and screening for novel therapeutics.
Your collaboration with Cliff Tabin and Andy McMahon was very successful, identifying the vertebrate hedgehog genes. Why do you think it was such a productive collaboration?
One simple answer is that Andy and Cliff are incredibly driven people. Whenever they set out to do something it is going to work, so being involved in a collaboration with them is almost a guarantee of success! However, once we found the hedgehog homologues and did the initial characterization, the collaboration fell apart almost as quickly as it started. Cliff has spoken at some length about this collaboration (Tabin, 2009), and in Kathy Weston’s book Blue Skies and Bench Space (Weston, 2013) he says that it is a shame that we didn’t collaborate more: a sentiment which I share. There were some tensions between our labs, but the three of us got on extremely well together. Through that collaboration they probably became two of my best scientific friends.
I believe there was some disagreement in naming one of the homologues Sonic hedgehog…
I didn’t know about the computer game, so I didn’t grab the significance: I just thought it was a silly name. People talk about silly names in Drosophila, but fly gene names are always pretty descriptive, whereas Sonic was not descriptive at all. So much so that I actually resisted even mentioning the name Sonic in our paper. In retrospect, this was probably a mistake, because our paper doesn’t get noticed as much since the name isn’t in the title. And it is one of the favourite gene names out there: Lewis Wolpert once commented ‘Phil, the one gene that my students remember is Sonic hedgehog’!
After spending most of your scientific career in the UK you moved to an A*STAR institute in Singapore. How does scientific research in Asia compare with that in Europe?
I don’t think it is as different as people imagine, but I can’t answer that question with any great authority since, although I have visited several Asian countries, I have only actually worked in one. Considering Singapore, the first thing to say is that the A*STAR institutes are very similar to Western institutes. The IMCB (Institute for Molecular and Cellular Biology) [the institute to which Phil first moved in Singapore] in particular is modelled on the LMB, so much so that when I arrived I really felt that I had gone back home! There is a certain level of topdown management of research strategy, but I don’t think it is that different from the way strategies are determined in Europe or the US – there are the same demands on politicians to justify the spending that they put into research. Maybe in Singapore there is a more extreme version of it, strategies are implemented more strictly, and things are expected to happen more quickly. For instance, the transition in A*STAR from the honeymoon period when you could do anything to having to do industrially relevant research was very sudden. It happened over a matter of months, while in Europe and the US it might have happened over a number of years. But in terms of the day-to-day practice of science, it is not so different.
You recently became the president of the ISDB. What are you aiming to achieve during your presidency?
The principal activity of the ISDB is organising the ISDB Congress every 4 years; we’ll be holding the next one in 2017 in Singapore. However, my predecessor Claudio Stern has really increased the profile and the activities of the ISDB. He has been very energetic in going around the world encouraging national communities to organise themselves into societies, and coming up with various funding schemes, such as the ISDB–Mechanisms of Development lecture. Claudio is quite a hard act to follow. I am going to try to follow his lead and continue to help communities to establish societies where they don’t exist – not least in Singapore, where we still only have a developmental biology ‘club’!
During your career you were not afraid to change lab, institute and even model organism to pursue your scientific interests. What is your advice for young scientists when making their career choices?
The best advice in science is always to do what you find really interesting. Maybe circumstances are very different now, and you do have to be a bit more directed and strategic. However, I can honestly say that I never thought about my career at all – I just did what I found interesting! Maybe I have just been incredibly lucky, but it is an approach that has paid off. I remember when I joined the ICRF (Imperial Cancer Research Fund) that I was told in no uncertain terms by other postdocs suspicious of a fly geneticist that there was no way I would ever get a position there – but then I spent 10 years of my career at the ICRF, ultimately as a Principal Scientist. I don’t think it is a good idea to try to follow trends and second guess what is going to put you in a ‘marketable’ position. Science requires so much hard work and dedication that if you are not passionate about what you are doing, you are not going to succeed.
What would people be surprised to find out about you?
I come from a strong agricultural background and am the only member of my family to have graduated from university. My grandfather was a dairyman and cattle dealer, and my happiest days as a teenager were spent working on my cousin’s arable farm outside Liverpool, picking potatoes and making hay! I also have several farming cousins in Massachusetts who I try to visit whenever I am in Boston. Also, I played guitar in a rock band when I was younger. My youngest daughter is a very good guitarist, so I still jam with her occasionally.
References
Ingham, P and Whittle, R. (1980). Trithorax: a new homoeotic mutation of Drosophila melanogaster causing transformations of abdominal and thoracic imaginal segments, Mol. Gen. Genet. 179, 607-614.
Tabin, C. (2009). Molecular Tools, Classic Questions – An Interview with Clifford Tabin. Interviewed by Richardson, Michael K. Int. J. Dev. Biol. 53, 725-731.
Weston, K. M. (2013). Blue Skies and Bench Space: Adventures in Cancer Research. Cold Spring Harbor, New York: Cold Spring Harbor Laboratory Press.
 (1 votes)
(1 votes)
 Loading...
Loading...
 (No Ratings Yet)
(No Ratings Yet) Familial dysautonomia (FD) is a germline autosomal recessive disease that is characterized by impaired peripheral sensory and sympathetic neuron development. The disease is known to be caused by mutations in the gene encoding Elp1 (also known as IKBKAP), but how Elp1 functions in neurons is unclear. Now, Warren Tourtellotte and colleagues investigate the role of Elp1 in mice (p.
Familial dysautonomia (FD) is a germline autosomal recessive disease that is characterized by impaired peripheral sensory and sympathetic neuron development. The disease is known to be caused by mutations in the gene encoding Elp1 (also known as IKBKAP), but how Elp1 functions in neurons is unclear. Now, Warren Tourtellotte and colleagues investigate the role of Elp1 in mice (p.  The recent advent of tools for manipulating and monitoring gene expression calls for efficient ways to document, access and analyse these gene expression patterns. Although a number of databases and gene expression atlases have been compiled in recent years, many of them are limited with regards to their content and utility. Here, Chris Doe and colleagues develop new software that overcomes these limitations (p.
The recent advent of tools for manipulating and monitoring gene expression calls for efficient ways to document, access and analyse these gene expression patterns. Although a number of databases and gene expression atlases have been compiled in recent years, many of them are limited with regards to their content and utility. Here, Chris Doe and colleagues develop new software that overcomes these limitations (p.  Haematopoiesis – the formation of blood cells – is regulated by a number of ubiquitous and tissue-specific transcription factors, but the extent of interplay between these factors is unclear. Sp1 is a transcription factor that is ubiquitously expressed and regulates the expression of thousands of genes, and it has been shown that Sp1-deficient mouse embryos die during early development. Now, on p.
Haematopoiesis – the formation of blood cells – is regulated by a number of ubiquitous and tissue-specific transcription factors, but the extent of interplay between these factors is unclear. Sp1 is a transcription factor that is ubiquitously expressed and regulates the expression of thousands of genes, and it has been shown that Sp1-deficient mouse embryos die during early development. Now, on p.  Root nodulation in plants is a form of de novoorganogenesis and involves the dedifferentiation of root cortical cells in response to rhizobia-derived factors. However, due to the complexity of this event, our understanding of the factors and mechanisms that initiate nodule formation is limited. Now, Takuya Suzaki and co-workers (p.
Root nodulation in plants is a form of de novoorganogenesis and involves the dedifferentiation of root cortical cells in response to rhizobia-derived factors. However, due to the complexity of this event, our understanding of the factors and mechanisms that initiate nodule formation is limited. Now, Takuya Suzaki and co-workers (p. 
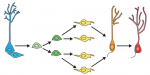 Granule neurons in the hippocampal dentate gyrus (DG) are known to be continuously generated throughout adult life, and the ongoing integration of newborn neurons into the existing hippocampal neural circuitry provides enhanced neuroplasticity, which plays a crucial role in learning and memory. In their Primer article, Gage and colleagues summarize the developmental principles that regulate the process of DG neurogenesis and discuss recent advances in harnessing these developmental cues to generate DG granule neurons from human pluripotent stem cells. See the Primer on p.
Granule neurons in the hippocampal dentate gyrus (DG) are known to be continuously generated throughout adult life, and the ongoing integration of newborn neurons into the existing hippocampal neural circuitry provides enhanced neuroplasticity, which plays a crucial role in learning and memory. In their Primer article, Gage and colleagues summarize the developmental principles that regulate the process of DG neurogenesis and discuss recent advances in harnessing these developmental cues to generate DG granule neurons from human pluripotent stem cells. See the Primer on p.  In pluripotent stem cells, the interplay between signaling cues, epigenetic regulators and transcription factors orchestrates developmental potency. Here, Maria-Elena Torres-Padilla and Ian Chambers review what is known about transcriptional heterogeneity in pluripotent stem cells, focusing on the underlying causes of heterogeneity and how transcriptional heterogeneity can be to the benefit of the whole stem cell population. See the Review on p.
In pluripotent stem cells, the interplay between signaling cues, epigenetic regulators and transcription factors orchestrates developmental potency. Here, Maria-Elena Torres-Padilla and Ian Chambers review what is known about transcriptional heterogeneity in pluripotent stem cells, focusing on the underlying causes of heterogeneity and how transcriptional heterogeneity can be to the benefit of the whole stem cell population. See the Review on p.