A postdoctoral fellowship for 3 years is available from October 2014 in Delphine DUPREZ’s team, located in the Developmental Biology laboratory in Paris (bio–dev.snv.jussieu.fr/). The team focuses on tendon and muscle development using animal models. The objective of the project is to better understand the molecular pathways involved in tendon development. The project will involve the use of the chick embryos and mesenchymal stem cells in 2-dimensional and 3-dimensional culture systems.
DUPREZ – Related references:
Guerquin et al.,(2013). J Clin Invest123, 3564-3576.
Lejard et al., (2011) J. Biol. Chem286(7), 5855-5867.
Wang et al., (2010) Dev Cell, 18, 643-654.
Applicants may be of any nationality and should have obtained the equivalent of a PhD less than 2 years. A background in developmental biology or/and an expertise in the use of mesenchymal stem cells will be an advantage.
Applications should be sent to Delphine DUPREZ to Delphine.duprez@upmc.fr
Applications should comprise the following:
– CV
– Description of previous research experiences, publication list and a personal statement describing research interests and career goals
– List of 3 names of referees including email address and telephone number
Any mammal who celebrated Mother’s Day earlier this month realizes how important mothers are for us and the tight bond between them and their children. Forget clean shirts and packed lunch every day; for us developmental biologists, there is no better reflection of this bond than the extraembryonic membranes that support the growth of the fetus in the uterus. These tissues, among other roles, serve as circulatory, digestive and excretory systems until we develop our own. As a matter of fact, they are so important that they begin to form even before the fetus itself – during the preimplantation period, before the embryo attaches to the uterus (check out the cartoon below). Therefore, the very first decisions cells need to make during a mammal’s life are whether to become part of the extraembryonic lineages (called trophectoderm and primitive endoderm at this stage) or become the foundation of the fetus (the epiblast).
Historically, the first of these decisions (whether or not to become trophectoderm) has received more attention by researchers. However, the second one, whether to become primitive endoderm or the pluripotent, embryonic epiblast, has only been studied in more detail over the last decade or so. A number of molecular markers and the cellular behaviors involved in this process have been described, although no transcription factor has yet been shown to be essential for primitive endoderm specification. In the Hadjantonakis lab, we have recently looked at the role of the main suspect – the GATA family transcription factor GATA6 – and we are publishing the results in the current issue of Developmental Cell [1].
Cartoon and timeline for the main stages of mouse preimplantation development, from the zygote (left) to the blastocyst (right). Cell lineages and their contributions later in development (boxes) are color coded: trophectoderm (green), primitive endoderm (blue), epiblast (red). Inner cell mass (purple). Implantation takes place approximately at day 4.5 of development.
This study is important for two reasons: one biological and the other one technical. As I mentioned earlier, this is the first study to show a transcription factor that is absolutely required for primitive endoderm specification. Gata6 null embryos completely lack primitive endoderm because all cells of the inner cell mass default prematurely to epiblast fate (the future fetus). Although they show no other phenotypic defect, these embryos die upon implantation, presumably because there is no tissue to support the growth of the epiblast. We also show that Gata6 heterozygous null embryos have reduced levels of GATA6 and, as a consequence, show a delay in primitive endoderm specification. Finally, this study clarifies the love triangle formed by GATA6, the pluripotency champion NANOG and the fibroblast growth factor (FGF) signaling pathway. FGF4 (or basic FGF/FGF2) is the instructive signal for primitive endoderm specification among inner cells in the blastocyst; however, as we show for the first time in this paper, GATA6 is necessary for cells to respond to FGF4, consequently downregulate NANOG, and acquire primitive endoderm identity. Our paper not only provides a mechanism for the induction of primitive endoderm fate, but also suggests that the relative levels of GATA6 to NANOG in each cell could tip the balance towards either an extraembryonic or an embryonic fate. How these levels change and how cells can measure them remains to be seen…
Proposed interaction between GATA6, NANOG and the FGF signaling pathway. Dashed lines represent hypothesized regulation, grey lines represent weak effects due to low protein concentrations. From Schrode N, Saiz N, Di Talia S, Hadjantonakis A-K (2014) GATA6 Levels Modulate Primitive Endoderm Cell Fate Choice and Timing in the Mouse Blastocyst. Dev Cell 29: 454–467
Our study is also exciting because of the approach we used to analyze the Gata6 allelic series. For those less familiar with it, the mouse preimplantation embryo is as good a model for (live) imaging and single-cell analysis as it is unfit for biochemistry. It is very small, with few cells, and can develop in vitro, without maternal support, in standard culture conditions. This means we can image and analyze the behavior of every single cell within the embryo using a rather simple experimental setup, making it an ideal model to study mammalian cell differentiation in situ. Despite these features, only recently have there been attempts at quantifying gene expression and doing single cell analyses, partly fueled by the technology that has become available [see references 2-4, or, more recently, 5-7, for examples].
In this work, we have used MINS, a piece of software recently developed in our lab [8] that can segment nuclei in series of confocal images with higher accuracy and lower manual input than any other software we have tried so far – including popular (and powerful) solutions like ImageJ or Imaris. MINS collects spatial coordinates and fluorescence intensity data for every channel on each cell, gives them a unique ID, and generates a data matrix for each embryo. After some manual curation for under/oversegmentation and to remove the occasional outlier (generally dead cells), the data is ready for you to do maths and statistics to your heart’s content. Being biologists, large data matrices send shivers down our spines, so we teamed up with physicist-turned-biologist Stefano Di Talia and, lo and behold, the numbers began to make sense! Furthermore, because MINS also collects positional information, we were able to relate changes in protein levels and cell identity to position within the embryo. To a certain extent, this kind of analysis allows us to bypass Western Blots and study protein expression in individual cells, while preserving positional information, which is lost when cells are disaggregated for gene expression analyses. This study is the first of several from our lab where we will apply this pipeline to three- and four- (time lapse) dimensional datasets to understand better the earliest cell fate decisions in mammalian development. We hope the implementation of this analysis pipeline by other labs will improve it and help generate even higher quality data in future studies. The combination of these advanced algorithms for image analysis with single-cell expression profiling techniques will let researchers study with high resolution the molecular mechanisms controlling cellular processes in situ, in intact tissues or embryos. For further proof that this is becoming a thing, have a look at these recent comments: 9, 10.
If you want to discuss or comment, do it below or you can reach me on twitter
References:
1.Schrode, N., Saiz, N., Di Talia, S., & Hadjantonakis, A. (2014). GATA6 Levels Modulate Primitive Endoderm Cell Fate Choice and Timing in the Mouse Blastocyst Developmental Cell, 29 (4), 454-467 DOI: 10.1016/j.devcel.2014.04.011
2.Kurimoto, K., Yabuta, Y., Ohinata, Y., Ono, Y., Uno, KD., Yamada, R., Ueda, H., & Saitou, M. (2006). An improved single-cell cDNA amplification method for efficient high-density oligonucleotide microarray analysis Nucleic Acids Research, 34 (5) DOI: 10.1093/nar/gkl050
3.Plusa, B., Piliszek, A., Frankenberg, S., Artus, J., & Hadjantonakis, A. (2008). Distinct sequential cell behaviours direct primitive endoderm formation in the mouse blastocyst Development, 135 (18), 3081-3091 DOI: 10.1242/dev.021519
4.Guo, G., Huss, M., Tong, G., Wang, C., Li Sun, L., Clarke, N., & Robson, P. (2010). Resolution of Cell Fate Decisions Revealed by Single-Cell Gene Expression Analysis from Zygote to Blastocyst Developmental Cell, 18 (4), 675-685 DOI: 10.1016/j.devcel.2010.02.012
5.Frum, T., Halbisen, M., Wang, C., Amiri, H., Robson, P., & Ralston, A. (2013). Oct4 Cell-Autonomously Promotes Primitive Endoderm Development in the Mouse Blastocyst Developmental Cell, 25 (6), 610-622 DOI: 10.1016/j.devcel.2013.05.004
6.Le Bin, G., Munoz-Descalzo, S., Kurowski, A., Leitch, H., Lou, X., Mansfield, W., Etienne-Dumeau, C., Grabole, N., Mulas, C., Niwa, H., Hadjantonakis, A., & Nichols, J. (2014). Oct4 is required for lineage priming in the developing inner cell mass of the mouse blastocyst Development, 141 (5), 1001-1010 DOI: 10.1242/dev.096875
7.Ohnishi, Y., Huber, W., Tsumura, A., Kang, M., Xenopoulos, P., Kurimoto, K., Oleś, A., Araúzo-Bravo, M., Saitou, M., Hadjantonakis, A., & Hiiragi, T. (2013). Cell-to-cell expression variability followed by signal reinforcement progressively segregates early mouse lineages Nature Cell Biology, 16 (1), 27-37 DOI: 10.1038/ncb2881
8.Lou, X., Kang, M., Xenopoulos, P., Muñoz-Descalzo, S., & Hadjantonakis, A. (2014). A Rapid and Efficient 2D/3D Nuclear Segmentation Method for Analysis of Early Mouse Embryo and Stem Cell Image Data Stem Cell Reports, 2 (3), 382-397 DOI: 10.1016/j.stemcr.2014.01.010
9.Wen, L., & Tang, F. (2014). Reconstructing Complex Tissues from Single-Cell Analyses Cell, 157 (4), 771-773 DOI: 10.1016/j.cell.2014.04.024
10.Robson, P. (2014). Deciphering Developmental Processes from Single-Cell Transcriptomes Developmental Cell, 29 (3), 260-261 DOI: 10.1016/j.devcel.2014.04.032
We are looking for a Research Assistant to join the Electron Microscope (EM) Facility based in the Sir William Dunn School of Pathology at the University of Oxford.
The successful candidate will assist the EM Facility Manager in two main areas. The first is the day-to-day running of the facility, which will include preparation of chemical solutions, ordering consumables and general lab maintenance. The second will be to undertake work on service projects, which will involve the preparation and imaging of biological specimens for EM, and also to train users in EM specimen preparation techniques (including ultramicrotomy) and on the electron microscopes. There will also be scope to develop the capabilities of the facility in advanced EM techniques, which will involve both independent and collaborative work with researchers.
The Dunn School EM Facility is one of two main hubs for electron microscopy of biological specimens at The University of Oxford. Here, researchers from many different departments across the University image everything from nanoparticles, proteins, viruses and bacteria to cells, tissue and whole organisms, as part of research in diverse range of disciplines, including biomedicine, biophysics, biochemistry and plant biology.
The Facility comprises an FEI Tecnai T12 Transmission Electron Microscope (TEM), a JEOL-6390 Scanning Electron Microscope (SEM) and a dedicated specimen preparation laboratory. This Facility provides its users with a broad range of EM options, from imaging negatively stained particulate samples and thin sections of resin-embedded cells and tissue at the ultrastructural level using TEM, to high resolution topgraphical imaging of cells and whole organisms (eg: C. elegans and Drosophila) with the SEM, through to more advanced techniques including 3D-SEM, EM tomography, correlative light and electron microscopy (CLEM), cryo-ultramicrotomy and protein localisation using immunogold or new genetic tags for EM. Development and optimisation of advanced EM techniques is a key priority for the Facility so that we can continue to provide our expanding user base with access to cutting edge EM methods and technologies.
The post is available as a full time fixed-term contract (Grade 6: £26,527 – £31,644 p.a.) for 2 years in the first instance. If you are interested in this role, and have the skills and experience we are looking for, please apply online. You will be required to upload a CV and supporting statement as part of your online application.
The closing date for applications is 12.00 midday on 13 June 2014. Interviews will be held in the week beginning 16 June 2014.
Decisions, decisions…aren’t those one of our main worries? It is certainly the everyday worry of a stem cell! Understanding stem cell decisions is a central question in the field: how do stem cells manage to keep the right balance between self-renewal (make identical copies of themselves) and differentiation (produce specialized cells)? How do stem cells choose between quiescence and proliferation? Good models to study these decision-making mechanisms are stem cells located in highly renewable tissues, such as the skin, the gut, the blood and the testes, since their specialized cell output is constant and huge.
It is currently known that in these tissues, some stem cells will stay quiescent in order to avoid premature exhaustion. This allows the long-term maintenance of the stem cell population and ensures that organ function is maintained throughout life. However, little is known about how these quiescent stem cells decide to stay quiescent.
In a recent study published in Development, Liao and colleagues showed that the protein Dnmt3l promotes the quiescence of spermatogonial stem cells (stem cells of the testes). They genetically engineered a mouse model in which they switched off the gene producing Dnmt3l (called Dnmt3l KO). They showed that spermatogonial stem cells in Dnmt3l deficient mice were abnormally distributed in the testes and that they proliferated more than in normal mice (wild type).
In this picture you can observe the testes of normal mice (wild type, top of the panel) and of Dnmt3l deficient mice (Dnmt3l KO, bottom of the panel). In wild type mice, the expression of PLZF, a marker for spermatogonial stem cells, is mainly perinuclear (ie: slightly around the nucleus identifiable by Hoescht 33342 in blue), a pattern specific to quiescent stem cells. In Dnmt3l KO mice, the expression of PLZF is mainly punctate (ie: small dots within the nucleus), a pattern specific to active/proliferating stem cells. From this, authors conclude that spermatogonial stem cells in Dnmt3l deficient mice proliferate more than in wild type mice, hence demonstrating a key role of Dnmt3l in regulating the quiescence/proliferation decisions of spermatogonial stem cells.
This study, among many others, is a step closer to a good understanding of how stem cells make decisions between quiescence and proliferation and/or self-renewal and differentiation. These decisions, when good, ensure the maintenance of healthy tissues throughout our lives…Bad decisions however, can lead to disease, and understanding how a stem cell makes the wrong decision is the first step to design a strategy to prevent it!
Picture credit:
Liao, H., Chen, W., Chen, Y., Kao, T., Tseng, Y., Lee, C., Chiu, Y., Lee, P., Lin, Q., Ching, Y., Hata, K., Cheng, W., Tsai, M., Sasaki, H., Ho, H., Wu, S., Huang, Y., Yen, P., & Lin, S. (2014). DNMT3L promotes quiescence in postnatal spermatogonial progenitor cells Development DOI: 10.1242/dev.105130
The research headed by Roger Gomis at IRB Barcelona, with the collaboration of Joan Massagué, describes that the loss of the suppressor RARRES3 promotes the colonization of breast cancer cells in the lung.
RARRES3 could prove to be a useful marker to identify patients with a greater risk of metastasis, as well as providing a target for the development of a specific treatment for preventive strategies after removal of the primary tumour.
A study published today in EMBO Molecular Medicine reveals that the loss of function of the gene RARRES3 in breast cancer cells promotes metastasis to the lung.
The research, headed by Roger Gomis, ICREA Professor at the Institute for Research in Biomedicine (IRB Barcelona), is the result of a collaboration between two IRB labs and Joan Massagué, at the Memorial Sloan Kettering Cancer Center in New York.
The scientists demonstrate that RARRES3 is suppressed in estrogen receptor-negative (ER-) breast cancer tumours, thus stimulating the later invasion of the cancer cells and conferring them “a greater malignant capacity,” says Gomis, head of the Growth control and cancer metastasis lab at the IRB.
This type of tumour accounts for between 20% and 30% of breast cancer cases, the metastasis of which usually occurs in the lung or other soft tissues. This stage of the disease is generally fatal. The studies have been performed in samples from mice and in cell lines and have been validated in 580 samples from primary breast cancer tumours.
The study describes that RARRES3 loss of function allows the breast tumour cell to develop the adhesion capacity of malignant cells when settling in lung tissue. Furthermore, after losing RARRES3 cells also lose differentiation capacity (specialization in a specific cell type), which facilitates the initiation of metastases in distant tissues.
“The transformation of a normal cell into an invasive tumour cell is not just about acquiring capacities but equally important is the loss of certain genes, such as RARRES3,” explains Roger Gomis.
The researchers have established that the detection of the loss of RARRES3 could provide a marker to identify patients with greater susceptibility to lung metastasis.
In addition, they uphold that, in order to enhance cell differentiation and to prevent stem cell capacity, which confers greater malignancy, some patients may benefit from treatment with retinoic acid to prevent metastasis after the removal of the original tumour.
“Treatments with retinoic acid have already been tested for other conditions. We consider that it would be interesting to develop specific treatments for this breast cancer subtype,” says Gomis.
Mónica Morales, postdoctoral researcher, and Enrique J. Arenas, “la Caixa” PhD student, both in Roger Gomis’ lab during the research, appear as first authors of the study. Xavier Salvatella, ICREA Professor and head of the Molecular Biophysics lab at IRB, contributed to revealing the key structure of RARRES3 that explains its dual role in the cell.
The study was partially supported by the BBVA Foundation, which since 2006 provides funding for metastasis research undertaken at IRB Barcelona, and by the Spanish Association Against Cancer” (acronym in Spanish AECC), the Catalan Government, and the Spanish Ministry of Economy and Competitiveness.
Reference article: RARRES3 suppresses breast cancer lung metastasis by regulating adhesion and differentiation
Mònica Morales, Enrique J. Arenas, Jelena Urosevic, Marc Guiu, Esther Fernández, Evarist Planet, R. Bryn Fenwick, Sonia Fernández-Ruiz, Xavier Salvatella, David Reverter, Arkaitz Carracedo, Joan Massagué, Roger R. Gomis. EMBO Mol Med. (2014). DOI: 10.15252/emmm.201303675
ROGER GOMIS, PhD,
This article was first published on the 27th of May 2014 in the news section of the IRB Barcelona website
Here are the highlights from the current issue of Development:
Enhanced maturation of induced neurons
The ability to reprogram human fibroblasts to neurons in vitro has opened up unprecedented opportunities in disease modelling and cellular therapeutics. Despite this breakthrough, a major challenge in the field is the limited phenotypic and functional maturation of the induced neurons (iNs). Now, on p.2216, Anna Philpott and colleagues report that cell cycle-dependent phosphorylation of key neural determinant Ascl1 affects neuronal maturation both in vivo during Xenopus development and in vitro during reprogramming of human fibroblasts to iNs. The authors show that Ascl1 can undergo phosphorylation at multiple serine-proline sites, and that this multisite phosphorylation represents a means to limit neurogenesis to appropriate levels during development. Disruption of this mechanism using a phosphomutant form of Ascl1 results in enhanced neuronal production in the early Xenopus embryo. Moreover, substitution of the phosphomutant Ascl1 for wild-type Ascl1 during lineage reprogramming not only increases the efficiency of conversion to iNs in the absence of reprogramming factor NeuroD, but also enhances the morphological and functional maturation of the cells. This exciting study suggests a way forward in addressing the roadblock of functional maturation during direct lineage reprogramming in vitro.
A-stroma-nomical role for Wnt in the ISC niche
The intestinal stem cell (ISC) niche is responsible for coordinating the ongoing maintenance and regeneration of the adult gut. Wnt signalling is crucial for stem cell maintenance in the ISC niche, but the source of Wnts remains unclear. Now, on p.2206 David Virshup and colleagues show that epithelial production of Wnt is dispensable for intestinal homeostasis in the murine gut in vivo, contrary to in vitro observations. The authors use genetic ablation studies to eliminate either Porcn or Wls, two crucial components of Wnt signalling, specifically in intestinal epithelial cells. The resulting mice from these experiments are phenotypically normal, but cannot form intestinal organoids in vitro when epithelial-derived Wnt is abolished. The authors show that the latter phenotype can be rescued by co-culturing the cells with purified intestinal stromal cells that endogenously produce both Wnt and R-spondin 3, suggesting a requirement for stromal-produced Wnt in gut homeostasis. Inhibition of Wnt signalling with the pan-Wnt inhibitor C59 results in impaired proliferation and regeneration following radiation injury in mice where epithelial Wnt is already ablated, confirming the importance of stromal-derived Wnts.
Bone repair: a fishy business
Complete bone regeneration following appendage amputation is remarkably efficient in zebrafish, but does not occur in mammals. Both groups, however, can repair bone fractures to varying extents, but whether a conserved cellular mechanism underpins both bone regeneration and repair in the adult zebrafish remains unclear. It is also unclear whether new tissue can be generated in other bony structures apart from the fin. In this issue (p.2225). Gilbert Weidinger and colleagues develop two new models of zebrafish bone repair in fin and in skull, and use these to investigate the mechanism by which bone is repaired. Their findings reveal that, like complete bone regeneration, bone repair in both models involves dedifferentiation of mature osteoblasts followed by proliferation, migration to the injury site and redifferentiation to form new bone. Using live fluorescent reporter-based imaging the authors are able to observe osteoblast differentiation dynamics, and confirm via lineage tracing that mature osteoblasts do indeed contribute to the formation of new bone following injuries in both the fin and the skull.
New start for embryonic nucleoli
Almost every living cell contains a nucleus and, within that, a nucleolus. It is commonly accepted that the nucleolus of somatic cells arises from the nucleolar precursor body (NPB), a large, compact nucleolus present in the oocyte, but whether this is true remains to be definitively shown. Now, on p.2255 Hirohisa Kyogoku and colleagues reveal that the NPB in the zygote, which is inherited from the oocyte, is not required for embryonic and full-term development in the mouse. Instead, the nucleolus in the developing embryo is formed de novo. Using micromanipulation techniques, the authors show that removal of the NPB from the late-stage zygote did not cause any disruption to normal development: the resulting zygote was able to develop to the blastocyst stage and upon implantation gave rise to phenotypically normal pups. Several cell divisions after NPB removal, the authors observed de novo formation of nucleoli from newly synthesized materials. Taken together, these results indicate that the NPB of late-stage zygotes is dispensable for subsequent embryonic development, and that the nucleoli of developing and somatic cells form from de novo synthesized material.
Plus…
Switching on sex
Christian Larney, Timothy Bailey and Peter Koopman review the transcriptional regulation of the testis-determining gene Sry. The authors present an integrated model for Sry regulation, and explain how this model functions to control sex determination in mammals. See the Review article on p.2195
Transcription factor heterogeneity in stem cells
Maria-Elena Torres-Padilla and Ian Chambers review what is known about transcriptional heterogeneity in pluripotent stem cells, focusing on the underlying causes of heterogeneity and how transcriptional heterogeneity can be to the benefit of the whole stem cell population. See the Review article on p.2173
Imaging beetle embryogenesis
This issue’s featured video shows the non-invasive, long-term imaging of a Tribolium embryo from several angles. Read the paper by Strobl and Stelzer to find out how a combination of light-sheet fluorescence microscopy and a new mounting method allowed such imaging, on p.2331
We are currently seeking applications for the role of Associate Reviews Editor for the stem cell field at Development – as a full time maternity cover position.
Joining an experienced, expanding and successful team, this is an exciting opportunity to make a significant contribution to one of the major journals in the field of developmental biology. Development publishes primary research articles, reviews and other front section content. Over recent years, the journal has expanded its focus in the stem cell field via the publication of high quality research and review articles, and the role of Associate Reviews Editor is an important part of our strategy in this growing area.
Applicants will hold a PhD, ideally in stem cell biology, although outstanding candidates with experience in other areas of developmental biology will be considered. Post-doctoral and/or previous editorial experience are desirable, although we will provide on-the-job training. The successful candidate will have a broad interest in science, the scientific community and publishing. Excellent interpersonal and literary skills, enthusiasm and commitment are also essential requirements for the position.
Core responsibilities:
• Commissioning, handling peer review and developmental editing of material for the front section of the journal
• Writing press releases, article highlights and material for Development’s community website ‘the Node’
• Representation of the journal at international conferences and within the wider scientific community
• Creative involvement in the journal’s development
The Associate Reviews Editor will work alongside an experienced in-house team, including the Executive Editor and current Reviews Editor, as well as with our international team of academic editors.
This maternity cover position provides an excellent opportunity to gain experience on a highly successful life-science journal, and offers an attractive salary and benefits. The position will be based in our office in Cambridge, and is a temporary role envisaged to last up to 12 months.
The Company of Biologists (www.biologists.com) is a not-for-profit organisation, publishing five journals in the biological sciences: the three established journals Development, Journal of Cell Science and The Journal of Experimental Biology, as well as two newer Open Access journals, Disease Models & Mechanisms and Biology Open. The organisation has an active programme of charitable giving for the further advancement of biological research, including travelling fellowships for junior scientists and contributions to academic societies and conferences.
Applicants should send a CV along with a covering letter that summarises their relevant experience and why they are enthusiastic about this opportunity, and states their salary expectations.
Applications should be sent by email no later than 23 June 2014 to miriam.ganczakowski@biologists.com
Informal queries to Miriam Ganczakowski on +44 (0)1223 426 164
Explaining new scientific concepts can be a daunting task for anyone involved in outreach. We are constantly trying to come up with ways to explain, show and describe theories and ideas step by step. I’ve recently stumbled across a new app (albeit only available on ipad) called Adobe Voice that could help out scientific communication.
Describing scientific research is like telling a story and Adobe Voice is an app made for just that. It combines on screen images of your choice with your own voice over in an elegant yet simple manner to guide the viewer through a thought process. It has many pre-set themes and background songs to choose from to help personalise your video, and I’m yet to find a combination that doesn’t look professional!
In developmental biology, visualisation of our concepts and ideas helps explain them, as description alone doesn’t convey the beauty of what we study everyday. These days, videos of the first hours of an embryo’s life are almost always included in any speaker’s presentation on development so why shouldn’t we extend this visualisation to our outreach? If, like me, you are not always able to visit as many schools as you’d like, creating these videos with adobe voice means you can send a part of yourself into classrooms virtually!
With Adobe Voice I have started what will hopefully be a mini-series resource that schools can pick up on. I’m focusing on the first ideas of genetics for a Key Stage 4 class (starting their GCSE curriculum). My first video took maybe an hour from concept to completion – I spent most of that learning how to draw a monkey – and I intend to make many more already.
With so many beautiful images available from developmental biology research, the field is made for showing off and these videos are a simple way of doing it justice. I’ve included my first video above and a link to my blog below where I’ll be putting up further videos over the coming weeks. If you have any suggestions for other videos I could make or other topics to cover please let me know, all feedback is welcome for this new endeavour into the video world!
The second video in this mini series I’m creating is now up! I’ve played around with the sound settings a bit and used only images included in the app itself to see how far I could stretch its applications to teaching scientific ideas.
This post is part of a series on science outreach. You can read the introduction to the series here and read other posts in this series here.
FULLY FUNDED PhD POSITION Regulation of immune cell migration during development in Drosophila
Seeking highly motivated, intelligent and creative student with lab experience and either a bachelor’s or a master’s degree to join a research group studying cell migration at IST Austria for their PhD.
The developmental cell biology team of Professor Dr. Daria Siekhaus is an ambitious new group at IST Austria working in the field of invasive migration of immune cells using Drosophila melanogaster. We genetically identify mutants that affect cellular barrier penetration in the embryo, and characterize them molecularly, cellularly and by live imaging. To complement our international team (working language is English), we seek applications for a graduate student to work on a project involving a mutant we have identified in a lipid metabolism enzyme that has effects on specific aspects of migration, apparently in penetrating a tissue barrier. We seek to understand how this protein functions during migration. The project would involve further genetic characterization of this mutant, live imaging of its migratory behaviors, and genetic and biochemical investigations of its function. This mutant may have a relationship to a known human disease gene and exploring that would also be part of the project as well as an investigation of its roles in vertebrate immune cell function in collaboration with the Sixt group.
IST Austria is a new dynamic interdisciplinary institute that seeks to bring the American graduate student model to Europe.
Must interview by early July 2014 and start in mid September 2014. Would present and defend a thesis proposal in a qualifying exam by the following December, and graduate within three to four years from that point.
For more information about IST Austria and the Siekhaus group please visit:
This funded graduate student position is available to any nationality. To apply and to ask any questions send an email to Dr. Daria Siekhaus (Daria.siekhaus@ist.ac.at). Include in application:
-CV and official transcript of Bachelors and Master’s (if applicable) grades
-Personal statement (up to 2 pages) about your scientific experience, interests and career goals
– Names and contact information of three people who can write a letter of recommendation
Every year we give the Node readers the chance to choose their favourite images from a selection of great pictures taken by the students of the Woods Hole Embryology course. Last year you chose 4 beautiful images that featured in the cover of Development, and now is time to make your choice again!
Below are 4 great images from last year’s course. Vote in the poll at the end of this post to choose the image that you would like to see in the cover of Development! You can see a bigger version by clicking on the images.
Voting will close noon GMT on June 9th.
1. Squid (Loligo pealeii) embryo stained with anti-acetylated tubulin (red), anti-serotonin (green), and DAPI (blue,nuclei). Imaged on a Zeiss LSM 700 confocal. This image was taken by Nathan Kenny (University of Oxford), Kathryn McClelland (Institute for Molecular Bioscience, University of Queensland), and Sophie Miller (University of Cambridge).
2. Annelid (Capitella teleta) larvae (stage 8), stained with Phalloidin (cyan, F-actin), anti-acetylated-tubulin (yellow), anti-FMRFamide (red) and Hoechst (purple, nuclei). Imaged on a Leica SP8 confocal. This image was taken by Poulomi Ray (Clemson University).
3. Live late stage embryo of a squid (Loligo pealeii). Imaged with a Zeiss SV8 and a SPOT Flex camera. This image was taken by Brijesh Kumar (Indian Institute of Technology, Kanpur).
4. Eye of a late stage squid (Loligo pealeii) embryo stained with Phalloidin (green, F-actin), anti-acetylated tubulin (red), and DAPI (blue, nuclei). Imaged on a Zeiss LSM 780 confocal. This image was taken by Ezgi Kunttas (Carnegie Mellon University).
 (No Ratings Yet)
(No Ratings Yet)

 (5 votes)
(5 votes) (1 votes)
(1 votes)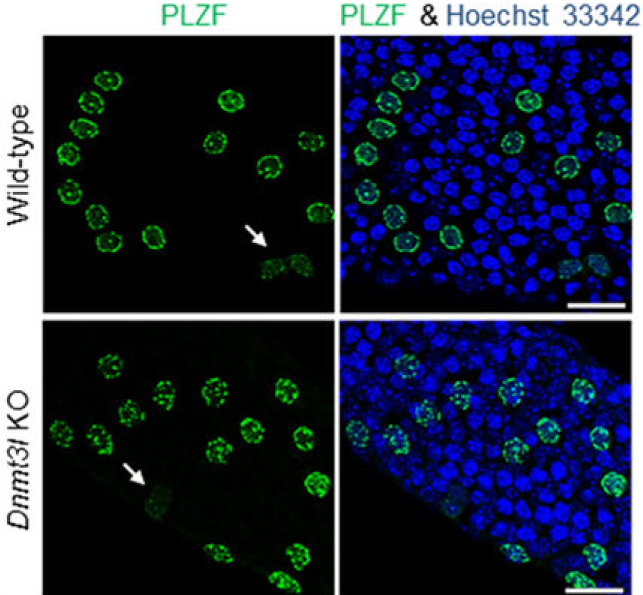


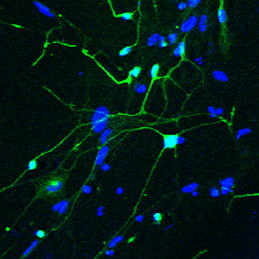 The ability to reprogram human fibroblasts to neurons in vitro has opened up unprecedented opportunities in disease modelling and cellular therapeutics. Despite this breakthrough, a major challenge in the field is the limited phenotypic and functional maturation of the induced neurons (iNs). Now, on p.
The ability to reprogram human fibroblasts to neurons in vitro has opened up unprecedented opportunities in disease modelling and cellular therapeutics. Despite this breakthrough, a major challenge in the field is the limited phenotypic and functional maturation of the induced neurons (iNs). Now, on p.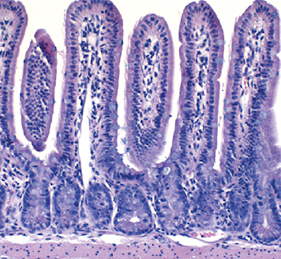 The intestinal stem cell (ISC) niche is responsible for coordinating the ongoing maintenance and regeneration of the adult gut. Wnt signalling is crucial for stem cell maintenance in the ISC niche, but the source of Wnts remains unclear. Now, on p.
The intestinal stem cell (ISC) niche is responsible for coordinating the ongoing maintenance and regeneration of the adult gut. Wnt signalling is crucial for stem cell maintenance in the ISC niche, but the source of Wnts remains unclear. Now, on p.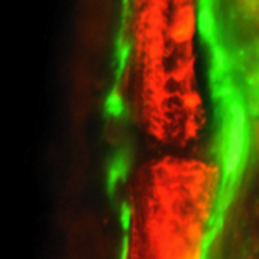 Complete bone regeneration following appendage amputation is remarkably efficient in zebrafish, but does not occur in mammals. Both groups, however, can repair bone fractures to varying extents, but whether a conserved cellular mechanism underpins both bone regeneration and repair in the adult zebrafish remains unclear. It is also unclear whether new tissue can be generated in other bony structures apart from the fin. In this issue (p.
Complete bone regeneration following appendage amputation is remarkably efficient in zebrafish, but does not occur in mammals. Both groups, however, can repair bone fractures to varying extents, but whether a conserved cellular mechanism underpins both bone regeneration and repair in the adult zebrafish remains unclear. It is also unclear whether new tissue can be generated in other bony structures apart from the fin. In this issue (p.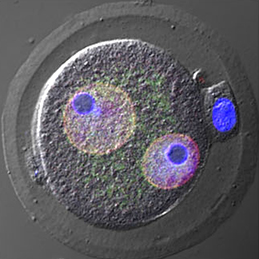 Almost every living cell contains a nucleus and, within that, a nucleolus. It is commonly accepted that the nucleolus of somatic cells arises from the nucleolar precursor body (NPB), a large, compact nucleolus present in the oocyte, but whether this is true remains to be definitively shown. Now, on p.
Almost every living cell contains a nucleus and, within that, a nucleolus. It is commonly accepted that the nucleolus of somatic cells arises from the nucleolar precursor body (NPB), a large, compact nucleolus present in the oocyte, but whether this is true remains to be definitively shown. Now, on p.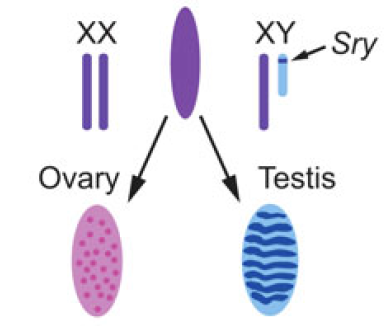 Christian Larney, Timothy Bailey and Peter Koopman review the transcriptional regulation of the testis-determining gene Sry. The authors present an integrated model for Sry regulation, and explain how this model functions to control sex determination in mammals. See the Review article on p.
Christian Larney, Timothy Bailey and Peter Koopman review the transcriptional regulation of the testis-determining gene Sry. The authors present an integrated model for Sry regulation, and explain how this model functions to control sex determination in mammals. See the Review article on p.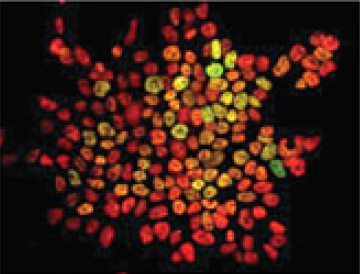 Maria-Elena Torres-Padilla and Ian Chambers review what is known about transcriptional heterogeneity in pluripotent stem cells, focusing on the underlying causes of heterogeneity and how transcriptional heterogeneity can be to the benefit of the whole stem cell population. See the Review article on p.
Maria-Elena Torres-Padilla and Ian Chambers review what is known about transcriptional heterogeneity in pluripotent stem cells, focusing on the underlying causes of heterogeneity and how transcriptional heterogeneity can be to the benefit of the whole stem cell population. See the Review article on p.