It is our birthday today! It is exactly 4 years since the Node was launched, and since then we have grown in users and readers every year! Thank you all for writing, commenting, rating and reading the Node! We hope you will join us in another year of great discussions, research, meetings, competitions, etc, etc, etc…
Birthday also means cake, so it is an appropriate time to share the video below. It shows how to cut a cake in the most scientific way, according to a 1906 paper in Nature by Francis Galton!
We have a lectureship available in Cell and Developmental Biology at the University of Bath, UK. Please share with anybody who you think might be interested.
Registration is now open for the Queenstown Molecular Biology meeting, Queenstown, New Zealand including the Developmental Biology and Reproduction satellite meeting.
August 28-29th 2014, Rydges Hotel, Queenstown, New Zealand
Sessions include: Reproduction, Infertility, Fate determination, Organ development, Developmental pathways in human disease and cancer, Neurodevelopment, Stem cells, Germ cells and Pluripotency.
Student speaker and poster prizes on offer thanks to Australia and New Zealand Society for Cellular and Developmental Biology (ANZSCDB) and Genetics Otago
The heat started to increase in Japan, as the rainy season approached and with it the high levels of temperature and humidity. But this was not an obstacle for scientists from all over Japan (and also some scientists from abroad) to meet in the great and beautiful city of Nagoya, in Aichi prefecture. Here took place the 47th Meeting of the Japanese Society of Developmental Biologists (27th-30th May 2014). The meeting was greatly organized by Masahiko Hibi-sensei, a professor in the University of Nagoya, who, as it happens, was a previous PI in my current institute, RIKEN Center for Developmental Biology.
The meeting embraced developmental biologists from a high variety of fields, and thus it was not that small albeit being a national meeting. So, it was organized in several parallel sessions, including some main Symposia, a couple of technical Workshops, and several sessions of contributed oral presentations (each about a common topic). Therefore, it was impossible attending to everything. I hope the reader forgives me if I focus mainly in what I’m interested in.
Hybernating hamster, picture courtesy of T. Bullmann.
The meeting opened a day before the official date (28th), with a satellite meeting in Japanese in the morning (to which I did not attend for obvious reasons) and three oral sessions in the afternoon (in English), one of them mixing topics on neural development, system biology, technological and theoretical approaches. For a start, and given that it was not the official day 1 (but day 0…), the room was not full, but still there were some discussion and even a cute picture of a hibernating hamster (see below), presented by Torsten Bullmann, of RIKEN QBiC, about his work on the protein tau and its role on the plasticity of dendrites.
It was a surprise for some of the audience, since it seemed that tau is a very well known marker for axons… but Bullmann explained that it depends on its phosphorylation state and thus you can use different antibodies against tau to mark either axons or dendrites. Other quite interesting talk was that of Kenneth Ho et al., also from RIKEN QBiC, about the Systems Science of Biological Dynamics (SSBD) database that they have created and to which any scientist can upload published data or download them, so that everyone can use them. You can find the database and more information about it here. This database looks quite good, and is distinct from other databases that contain just sequence information. A set of tools to work with the images is also integrated into the database, such as ImageJ utilities. You can have a look at the database also in this video:
On the second day (official 1st day), I attended to the joint symposium between the Spanish Society for Developmental Biology (SEBD, standing for the Spanish name of the society) and the Japanese one. This was the first time that the JSDB held a joint symposium with a society from abroad, and I would say that it was a success. Great scientists from Spain joined the meeting, both senior and young. The talk by Ángela Nieto, from the Institute for Neurosciences in Alicante and president of the SEBD, on the role of snail and other transcription factors in epithelial-to-mesenchymal and mesenchymal-to-epithelial transitions, not only during development but also during the metastatic process of cancer, woke up the curiosity of the audience in the early morning. Have a look to this great review by Nieto about this topic here. Of much interest was also the talk by Miguel Torres, from the National Center for Cardiovascular Research (CNIC) in Madrid, on how cells compete with each other to contribute to the embryonic development of mammals; and that of María Abad, from the Spanish National Cancer Research Centre (CNIO), also in Madrid, who talked about the in vivo generation of iPS cells. You can check the work by Torres here, and that of Abad, here. Ana Gradilla, a Mexican researcher who belongs to the SEBD, presented her work done at the Center for Molecular Biology Severo Ochoa (Madrid) about the very hot topic of the distribution of morphogens within exovesicles through cytonemes in Drosophila. The discussion on this work (check it out here) was also continued during a nice nijikai (Japanese word for after-party), the second day after the reception.
One important feature of this meeting was the high number of talks. Masahiko Hibi, the organizer of the meeting, said that the aim of the meeting was to give as many chances as possible to young researchers to give a talk. In this regard, there were two sessions of flash talks, one on each of the first two days, of 3 minutes of duration where the presenters had to introduce their work, and later on continue the discussions with those interested in the poster session. It was actually a success, since I haven’t seen such a lively poster session in any meeting so far. I’d like to highlight that of Yuichiro Hara, from RIKEN CDB, who presented about transcriptomic and genomic resources of the Madagascar ground gecko, a very interesting emerging model organism. They are now constructing a database, Reptiliomix, which contains these transcriptomics resources. Keep an eye on their lab website about the anticipated release of the web server.
After the flash talks I attended one of the workshops scheduled in the meeting (at the same time that two very nice oral presentation sessions, about Early Embryogenesis and Evo-Devo, and about Morphology – I wish I could have cloned myself to attend those-). I attended the workshop because I had to give a contributed talk there. This workshop was entitled “New Genome Technologies in Developmental Biology” and was organized by Atsuo Kawahara, from Yamanashi University and RIKEN QBiC, and by Takashi Yamamoto, from Hiroshima University. The workshop was basically focused in the most recent genome editing technology, such as the CRISPR/Cas9 system, a topic that was very present during the whole meeting, highlighting the importance of these very new techniques. However, my talk was about a comparative transcriptomics analysis between turtle and chicken tissues, including the carapacial ridge, the embryo’s structure controlling the shell formation.
The second day started with the two plenary lectures of the meeting. The first one, by Alex Schier from Harvard University, was about the role of a newly described gene, toddler, in the early embryogenesis of zebrafish. The second talk was by Hans Clevers, from the Hubrecht Institute in the Netherlands who described Lgr5 as a marker for stem cells in the crypts of the intestinal epithelium. Clevers’ team could also generate intestinal organoids by controlling the expression of Lgr5, technology developed by this postdoctoral fellow Toshiro Sato. Clevers delighted the audience with beautiful animations, including those that represented clonal crypts from cells expressing different fluorescent proteins… eventually resulting in colorful intestinal epithelia. Both plenary talks were followed by many questions from the audience.
The afternoon of the second day was also followed by flash talks presentations, and after that the second workshop (“Frontiers in Developmental Biology by Unique Approaches”) and two parallel oral presentation sessions. In this case I decided to attend one of the oral sessions, about the Gene Expression and Epigenetics, where I attended an interesting talk about the differences in Shh regulation between chicken and mouse, by Takanori Amano, from the National Institute of Genetics of Japan.
Since a meeting does not consist only of science, but also of socializing events among scientists, the second day was finished by a very nice reception in a hotel near the meeting venue. It was a very relaxing time, and I could finally enjoy some time with my Spanish colleagues and discuss, among other things, about the situation of science in our country (not a very hopeful future, I would say). Some announcements that you might be interested in were about the next year’s JSDB meeting, to be held in Tsukuba, and organized by Hiroshi Wada, from the Tsukuba University; Ángela Nieto, as the president of the SEBD, announced the next meeting of the Spanish Society (together with the Portuguese Society of Developmental Biology) will be held in Madrid this year from October 13th to 15th, and it will be in association with the JSDB (there will be a couple of fellowships for young researchers to attend, so don’t forget to apply if you plan to attend!). Also, the next year’s JSDB meeting will hold a joint symposium with the Dutch Society for Developmental Biology (see this past post in the Node), in the same way that this year was with the Spanish counterpart.
The last day had 6 symposia, 3 in the morning and 3 in the afternoon. In the morning, I attended the symposium of the Asia-Pacific Developmental Biology Network, to have a glimpse of what is done in the region. I attended the talk of Xinhua Lin, at the Institute of Zoology from the Chinese Academy of Science, about tissue homeostasis by gut stem cells in Drosophila. And, finally, in the afternoon I went to the talk given by Benny Shilo, from the Weizmann Institute of Science in Israel, about the dorso-ventral patterning of the Drosophila embryo.
Overall, it was a nice meeting, not too small, and not too big. In my case, I have been working for almost four years in Japan, and it has been my first national meeting, what have allowed me to get an idea of what Japan is up to. Given the fact that I am actually thinking about continuing my scientific career here, I could learn about different institutes, universities and researchers with whom I can collaborate in the future. However, the atmosphere is much more international than I expected, and thus even if you are not working in Japan, attending this meeting is definitely worthy. So, keep an eye on the upcoming meeting in 2015 in Tsukuba, and come if you have the chance. You will not regret it!
Last month 39 people from around the world gathered together in the flagship European Molecular Biology Laboratory (EMBL) in Heidelberg, Germany to take part in the Master Course on Bioimage Data Analysis. This was the third edition of the course that had previously been held in Heidelberg and Barcelona, and is aimed at training scientists to meet the growing need to extract measurable and quantitative data from biological images. In this latest incarnation there was a meeting component (invited speaker talks, short talks selected from abstracts and a poster session), a practical component, and a strong “community building” component.
Takeo Kanade from the robotics institute at Carnegie Mellon University started off the course with a fascinating keynote lecture that not only explained some technical aspects of segmentation of biological images, he also gave a great review of the key elements that make a computer capable of “seeing”. In addition, lectures were presented by pioneering researchers such as Fred Hemprecht from the Heidelberg Collaboratory for Image Processing (HCI), Ivo Sbalzarini from the Max Plank Institute of Molecular Cell Biology and Genetics, Nadine Peyriéras from the CNRS, France and Christophe Zimmer from the Institut Pasteur. Diverse aspects of imaging were presented in these lectures, as well as radically different approaches to image analysis. However, all of them had a common thread; how do we teach a computer to “see” the information contained in digital image so that we can quantify this information in a way that is biologically meaningful.
Two underlying themes emerged from these talks. The first focused on how scientists can more easily make computers “understand” what is that they are “seeing” by providing the computers with models of both, the world, and how it appears on the other side of a microscope. Giving computers both of these models as well as the laws that rule them (-i.e. a cell can only divide into two or a nucleus cannot leave a cell-) allows computers to “see” better, requiring less human intervention and data curation. The second theme focused on how to make software designed to quantify images more user friendly. These talks focused on how tool creators (those that design the software) are minimizing the need to change a variety of parameters inside the different tools (known as parameterize) in order to obtain the desired outcome. This is important because unless the user really understands how the tool was implemented, using an application resembles more a leap of faith than a scientific decision. By incorporating computer learning with representative training sets, programmers and tool developers are simplifying the way in which the user applies a given tool, having full control of the quantification process.
The classroom component of the course consisted of structured exercises in which the students learned how to apply the different strategies for segmentation and data analysis using real biological data. Students used the image analysis platform Fiji (ImageJ) and Matlab, and R for part of the analysis. With these tools, students implemented workflows that exemplify some of the most common tasks in image analysis such as the segmentation and visualization of complex 3D structures, tracking particles and cell movements, and analysis of the speed and directionality of biological movements big and small, among others. In addition, students got “insider information” from the experts in the field, regarding how and why they use different strategies (plugins, tools and macros) to accomplish the desired image restoration and segmentation required to quantify a given feature.
Finally, but not less importantly, the attendees got to present their work in short talks and a poster session. This component of the course had a dual goal. On one hand it exemplified how diverse problems in biology are converging in the need to generate quantitative data and how the use of quantitative microscopy will clearly be a first line tool for biologists in years to come. On the other, it generated a sense of community amongst the course participants, imparting the philosophy of the course: to foment and energize the next generation of Image Analysts as members of a growing community that not only applies but also generates tools and research that will propel the specialty well into the future.
After serving on some academic selection committees recently, I’m worried about the future of some of our young scientists. Especially concerning are the number of applications where the candidate, pursuing a academic or research career, does not seem to have a understanding of what is required to put together a stand-out application for a position, and it in some cases may even be to late for them to be competitive for such positions.
It is getting tough out there on the academic job front; each position may receive well over 50-100 applications (even getting into the hundreds at larger high profile Universities).
How can we help our PhD students who want a career in academia? They need to get on top of things early on and learn what they need to achieve to be considered for research and group leader positions.
Job applications are becoming like grant applications – in a pool of excellent candidates the selection committee will look for a reason not to put you through to the short list. How are you going to make your CV and application stand out in a competitive field?
Below I have listed a few tips to get the discussion started.
“The post-doc position”
Outline your Research expertise and interests
An overview of your research interests (what about this research excites you?), can also (briefly) outline your previous research projects and their outcomes
Publications – vital
This could be split into manuscripts in preparation, manuscripts under review and accepted/published. It is critical to publish during your PhD. Your PI is likely to be very busy so it may be up to you to push to get the first drafts of the manuscript together. Plus those employers that are looking for post-doc want to see evidence that you can write!
Qualifications and Employment record
Include expected date of thesis completion or examination.
Presentations:
Contributed and invited talks at meetings/conferences and at departmental level. Never turn down opportunities to speak, it is your best shot at getting you and your research noticed. As they say, every talk is a job talk.
Laboratory skills
Describe the skills/techniques that you are proficient in and have experience with. The employer may be looking for new skills you can bring to the group as well as experience required for the project.
Teaching and mentoring experience
Evidence of teaching and mentorship can be important.
This may include student demonstrating and guest lectures. List the students that you helped to supervise and the relevant output (eg thesis or if their data ended up in your publication).
Grants, awards, scholarships
List any travel awards, PhD scholarship, anything you applied for $ and got it. Any prizes at conferences and meetings = Peer esteem.
Professional activities and skill development
Workshops, reviewing grant applications, committee work, outreach activities. Take the opportunities to be involved all these activities when they are offered or seek them out yourself.
Professional memberships – Membership to societies. Join up – it is often very cheap for students and early career researchers plus they offer travel grants and awards. The COVERLETTER – Please write one and address the advertised project, otherwise it looks like you are just applying for everything. Be enthusiastic about the proposed project/research area and address how your skill sets met the selection criteria. If you don’t have all the skills listed (and it is rare to have them all), then you can address this to say why you aren’t proficient in this area, and refer to your ability to quickly pick up new skills.
“Lecturership position”
Publications (vital):
The number of publications since PhD is often used as a marker of research output (often around 2 publications per year is considered great but this will vary depending upon the research field). Impact factor is also taken into consideration eg a fewer number of high impact papers can be weighted similarly to a CV with an overall higher number of publications. Where possible you might want to include impact factor, number of citations of each paper, H-index, ranking of the journal within your field of research (Eg this journal is ranked 2nd out of 50 journals in the field of developmental biology) = essentially showing the measure of the impact of your work in the field of research.
For multi-author publications – what was your contribution? (this can be important if you aren’t the first or last author in a long list of authors, even if it is a Nature or Science paper).
Previous grant funding
Provide evidence of research grant success – this proves you have ideas and they are fundable. It is expensive to do research, you need to show you can fund your research (and bring some extra income into the department). These don’t have to be large grants; keep an eye out for smaller funding opportunities to get you your first grants as PI.
Research interests and projects
You need to outline the research directions for the programme of research you will establish in your new group. Include how they fit with the department (and existing facilities) and other members of the university. What funds will you apply for? Do you have any established collaborations?
Leader qualities
Examples of how you have lead research projects/programmes in the past.
Supervision of research students – what was the outcome (eg did they contribute to publications?)
Teaching
Develop a teaching profile or teaching philosophy. Why do you want to teach, what kind of teacher do you want to be? Include any evidence of teaching experience. Look at the courses within the department you are applying, discuss what courses can you contribute too.
Other helpful additions to the CV
Any community engagement or outreach activities.
Presentations such as conferences but include others too and note if you were an invited speaker. University service such as work on committees (there will be a lot of this if you get the job!) Evidence that you understand the compliance issues that must be met and administrated by you as a group leader such as animal ethics, lab safety, biological compliance.
Your employment record – if you have any career gaps it may help to explain them.
The Coverletter. Do one!
Why do you want to move to this department/university? Specifically address the selection criteria.
The Final word –It is going to take planning! Start planning early in your career for the position you want in 5 years time. Don’t turn down opportunities and take a few risks.
Electroporation: an efficient technique for embryologists
During embryonic development, the specification of different cell types giving rise to the future organs involves a precise spatiotemporal regulation of cell proliferation, migration, and differentiation. Studying these processes requires tools to manipulate gene expression locally in the developing embryo.To this aim, embryologists have widely used the technique of electroporation, consisting in the delivery of exogenous molecules (such as nucleic acids) into targeted cells through electric permeation of the plasma membrane(Calegari et al., 2004; Escoffre et al., 2009; Nakamura and Funahashi, 2012; Swartz et al., 2001). Localised electroporation is achieved by two main approaches depending on the position and the geometry of the targeted tissue (Fig. 1). If the zone of interest surrounds a natural body cavity, one can generate an electric field with large electrodes and inject locally the exogenous molecule (Fig. 1a; Itasaki et al., 1999; Soares et al., 2008). Otherwise, the electroporation solution is homogeneously applied and the electric field is then spatially restricted by, for instance, using needle-shaped electrodes placed in close proximity to the targeted area (Fig. 1b; Davidson et al., 2003; Momose et al., 1999). One important drawback of this second strategy is that electrolysis occurring at the needles surface during pulse application generates harmful chemical species that may result in cell damage and poor embryo survival (Kim et al., 2008; Wang et al., 2010; Wang and Lee, 2013).
Figure 1. Strategies for localised electroporation in early post-implantation mouse embryos. (a) The first technique involves a uniform electric field and a localized injection of nucleic acids. (b) The second technique relies on a localized electric field and a homogenous nucleic acid solution. (c) Our technique involves dielectric guides where electric field is spatially restricted inside confined channels filled with electroporation buffer.
When embryology meets microfabrication
To circumvent these problems, we developed a system where the electric field generated by large electrodes is conveyed to the targeted zone by narrow channels, also known as dielectric guides, filled with electrolyte (Fig. 1c; Mazari et al., 2014). In this way, efficient electroporation with reduced cell damage is obtained as the electroporated tissue is exposed to a confined electric field while lying far away from the electrode. Moreover, the sample is immobilized by suction in front of the channel, therefore obviating the need for micromanipulation. Interestingly, dielectric guide-based electroporation devices for single cells in culture had been successfully adapted to the on-chip format, with microfabricated electrodes and fluidic channels (Fox et al., 2006; Wang et al., 2010; Wang and Lee, 2013). To design similar tools able to target small embryos or organ explants, the most important parameters are the dimensions of the dielectric guide. Indeed, transfecting as few as 4 or 5 cells of a specific tissue, which approximately represent an average surface of tens square micrometers, requires the guide aperture to be of a similar size. Derived from the microchip industry, microfabrication techniques are particularly well-suited to create microstructures. Through a straightforward and quite simple process, these microengineering techniques then enable the reproducible production of defined shapes and positions at the micrometer scale.
An optimised and collaborative approach
As a proof of principle, we chose to target small cell populations in the visceral endoderm (VE), the outer epithelium of mouse embryos at embryonic day 5.5 of development (E5.5) (Fig. 2a-b). This is a critical time for the establishment of embryonic polarity, with distal VE (DVE) cells undergoing a stereotypical migration that will define the future anterior side (Takaoka and Hamada, 2012). However, existing electroporation procedures for this stage face poor embryo survival and poor reproducibility. First, we conceived an electrical model of E5.5 embryos and used it in numerical simulations to compare the outcome of different electroporation strategies. This analysis demonstrated that the proposed dielectric guides device would permeabilize a more restricted area than current electroporation systems. In addition, we experimentally and systematically investigated pore generation and cell viability to determine the best electrical conditions for efficient electroporation of DVE cells. Constant optimisation by alternative simulations and experimental tests, resulted in the development of dielectric guide-based devices and associated protocols to locally electroporate E5.5 mouse embryos in an efficient, reproducible, and safe way (Fig. 2a-c).
Figure 2. Use of the dielectric guide-based device on an E5.5 embryo. (a) View of the whole system included between the electrodes. An E5.5 embryo (blue arrow) is positioned with its DVE abutting the dielectric guide connected to the cathode, so as to target precisely this tissue. (b) Close-up view of the same embryo. (c) Image of a transgenic Hex-GFP E5.5 embryo electroporated in the DVE with pCAG-mCherry DNA. (d) Close-up view of migrating mCherry expressing cells. The red arrows show cell projections.
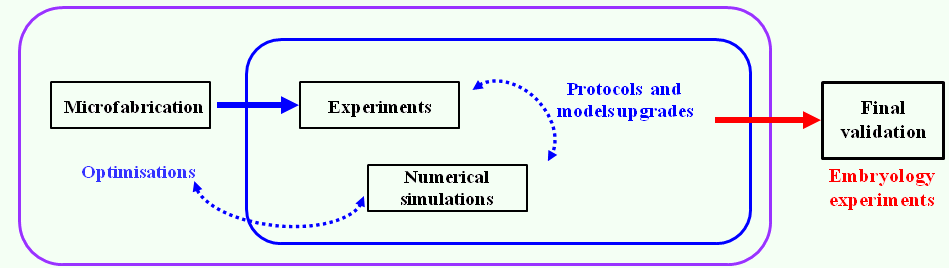
We combined our electroporation technique, transgenic mouse lines, and live imaging to study the behavior of VE cells during DVE migration between E5.5 and E6.0 (Fig. 2c). We were able to specifically label subpopulations of follower DVE cells that so far had been difficult to visualize, and to monitor the production of dynamic cellular projections during their migration (Fig. 2d). Furthermore, we demonstrated that our microdevice can be used to electroporate tissues in a wide range of embryonic contexts just by adapting the guides dimensions. A distinctive feature of our work lies in the proposed interdisciplinary approach that brings together expertise from biology to physics. Our strategy coupling numerical simulations, prototype microfabrication, and in vivo testing, provides an optimized and rigorous framework for the design of other tailor-made electroporation devices (Fig. 3). We hope that this work will provide a stimulating example ofthe electrifying interest of microfabrication approaches to developmental biologists.
Figure 3. Interdisciplinary and auto-optimised strategy used to conceive a dielectric guide-based tool to electroporate locally, reproducibly, and safely small cells populations from external tissues of embryos or organ explants.
Bibliography
Calegari, F., Marzesco, A., Kittler, R., Buchholz, F., & Huttner, W. (2004). Tissue-specific RNA interference in post-implantation mouse embryos using directional electroporation and whole embryo culture Differentiation, 72 (2-3), 92-102 DOI: 10.1111/j.1432-0436.2004.07202002.x
Davidson, B., Tsang, T., Khoo, P., Gad, J., & Tam, P. (2003). Introduction of cell markers into germ layer tissues of the mouse gastrula by whole embryo electroporation genesis, 35 (1), 57-62 DOI: 10.1002/gene.10166
Escoffre, J., Portet, T., Wasungu, L., Teissié, J., Dean, D., & Rols, M. (2009). What is (Still not) Known of the Mechanism by Which Electroporation Mediates Gene Transfer and Expression in Cells and Tissues Molecular Biotechnology, 41 (3), 286-295 DOI: 10.1007/s12033-008-9121-0
Fox, M., Esveld, D., Valero, A., Luttge, R., Mastwijk, H., Bartels, P., Berg, A., & Boom, R. (2006). Electroporation of cells in microfluidic devices: a review Analytical and Bioanalytical Chemistry, 385 (3), 474-485 DOI: 10.1007/s00216-006-0327-3
Itasaki N, Bel-Vialar S, & Krumlauf R (1999). ‘Shocking’ developments in chick embryology: electroporation and in ovo gene expression. Nature cell biology, 1 (8) PMID: 10587659
Kim, J., Cho, K., Shin, M., Lee, W., Jung, N., Chung, C., & Chang, J. (2008). A novel electroporation method using a capillary and wire-type electrode Biosensors and Bioelectronics, 23 (9), 1353-1360 DOI: 10.1016/j.bios.2007.12.009
Mazari, E., Zhao, X., Migeotte, I., Collignon, J., Gosse, C., & Perea-Gomez, A. (2014). A microdevice to locally electroporate embryos with high efficiency and reduced cell damage Development, 141 (11), 2349-2359 DOI: 10.1242/dev.106633
Momose, T., Tonegawa, +., Takeuchi, J., Ogawa, H., Umesono, K., & Yasuda, K. (1999). Efficient targeting of gene expression in chick embryos by microelectroporation Development, Growth and Differentiation, 41 (3), 335-344 DOI: 10.1046/j.1440-169X.1999.413437.x
Nakamura, H., & Funahashi, J. (2013). Electroporation: Past, present and future Development, Growth & Differentiation, 55 (1), 15-19 DOI: 10.1111/dgd.12012
Soares, M., Torres-Padilla, M., & Zernicka-Goetz, M. (2008). Bone morphogenetic protein 4 signaling regulates development of the anterior visceral endoderm in the mouse embryo Development, Growth & Differentiation, 50 (7), 615-621 DOI: 10.1111/j.1440-169X.2008.01059.x
Swartz, M., Eberhart, J., Mastick, G., & Krull, C. (2001). Sparking New Frontiers: Using in Vivo Electroporation for Genetic Manipulations Developmental Biology, 233 (1), 13-21 DOI: 10.1006/dbio.2001.0181
Takaoka, K., & Hamada, H. (2012). Cell fate decisions and axis determination in the early mouse embryo Development, 139 (1), 3-14 DOI: 10.1242/dev.060095
Wang, M., Orwar, O., Olofsson, J., & Weber, S. (2010). Single-cell electroporation Analytical and Bioanalytical Chemistry, 397 (8), 3235-3248 DOI: 10.1007/s00216-010-3744-2
Wang, S., & Lee, L. (2013). Micro-/nanofluidics based cell electroporation Biomicrofluidics, 7 (1) DOI: 10.1063/1.4774071
Here is June’s round-up of some of the interesting content that we spotted around the internet:
News & Research:
– Fly developmental geneticist Walter Gehring sadly passed away this month. His influential research career included the discovery of the homeobox genes.
– This last month saw new updates in the STAP case controversy- Haruko Obokata agreed to retract both papers, while a committee investigating the case has made recommendations that could potentially lead to the closing down of the RIKEN Centre for Developmental Biology.
– Jordan passed a law to control research and therapy using human stem cells derived from embryos, the first such legislation in the region. In Nature, Elena Cattaneo and Gilberto Corbellini wrote about their fight against pseudoscience in stem cell treatments in Italy.
– Mosaic, published by the Wellcome Trust, published a great piece this month about ageing and development.
– Model organisms at the MPI-CBG in Dresden are not just used for research but also to predict footbal scores- here are the scientific oracles for this World Cup! [in German] And if you are following the World Cup, here is how football compares with a PhD by PHD comics.
– And the Weigel lab (Max Planck Institute for Developmental Biology in Tübingen, Germany) continues the tradition of science-themed parodies of popular songs by releasing their ‘Weigel style’ video as part of their take over of the eLife twitter account:
Keep up with this and other content, including all Node posts and deadlines of coming meetings and jobs, by following the Node on Twitter.
Once upon a time, physicists got curious about the cytoskeleton. They characterised the cytoskeleton – using tools of soft matter, statistical and polymer physics – as a mesoscale material whose physical properties govern its dynamics. They showed that the cytoskeleton is an interconnected scaffold that, depending on the time scale, can behave like a shape-morphing gel (slow) or like an elastic polymer network (fast)1. Concurrently, biochemists showed how molecules interact to mediate the architecture of this scaffold. Together, biochemists and physicists have illustrated an emergent picture of comprehensive cytoskeletal function; they enabled a fuller comprehension of, for example, cell migration2, maintenance of cell shape3, and morphogenesis4. Essentially, the two lines of research linking the molecular scale to the polymer scale in the cytoskeleton have amplified one another in an entirely co-dependent manner.
Interestingly, if the cytoskeleton exists solely to maintain cell integrity, and drive migration and molecular traffic flow, one would think cytoskeletal polymers would begin and end at the cellular surface. However, they do not: they are vitally connected, through nesprins, lamins and the LINC complex5,6, to the nucleus and the genetic machinery of the cell7. Why is the genetic machinery of the cell coupled to forces from the outside world? This is an especially vexing question to ask of embryonic stem (ES) cells, given that they lack the Lamin A/C protein8 that largely provides the nucleus its mechanical integrity9. Observe below the dramatic shape changes of the ES cell nucleus within one minute (click to play):
These shape changes are generated by the cytoskeleton. However, some might say the cell is devoted to a pre-ordained genetic program independent of the outside world modulo chemical signaling. Then why does the nucleus coordinate its shape with cytoskeletal dynamics? One potential explanation is mechanotransduction, which posits that forces from the outside world are transduced into changes in cell function. An idea within the mechanotransduction framework is that cytoskeleton-driven nuclear shape changes and subsequent rearrangements of chromatin precipitate changes in gene expression. It is now high time – in the days of CHiP and HiC highlighting chromosomal molecular interactions10,11 – for physicists to do for the nucleus what they have done for the cytoskeleton: provide a material foundation.
Against this backdrop, we set out to understand how ES cell nuclei respond to forces.
Specifically, we wished to know how nuclei of ES cells exiting pluripotency respond to forces. We investigated mouse ES cells maintained in a naïve pluripotent state using inhibitors described elsewhere12. By removing the inhibitors from the medium, ES cells begin to differentiate, and after approximately one day in transition, they exit pluripotency and prime for differentiation. We were especially interested in comparing these transition ES cells – the ones that are exiting the pluripotent state – to naïve pluripotent ES cells. Details of how we defined naïve and transition ES cells can be found in our recent work published in Nature Materials10.
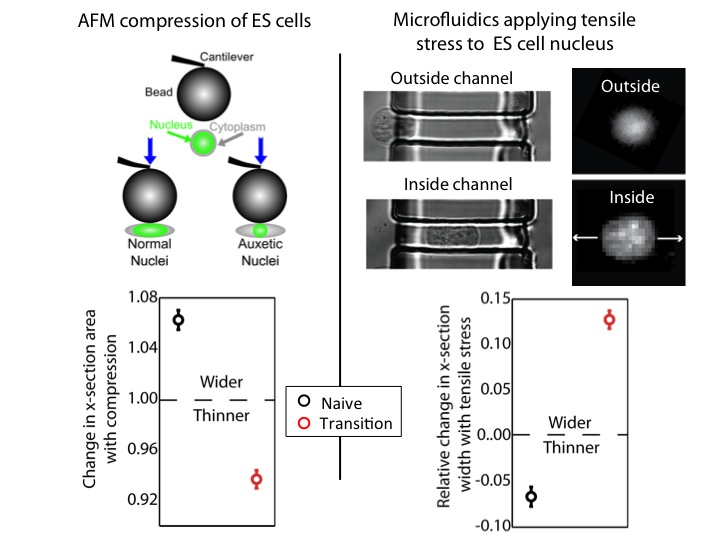
We used an atomic force microscope (AFM) to compress naïve and transition ES cells. We found that the nuclei of transition ES cells, in contrast to naïve ES cells, shrank in cross-section upon compression. This counters expectations. If one presses, for instance, on a ball, couch cushion, or plastics, they would be surprised if these items shrank in cross-section when they press. Nevertheless, that’s what happens to the transition ES cell nucleus.
For further exploration, we tested what happens when tensile stresses are applied via the cytoskeleton. We developed a microfluidic system comprising channels smaller in cross-section than ES cells, but larger than average nuclei. The cytoplasm elongates, and applies tensile forces to the nucleus. Ordinarily, with applied tensile force a material, such as a rubber band, becomes thinner in cross-section and approximately conserves volume. This is what occurs with naïve nuclei, and nuclei of other cell types we investigated (including lineage primed cells). However, transition nuclei – against anticipation – expand in cross-section with applied tensile stress; indeed, the nuclear volume increases by up to 50%.
At left, we show that by compressing transition, but not naive, ES cells with AFM, we see that the nucleus shrinks. At right, we show that by using our microfluidic system to stretch cells and apply tensile forces to the nucleus, the nucleus gets wider in cross-section in transition ES cells, but not naive ES cells.
At left, we show that by compressing transition, but not naive, ES cells with AFM, we see that the nucleus shrinks. At right, we show that by using our microfluidic system to stretch cells and apply tensile forces to the nucleus, the nucleus gets wider in cross-section in transition ES cells, but not naive ES cells.
A material that expands in cross-section when stretched and shrinks in cross-section when compressed is called auxetic13. It is an unusual material property (one must imagine a rubber band getting fatter when stretched) not generally observed in biological materials. It is used for materials like bulletproof vests and soundproofing, due to its ability to immediately absorb impact energy throughout the material instead of just at the impact point. It is also a useful property for materials used for absorption – like sponges – due to the large volume gain and consequent opening of space resulting from a stretch (consider a sponge mop that is pulled, expanding potential fluid pockets in the material).
We investigated possible mechanisms for this unusual nuclear property. We looked for differences in nuclear envelope proteins such as Lamin B1 and Nesprins, but found no meaningful difference between transition and naïve ES cells. However, electron microscopy images showed that transition nuclei were more granular and less dense than naïve nuclei. Therefore, we induced chromatin decondensation using Trichostatin A (TSA)14. With TSA, we provoked an auxetic nucleus in naïve ES cells. This led us to conclude that auxetic nuclei are due, at least in part, to increased global chromatin decondensation in transition ES cells. This is a relatively surprising result from a biological perspective, given the conventional wisdom that naïve ES cells are in state of minimum global chromatin decondensation. However, given that auxetic materials tend to be low density, it was entirely predictable from the standpoint of materials science.
Finally, we addressed the potential function of an auxetic nucleus. We suspected that the function might lie in the dramatic absorptive properties implied by auxeticity. To explore this potential function, we first loaded both transition and naïve ES cells with fluorescein, which in cells becomes more highly concentrated in the cytoplasm than the nucleus. We used our microfluidic technique to apply tensile stresses to the nucleus, and observed fluorescein flowing from the cytoplasm into the nucleus in the transition ES cells, but not in the naïve ES cells. This result indicates that auxeticity opens up porous space in transition nuclei. Coming back to the possible function of an auxetic nucleus – and auxetic sponges – we now hypothesise that auxeticity may prove to be a mode of mechanotransduction. Tensile stresses would lower the free energy barrier associated with nuclear translocation, delivering differentiation specific signaling factors from the cytoplasm to the nucleus in the differentiating ES cell. This is an especially compelling idea given the transition of epiblast from a compressed epithelial phenotype to a spread mesenchymal phenotype after implantation, when they are primed for differentiation.
The idea of a nuclear sponge facilitating an increased differentiative capacity is an enticing prospect for the function of auxeticity, but there are other possibilities. We noticed with our AFM studies that transition ES cells (not naïve ES cells) stiffened with increased compression, as one would expect with an auxetic material. Possibly, this stress-driven stiffening (and conversely softening with tensile stress) could have a significant effect on dynamic sorting of nascent tissue layers within the developing embryo.
Ultimately, there is much work to be done to understand the how and why of auxetic nuclei in ES cells. It is, however, exemplary of new discoveries in biology made by applying physics to biological systems. We hope this discovery will be coupled to emerging awareness of molecular and mechanical signaling in the nucleus to generate a richer comprehension of pluripotency, differentiation, and embryogenesis.
1. Deng, L., Trepat, X., Butler, J., Millet, E., Morgan, K., Weitz, D., & Fredberg, J. (2006). Fast and slow dynamics of the cytoskeleton Nature Materials, 5 (8), 636-640 DOI: 10.1038/nmat1685
2. Gardel, M., Schneider, I., Aratyn-Schaus,, Y., & Waterman, C. (2010). Mechanical Integration of Actin and Adhesion Dynamics in Cell Migration Annual Review of Cell and Developmental Biology, 26 (1), 315-333 DOI: 10.1146/annurev.cellbio.011209.122036
3. Paluch, E., & Heisenberg, C. (2009). Biology and Physics of Cell Shape Changes in Development Current Biology, 19 (17) DOI: 10.1016/j.cub.2009.07.029
4. Lecuit, T., Lenne, P., & Munro, E. (2011). Force Generation, Transmission, and Integration during Cell and Tissue Morphogenesis Annual Review of Cell and Developmental Biology, 27 (1), 157-184 DOI: 10.1146/annurev-cellbio-100109-104027
5. Lombardi, M., Jaalouk, D., Shanahan, C., Burke, B., Roux, K., & Lammerding, J. (2011). The Interaction between Nesprins and Sun Proteins at the Nuclear Envelope Is Critical for Force Transmission between the Nucleus and Cytoskeleton Journal of Biological Chemistry, 286 (30), 26743-26753 DOI: 10.1074/jbc.M111.233700
6. Swift, J., Ivanovska, I., Buxboim, A., Harada, T., Dingal, P., Pinter, J., Pajerowski, J., Spinler, K., Shin, J., Tewari, M., Rehfeldt, F., Speicher, D., & Discher, D. (2013). Nuclear Lamin-A Scales with Tissue Stiffness and Enhances Matrix-Directed Differentiation Science, 341 (6149), 1240104-1240104 DOI: 10.1126/science.1240104
7. Kind, J., Pagie, L., Ortabozkoyun, H., Boyle, S., de Vries, S., Janssen, H., Amendola, M., Nolen, L., Bickmore, W., & van Steensel, B. (2013). Single-Cell Dynamics of Genome-Nuclear Lamina Interactions Cell, 153 (1), 178-192 DOI: 10.1016/j.cell.2013.02.028
8. Pagliara, S., Franze, K., McClain, C., Wylde, G., Fisher, C., Franklin, R., Kabla, A., Keyser, U., & Chalut, K. (2014). Auxetic nuclei in embryonic stem cells exiting pluripotency Nature Materials, 13 (6), 638-644 DOI: 10.1038/nmat3943
9. Pajerowski, J., Dahl, K., Zhong, F., Sammak, P., & Discher, D. (2007). Physical plasticity of the nucleus in stem cell differentiation Proceedings of the National Academy of Sciences, 104 (40), 15619-15624 DOI: 10.1073/pnas.0702576104
10. Bickmore, W., & van Steensel, B. (2013). Genome Architecture: Domain Organization of Interphase Chromosomes Cell, 152 (6), 1270-1284 DOI: 10.1016/j.cell.2013.02.001
11. Nagano, T., Lubling, Y., Stevens, T., Schoenfelder, S., Yaffe, E., Dean, W., Laue, E., Tanay, A., & Fraser, P. (2013). Single-cell Hi-C reveals cell-to-cell variability in chromosome structure Nature, 502 (7469), 59-64 DOI: 10.1038/nature12593
12. Ying, Q., Wray, J., Nichols, J., Batlle-Morera, L., Doble, B., Woodgett, J., Cohen, P., & Smith, A. (2008). The ground state of embryonic stem cell self-renewal Nature, 453 (7194), 519-523 DOI: 10.1038/nature06968
13. Evans, K.E., & Alderson, A. (2000). Auxetic Materials: Functional Materials and Structures from Lateral Thinking! Advanced materials, 12 (9), 617-628
14. Chalut, K., Höpfler, M., Lautenschläger, F., Boyde, L., Chan, C., Ekpenyong, A., Martinez-Arias, A., & Guck, J. (2012). Chromatin Decondensation and Nuclear Softening Accompany Nanog Downregulation in Embryonic Stem Cells Biophysical Journal, 103 (10), 2060-2070 DOI: 10.1016/j.bpj.2012.10.015
This story starts with me shaking and nervous and stumbling through my mid-term thesis committee meeting presentation. Not only was I presenting the results from the first half of my PhD but on top of that, I was presenting this work to Francois Guillemot, basically a legend in the field of developmental neurobiology. Luckily for me, my nervousness was completely unfounded and I quickly learned that Francois is not only incredibly knowledge but also very friendly and not at all intimidating. I presented the data I obtained that demonstrates an essential role for microRNA-9 (miR-9) in regulating the quiescent state of neural stem cells (NSCs) in the adult zebrafish pallium. After providing me with a lot of helpful insight and advice he generously offered to have me come to his lab to look at miR-9 in the NSCs of the adult mouse brain. Additionally, his lab had recently optimized a protocol to induce quiescence in vitro in primary adult hippocampal derived NSCs (AH NSCs) which would be a great model in which to test the expression of miR-9 in quiescent versus activated NSCs.
I immediately accepted his offer without hesitation but I soon realized that traveling to and staying in London might end up being quite expensive. Luckily for me, the money provided by the Development Travelling Fellowship enabled me to spend 3 weeks in the Guillemot lab at the NIMR in Mill Hill.
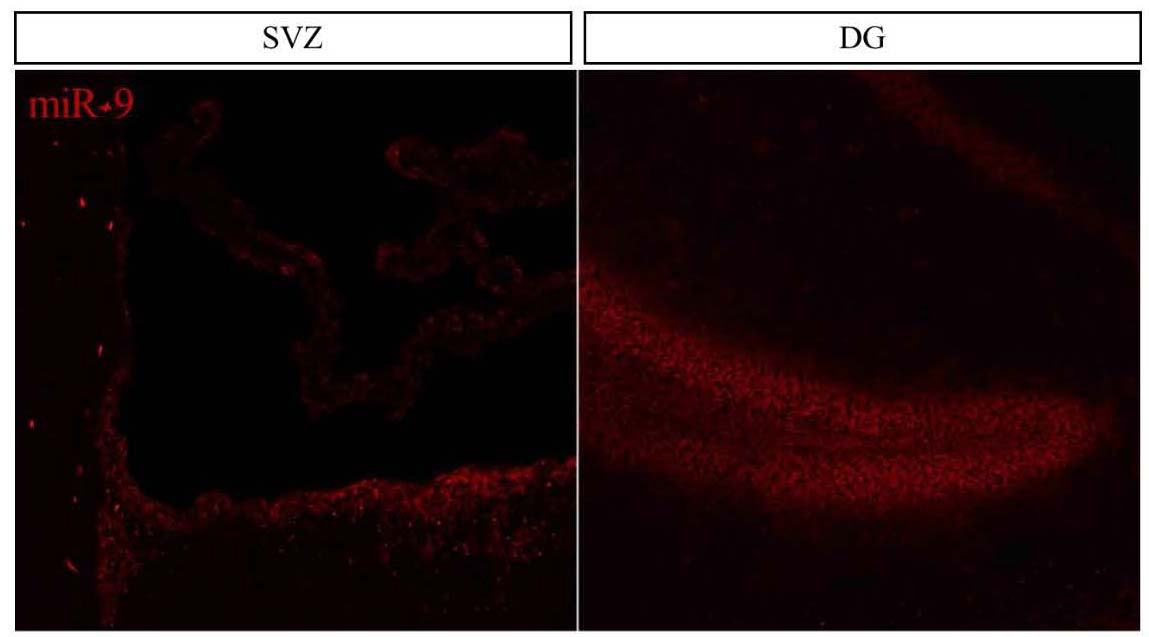
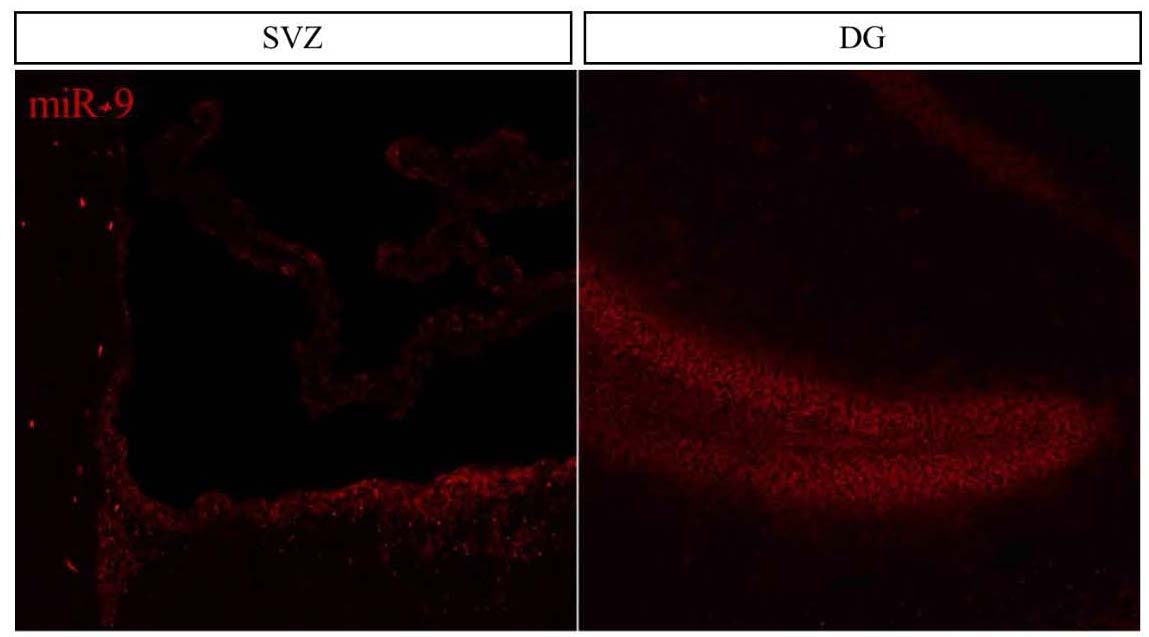
The time I spent at the NIMR in Mill Hill proved to be extremely fruitful and scientifically enriching. Upon my arrival in the Guillemot lab I was greeted by an incredibly friendly group of people and I knew that I would have a great time in the lab. Additionally, all of the mouse samples and cells were ready for me so no time was wasted and I was immediately able to start my search for miR-9 in the mouse brain. Everyone in the lab was incredibly organized and helpful which enabled me to complete a tremendous amount of work and in the end I obtained exciting results showing that miR-9 is highly expressed in the dentate gyrus (DG) and subventricular zone (SVZ), the two germinal zones of the adult mouse brain (see figure below), and also that miR-9 is much more abundant in quiescent NSCs compared with proliferating NSCs in vitro. Additionally, the NIMR is an incredibly stimulating scientific environment and I had the good fortune to attend a number of very interesting lab and departmental meetings and seminars. All of this on top of scientific discussions at the pub!
Overall, thanks to the generous Development Travelling Fellowship, I had an incredibly productive and scientifically stimulating 3 weeks in Mill Hill. I was able to obtain results that will be of great benefit to my PhD project, I learned a number of useful techniques and most importantly, I met a bunch of interesting young scientists that I am looking forward to seeing again soon!

 (1 votes)
(1 votes) (No Ratings Yet)
(No Ratings Yet)

 (5 votes)
(5 votes)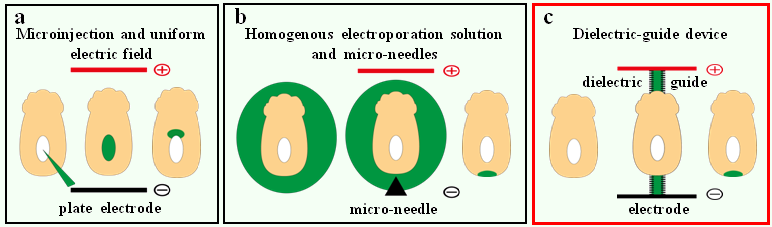


{kind=link}
{kind=link}
{kind=link}
{kind=link}