We welcome research studies, reviews, but also preliminary data and/or unpublished data that accompany a recent published article. The call for contributors is now open, and authors can submit an abstract and/or directly a manuscript for consideration. Please feel free to forward this email to colleagues who might also be interested in contributing to this special issue.
Best wishes,
Cédric & Anupama
————-
Cédric Finet, PhD National University of Singapore
Meeting Description: Stowers Research Conferences are biennial meetings that employ an innovative trainee-centered approach to inspire creative thought and collaboration in the biological sciences. Drawing on a diversity of experimental systems, the 2023 Stem and Regenerative Biology meeting will feature a broad range of perspectives on stem cell dynamics in development, homeostasis, and regeneration. We are excited to bring together thought leaders in this fascinating area of research to share new ideas, discover common themes, and forge new relationships.
When are you stepping down from the Harvard Medical Postdoc Association (HMPA) Chair position to focus on your science? This recurring question posed by a few colleagues over the last two years of my term has led me to introspect what I consider to be science. For some, it may be a fair question, as postdoc is crunch time to make or break in competitive academia, so engaging in academic community activities can be considered a futile distraction, no matter the purpose. While solely focusing on the research and getting published might be the pragmatic and effective strategy to secure a faculty position and reinforce what it means to be a successful postdoc, I see a larger disconnect with the purpose and practice of science–the dichotomy of academic culture.
Engaging in community service during my postdoc is neither my first involvement nor the first time when doctrinaire questions are posed to me. I am passionate about social justice and have been an active community member in leadership roles since high school. However, at every academic stage–B.S., M.S., PhD, and Postdoc–I have been made to feel I am not doing enough scientifically. I have been told community engagement is an extracurricular, a hobby, and an interference. I have been told to focus on my science because it is a rare opportunity that I have gotten to better my life. I have been told I am just a social butterfly. I have been told I could do ‘all this stuff’ once I have a faculty position. No wonder I feel trained to experience guilt and remorse for engaging in the community. I have constantly struggled with the expectation to fit in the academic binary–choosing academic scholarship for career progression over social justice in the academic community. I have felt the quality of my scientific work is in question because of my involvement in the scientific community. I must work harder to prove my scientific interests or let community work go. Some of my friends and colleagues feel similarly. A Black friend who is also a postdoc and actively involved in the community was recently told by their postdoc mentor that their academic excellence is not enough, meaning they are not good scientifically. It makes me think, what do we value in science? What does success mean in science? While some may argue about the extent of engagement or time management during postdoc or any other academic stage, I posit what it means to be a postdoc or rather a scientist today.
The practice of science is not just the human pursuit of knowledge and truth with rational analysis in isolation. It is a social enterprise–with diverse ideas, identities, and experiences–embedded in larger social and historical contexts, thus providing direction, substance, and meaning to our discoveries. The leaps and quests to understanding the natural world, such as cancer or water on Mars, are immersed into humanity. Therefore, the purpose of science is dynamically linked to society and its people. But what exactly is the role of scientists, then? Is it only conducting objective research and sharing the findings with the scientific community? Is it also engaging with the scientific community, policy, decision-makers, and the public? It reminds me of popular debates such as ‘Should scientists be activists?’ and ‘Should scientists be value-driven and opinionated?’. To me and many other people, science has inherent political dimensions. Indeed, science and technology studies argue that such epistemological boundaries of science and activism are problematic. Like any other social organisation, we are embedded in orderly structures of power and inequities irrespective of our rationale and logic. For example, the decisions of who gets to be an author on a research paper or which lab member goes to a conference may have influences of relative hierarchies or wealth like any social organisation. We have decades of data to support that hierarchies and biases exist in the academic system. We understand that systemic discriminatory policies and practices exist that perpetuate imperial legacies and need change. However, we let Western epistemological boundaries and frameworks shape practices, policies, and behaviours discouraging scientists from engaging in community activism. Such systemic dissociation between scientists and community members inhibits effective accountability, transparency, and responsibility in our academic system. In my understanding, cultivating, and propagating a system where we systemically continue to set the narrative for students and early career researchers (ECRs) that scholarship matters and not the community is flawed. Unuttered narratives of research priority over shared responsibilities to decolonise academic structures are flawed. The definition of scientific success is flawed.
Recently, Holden Thorp, in his remarkable editor’s blog on ‘It Matters Who Does Science’ brought up an important fact which resonated so much–scientists are conspicuously humans and we, as scientists, should embrace our humanity. Thus, instead of waiting for community engagement and conversations of bettering academia when we reach tenure, shouldn’t we set the narratives that research and scholarship are important at any academic stage, but so is community engagement–who and how do we do science? Shouldn’t we create conditions for more conducive work in meeting the needs of our people who drive science and limiting disparities that hinder the inclusion of marginalised scholars?
One of my colleagues, friends, and mentor, Akankshi Munjal, a faculty member now, supported me once during a guilt trip. She said, you should not see your postdoc as just biological research. Addressing important issues of equity and inclusion in our academic spaces is difficult work that is long ignored. You should see it as a part of your holistic postdoc experience. With shifts in academia, we are starting to see the light, where we have started to consider community work, for example, during faculty recruitment. This conversation helped me redefine my postdoc and restructure my expectations as I was constantly feeling guilty before and was considering myself a failure for not working hard during my postdoc while I was. Although I have always thought, support from Akankshi and my introspection helped me register that being a scientist is more than science itself.
Finding a mentor and community who identifies you as human and then kindly supplements you with scientific direction, community engagement, professional space, and other unique needs to be the best version of yourself can be life changing. I am fortunate to have Sean Megason as my mentor and, among others in other departments, a champion of my community work. I now take pride in doing community work in academia along with research–what I call science–and resist obstinate notions of science. I am vocal about compensation and incentives to normalise such work, so it is not disproportionately carried out, especially by marginalised identities. I do not know if I will pursue a tenure-track faculty job, but I am happy with where I am now and what I perceive of science. To be honest, I would love to teach, mentor, and conduct research. But I am finding it hard to navigate a faculty position that values my skills, expertise, and experiences, not just my academic scholarship.
Imposter syndrome is something I think all graduate students can relate to. While I’ve had my fair share of struggles with imposter syndrome in my academic career, I want to talk about a different kind of imposter syndrome, the one I felt for 25 years of my life.
Being born a woman, something always felt off. I never felt comfortable in my own skin and any attempt to appear feminine felt like wearing a mask or putting on an act. I was blessed with loving, adoring parents who always told me they just wanted me to be happy in life. I excelled in school and enjoyed learning. I played softball throughout childhood and high school and I loved the friends I made through it who became like family to me. Despite all the reasons to be happy in my life, I always felt like true happiness was just out of my grasp.
Since I was young, I had wanted to be a boy, but it was never something I thought was really possible. I was born a girl. It was the role I was cast into, so it was the role I had to play for everyone. I didn’t have the resources or the representation to understand that I was transgender or that I could live life as the man I wanted to be. By the time I had the knowledge and life experience to accept I was trans, I still wasn’t ready to make this giant change and come out to everyone as such. I had played the role of woman for so long that I was afraid there was no way to stop now. Even if I did, would the people I love accept my truth? I had so much fear, so I just held it in. I let the truth sit and eat at me, secretly knowing I’d be much happier if everyone just saw me as a man.
It wasn’t until I was 25, halfway through my second year pursuing a PhD in Genetics and Epigenetics, that I was finally ready to live for my own happiness. There isn’t one defining moment or experience that I can point to that made me finally ready to come out, I just wanted to be happy. After my now-fiancée, my PhD mentor, Dr. Swathi Arur, was actually the first person I came out to as a man. It felt far less daunting than the task of coming out to my parents, and I wanted my lab, who I saw and worked with every day, to know my truth. Swathi was supportive from the start, which if I’m being honest, I knew she would be. My lab was the same. I was absolutely terrified to tell them, but they were so happy for me. They accepted my truth and felt honored that I felt safe to share it with them. And lucky for me, I’ve received immense support from my immediate community and in graduate school since coming out. It’s not lost on me how many transgender people do not receive this same support, so I want my friends and family to know how much their love and acceptance means to me.
Since coming out, I feel like I’ve been able to thrive, despite the obvious challenges of being in graduate school and pursuing a PhD. For one, I no longer feel like I am wearing a mask around people, trying to be someone I’m not. I held so much anxiety inside me from trying to pretend to be a woman, that I didn’t even realize it was possible to live without that constant anxiety until coming out. Simply put, I feel so much more at ease with myself and in my body. That’s not to say there haven’t been stressful or awkward moments during my transition. There’s been the occasional accidental misgendering from colleagues. There was navigating doctor’s appointments and learning how to inject myself with testosterone. And there were bureaucratic piles of paperwork to get my name legally changed. So. Much. Paperwork. But it really was the best thing I could have ever done for myself. Transitioning has brought so much joy to my everyday life, and at the risk of sounding corny, it’s helped me to see that life truly is a blessing.
“It is clearly obvious that had they worked together, had that magic gone okay, then we would not talk about Watson and Crick at all. We would talk about the Franklin-Wilkins or Wilkins-Franklin structure.” Professor Matthew Cobb, University of Manchester
In the latest episode of the Genetics Unzipped podcast, Kat Arney chats with Professor Matthew Cobb about what really happened between James Watson, Francis Crick and Rosalind Franklin during the discovery of the double helix structure of DNA.
Anyone who has spent time doing research knows that being a scientist is not always sunshine and rainbows. It is great to see people announcing on Twitter that they published a new paper, or that they received a new grant, but what is less talked about are those darker days of frustrations that many researchers encounter, or the various barriers that many have to overcome.
That is why the Node has created a new blog series called ‘Honest conversations’. The aim is for people to write about the ups and downs in their academic journey and create frank conversations around less-discussed topics on the Node, such as unconventional career paths, mental health, and equity, diversity and inclusion in science.
Some of you may wonder, why does the Node think it’s important to create such a blog series? That’s because scientific research doesn’t just happen on its own; it is made possible by scientists, who are “conspicuously and magnificently human—with all the assets and flaws that humans possess”, as Holden Thorp has eloquently expressed in his Editorial ‘It matters who does science’, “…and that means that who those individuals are, and the backgrounds they bring to their work, have a profound influence on the quality of the end result.” We hope by talking more about these less-discussed topics, upcoming and established scientists alike can find reassurance and flourish in this profoundly rewarding career.
There is no strict format or word limit for these posts, but we particularly encourage posts about:
Navigating an academic career: were there moments of self-doubt of whether you’re cut out for academia? Do you have an unconventional ‘squiggly’ career path?
Mental health issues in the scientific community
Equality, diversity and inclusion in science
Troubleshooting: you can write about a tricky problem that you or your lab have been trying to troubleshoot for ages. Maybe someone from the other side of the world can help
Do you have an idea for a ‘Honest conversation’ post? Get in touch by emailing us!
Valuable new blog series from @the_Node Honest Conversations
Putting the life back into life sciences by providing a forum to discuss aspects of academic life that don't often get discussedhttps://t.co/829v8s8DfB
Our August webinar was chaired by Development Editor Cassandra Extavour (Harvard University) and featured talks from three early-career researchers studying evo-devo. Below are the recordings of the talks.
Hannah Gruner (Swarthmore College)
Talk and Q&A by Hannah Gruner
Seth Donoughe (University of Chicago)
Talk and Q&A by Seth Donoughe
Allan Carrillo-Baltodano (Queen Mary University)
Talk and Q&A by Allan Carrillo-Baltodano (No Ratings Yet) Loading...
We are pleased to announce a workshop on cell-based computational modelling within Chaste, an open-source multi-scale framework for the computational modelling of biological systems. This software library consists of object-oriented C++ code. The library enables general simulations to be undertaken and includes tools to visualise, analyse and automatically curate and store simulation results, thus expediting model development.
The workshop will introduce Chaste’s functionality, and showcase recent additions to the codebase, including varied cell-based modelling approaches and a Python interface. Through a combination of lectures, case studies, and hands-on practicals, participants will learn how to implement and run simulations of several model biological systems, focusing on applications in cell and developmental biology. Time is also reserved for participants to receive help and tuition in how to apply Chaste to novel biological problems, including their own research.
This workshop is aimed at anyone with an interest in developing, or learning more about multi-scale computational cell based models. In order to get the most out of the workshop participants should have some knowledge of C++ and some experience of mathematical modelling in the life sciences.
We have some funding to cover accommodation and travel within the UK for PhD students and early career researchers attending this workshop. Further details for how to apply for funding may be found on the online application form.
Biology Open (BiO) has appointed a new Editor-in-Chief, Daniel Gorelick. Dan is currently Associate Professor in the Center for Precision Environmental Health, and Department of Molecular and Cellular Biology at the Baylor College of Medicine in Houston, Texas, USA. He is also Director of the Zebrafish Advanced Technology Core Facility. Dan’s lab studies how endocrine-disrupting chemicals and related toxicants influence embryonic development.
In this interview, Rachel, the Manager Editor of BiO, talks to Dan about his background in development biology, his views on open access publishing, and his vision for BiO as the new Editor-in-Chief. Dan also mentions that one of his scientific heroes is Cassandra Extavour, who many of the Node and Development readers will be familiar with!
Dan was very excited when he found the elusive zebrafish special issue of Development in the head office of The Company of Biologists! (No Ratings Yet) Loading...
In this SciArt profile, we spoke to Deepti Trivedi, who has a background in neurogenetics in flies and mice, and does drawings inspired by Drosophila research.
Can you tell us about your background and what you work on now?
I grew up in Delhi, where my parents had set up a life, moving from rural part of central India, right around the time I was born. My dad was an ecotoxicologist working for the Government and mom a homemaker. Growing up I was a shy, yet confident kid. After my schooling, I took up Biochemistry at University of Delhi. My first experience with real research happened, when by alignment of several coincidences, I ended up in Bangalore to do Masters by research in a Drosophila neurogenetics lab. I can safely say, this was the first time, I knew that I wanted to be a researcher. This was a really exciting time for me to be able to ask open ended questions and wonder how they could be answered through experiments.
I was one of the beneficiaries of the travel award that The Company of Biologists offers to young researchers, using which I went to USA (SUNY, Buffalo and CSHL) for 6 months to learn electrophysiology. I used this in my Masters’ thesis work. By this time, I knew I wanted to do a PhD. I did a PhD from University of Cambridge as a Gates Cambridge Scholar where I studied phototransduction in flies. After PhD, I went to USA for a postdoctoral training (UCLA and UTMB) further studying neurogenetics in flies and mice. For past 9 years, back in India, at National Centre for Biological science, Bangalore, I set up CRISPR Cas9 based genome engineering services in flies and have been managing a core research facility.
Morgan and flies In 2021, the twitterverse decided to celebrate November 18th as Fly day, owing to the fact that w1118 fly mutant was isolated on this day. Deepti did a series of art work on this day, one of them is “Morgan and Flies” depicted here.
Were you always going to be a scientist?
Although I don’t really recall how I got interested in “science”, I feel it seamlessly integrated into the fabric of my life. Growing up, a lot of discussions at home surrounded topics of nature, plants, animals and habitats. There was a general tendency to approach regular life discussion, such as cooking or exercising, with scientific logic. There wasn’t much discussion about religion, politics, history, business, money or any other topics. So, looking back, the general outlook was already quite skewed towards science. However, taking it seriously as a career choice did not come that obviously. In fact, it was drawing and art that I spent a lot of time in. I remember wanting to be an architect, an artist or a fashion designer at some point. All these interests were deeply rooted in the fact that I really enjoyed the process of drawing and working with my hands, crafting little knick-knacks. In India, at that time, the path to pursuing art as a subject lay solely within the humanities program. This meant delving into social sciences like history, geography, economics, politics, and languages, in which I had no interest, at that time. On the other hand, I loved maths and sciences. It also helped that I had fantastic science teachers in school and then in college. Although it was no brainer to take up sciences for higher studies, I was not sure what exactly I wanted to do in sciences.
Looking back, most of my life decisions have been on the go, with no long term career goals that I have passionately pursued. However, I have always gotten extremely excited about short term goals and have pursued them with great rigour and enthusiasm. As a scientist who runs a core research facility, a shared resource for many scientist, I feel, I have found exactly the kind of work that I like. I help scientists in their passionate pursuits to scientific discovery by making a small contribution to their research programs.
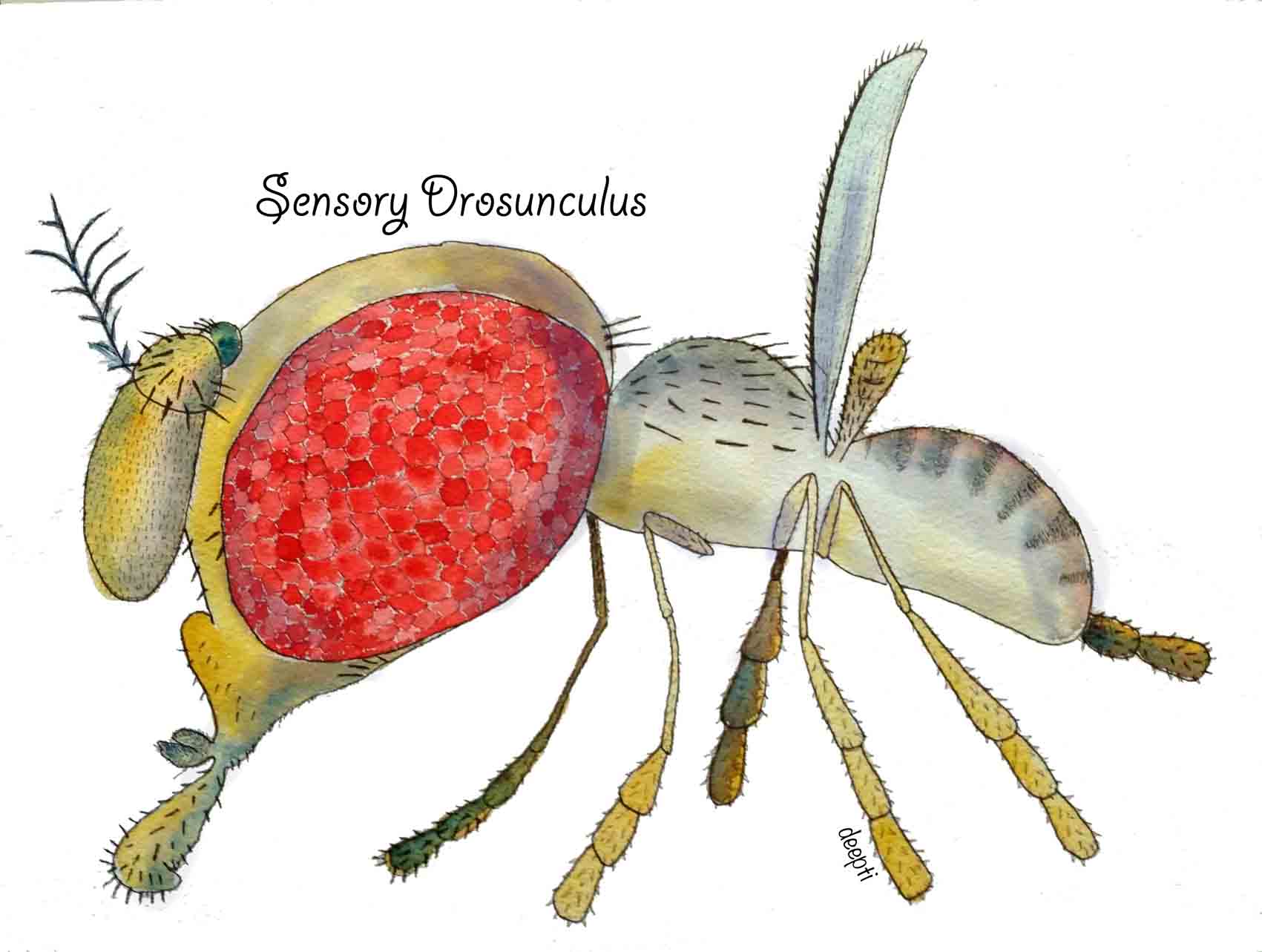
Sensory drosunculus Just like how sensory homunculus is topographic representation of sensory neuronal distribution, Deepti made a “Sensory Drosunculus” that represents that distribution of sensory neurons on Drosophila body.
And what about art – have you always enjoyed it?
As I said, I remember doing art much more vividly from my childhood than science, perhaps because there are no artists in the family. The feeling that you are doing something unique and getting appreciated for it, is kind of nice for a little kid. I doodled all the time – while studying, while listening to radio, while watch television, while on holidays with parents, inside text books, everywhere. I took part in all the art activities at school, and in the community. However, at some point during my late teens (as I picked up science that I suddenly became too passionate about), I stopped doing art completely. I picked it up again only towards the end of my PhD at Cambridge.
I now realise that art is not something I like to do all the time. I go through these phases when I do a lot of it, and then I don’t do any. Great that I didn’t choose it as a career, otherwise I would go through a lot of famines.
What or who are your most important artistic influences?
Nature is the greatest influencer. I am also deeply influenced by traditional art making and art practices.
As I mentioned there is always a long pauses which can last up to months before I pick up art and draw or paint something. However, it doesn’t mean that I am not thinking about art or in artistic way. An art process requires a lot of thinking for me and I pick up pen and paper only when an idea has clicked and has made an impact on me for a few days and stayed there and compelled me to go ahead with the idea. I usually have thought through what exactly I want to make, which medium I want to use, what size, what colour scheme etc. The actual work may require just 10 minutes, or 10 hours, but it is an immersive experience and there is enough energy that I can sit through the night finishing a piece if it requires. Once I am done, I am done and may not touch art for several months. I don’t repeat an idea or make several iterations of the same piece. It is usually a one-time process for me as if to go through a once in a life time experience and back. However, it is a deeply satisfying process.
Sometimes, I am asked to do something by friends or colleagues, which requires rounds of corrections or changes. Those are not the things that I enjoy. One more reason why this could not have been a sound career choice for me.
Circle of Life “From the day we arrive on the planet And, blinking, step into the sun There’s more to see than can ever be seen More to do than can ever be done” From “Circle of life” on which this painting is based. Drosophila has certainly been instrumental for us to see more than ever could be done otherwise.
Does your art influence your science at all, or are they separate worlds?
I am sure it does influence, but in a more sublime way rather than tangible. Both art and science, as passionate pursuits of truth, are deeply creative and immersive processes. Both require you to observe something as they are, but also to wonder what could be. I have always used drawing as a tool to understand scientific concepts, and also sometimes to avoid distractions to focus. I am a visual person and so if I am to listen to something, I usually use drawing to understand what is being said.
I feel art is more open, more non-restrictive. There are no rules to follow and so it also gives me a respite when things are not working as one wants in science or life.
What are you thinking of working on next?
There are many ideas, mostly revolving around science outreach for younger audience. I have a preteen kid and I think a lot about catering to that age group kids through art and science. I have still not pinpointed what exactly it is and hence currently going through an artistic lull period.

 (No Ratings Yet)
(No Ratings Yet) (11 votes)
(11 votes)