“I think it’s really important we have to do science that serves all. Because otherwise we’re not going to provide betterment for all, which must be of course, the goal with our science.”
Dr Cecilia Lindgren
In the latest episode of the Genetics Unzipped podcast, we’re chatting with two of this year’s Genetics Society award winners – Cecilia Lindgren, who’s an expert on the genetics of obesity and metabolic disorders, and Lucy van Dorp, who has spent the past three years tracing the spread of SARS-CoV-2 around the world.
One of the most fascinating observations that comes from comparing mammalian development is the difference in developmental tempo across species (Ebisuya & Briscoe, 2018). Mice and humans develop through a series of stereotypical events requiring conserved molecular pathways. Yet, embryogenesis takes around 60 days in humans and 20 days in mice. How mice generate similar-sized embryos containing the same structures as humans in only half the time remains unknown. Moreover, for other unconventional mammalian species, our knowledge is even more limited.
The Ebisuya lab has been addressing this question using the segmentation clock as a model system. The segmentation clock is the oscillatory gene expression found in the cells of the pre-somitic mesoderm (PSM) that controls the periodic formation of vertebrate body segments. These oscillations are cell-autonomous, and their period differs across species: around 30 min in zebrafish, 90 min in chicken, 100 min in snake, 2 hours in mouse and 5 hours in human (Matsuda et al, 2020b; Gomez et al, 2008). In our past study, in vitro recapitulation of the segmentation clock using mouse and human pluripotent stem cells (PSCs) revealed that differences in the biochemical reaction speeds, including protein degradation rates and gene expression delays, are responsible for the 2-3 times slower tempo of the human clock compared to that of the mouse (Matsuda et al, 2020a). However, whether this constitutes a general mechanism of developmental time control across mammals remained to be determined. In this study, we used PSCs to recapitulate in vitro the segmentation clock of four novel mammalian species in addition to the mouse and human: marmoset, rabbit, cattle and rhinoceros. We then used this “stem cell zoo” platform to systematically investigate the general mechanism behind the interspecies differences in developmental tempo (Lázaro et al, 2023).
The bigger the better?
Based on the results obtained comparing mouse and human developmental time, our first hypothesis was that the slower tempo of human was due to its bigger size. Several biological processes such as metabolic rates or gestation periods are known to scale with body weight. Larger animals tend to have slower metabolism, longer gestation, extended lifespan and scale most of their physical and biological properties to match their big size. Therefore, initially it made sense that developmental tempo could be regulated by similar rules. For this reason, to extend our zoo we wanted to have the largest mammal we could possibly get stem cells from. This animal turned out to be the southern white rhinoceros.
The question most people ask is: how did you manage to get rhinoceros cells? This is thanks to the work of Prof. Thomas B. Hildebrandt and colleagues in trying to save the northern white rhinoceros from extinction. For this, they have derived high quality embryonic stem cell lines of rhinoceros which can now be used for different studies (Hildebrandt et al, 2018). The first thing we did for this project was to obtain the rhinoceros stem cells and differentiate them into PSM. To our surprise, despite rhinoceros being much larger than human, their PSM cells showed a faster tempo. We then thought that maybe the slower tempo of human was due to a primate specific feature. Therefore, we searched the primate literature and found studies describing the very slow development of the common marmoset monkey. Common marmoset is a very small primate with a longer embryogenesis length than human. We obtained marmoset PSCs and, after differentiating them to PSM, we confirmed that their tempo was indeed slower than that of human. With the examples of rhinoceros and marmoset, it started to be evident that early developmental time could be uncoupled from the animal body weight, proving wrong our initial hypothesis.
We completed the zoo with rabbit and cattle cells to have a more complete phylogenetic representation of our species. Overall, our zoo contains species with adult body weights spanning from 50 grams to 2 tonnes, and gestation lengths ranging from 20 days to 17 months. These species belong to three distinct phylogenetic clades: Primates (marmoset and human), Glires (mouse and rabbit) and Ungulates (cattle and rhinoceros), constituting a diverse sampling of mammalian species unprecedented for developmental studies.
The stem cell zoo
Advancement in PSC technologies opens up new possibilities for broadening our understanding of mammalian development beyond traditional human and mouse models. The utilization of in vitro models representing various species poses a unique opportunity to conduct interspecies comparisons of cell- and tissue-autonomous processes (Figure 1). By recapitulating the segmentation clock of six mammalian species, we observed that the oscillatory period did not scale with the animal body weight but with the embryogenesis length. The biochemical kinetics of the core clock gene HES7 displayed clear scaling with the species-specific segmentation clock period. However, the cellular metabolic rates did not show an evident correlation. Instead, genes involving biochemical reactions showed an expression pattern that scales with the segmentation clock period.
Figure 1: Schematic illustration of the differentiation of mammalian PSCs towards PSM. Cells differentiated under similar culture conditions show species-specific segmentation clock periods.
A zoo of possibilities
In this study, we have focused on establishing correlations between developmental time and the different cellular parameters across species. In the future, we would like to test our hypothesis by establishing causal relationships between these processes, trying to better understand the genetic control of species-specific tempo establishment. Additionally, the stem cell zoo opens up possibilities to investigate a plethora of developmental processes across species. Other projects we have ongoing in the lab are the study of interspecies differences in brain development or heart beat rate determination. The use of stem cells allows us to study animals that are normally inaccessible in a lab but have particular features that make them interesting. We hope that the expansion of the stem cell zoo will spark further comparative studies across species.
Access the article
Jorge Lázaro, Maria Costanzo, Marina Sanaki-Matsumiya, Charles Girardot, Masafumi Hayashi, Katsuhiko Hayashi, Sebastian Diecke, Thomas B. Hildebrandt, Giovanna Lazzari, Jun Wu, Stoyan Petkov, Rüdiger Behr, Vikas Trivedi, Mitsuhiro Matsuda, Miki Ebisuya. – A stem cell zoo uncovers intracellular scaling of developmental tempo across mammals. Cell Stem Cell. 2023 Jul 6; 30: 938-949.e7
References
Ebisuya M & Briscoe J (2018) What does time mean in development? Dev 145
Gomez C, Özbudak EM, Wunderlich J, Baumann D, Lewis J & Pourquié O (2008) Control of segment number in vertebrate embryos. Nature 454: 335–339
Hildebrandt TB, Hermes R, Colleoni S, Diecke S, Holtze S, Renfree MB, Stejskal J, Hayashi K, Drukker M, Loi P, et al (2018) Embryos and embryonic stem cells from the white rhinoceros. Nat Commun 9: 1–9
Lázaro J, Costanzo M, Sanaki-Matsumiya M, Girardot C, Hayashi M, Hayashi K, Diecke S, Hildebrandt TB, Lazzari G, Wu J, et al (2023) A stem cell zoo uncovers intracellular scaling of developmental tempo across mammals. Cell Stem Cell 30: 938-949.e7
Matsuda M, Hayashi H, Garcia-Ojalvo J, Yoshioka-Kobayashi K, Kageyama R, Yamanaka Y, Ikeya M, Toguchida J, Alev C & Ebisuya M (2020a) Species-specific segmentation clock periods are due to differential biochemical reaction speeds. Science (80- ) 369: 1450–1455
Matsuda M, Yamanaka Y, Uemura M, Osawa M, Saito MK, Nagahashi A, Nishio M, Guo L, Ikegawa S, Sakurai S, et al (2020b) Recapitulating the human segmentation clock with pluripotent stem cells. Nature 580: 124–129
Organizing an art exhibition, especially a solo one, is an intimidating but truly exciting experience. For me, as a scientist and artist, the journey of curating a science art exhibition has been a transformative one. I am currently organizing a science art exhibition that will display the intricate world of developmental biology (in zebrafish) through the lens of microscopy and will be hosted from November 2023 to January 2024 in Austria. In this blog post, I will share the challenges, triumphs, and lessons I learned while organizing the exhibition until the time of writing, which is four months before the opening event.
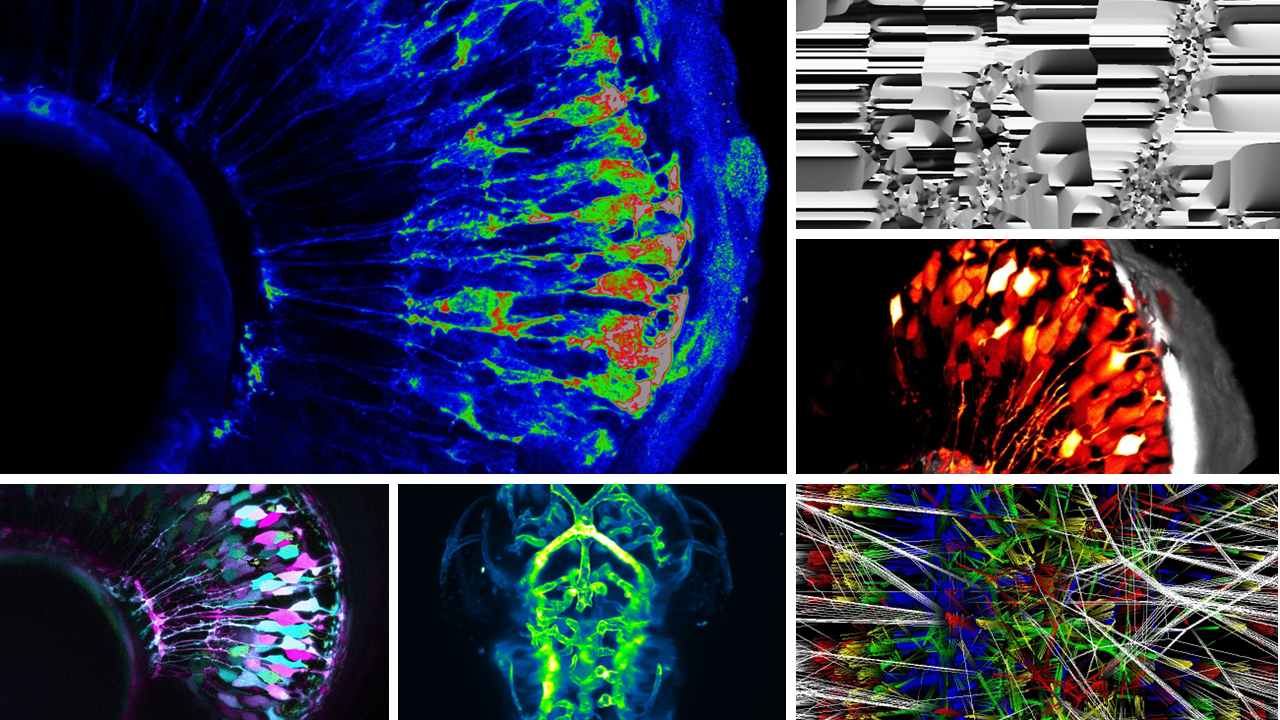
Overview of a selection of microscopy science art pieces.
The Fascination of Developmental Biology in Zebrafish
Throughout my education, I was fascinated by microscopy and developmental biology. The very fact that it is possible to light up cells in the dark and watch them move, divide, and interact in real-time is still fascinating to me. Thus, to share this fascinating world and discover new things, I shifted my career path in 2022 from academia to having my own company where we combine science and art, creating science communication and art.
For the end of 2023, we are planning a science art exhibition in Austria, where we plan to showcase the breathtaking beauty and complexity of developmental biology. To achieve this, the central artworks are a collection of 12 microscopy science art pieces. These artworks reveal the exquisite details of cells that are smaller than a single spider silk strand on canvases that are up to 1 meter in size. Using vibrant colours on black canvas and frames, I aimed to establish high-contrast works which truly emphasize the detail of the cells and tissues on display.
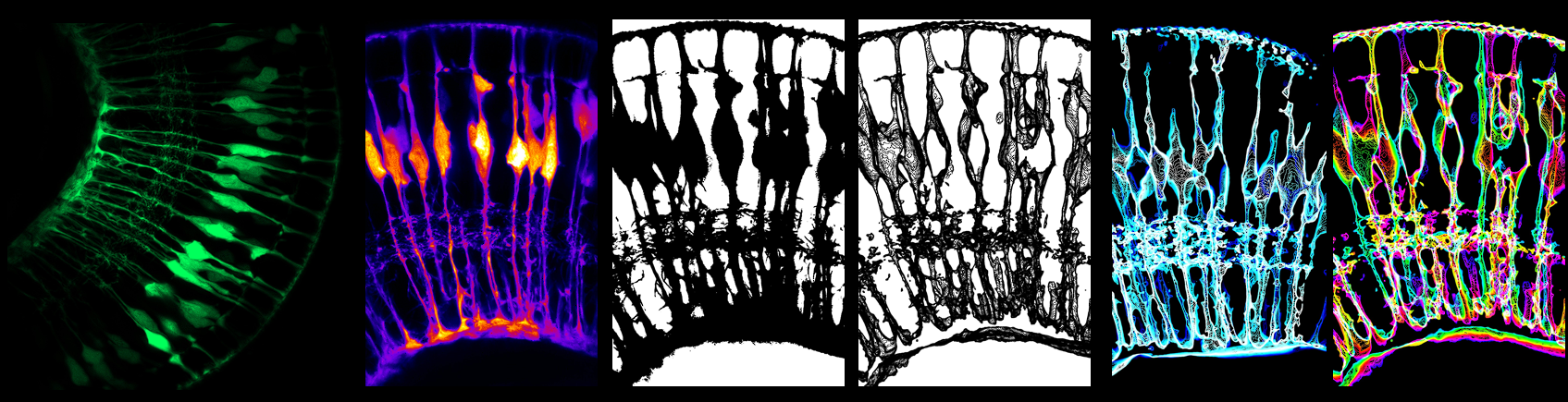
Example workflow of the science art process transforming a 3D microscopy dataset (left) to science art (right) using various image analysis steps (middle).
Exploring Various Artistic Mediums
In addition to the central microscopy science art pieces, the exhibition will feature a diverse range of artworks that have contributed to developing my art style, techniques, palettes, and composition. To offer a modern perspective on the subject matter, five contemporary science art pieces of acrylic on canvas are part of the exhibition. Alongside these, the exhibit will contain ink and watercolour works on paper as the technical requirements for these are very complementary to the acrylics on canvas pieces. The topics explored there, are portrait, landscape, and architecture. By incorporating different visual art mediums, I sought to present a comprehensive exploration of the connection between science and art, and how exploring different avenues of topics and techniques can help fine-tune others.
Establishing the Exhibition Theme
One of the primary challenges I faced was pulling together a cohesive exhibition theme. It required careful consideration and collaboration with the gallery, Hofinger BilderRahmen Tirol. Together, we worked on topics, positioning, and carefully selecting manufactured frames to ensure a seamless presentation. Through numerous discussions and consultations, we aimed to create an immersive experience that would captivate viewers and communicate the essence of developmental biology in zebrafish. My biggest tip here is to trust the expertise and experience of the gallery and framers.
Seeing your research come alive by transforming it from a digital image to a framed and mounted art piece is quite an experience.
Navigating the Artistic Learning Curve
As an individual with an academic background in science and recently transitioning to a small business owner and artist, the journey from concept to execution was a steep learning curve. Building my own company and transitioning from a scientific mindset to that of an entrepreneur and artist required especially a significant shift in perspective. Be it price negotiation, international logistics, or business networking – it is very different to academia.
The art pieces shown reflect this range, and only by embracing this path of being a scientist and artist I could merge different collections and works harmoniously into one cohesive and impactful exhibition.
Embracing the Art of Science Communication and Marketing
To ensure the success of the exhibition, early collaboration with others was essential. Together, with the gallery, colleagues, and family, we established effective marketing strategies, meticulously handpicked materials, debated font choices, and added personal touches such as handwritten notes. By going above and beyond traditional marketing techniques, we aim to create a unique experience for visitors, drawing them into the captivating world of science art. This is particularly important as we want to reach the wider public, friends, family, and academic colleagues alike. For example, choosing the right balance of science communication language is a challenging task – we want it to be scientifically correct but don’t drown the wider public in unnecessary scientific detail. My biggest tip, let a variety of people read the text and ask them to provide feedback.
Making the Invisible Visible
The process of curating an exhibition from scratch resonated deeply with the essence of microscopy itself—making the invisible visible. As a developmental biologist, my work revolved around uncovering the hidden wonders of life. Similarly, organizing an exhibition allowed me to bring together artworks and showcase the unexplored beauty that lies within the intricate world of zebrafish developmental biology.
But to be honest, it is hard work. For me, defining milestones (such as selecting the art pieces, or sending out the first invitations) is incredibly important to keep up the sense of achievement while working towards the exhibition opening.
We still have about four months before the opening, but the to-do lists seem to get longer than shorter. Why? Because we can now switch from higher level decisions (e.g. art pieces and dates) to smaller level decisions (e.g. organizing beamers and picking out food). Thus, decisions started to fan out like a tree. Where the exhibition is the trunk, the branches are higher-level decisions, and the smaller-level decisions are the leaves.
Pro Tips Summary:
Think about your why and hold on to it. E.g. for me, it is the love for sharing microscopy and science art
Explore different topics and techniques as they might fine-tune each other. E.g. digital microscopy, acrylics on canvas, watercolour on paper
Trust the years of experience a gallery and framers bring to the table. E.g. establishing contrast for microscopy art
Embrace that science and art are not exclusive. There will be some challenges and contradictions, but they are not necessarily excluding other things.
Marketing is a must, but marketing science art is particularly interesting as you want to speak to a variety of people alike. E.g. carefully consider word choices
Enjoy the process! As with every project, there are ups and downs, but that’s why considering your “Why” is so important.
Conclusion
Organizing a solo science art exhibition has so far been an unforgettable journey. I am incredibly lucky to be able to blend my passion for science and art and do this in full support of the gallery, family, friends, and colleagues. The challenges faced along the way, from establishing a cohesive theme to navigating the nuances of marketing, only served to enhance the outcome. By merging scientific expertise with artistic expression, I aim to inspire viewers and ignite a sense of wonder about the hidden realms of science and life itself. Through the medium of art, I want to bring the invisible to life, weaving together a collection that celebrates the fascination and beauty of microscopy and science.
The Inclusive STEM Teaching Project is a 6 week course designed to advance the awareness, self-efficacy, and the ability of faculty, postdocs, and doctoral students to cultivate inclusive STEM learning environments for all their students and to develop themselves as reflective, inclusive practitioners.
This is a free, online course (run through edX) that provides an excellent overview of inclusive STEM teaching. We highly recommend this course to all who teach or might teach.
Examine issues of diversity, equity and inclusion in higher education
Reflect on their own and their students’ identities and experiences
Question their assumptions about all aspects of instruction
Identify and implement learner-centered structures and strategies
Apply principles of evidence-based inclusive teaching
Use their student learning data and feedback to inform pedagogical and curricular choices
———-
This article is reposted from the ROOT & SHOOT blog (https://rootandshoot.org/the-inclusive-stem-teaching-project/). ROOT & SHOOT is a NSF-funded Research Coordination Network granted to seven plant science organizations that are seeking to “Root Out Oppression Together & SHare Our Outcomes Transparently”. In other words, we recognize that science as traditionally practiced is inherently exclusionary, and we are exploring strategies to build a more inclusive scientific culture. We invite you to visit our project home page, check out our Resources page, and follow us on Twitter (@RootAndShootRCN).
Reflections on a social media workshop that was part of the Society for Experimental Biology Centenary Conference 2023.
The use and availability of social media platforms are constantly changing, as the recent launch of Threads by Meta Platforms and our own server on Mastodon – biologists.social – nicely exemplify. For years, Twitter has been the go-to platform for biologists to share and discuss recent scientific advances, but this may well change. Regardless of what may – or may not – happen, a refresher on the different ways to use social media to engage with other scientists (and the public) seemed timely.
At the recent SEB Centenary Conference in Edinburgh (UK), Isabel Mendoza (Global Plant Council), Geraint Parry (Global Plant Council & Association of Applied Biologists) and Mary Williams (American Society of Plant Biology) teamed up to provide a workshop with exactly this in mind. They named it: “Science and Social Media. Are they really compatible? Developing Effective Science Communication Strategies”.
To start things off, Isabel Mendoza showed us (= the participants) some user statistics in order to rank the available social media platforms. This re-affirmed the notion that the platforms we often use as biologists don’t necessarily have the widest reach. Why not use Pinterest, for example, which may actually be a very efficient platform to reach new audiences?
When reaching out to new (and existing!) audiences, thinking about their needs is crucial. How can you make sure that you communicate in a way that is inclusive? The workshop organisers had plenty of examples – such as the lack of ALT-text, or wrong use of hashtags/emojis – that quickly make things inaccessible to people with marginalized identities. Even with all the right intentions…
The topic of inclusivity was actually discussed in the context of a larger movement that drives a cultural change in the plant sciences, known as the ROOT & SHOOT project. This 5-year project, combining the expertise of seven plant science organisations, aims to “remove barriers for individuals historically excluded from science based on gender, gender identity, disability status, sexual orientation, ethnicity, or race”. The title of the project refers to the goals to “Root Out Oppression Together and SHare Our Outcomes Transparently” (ROOT & SHOOT). A wonderful initiative that will hopefully be followed by many others within and outside the biological community.
There wasn’t just one take-away message from this SEB workshop. Rather, there were many different things to consider. One particular consideration that struck a chord with the participants was this: if you’re still on the fence about using social media as a scientist, there is never an ideal time to join – so why not just dive in straight away? One of the workshop participants took this advice to heart and created her Twitter profile on the spot – that’s the spirit! 💪
After a pandemic related hiatus the MBL Gene regulatory networks (GRNs) course at Woods Hole is back. It’s been refreshed with new course directors and faculty and is better than ever. It covers experimental and computational methods used to study GRNs, through highly interactive lectures, discussions, group projects, and practical tutorials.
Gene regulatory networks (GRNs) are key to the genomic control of development in animals and plants. To study GRNs requires insights from various research fields, including systems biology, developmental and evolutionary biology, as well as functional genomics, and provides an integrative approach to fundamental research questions in biology. This course introduces the concepts of GRNs, and teaches experimental and computational methods used to study them, through highly interactive lectures, discussions, group projects, and practical tutorials. We will cover a broad range of topics, including transcriptional control systems, the structural organization of hierarchical networks, developmental functions of GRN circuit modules, GRN evolution, and computational modelling using BioTapestry as well as Boolean and quantitative mathematical approaches. Students will learn how to generate GRN models based on data extracted from the literature, and will generate computational models to analyze dynamic circuit behavior. We will present and discuss a broad range of experimental approaches and how they are effectively used for studying gene regulation and developmental GRNs. Examples of experimentally solved developmental GRNs from a variety of organisms, such as flies, sea urchins, frogs, chicken, and mice, will be explored. Students are encouraged to share their research projects in a poster session, and to discuss with course faculty how to apply the approaches taught in the course to their own research questions. The course is intended for advanced graduate students, postdoctoral scholars, and faculty.
Course Faculty and Lecturers Leslie Babonis, Cornell Sarah Tulin, Canisius College Crystal Rogers, UC Davis Scott Barolo, University of Muchigan Doug Erwin, Smithsonian Mark Rebeiz, University of Pittsburgh William Longabaugh, Institute for Systems Biology Paola Oliveri, University College London Roberto Feuda, University of Leicester Nipam Patel, Marine Biological Laboratory Zeba Wunderlich, Boston University Hui-Chun Lu (grace), University College London James Briscoe, Crick Institute Andrea Streit, King’s College London Megan Martik, UC Berkeley
In this latest SciArt profile, we spoke with Kimberley Snowden, who has a background in plant molecular and developmental biology and makes science-inspired polymer clay artwork.
Can you tell us about your background and what you work on now?
I have a background in plant molecular and developmental biology. I enjoy working out at a mechanistic level how plants get their very complex forms. In particular, I have worked on plant shape or architecture, and the molecular genetic controls of plant branching for a while now. While we understand a lot about the hormones and signals that contribute to the development of branches, there is still a lot to learn, particularly at the level of the whole plant.
Leaf cross section used on a tea-light holder. A complex polymer clay cane was made to represent the cellular organisation of a leaf, including the epidermis, mesophyll, vascular bundles and guard cells. Slices of the cane were used here to cover a tea-light holder. The air spaces of the leaf were made using translucent clay.
Were you always going to be a scientist?
Yes I think so. From a reasonably early age I enjoyed the idea that we can understand the world around us using science. That if we collected data and used some logic, then perhaps we could figure out how things work. The idea that science can help us solve problems and help advance the human race is very appealing to me. Although as I’ve gotten older I’ve come to appreciate our problems require so much more than science to give us enduring solutions.
Root tip This cane is a stylised representation of the cellular organisation of a plant root tip.
And what about art – have you always enjoyed it?
I never felt I had much aptitude for art for much of my life. During my postdoc years I lived in a few very beige apartments and became very drawn to environments and art that were full of colour. I had dabbled in pottery on and off for many years, but had been frustrated that I couldn’t get the detail or colour that I wanted to. And then by chance in 2017 I stumbled into the world of polymer clay, being captivated by the colours and possibilities of this medium. Along the way I learned cane making, and have been hooked ever since.
Meristem Cane This is a full-sized cane before reduction (approximately 12 cm wide). The subject is a plant shoot tip, with the darker cells portraying the shoot apical meristem along with two axillary meristems in the developing leaf axils. This one is probably the closest to the science I do!
What or who are your most important artistic influences?
I’ve always been drawn to tactile types of art, including ceramics, glass, and sculpture. In general I like colour and abstract art, but actually many different art styles and forms appeal to me, so it’s hard to narrow that down. In the polymer clay world I am inspired by many artists, and would suggest to those unfamiliar with this medium to check out the works of people like Kathleen Dustin, Georg Dinkel, and Adam Thomas Rees. On a more personal level, a lot of the skills I learned to make canes came from the generosity of artists like Fiona Abel-Smith.
Animal cell cane This is a polymer clay cane portraying an animal cell. This cane has been reduced and is about 3 cm wide, while the original cane started out about 11 cm wide.
How do you make your art?
I start with a rough sketch of my subject, often based on looking at many images of whatever that might be. Sometimes I spend time learning about the subject as well, so that I feel I know a bit more than what I might see in photographs. Then I need to work out the level of detail needed, and spend time thinking about how to achieve that, or if it is achievable at all at the scale I work at. Some things that might be easy to draw can be very difficult to convert to a cane format. An example of that is when I recently made a version of Hokusai’s Great Wave off Kanagawa in cane form – there the small details of the wavelets took me a few attempts and approaches to get something I was happy with. Consideration of contrast and colour combinations is also very important, and not something I feel I have mastered yet. Then I usually build the components of the cane, incorporating blends, and a few other different methods before reducing those to a size that can be used in the cane. Assembly of the components into the cane happens next and requires components to be sized appropriately and for everything to be fitted well together. Rushing that aspect usually causes a lot of problems in the final cane, as the physics of reduction mean that clay will move into any gaps causing distortion. My complex canes are often built around 12 cm in diameter and 4 or 5 cm in height, and once finished the whole cane is reduced down to something more like 3 cm in diameter. Final items are made with slices of my canes, which are then cured by baking in a domestic oven.
Diatom picture. This one is based on a photograph by Anatoly Mikhaltsov of a fossil diatom from New Zealand.
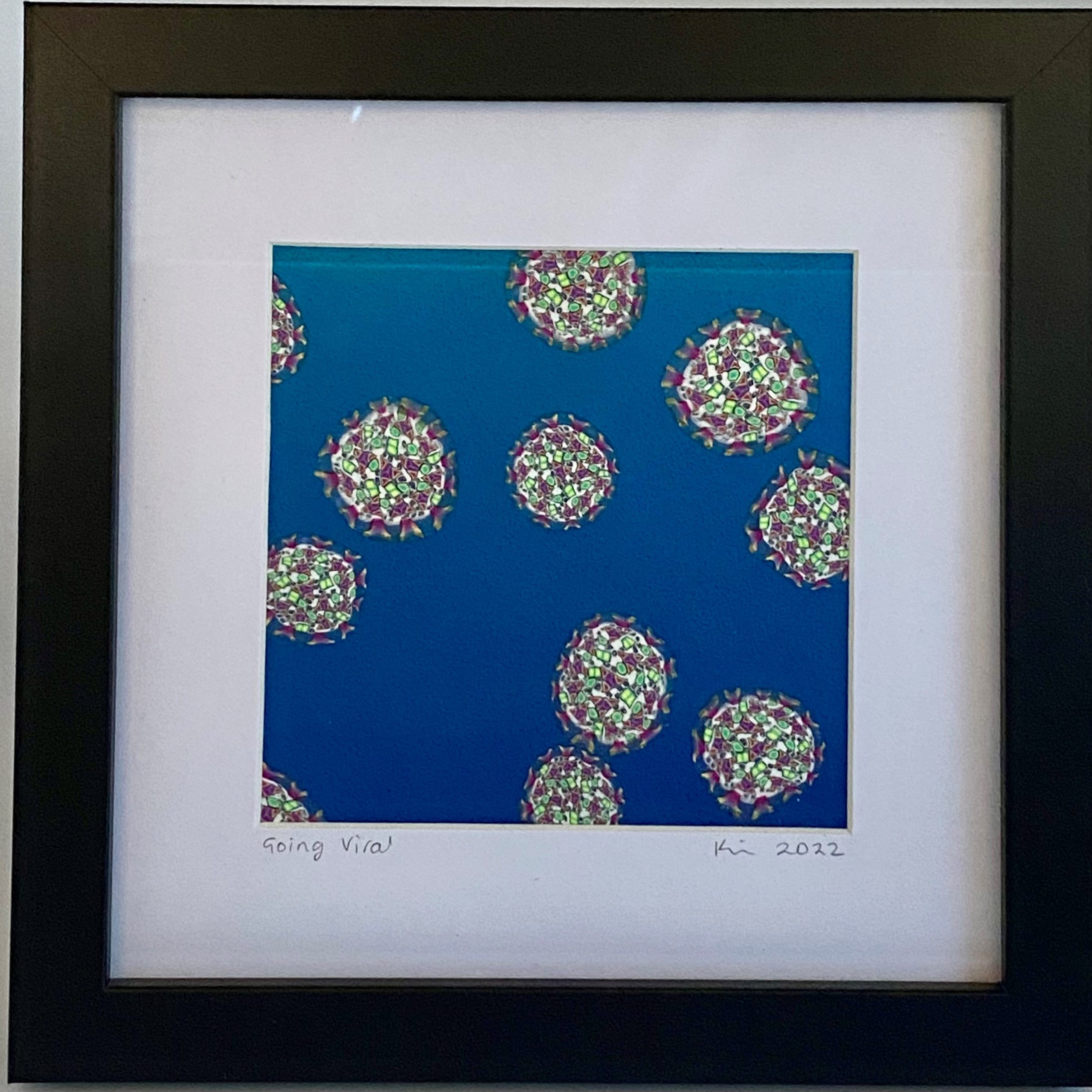
Going viral. While in lockdown in 2020, I made a cane that was my version of a SARS-CoV-2 virus. A couple of years later I used slices to assemble this piece.
Does your art influence your science at all, or are they separate worlds?
I think it is more that my science influences my art. My science background has taught me to be detail oriented which often results in me putting too much detail into my art. I have to balance portraying what is real versus what I think could convey the essence of a subject. An example of that would be my animal cell cane. There are no animal cells that look exactly like that, and the scale of different organelles is all over the place, which was necessary so that they could still be recognisable in the finished piece. In spite of that most people would still recognise it as being a cell.
What are you thinking of working on next?
I haven’t decided, and I’m always open to ideas! A few of my science canes have come from suggestions by other scientists. Some science subjects don’t lend themselves easily to a cane design, and so I sometimes think about a possible subject a long time before I start on it. Currently I have another native New Zealand flower in mind as one possibility. In the meantime, I often make kaleidoscope canes when I’m between complex cane projects, which I enjoy because of the combination of symmetry and often quite organic designs.
Native birds and plants of New Zealand. These small artworks are each approximately 10 cm square. They are made entirely of polymer clay (except a small bit of embedded wire for hanging). They each show a native New Zealand bird with a background appropriate to where these birds are found. Top left is a kererū (native wood pigeon) with a background of ngutu kākā flowers and foliage. Top right shows a pīwakawaka (also known as a fantail) with a background of kōpukapuka (also known as Chathan Islands forget-me-not, although it is not a true forget-me-not). On the bottom left is a kea (a very smart alpine parrot), with a more abstract background (working with alpine colours in a kaleidoscope format), and on the bottom right is a tūī with a background of kōwhai flowers and foliage. In the wild our kea, ngutu kākā and kōpukapuka are threatened from a conservation stand-point.Making of a complex cane. This montage shows a few of the steps involved in making a complex polymer clay cane. The top left panel shows the beginnings of the wave construction, the top middle shows construction about half way through and the top right shows the completed cane at full size (approximately 11 cm in diameter). On the middle right, you can see the cane during reduction, and the bottom right image shows the completed cane with the interior showing (here the cane is approximately 3 cm in diameter). The large image on the left shows a completed pin constructed from a slice of this cane.
Alessandra M. Norris, Ambili Bai Appu, Connor D. Johnson, Lylybell Y. Zhou, David W. McKellar, Marie-Ange Renault, David Hammers, Benjamin D. Cosgrove, Daniel Kopinke
Nikola Sekulovski, Jenna C. Wettstein, Amber E. Carleton, Linnea E. Taniguchi, Xiaolong Ma, Sridhar Rao, Jenna K. Schmidt, Thaddeus G. Golos, Chien-Wei Lin, Kenichiro Taniguchi
Eduardo D. Gigante, Katarzyna M. Piekarz, Alexandra Gurgis, Leslie Cohen, Florian Razy-Krajka, Sydney Popsuj, Hussan S. Ali, Shruthi Mohana Sundaram, Alberto Stolfi
Marta Moreno-Oñate, Lourdes Gallardo-Fuentes, Pedro M. Martínez-García, Silvia Naranjo, Sandra Jiménez-Gancedo, José L. Gómez-Skarmeta, Juan J. Tena, José M. Santos-Pereira
John R Klem, Tae-Hwi Schwantes-An, Marco Abreu, Michael Suttie, Raeden Gray, Hieu Vo, Grace Conley, Tatiana M Foroud, Leah Wetherill, CIFASD, charles B lovely
Teresa Krammer, Hannah T. Stuart, Elena Gromberg, Keisuke Ishihara, Manuela Melchionda, Jingkui Wang, Elena Costantini, Stefanie Lehr, Dillon Cislo, Laura Arbanas, Alexandra Hörmann, Ralph A. Neumüller, Nicola Elvassore, Eric Siggia, James Briscoe, Anna Kicheva, Elly M. Tanaak
Wilma Tixi, Maricela Maldonado, Ya-Ting Chang, Amy Chiu, Wilson Yeung, Nazia Parveen, Michael Nelson, Ryan Hart, Shihao Wang, Wu Jih Hsu, Patrick Fueger, Janel L. Kopp, Mark O. Huising, Sangeeta Dhawan, Hung-Ping Shih
An Chengrui, Alison M. Farley, Sam Palmer, Dong Liu, Anastasia I. Kousa, Paul Rouse, Viktoria Major, Joanna Sweetman, Jan Morys, Andrea Corsinotti, Jennifer Nichols, Jan Ure, Renee McLay, Luke Boulter, S. Jon Chapman, Simon R. Tomlinson, C. Clare Blackburn
Tania L Gonzalez, Sahar Wertheimer, Amy E Flowers, Yizhou Wang, Chintda Santiskulvong, Ekaterina L Clark, Caroline A Jefferies, Kate Lawrenson, Jessica L Chan, Nikhil V Joshi, Yazhen Zhu, Hsian-Rong Tseng, S. Ananth Karumanchi, John Williams III, Margareta D Pisarska
Ricardo Fuentes, Florence Marlow, Elliott Abrams, Hong Zhang, Manami Kobayashi, TriptiGupta, Lee Kapp, Zachary DiNardo, Felipe Montecinos-Franjola, William Vought, Charles Vejnar, Antonio E Giraldez, Mary C Mullins
Sudha Sunil Rajderkar, Kitt Paraiso, Maria Luisa Amaral, Michael Kosicki, Laura E. Cook, Fabrice Darbellay, Cailyn H. Spurrell, Marco Osterwalder, Yiwen Zhu, Han Wu, Sarah Yasmeen Afzal, Matthew J. Blow, Guy Kelman, Iros Barozzi, Yoko Fukuda-Yuzawa, Jennifer A. Akiyama, Veena Afzal, Stella Tran, Ingrid Plajzer-Frick, Catherine S. Novak, Momoe Kato, Riana D. Hunter, Kianna von Maydell, Allen Wang, Lin Lin, Sebastian Preissl, Steven Lisgo, Bing Ren, Diane E. Dickel, Len A. Pennacchio, Axel Visel
Rinaldo Catta-Preta, Susan Lindtner, Athena Ypsilanti, James Price, Armen Abnousi, Linda Su-Feher, Yurong Wang, Ivan Juric, Ian R. Jones, Jennifer A. Akiyama, Ming Hu, Yin Shen, Axel Visel, Len A. Pennacchio, Diane Dickel, John L R Rubenstein, Alex S Nord
Andrew J. Aman, Lauren M. Saunders, August A. Carr, Sanjay R. Srivatsan, Colten D. Eberhard, Blake Carrington, Dawn Watkins-Chow, William J. Pavan, Cole Trapnell, David M. Parichy
Ricardo Fuentes, Florence L. Marlow, Elliott W. Abrams, Hong Zhang, Manami Kobayashi, Tripti Gupta, Lee D. Kapp, Zachary DiNardo, Ronald Heller, Ruth Cisternas, Felipe Montecinos-Franjola, William Vought, Mary C. Mullins
Richard Francis, Jovenal T San Agustin, Heather L. Szabo Rogers, Cheng Cui, Julie A. Jonassen, Thibaut Eguether, John A. Follit, Cecilia W. Lo, Gregory J. Pazour
Laura Massoz, David Bergemann, Arnaud Lavergne, Célia Reynders, Caroline Désiront, Chiara Goossens, Lydie Flasse, Bernard Peers, Marianne L. Voz, Isabelle Manfroid
Georgios Tsissios, Anthony Sallese, J. Raul Perez-Estrada, Jared A. Tangeman, Weihao Chen, Byran Smucker, Sophia C. Ratvasky, Erika Grajales-Esquivel, Arielle Martinez, Kimberly J. Visser, Alberto Joven Araus, Hui Wang, Andras Simon, Maximina H. Yun, Katia Del Rio-Tsonis
Fabrizio E. Mancini, Paul Evelyn Alexander Humphreys, Steven Woods, Nicola Bates, Sara Cuvertino, Julieta O’Flaherty, Leela Biant, Marco A.N. Domingos, Susan J. Kimber
Joshua Hislop, Amir Alavi, Qi Song, Rayna Schoenberger, Kamyar Keshavarz F., Ryan LeGraw, Jeremy Velazquez, Tahere Mokhtari, Mohammad Nasser Taheri, Matthew Rytel, Susana M Chuva de Sousa Lopes, Simon Watkins, Donna Stolz, Samira Kiani, Berna Sozen, Ziv Bar-Joseph, Mo R. Ebrahimkhani
Kiryu K. Yap, Jan Schröder, Yi-Wen Gerrand, Anne M. Kong, Adrian M. Fox, Brett Knowles, Simon W. Banting, Andrew G. Elefanty, Eduoard G. Stanley, George C. Yeoh, Glen P. Lockwood, Victoria C. Cogger, Wayne A. Morrison, Jose M. Polo, Geraldine M. Mitchell
Christian Wiese, Miriam Abele, Benjamin Al, Melina Altmann, Alexander Steiner, Nils Kalbfuss, Alexander Strohmayr, Raksha Ravikumar, Chan Ho Park, Barbara Brunschweiger, Chen Meng, Eva Facher, David W. Ehrhardt, Pascal Falter-Braun, Zhi-Yong Wang, Christina Ludwig, Farhah F. Assaad
Facundo Romani, Susanna Sauret-Güeto, Marius Rebmann, Davide Annese, Ignacy Bonter, Marta Tomaselli, Tom Dierschke, Mihails Delmans, Eftychios Frangedakis, Linda Silvestri, Jenna Rever, John L Bowman, Ignacio Romani, Jim Haseloff
Martina Jablonski, Guillermina M Luque, Matias Gomez-Elias, Claudia Sanchez-Cardenas, Xinran Xu, Jose L de la Vega-Beltran, Gabriel Corkidi, Alejandro Linares, Victor Abonza, Dario Krapf, Diego Krapf, Alberto Darszon, Adan Guerrero, Mariano G Buffone
Anna E. Williamson, Sanuri Liyanage, Mohammadhossein Hassanshahi, Malathi S.I. Dona, Deborah Toledo-Flores, Dang X.A. Tran, Catherine Dimasi, Nisha Schwarz, Sanuja Fernando, Thalia Salagaras, Aaron Long, Jan Kazenwadel, Natasha L. Harvey, Grant R. Drummond, Antony Vinh, Vashe Chandrakanthan, Ashish Misra, Zoltan Neufeld, Joanne T.M. Tan, Luciano Martelotto, Jose M. Polo, Claudine S. Bonder, Alexander R. Pinto, Shiwani Sharma, Stephen J. Nicholls, Christina A. Bursill, Peter J. Psaltis
The conference will for the first time since COVID-19 bring together the research community using avian models in research. We have an exciting line-up of top international speakers covering topics like stem cells, developmental biology, new technologies, immunology, comparative genomics and evolution, and the use of avian models for medical research. Besides the keynote speakers, the programme features short and flash talks selected from the abstracts, poster sessions, a plenary discussion on avian models in research, and a PI meeting to discuss the development of resources for the international avian research community.
Outside the conference Portsmouth and the South coast of England provide a range of attractive sights to visit. Portsmouth can easily be reached by train and from London Heathrow and Gatwick airports and Southampton airport. It also has ferry links with the Channel Islands, France and Spain.
We are very much looking forward to welcoming the avian model research community in Portsmouth. Please spread the word!
Excellent Unit of Wildlife Stem Cells (eWIS) is housed in two remarkable locations: Wildlife Reproductive Innovation Center (WRIC) atKhao Kheow Open Zoo (as part of Zoological Part Organization of Thailand) and Department of Biology, Faculty of Science, Burapha University. Both reside in Chonburi Province, Thailand.
Research summary
The eWIS initiative brings together a team of stem cell scientists and veterinarians, forging connections between universities and zoos. The primary objective is to explore the immense potential of stem cell technology in establishing a biobank comprised of stem cells derived from endangered mammalian and avian species. Our research focuses on species that require urgent preservation efforts to safeguard genetic diversity through the collection of viable cells capable of generating entire animal bodies, known as induced pluripotent stem cells (iPSCs). Additionally, we are striving to generate wild felid/ domestic and non-domestic cat embryonic stem cells.
Our primary focus lies in wild felid species, including fishing cat (Prionailurus viverrinus) and clouded leopard (Neofelis nebulosa), and domestic cat (Felis catus) as a model. Our goal is to generate iPSCs from these wild cats, unravelling the possibilities of utilizing iPSCs for conservation purposes through in vitro gametogenesis, animal cloning, and assisted reproductive technology (ART). We are also devoted to establishing wild avian iPSCs, with specific attention given to hornbills and peafowl.
Furthermore, we employ a range of evolutionary tools to delve into the conservation and divergence of pluripotency networks across various vertebrate species. In particular, our research involves studying Oct4 homologues to comprehend the effects of evolutionary changes on conserved and diverged Oct4 functions. The profound insights gained from understanding the Oct4 network and early developmental programs contribute to translational approaches, aimed at overcoming reprogramming barriers in wild species.
The eWIS research team primarily works within one of the largest zoos in the world, “Khao Kheow Open Zoo,” and Burapha University in Chonburi, Thailand. From left to right: Pawares, Chichakon, Pitiwut, Woranop, Ampika, Rattanaporn, Santhita, Nattakorn. (Photo credit : Nattawut Dueanjam).
Lab roll call
Santhita: Researcher – Managing routine lab works/paper works at WRIC, bridging zoo units to collect wild animal tissues (mostly dead) to eWIS; Organizing biobank at the zoo
Pathira : Lab Manager/Researcher – Managing routine lab works/paper works/ordering at eWIS lab at Department of Biology; Generating barnacle collection
Pawares: Pre-doc student – searching cat ovaries and testes around the city!, IVF of cat embryos, cat pluripotency explorer
Nattakorn: Master student – primary culture of tissues from live and dead mammals and birds; RNA-based reprogramming; bioinformatics on pluripotency network
Ratchapon: Master student – Wild avian/chicken reprogramming and understanding the network within avian pluripotency
Pitiwut & Rattanaporn & Phanpimon: BSc students – characterization of felid pluripotency and cat IVF/IVM
Chichakon & Phornchanok: BSc students – avian reprogramming: testing various sets of reprogramming factors
Ampika: PI – advice the team, developing ART research, searching research grant, building capacity of the team and strengthen research network
Woranop: PI – teaching, admin jobs, pipetting, being Head (meeting job) & Biology program director, making connections between organizations
Favourite technique, and why
Woranop: Cellular reprogramming is my favourite tool. It is fascinating of how these felid cells can uptake reprogramming factors and transform the morphology and reverse cell fate! I love immunostaining of pre-implantation embryos from IVF- this is stunning how a single cell can become diversified in forms and functions! Recently, I discover myself to enjoy working on structural models predicted from AlphaFold2 which helps understand the evolution of proteins orchestrating the development.
Ampika: ARTs including AI, IVF and cloning are my favourite techniques for enhancing breeding conservation program of important species such as fishing cat, clouded leopard, flat-headed cat (Prionailurus planiceps), Asian golden cat (Catopuma temminckii), Eld’s deer (Rucervus eldii) and Asian elephant (Elephas maximus). Developing cryopreservation for establishment of “Viable Cell Bank” is crucial as it is important as insurance against loss or extinct of endangered species. These techniques are valuable tools that we use to sustain the genetic diversity of our endemic and global important species.
Fishing cat, one of the wild felid species, is currently in a vulnerable status due to wetland destruction. The Zoological Park Organization of Thailand (ZPOT) prioritizes research efforts with universities to save fishing cats via assisted reproductive technology, particularly artificial insemination. We are utilizing stem cell technology to establish a biobank and perform cellular reprogramming in the Excellent Unit of Wildlife Stem Cells (eWIS). (Photo credit: Piyapong Chinnadate)
Apart from your own research, what are you most excited about in developmental and stem cell biology?
Woranop: It is amazing how cells know how to reorganize themselves into specific shapes and transform into particular lineages. The technology to generate synthetic embryos, based on our understanding of cell potency and differentiation, is incredibly exciting. I am curious to see how this technology will merge with our understanding of wild stem cells and eventually contribute to saving wild animals.
Ampika: It is very exciting to see how we can develop stable stem cell colonies and differentiate them into various cell types that can be applied for medical treatments and reproductive purposes. This capability is incredibly useful for sustaining wildlife populations in the future!
How do you approach managing your group and all the different tasks required in your job?
Woranop: Everything is all about team work and communication! Regular meet-up helps a lot. We also frequently hang out for lunch or dinner, creating a comfortable environment for students to freely talk and share ideas.
Ampika: Update and discuss in anytime and set up a small meeting at least once a month.
What is the best thing about where you work?
Woranop : I enjoy working multidisciplinary fields of biology at our department and visiting the zoo every time I go to the eWIS lab. It allows me to appreciate animals and nature.
Ampika : The best part of our work is that we apply all our knowledge to prevent our wildlife from facing the brink of extinction! Our work supports not only ex situ management but also in situ conservation.
Santhita: At the zoo, we have access to a diverse range of wild animal species, which enables us to quickly collect samples for our work after the animals die or during health checks.
Pathira: The department admin staffs – we can ask for almost everything from lab materials to animal/plant specimens.
Pawares: Working with developing embryos often fills my heart with boundless excitement.
Nattakorn: I love being a TA and teaching undergraduate students! It provides me with opportunities to help other young students develop a passion for science through activities like science camps. It also allows me to support them in navigating university courses and life.
Ratchapon: The teachers in our department are very friendly, and we can contact them to get advice anytime.
Pitiwut: Everyone here is a bundle of joy- we can engage in conversations about anything, no matter how unrelated to work it may be!
Rattanaporn: Learning new things all the time
Phanpimon: Our advisors are the best! They always provide great support.
Chichakon:There are plenty of opportunities to engage in biology research starting from my third year! I love the open exchange of ideas we have in the lab.
Phornchanok: The great friendships in the lab make everything easier.
Purachet: Enjoy Sukiyaki parties with admin staffs in the department lunch breaks!
eWIS research team
What is there to do outside of the lab?
Woranop : I teach undergraduate and graduate students, and it brings me joy to share the fascinating stories of developmental biology with them outside the lab. We are also located near Bangsaen Beach! I often hang out with our team and visit other beaches along the eastern coast of Thailand.
Ampika : Since we work with 6 zoos nationwide and the Elephant Kingdom Project, it is very enjoyable to explore the places and talk to local people. And of course, Bangsaen is my favourite place to R&R!
Santhita: Sometimes in the evening, we take walks around the zoo to be close to nature. Nearby, there are tourist attractions such as Chan Ta Then waterfall and Bang Pra reservoir.
Pathira: Hangout at the zoo, Capybara watching! & definitely, Bangsaen is a right place for seafood, in particular grilled shrimp/squid
Pawares: I help organize department events and take care of our small colonies of chickens!
Nattakorn: I enjoy playing online games while waiting for my phylogenetic analyses! The beautiful Bangsaen beach also provides great help during exam or lab meeting crises
Ratchapon: I assist the department’s administrative staffs for specific events and serve as a TA to help Woranop teach the Developmental Biology Lab.
Pitiwut:I keep myself fit and healthy by doing cardio exercises at the nearby gym! I think it helps clear my thoughts a lot after a long day of studying and working in the lab.
Rattanaporn: Moo-gata (Traditional Thai BBQ!) is a must here.
Phanpimon: Jet skiing, running on the beach, or even feeding the monkeys on Khao Sam Muk hill.
Chichakon: I love designing art works for my lab and also others in the department.
Phornchanok: We can do a lot of things in Bangsaen – shopping, visiting cafes, having dinner, and enjoying the beach!
Purachet: Outside of class and the lab, I spend most of my time in the department studio, where I assist with photography and collect biological samples for departmental teaching.
Browse through other ‘Lab meeting’ posts featuring developmental and stem cell biology labs around the world.

 (No Ratings Yet)
(No Ratings Yet)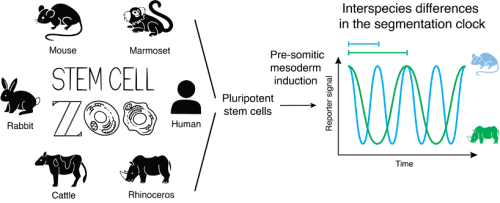
 (3 votes)
(3 votes)