Epithelial to mesenchymal transition (EMT) is an essential process in multiple steps of embryogenic morphogenesis and various pathological conditions. As an example, EMT is involved in gastrulation and neural crest cell (NCC) development during embryogenesis. EMT is also crucial for wound healing, tissue fibrosis and cancer progression. In addition to the cellular and molecular changes facilitating the transformation from epithelial cells into mesenchymal cells, EMT has also been associated with stemness, therapeutic resistance and tumor heterogeneity specifically in the context of malignancy. Due to the inherent differences between different species, cell types and biological contexts, there are variations within phenotypic changes and the underlying molecular mechanisms of different EMT programs.
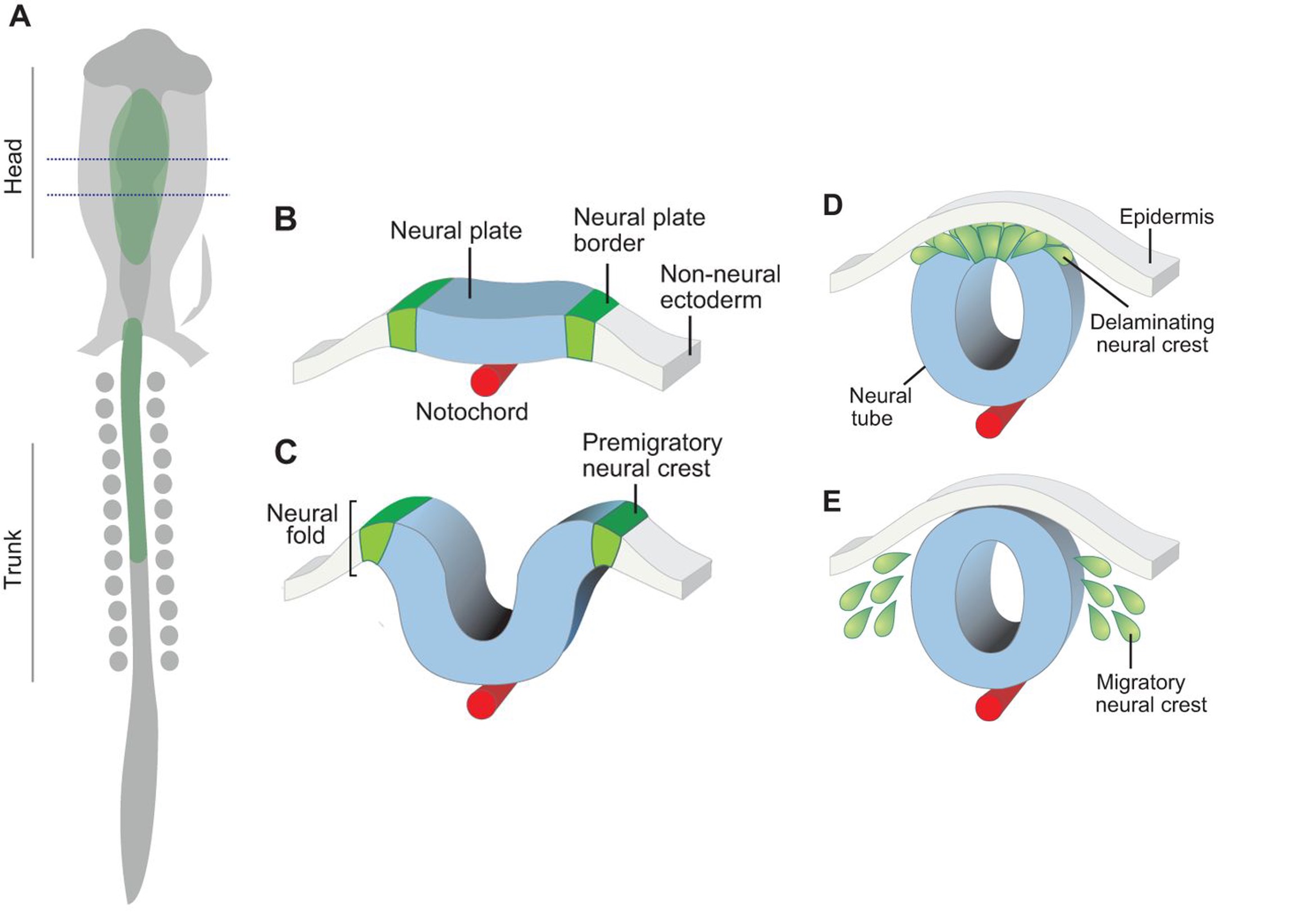
NCC are an embryonic progenitor cell population that gives rise to numerous cell types and tissues, such as craniofacial bone and cartilage, in vertebrates. Pre-migratory NCCs delaminate from the neuroepithelium via EMT, following which NCCs migrate throughout the embryo and undergo differentiation. Currently, we have limited understanding of the EMT process that gives rise to migratory NCCs in mammals because many NCC EMT related findings in non-mammalian species have not been successfully replicated in mammalian species.
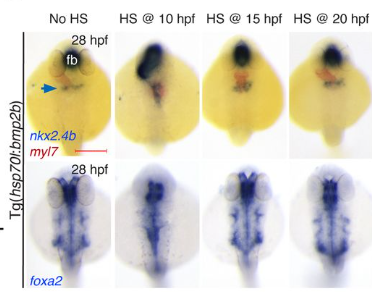
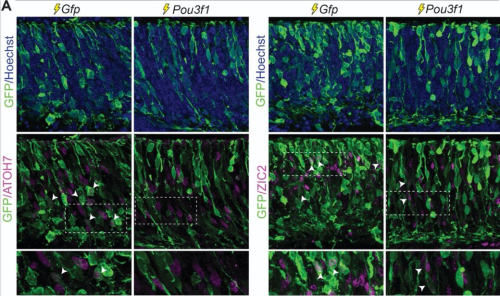
Transcriptional factor Twist1 is one of the major master regulators shown to be involved and to play an important role in EMT throughout both development and carcinoma progression. Previous studies on the role of Twist1 during mammalian NCC development using various mouse models have been thoroughly summarized and reviewed (Zhao and Trainor, 2022). In short, Twist1 null mice exhibit embryonic lethality around E11.5 associated with craniofacial defects such as malformed branchial arches and facial primordia (Chen et al.,2007; Chen and Behringer, 1995). Upon careful experimental testing, these phenotypes were believed to be caused by abnormality of NCC migration and differentiation. Consistent with these in vivo findings, mutations in Twist1 in humans lead to Saethre-Chotzen syndrome, which is characterized by craniosynostosis and cleft palate. In a recent publication “Twist1 Interacts with Beta/Delta-Catenins During Neural Tube Development and Regulates Fate Transition in Cranial Neural Crest Cells”, however, Bertol and her colleagues further depict the neuroectodermal expression profile of Twist1 during early mouse embryogenesis and illustrate potential functions of Twist1 in mouse cranial NCC delamination and EMT.
Key findings
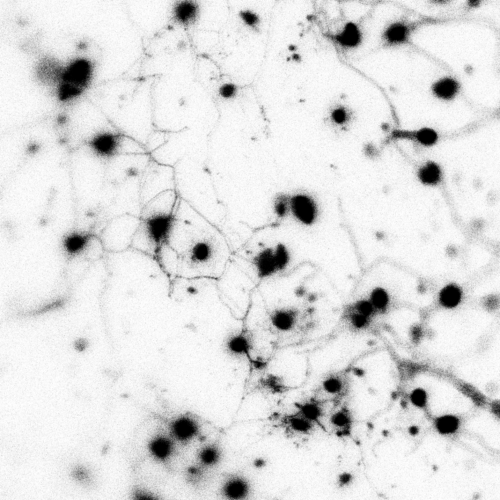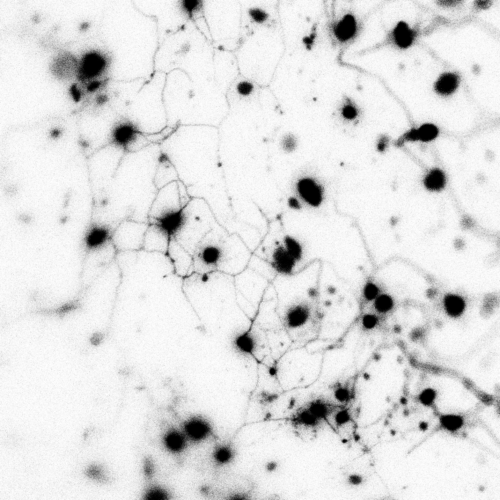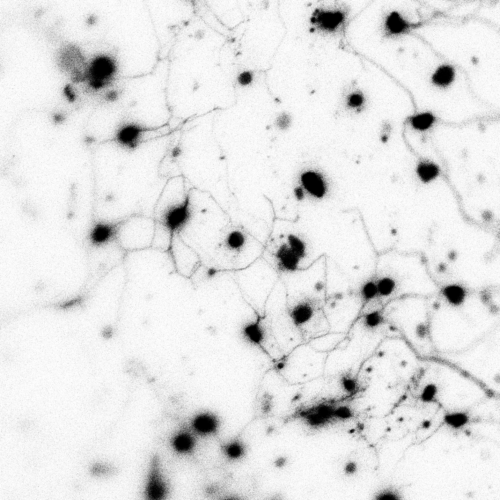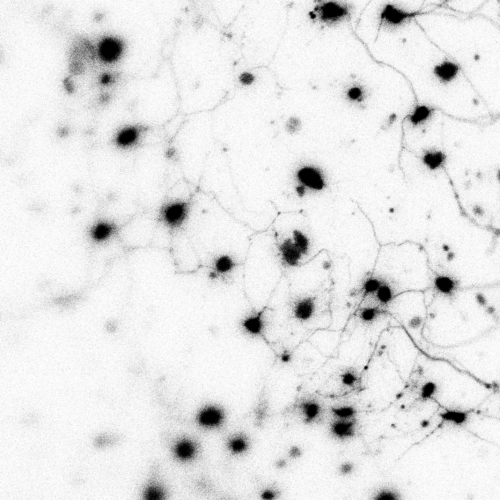
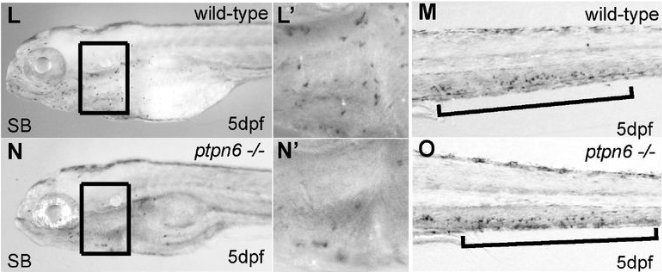
During mouse embryogenesis between E8.5 and E9.5, Twist1 is detected in vesicle-like structures on the apical side of the neuroepithelium/neural plate. Interestingly, such apical expression of Twist1 coincides with the expression pattern of B-catenin and Claudin-1 suggesting an association of Twist1 with adherens and tight junctions in the neuroepithelium. Furthermore, a physical interaction between cytosolic Twist1 and B-catenin is demonstrated by co-immunoprecipitation. When Twist1 is deleted in whole embryos, apical B-catenin in vesicle like structures become diffused and mostly cytosolic in the apical neuroepithelial cells of the neural plate.
Consistent with other studies, Twist1 is also found to be expressed in migratory cranial NCCs at E8.5, E9.5 and E10.5. When Twist1 is conditionally deleted in cranial premigratory NCCs at early E8.5, cranial migratory NCCs are observed throughout the embryos, but there is a fewer number of migratory NCCs in the frontonasal and pharyngeal processes between E9.5 and E11.5. This observation is later confirmed by severe frontonasal prominence defects and neural tube closure abnormalities.
Examinations of remaining post-delamination migratory NCCs in neural tube explants from Twist1 conditional knockout mice reveal that the majority of migratory NCCs exhibits epithelial morphologies, significant cell-cell adhesions and continuous junctional signals of ZO1. Moreover, migratory cranial NCCs in vivo show increased E-cadherin expression, and Specc1 (an actomyosin cytoskeleton regulator) expression is reduced in the hindbrain and first pharyngeal arch. These data indicate disrupted EMT during the delamination of cranial NCCs in an absence of Twist1 expression.
To study the importance of Twist1 phosphorylation in craniofacial tissue development, the researchers have also generated four Twist1 phospho-incompetent mouse lines. Phenotypic characterizations of these mutants demonstrate that S18/20 and S68 phosphorylation sites are critical for craniofacial development.
In summary, the paper contributes a valuable collection of data to fill our knowledge gap of how NCC delamination and EMT are regulated in mammalian species. To my knowledge, this is the first publication that directly studies the role of Twist1 specifically in early NCC development via using Wnt1-Cre and Wnt1-Cre2 driven conditional knockout mouse models. Although I find some parts of the paper slightly confusing regarding the interpretation of certain data and the relevance of IRF6 data to the rest of the paper, the data itself is still very intriguing and thought-provoking. Interestingly, Zeb2 null mutant mouse embryos exhibit similar phenotypes of persistent E-cadherin expression in migratory cranial NCCs (Putte et al., 2003). Similar to Twist1, neither Snail1 or Zeb2 conditional knockout in pre-migratory NCCs completely inhibits NCC delamination and EMT (Murray and Gridley, 2006; Rogers et al., 2013). These previous findings in combination with the proposed function of Twist1 in the completion of mouse cranial NCC EMT could suggest that perhaps EMT master regulators act synergistically in waves to promote the complete transition from neuroepithelial cells to mesenchymal migratory NCCs.
A morning excursion to the local tidepools – Instructor Nat Clarke (lower right) identifying species for students and guest Dan Rokhsar (photo credit: Azalea Martinez Jaimes).
In June 2022, I had the pleasure of teaching a short course on comparative embryology with Chris Lowe and Laurent Formery at Stanford University’s Hopkins Marine Station. Our mission: to take a mix of grad students and postdocs from disciplines across the biosciences, introduce them to diverse developmental mechanisms in a broad sampling of organisms spanning the animal tree of life, and then release them to pursue research projects of their own design.
What is the value of a course like this? And why are marine stations an ideal setting? Simply put, animal diversity showcases countless natural experiments – evolutionary experiments in diverse body plans, novel cell types, complex life history strategies, and more. Since life evolved in the ocean, the seaside environment allows unrivaled exploration of this biodiversity: where else offers ready access to embryos of 15 animal phyla within steps of the classroom?
I could, of course, describe at length what students might get out of such an experience, but instead I’ll give it to you directly from the source. Below, a few of our students, and my co-instructor, share their reflections and favorite moments from the course.
Student Perspectives:
Charlotte Brannon:
A colony of a red encrusting bryozoan growing on a shell (left; photo credit: Charlotte Brannon), with a close-up view of individuals enclosed in their ‘houses’ (center; photo credit: Lauren Lubeck). A confocal micrograph of individual zooids of another bryozoan species (right; photo credit: Joel Erberich)
There is so much biology hiding in non-model organisms, especially in the ocean, and it was enlightening to immerse myself in it during this course. Partly, this happened through tidepooling and digging for worms in mud flats. Tidepooling was surprisingly challenging at first. Our instructors could pick up a rock, identify five species on it, and tell you anything you wanted to know about any of them. Meanwhile, the most exciting thing I could find was a floating, white creature which turned out to be… a seagull feather. I ended up collecting a seemingly boring shell with some red stuff encrusted on it. Surprisingly, a quick look under the dissection scope revealed this crusty red stuff to be a Bryozoan colony! Unknowingly, I had collected a fascinating marine invertebrate. Similarly, when we later visited mud flats, a classmate and I found what seemed to us like a very average worm. We later learned that it was actually Leptosynapta albicans – the burrowing sea cucumber. I learned my lesson: when immersed in nature, you must actively try not to find something interesting.
Student Lauren Lubeck trying her hand at microinjection (left). At right, a single blastomere injection of a two-cell nudibranch embryo of the species Berghia stephanieae (photo credit: Nat Clarke)
Exposure to a wide range of marine invertebrates made me more excited about developmental biology because it highlighted how much we don’t know! I am excited about all that non-model organisms can teach us, and eager to explore the mechanics of development in a range of systems. Of course, this is easier said than done. As we learned, having the right tools to work with an organism is crucial, and sometimes that means building your own. The short timeline of the course forced us to be creative with our resources and think carefully about the appropriate tools. I also appreciated seeing multiple approaches to the same experiment. For example, our guest instructor, Brady Weissbourd, demonstrated his strategy for injecting Clytia eggs, which differed in many subtle ways from our instructors’ approach for injecting echinoderm eggs. This made me realize how important it is to tailor your experimental approaches to your organism.
Lauren Lubeck:
As a marine research station, Hopkins Marine Station is one of only a few special places where scientists can collect, observe, and perform experiments on a wide diversity of marine invertebrates. It was an incredible experience to be immersed in the marine environment at Hopkins. Unsurprisingly, almost every guest instructor wanted to spend time outside looking for their favorite organisms, and the students went tidepooling on our own many times. We learned the importance of understanding the ecology of our target species. Want choanoflagellates? Look for small, dirty looking pools. Want acoels? Turn over rocks in the sandy sections of tidepools. Want fat innkeeper worms? Look for the holes marking the entrance to their U-shaped burrows. Working where our favorite intertidal invertebrates live created a unique opportunity to learn more about them.
Circulating blood in the tunicate, Botryllus (A. Jaimes)
Nerve net of a transgenic jellyfish (N. Martinez)
A highlight for me was the connections I made with fellow students. Each student arrived with specific interests and biological questions in mind. I loved that we all found ways to investigate our favorite questions while using species that were new to us. While each of us pursued our original question, we also were spontaneously inspired by a new animal or phenomenon we encountered. For example, Nabor Vazquez Martinez usually studies aspects of the nervous system in C. elegans, but he found Brady Weissbourd’s Clytia jellyfish fascinating and decided to examine their nervous system too. Azalia Martinez Jaimes is interested in stem cell differentiation and found an interesting model in the tunicate Botryllus, which continuously builds new adults from growing buds filled with stem cells.
A child observing Chrysaora sea nettles in the jellies exhibit at the Monterey Bay Aquarium (photo credit: Nat Clarke)
The “Behind the Pipes” tour of the Monterey Bay Aquarium was another highlight. As we were led through the ctenophore facility by Senior Aquarist Wyatt Patry, I learned that they grow the same algae we were feeding our larvae in class, but I was amazed by the massive scale of production. Once we entered the main exhibit to see the ctenophores and cnidarians on display, I was overcome with a combination of awe and excitement. It was moving to see how our research interests meshed with the education and outreach of the Monterey Bay Aquarium.
A. S. Jijumon:
“In all things of nature there is something of the marvelous” — This is a quote from Aristotle that I saw at the Monterey Bay Aquarium during our class visit. In this course, I realized that there is much to learn and explore in marine organisms, and that it can be more straightforward to make novel and important observations in unexplored areas.
Sea cucumber ossicles (photo credit: Charlotte Brannon). DIC image and a confocal micrograph of a sea cucumber larva, DAPI (blue), tubulin (green), phalloidin (red) (photo credit: AS Jijumon).
I did my undergraduate education in India, and what I studied in my developmental biology classes were mostly theories and text book images. Through this course, for the first time, I got a practical demonstration of the developmental stages of multiple marine invertebrates and observed their morphogenesis over time. I got a chance to really experience the origin of classical experiments in embryology and developmental biology. Observing the wide diversity of organisms we could collect straight from tidepools, and then performing wet lab experiments on these creatures using microscopy and molecular tools was a fantastic experience.
Image of a sand dollar (Dendraster excentricus) egg during fertilization (photo credit: AS Jijumon).
My favorite observation was with the sand dollars (Dendraster excentricus) we worked with on our first day. We collected eggs and sperm by injecting KCl into their gonads. Subsequently, I added diluted sperm onto an egg and watched on the microscope. That was the first time I witnessed the event of fertilization occur in real life, and I felt goosebumps and got the impression that I acquired the power to manipulate life, which was a memorable moment. I would like to share this experience with other science enthusiasts in the future.
Instructor perspective – Laurent Formery:
Confocal micrograph of a nudibranch, Corambe sp. DAPI (cyan), phalloidin (yellow) and WGA (pink). (Photo credit: Joel Erberich)
This course was one of my first teaching experiences, and it was an awesome one. We wanted to promote exploration and experimentation using the incredible resource that we had right outside the classroom – the ocean. From tidepools, mudflats and plankton nets we collected species spanning over a dozen animal phyla (and some of our closest unicellular relatives, too), and we spawned, observed and manipulated them in the classroom. The combinations of the students’ unique skill sets and the array of animals that we collected generated a profusion of discovery – applying well-developed techniques to new questions in new species. During this process I personally learned much more than what I could teach to the students, making this course an enriching experience for me as well. The main message of the course we tried to emphasize was the astonishing diversity of biochemical processes, developmental mechanisms, and ecological strategies waiting to be discovered and documented right there in the ocean. This underscores the importance of protecting the endangered biodiversity of our coastlines, but also the value of supporting basic exploratory research outside the handful of classical biological model systems. One of my favorite examples highlighting the importance of exploring non-model systems was brilliantly told by Dan Rokhsar during his genomics lecture to the class. The recent chromosome mapping of non-model species such as the scallop Patinopecten yessoensis and the jellyfish Rhopilema esculentum enabled the discovery of a fundamental feature of animal evolution: the arrangement of genes on chromosomes (referred to as synteny) is highly conserved and can be traced back to the roots of metazoans (Simakov et al., 2022). The few exceptions to that rule, in which macrosynteny has undergone independent and major reorganization events, are curiously distributed among the metazoan tree: they include the fruit fly Drosophila melanogaster, the nematode Caenorhabditis elegans, and the entire vertebrate clade.
Acknowledgements: A course like this simply cannot run without the support of a team of deeply invested staff and instructors. We want to give a special “thank you!,” to the staff at Hopkins, and to all of our guest instructors – Dan Rokhsar, David Booth, Flora Rutaganira, Christina Zakas, Bo Wang, Brady Weissbourd, Ryan York, Wyatt Patry, Deidre Lyons, Jessica Goodheart, Dominique Bergmann – for making this year’s course a success. We also thank the departments of Biology, Developmental Biology, and BioEngineering, at Stanford for financial support, and Molecular Instruments and Luxendo for generously providing equipment and reagents for the course.
Confocal micrograph of a market squid, Loligo opalescens, stained with DAPI (grey) and HCR probes for sodium channel (yellow) and gluatamate receptor (magenta). (Photo credit: Nat Clarke)
CITED2 IS A CONSERVED REGULATOR OF DEEP HEMOCHORIAL PLACENTATION Marija Kuna, Pramod Dhakal, Khursheed Iqbal, Esteban M. Dominguez, Lindsey N. Kent, Masanaga Muto, Ayelen Moreno-Irusta, Keisuke Kozai, Kaela M. Varberg, Hiroaki Okae, Takahiro Arima, Henry M. Sucov, Michael J. Soares
AKT1-FOXO4 AXIS REGULATES HEMOCHORIAL PLACENTATION Keisuke Kozai, Ayelen Moreno-Irusta, Khursheed Iqbal, Mae-Lan Winchester, Regan L. Scott, Mikaela E. Simon, Masanaga Muto, Marc R. Parrish, Michael J. Soares
Visuomotor anomalies in achiasmatic mice expressing a transfer-defective Vax1 mutant Kwang Wook Min, Namsuk Kim, Jae Hoon Lee, Younghoon Sung, Museong Kim, Eun Jung Lee, Jong-Myeong Kim, Jae-Hyun Kim, Jaeyoung Lee, Wonjin Cho, Jee Myung Yang, Nury Kim, Jaehoon Kim, C. Justin Lee, Young-Gyun Park, Seung-Hee Lee, Han-Woong Lee, Jin Woo Kim
A Development-Inspired Niche for Homeostatic Human Mini-Intestines Charlie J. Childs, Emily M. Holloway, Caden W. Sweet, Yu-Hwai Tsai, Angeline Wu, Joshua H. Wu, Oscar Pellón Cardenas, Meghan M. Capeling, Madeline Eiken, Rachel Zwick, Brisa Palikuqi, Coralie Trentesaux, Charles Zhang, Ian Glass, Claudia Loebel, Qianhui Yu, J. Gray Camp, Jonathan Z. Sexton, Ophir Klein, Michael P. Verzi, Jason R. Spence
Reconstituting human somitogenesis in vitro Yoshihiro Yamanaka, Kumiko Yoshioka-Kobayashi, Sofiane Hamidi, Sirajam Munira, Kazunori Sunadome, Yi Zhang, Yuzuru Kurokawa, Ai Mieda, Jamie L. Thompson, Janet Kerwin, Steven Lisgo, Takuya Yamamoto, Naomi Moris, Alfonso Martinez-Arias, Taro Tsujimura, Cantas Alev
Reconstructing human Brown Fat developmental trajectory in vitro Jyoti Rao, Jerome Chal, Fabio Marchianò, Chih-Hao Wang, Ziad Al Tanoury, Svetlana Gapon, Yannis Djeffal, Alicia Mayeuf-Louchart, Ian Glass, Elizabeth M. Sefton, Bianca Habermann, Gabrielle Kardon, Fiona M. Watt, Yu-Hua Tseng, Olivier Pourquié
Generation of functional hepatocytes by forward programming with nuclear receptors Rute A. Tomaz, Ekaterini D. Zacharis, Fabian Bachinger, Annabelle Wurmser, Daniel Yamamoto, Sandra Petrus-Reurer, Carola M. Morell, Dominika Dziedzicka, Brandon T. Wesley, Imbisaat Geti, Charis-Patricia Segeritz, Miguel Cardoso de Brito, Mariya Chhatriwala, Daniel Ortmann, Kourosh Saeb-Parsy, Ludovic Vallier
New Hydra genomes reveal conserved principles of hydrozoan transcriptional regulation Jack F. Cazet, Stefan Siebert, Hannah Morris Little, Philip Bertemes, Abby S. Primack, Peter Ladurner, Matthias Achrainer, Mark T. Fredriksen, R. Travis Moreland, Sumeeta Singh, Suiyuan Zhang, Tyra G. Wolfsberg, Christine E. Schnitzler, Andreas D. Baxevanis, Oleg Simakov, Bert Hobmayer, Celina E. Juliano
Ten Simple Rules for Using Public Data for Your Research Vishal Oza , Jordan Whitlock , Elizabeth Wilk , Angelina Uno-Antonison , Brandon Wilk , Manavalan Gajapathy , Timothy Howton , Austyn Trull , Lara Ianov , Elizabeth Worthey , Brittany Lasseigne
8 July 2022 sees the full launch of Microscopya, a video game that takes its player inside the wonderfully complex world of the cell. One of the creators of Microscopya is Beata Mierzwa, who we heard from in one of the earliest SciArt features on the Node. You can read more about the game, its aims and its creators in the press release posted on FocalPlane.
Having tried the game with my family, I can confirm that it appeals to all ages with its beautiful visuals and soundtrack. Whilst younger children might not pick up on all the details (we had a dinner table conversation about a ‘metrotron’ being the powerhouse of the cell!), they loved the microtubule roads and playing the games to win trophies.
You can find out more about Microscopya in the following places:
The overall aim of this call is to support proposals that:
Develop the next generation of non-animal technologies that mimic the physiological environment enabling a whole system/ multi-system approach for discovery and translational science.
Enhance the capacity and confidence in non-animal technologies.
Establish partnerships between academia, the SME sector, and industry.
Proposals must fall within the BBSRC’s remit to qualify for funding and the research supported should have the realistic possibility of replacing the use of specific in vivo models or animal studies in line with the NC3Rs mission. Projects can run for up to 24 months and should begin by January 2023. Awards are for up to £250k full economic cost and the BBSRC and NC3Rs will fund 80% of the full economic cost. The competition will close on 8 September.
If you are interested in applying for the call, don’t miss the applicant webinar on Thursday 14 July at 10am (BST), where you will have the chance to ask any questions you may have about this new funding opportunity.
After attending several virtual conferences over the past two years, EMBO/EMBL symposium on Mechanobiology in development and disease, held at EMBL Heidelberg between the 15th to 18th of May 2022, was my first in-person meeting since Covid-19 hit. I was excited and equally nervous because my abstract had been selected for an oral presentation at this esteemed platform. This meeting was held in a hybrid format involving participants on-site as well as virtually.
Invited talks were mixed with selected short talks, followed by a “Meeting with speakers” at the end of each session. Also, days 2 and 3 culminated with flash talks by enthusiastic poster presenters, which were refreshing. Some of the flash talk presenters used innovative and hilarious ways to attract the audience to their posters, thus keeping us interested and curious even towards the end of a very busy day. Researchers, enjoying their drinks, congregated around the posters, which were displayed along the distinctive DNA double-helix-shaped staircases of the EMBL building. The virtual attendees also participated with their queries after each presentation via a chairperson for the session. A few highlights from the meeting which captivated me the most were as follows:
Tissue and organ morphogenesis: Day 1 of the meeting started with a wonderful keynote presentation by Ed Munro (University of Chicago, USA) on tissue morphogenesis in an Ascidian, Ciona robusta. He showed mesmerizing movies of neural tube zippering caused by concurrent events of tissue contraction and relaxation at the anterior and posterior end, respectively. Jean-Leon Maitre (Institut Curie, France) showed how hydrostatic pressure fractures the adhesion between cell contacts forming microlumens that coarse with time due to the contractile cell surface. These microlumens eventually fuse to form a single large fluid-filled cavity known as blastocoel in an early-stage mammalian embryo. Amy Shyer (Rockefeller University, USA) discussed the self-organization of tissue patterns in dermis explants which results due to cell-intrinsic contractility and the mechanical properties of the underlying extracellular matrix.
Nuclear mechanotransduction: The meeting revealed multiple exciting discoveries related to nuclear mechanotransduction i.e. how the “brain of the cell” i.e. the nucleus responds to different mechanical environments. The extracellular matrix transduces force to the nucleus through focal adhesions (connecting ECM to the cell surface) and the cytoskeleton (connecting focal adhesions to the nuclear envelop). Pere Roca-Cusachs (Institute for Bioengineering of Catalonia and Universitat de Barcelona, Spain) demonstrated how the stiffness of ECM modulates nuclear shape and pore size thereby affecting the nuclear import/export of a transcriptional regulator known as YAP. Windie Hoefs (Francis Crick Institute) shared her work on the opposing roles of cell-cell adhesion and cell-matrix adhesion in regulating the nuclear localization of YAP when the cells are mechanically stretched. Alice Willart (Institut Curie, France) showed variations in nuclear volume under different magnitudes of mechanical confinement, while Adel Al Jord (Collège de France, France) discussed the role of cytoplasmic forces in organizing nuclear condensates, nucleoli, and mRNA splicing in mammalian and fly oocytes and showed their significance in maintaining organism fertility.
Tissue repair, maintenance, and disease: Genomic instabilities or aberrations can lead to a myriad of deleterious consequences such as cancer and apoptosis. Rong Li (Mechanobiology Institute, Singapore) showed that mitotic errors cause hypo-osmotic stress. Aneuploid cells i.e., cells with an abnormal number of chromosomes, have proteome imbalance which results in the influx of water due to an osmotic pressure difference between the cytoplasm and extracellular environment. Consequently, aneuploid cells become much larger and stiffer than normal cells. While it is well-established that the clearance of apoptotic cells is carried out by immune cells, the mechanism of their removal during early embryogenesis had remain unexplored. Verena Ruprecht’s (Centre for Genomic Regulation, Barcelona, Spain) research revealed an unexpected role of epithelial cells as “phagocytes” that scavenge apoptotic cells during vertebrate embryogenesis, using specialized structures: phagocytic cups and epithelial arms. Tissue homeostasis and development require clearance of dead or unwanted cells as well as repairing of injured tissues. Yalan Mao (University College London, UK) discussed the multiple ways tissues employ to heal wounds, such as tissue fluidity and an altered 3D cell shape. Jochen Guck (Max Planck Institute for the Science of Light, Germany) reported a high-throughput method for screening the mechanical properties of blood cells such as cell size and stiffness, using a microfluidic device, which could be used for the prognosis or the analysis of the phenotypic consequence of diseases such as Covid-19 and cancer.
Besides the intensive scientific talks and discussions, there was a relaxed and calming musical night entertaining all the participants.
I enjoyed all aspects of the meeting, including talks, coffee break discussions, poster sessions, and the food. I would like to thank all the organizers (Alba Diz-Muñoz (EMBL Heidelberg, Germany), Carl-Philipp Heisenberg (Institute of Science And Technology, Austria), Prisca Liberali (Friedrich Miescher Institute for Biomedical Research, Switzerland), Anđela Šarić (University College London, UK), Xavier Trepat (Institute for Bioengineering of Catalonia, Spain) for organizing an excellent meeting covering various aspects of mechanobiology in multiple contexts, from molecular to organism level. Also, I would like to acknowledge Nathalie, the coordination officer, who answered everyone’s queries very patiently before, during, and after the meeting, and made the entire conference run smoothly without any technical glitches.
Nathalie Sneider, Conference coordinator, standing in front of the conference hall
I look forward to more such meetings in the future and would like to recommend this meeting to anyone who would like to pursue their research or is currently working in the field of mechanobiology.
Staff Scientist, University of Chicago, United States.
Charles Darwin, Primo Levi, Rachel Carson, John MacPhee, Ed Yong, Carl Sagan, and Lewis Thomas have all written creatively and eloquently about the natural world and the practice of doing science. In so doing they have changed the way people think about our world, and about scientists.
This past June, The Company of Biologists ran a Workshop called “Creative Science Writing.” The group numbered a dozen mentors (published writers of scientific non-fiction, fiction, and poetry—some full-time writers, many others holding academic positions) and about fifteen mentees (lab techs, recent PhDs, post-docs, professors, staff scientists, freelance writers and journalists, and a playwright, among others).
After we’d found our rooms in the historic Wiston House (I was told mine were in a converted brewery), we gathered in the large conference room with its long, U-shaped table, carrying in name-cards to display in front of us. A microphone sat at each place, making it feel a bit like a small United Nations. We went round the U introducing ourselves by means of an object of special meaning we’d been asked to bring. Those who’d gotten the memo after already being en route to the Workshop admirably evoked such objects by words alone. Already we were telling stories, and it was notable the extent to which people shared quite personal ones right at the outset. This set the tone for the Workshop, which continued in that vein throughout.
Kat Arney, science writer and broadcaster, started off our discussion of Narrative with the observation, “Stuff has to have a beginning, a middle, and an end, and not everyone does that.” We then had a fair bit of discussion of peoples’ practices of outlining work in detail vs. seat-of-the-pants composition, primarily related to non-fiction books. Matthew Cobb, professor of Zoology and writer of non-fiction books, concluded, “No plan survives first engagement with the enemy.” Our resident playwright, Raegan Payne, describing what not to tell, offered the tag, “Come in late and leave early.”
During the Workshop, we broke into smaller groups to discuss pre-distributed published essays and writing submitted by mentees. We also had blocks of time to write or revise. Interspersed with this full schedule were delicious meals prepared by the world-class chefs of Wiston House, tea breaks, and evening drinks and conversation. We were free to wander the extensive and beautiful grounds of Wiston House and held many of our small-group discussions out of doors. The rotating sets of small groups made it easy to get to know one another quickly, with the scale of the meeting making it easy to interact with everyone in the end whilst promoting a very pleasant and collegial atmosphere.
We were lucky to get to talk by zoom to the writer of one of the published pieces, Karen Joy Fowler, and she gave us a lot of insight into her individual writing process. She ended with a cautionary note to the mentees, who were about to hear feedback on their own writing: Never change what you’ve written based on someone else’s critique, unless you fully believe it will get you closer to your intentions.
That being said, the feedback sessions on our submitted pieces were quite supportive as well as offering detailed suggestions and thoughtful critiques. The diversity of writing styles and structures was impressive, and we learned a good deal from one another. One of the most unexpected lessons I took away from the Workshop was that an essential ingredient for a piece that worked is emotional openness and honesty, and a great many pieces displayed that virtue.
Although the majority of attendees had come to work on their non-fiction writing skills, by the end of the Workshop many had become enthusiastic about incorporating more elements from storytelling and fiction into their narratives, or had even decided to try their hands at fiction or poetry.
One of the organizers, Enrico Coen, professor and writer, summed up the Workshop thusly: “My conclusion is that everyone here is slightly barmy.” He added that this will be a great comfort to him as he returns to his peers in the scientific world, knowing that he has all of us as a community. Many of the attendees shared this feeling of having gained a sense of kinship with this group of science writers of all spots and stripes.
For this month’s SciArt profile, we caught up with Bob Goldstein, a professor at UNC Chapel Hill, who uses various printmaking techniques to produce unique posters advertising scientists’ talks, as well as other art that is shown in galleries and held by permanent collections including New York’s Poster House museum.
Where are you originally from and what do you work on now?
I was born in suburban New York, and trained in Texas, England, and California before starting my job in North Carolina. I run a lab where people use C. elegans to discover fundamental mechanisms in cell and developmental biology, plus we’re developing tardigrades as an emerging model system to study a few different things.
Dan Rokhsar Gig poster
Were you always going to be a scientist?
No – I confess that I didn’t really know what scientists did until late in college, when I took a course that focused on experiments. Before then, I had viewed science cynically as just a giant mountain of facts. It was during the course that I realized that science involved creative people devising clever solutions to answer fascinating questions. I was an instant convert, although it took about two years of grad school before I developed the skills and discipline to make any experiments work. I considered a lot of other careers during those two years.
Gig poster – Elke Ober, MBL Woods Hole
And what about art – have you always enjoyed it?
I’ve long enjoyed going to see art and making things with my hands. Parenting meant making things often – both of my kids are very creative, and when they were young, they loved making things that we did not know how to make. We did that just about every weekend. As they grew up and developed other interests, I kept making things on weekends. It took me a while to call any of it art. Like lots of people who make art, I have constructive urges, and acting on them can feel cathartic to me, much as I expect it feels for people who act on destructive urges.
Photogravure diptych inspired by Rita Levi-Montalcini. Photo credit: Lindsay Metivier
What or who are your most important artistic influences?
When I began screen printing, artists who made gig posters for bands were influences. Now I co-teach an unusual course with an Art professor, Beth Grabowski, who literally wrote the book (or a book) on printmaking, together with Duke University Art professor Bill Fick. They’ve been strong influences, along with other local printmakers that form a friendly and interactive community. And these days I think a lot about the old Italian poster artist Leonetto Cappiello, printmaker Delita Martin, and lots of people who make subversive public art.
Lino cut print – Drosophila
How do you make your art?
I use mostly screen printing, which involves using a squeegee to push ink onto paper through complex stencils supported by a fabric mesh. And I use other printmaking techniques – woodcut, linoleum cut, photogravure, intaglio – by standard and experimental methods, along with a little drawing and painting.
Still from the movie ‘Emergency’
Does your art influence your science at all, or are they separate worlds?
More and more I see ways that art influences my science, mostly in how I look at microscopy images and videos – and what I notice in them. And the converse is frequent: Science often influences my art. For years I’ve made gig posters for scientists’ talks. About two dozen of them just appeared in scenes of a movie called Emergency. And I’ve used Python programming to develop new ways for me to convert photos and drawings into patterns of dots that I can screen print.
Working with students, screen printing art onto windows of UNC’s Genome Science Building
What are you thinking of working on next?
I keep a long list of ideas in the Notes app on my iPhone, which helps me compartmentalise: I make art on weekends, and I set aside art ideas during the week when I’m focused on science. Only the Notes app knows what’s up next!
Thanks to Bob and all the other SciArtists we have featured so far. You can find the full list here. We’re always on the lookout for new people to feature in this series – whatever kind of art you do, from sculpture to embroidery to music to drawing, if you want to share it with the community just email thenode@biologists.com (nominations are also welcome!)
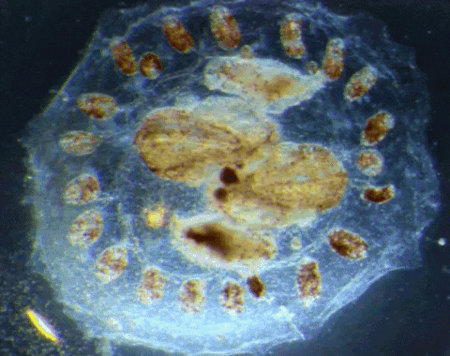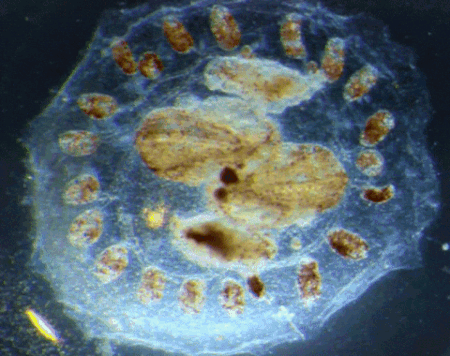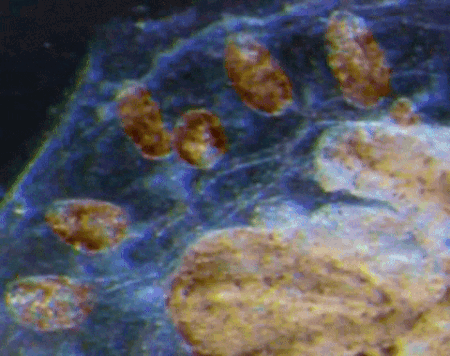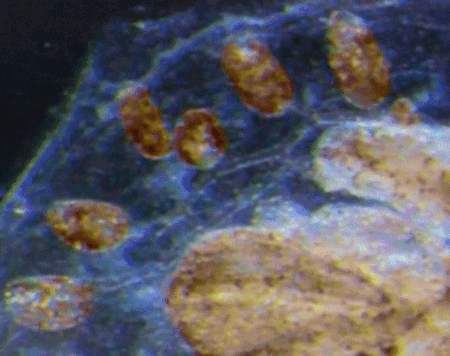
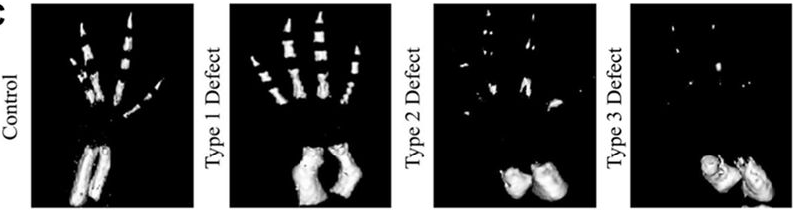
“Pay more attention to your hands, because not only can they perform microscopic miracles, they can also tell you who you were before you were even born”
Dr Sally Le Page
In the latest episode of the Genetics Unzipped podcast, we’re looking at the stories at your fingertips. Presenter, Dr Sally Le Page, uncovers how excrement espionage could bring down a superpower, and unearths a 100 year old family secret. But it’s not just genetic fingerprinting we’re interested in. We also grasp the genetics of fingerprints, and what they tell us about our early life in the womb.
If you enjoy the show, please do rate and review on Apple podcasts and help to spread the word on social media. And you can always send feedback and suggestions for future episodes and guests to podcast@geneticsunzipped.com Follow us on Twitter – @geneticsunzip
On June 8th 2022, a paper titled “Trunk Neural Crest Migratory Position and Asymmetric Division Predict Terminal Differentiation” was published on Frontiers in Cell and Developmental Biology. This is the second paper published by this research group on the topic of zebrafish trunk neural crest cell (NCC) migration in the past three months. Prior knowledge of their previous publication (Alhashem et al., 2022) is not required to understand or interpret this paper; however, I would like to prepare you with some general knowledge regarding the topic of neural crest migration/differentiation studied in this paper and mention a few key relevant findings from the previous paper.
NCC are an embryonic progenitor cell population that gives rise to numerous cell types and tissues in vertebrates. Pre-migratory NCC delaminate from the neural tube via epithelial to mesenchymal transition, following which NCC migrate throughout the embryo and undergo differentiation. NCC in the embryo trunk regions give rise to melanocytes, neurons and glia including Schwann cells and chromaffin cells. Migratory NCC streams consist of leader cells and follower cells. Leader cells initiate movement and remain at the front of the migratory NCC population to provide signals for the directionality of the population. Follower cells track leader cells’ movement and maintain cell-cell interactions necessary for migration.
Figure 1. Schematic dorsal view of a chicken embryo and cross-sectional view of NCC delamination and migration. Adapted from Simões-Costa and Bronner, 2015.
In Alhashem et al., 2022, researchers show that
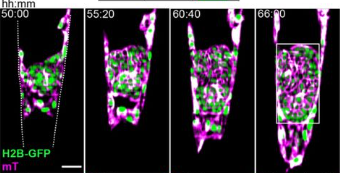
In each body segment, a single progenitor undergoes an asymmetric cell division to form a larger daughter cell, which later becomes a leader cell, and a smaller daughter cell, which migrates as a follower cell. The other progenitors undergo symmetric cell divisions to give rise to two follower cells.
Leader and follower cells progress asynchronously through cell cycle during early migration: leader cells divide during migration at neural tube/notochord boundaries, but follower cells divide prior to migration initiation in the most dorsal region of somites.
The total length of cell cycle is similar between leaders and followers during migration (no significant differences). However, leaders initiate migration during S phase, but followers initiate migration during G1 phase. Moreover, leader cells remain longer in S phase while followers spend longer periods of time in G1 phase in each cell cycle.
Cell cycle progression is required for trunk NCC migration in zebrafish and is regulated by Notch signaling.
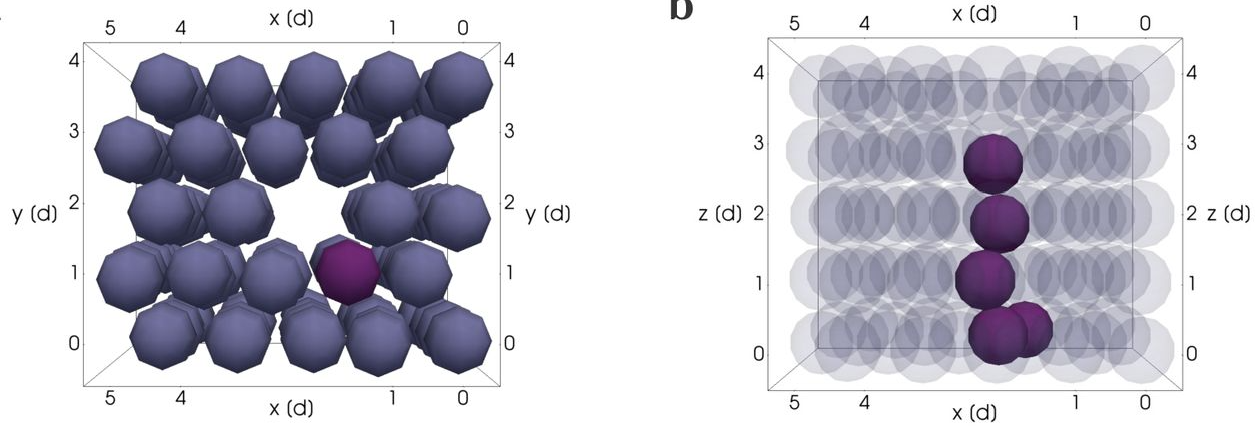
In the most recently published paper, researchers focus on later phases of trunk NCC migration when differentiation starts to occur. During migration, follower and leader cells maintain their relative positions within the migration chain. As a result, the position of migratory NCCs within a migratory chain correlates with their fates. When reaching the end of migration, leader cells divide perpendicular to the migration plane and give rise to a distal and proximal daughter cell. The distal cell remains at the most ventral position of the chain (lateral to the dorsal aorta) and differentiates to form the Sympathetic Chain Ganglia (SCG). The proximal cell stays dorsal to the SCG area and differentiates into Schwann cells. Interestingly, leader cell division is asymmetric so that the distal cell is bigger than the proximal cell sibling. On the other hand, followers divide parallel to the migration direction and their progenies populate the dorsal root ganglia (adjacent to the neural tube). In contrast to leaders, follower cell division is symmetrical generating two cells of similar sizes. Although it is beyond the scope of this study to address the mechanism regulating the pattern of cell division in leaders versus followers, it is discussed that an asymmetric distribution of Notch signaling in daughter cells after leader cell division could control identity allocation.
Another interesting observation demonstrated in the paper is that leader cells already express phox2bb, a noradrenergic marker, during migration (prior to reaching the destination for neuronal differentiation). This data suggests an early activation of transcriptional programs associated with lineage restriction in migratory NCC. How and when migratory NCCs become fate restricted has always been a major question in the field. The authors believe that this data supports the cyclical fate restriction model (Kelsh et al., 2021), which proposes that NCC can move between unstable sub-states biased towards specific differentiation outcomes. The transient differentiation biases can be determined by the expression of key fate determination receptors, the activity of the determination receptors, local environmental cues, and etc. Another recently published review paper also addresses and summarizes theories of NCC fate determination from existing single cell transcriptomics and multi-omics data (Erickson et al., 2022). It is worth a read if you would like to have a more up-to-date comprehensive view of this topic.
I personally enjoyed reading this paper because it demonstrates a strong link between two essential cellular activities: proliferation and differentiation. Cell division is so prominent during development that sometimes we don’t consider its possible interactions with other cellular processes. I also think most figures did a good job of illustrating the key points described in the texts. I especially like figure 4 and the 3D rendering that they did to show symmetric versus asymmetric division. It is a short and straightforward paper, but it leaves many intriguing topics to discuss and think about.
 (1 votes)
(1 votes)


 (No Ratings Yet)
(No Ratings Yet)