Read on for our news roundup of the past two weeks, with an emphasis on what has caught our eyes on twitter. We have also include a list of meetings with upcoming deadline, a selection of preLights and finish with our favourite April fools jokes .
Doing better as a community
There have been a few tweets in the last fortnight discussing some of the negative aspects of our research culture with a call to do better in the future. Lets make that happen!
Reviewing papers
Paper Reviewers: STOP asking for nickel and dime, incremental experiments that don’t advance the overall story. This kind of shit is strangling our trainees and accomplishes nothing but making you feel smart and slowing the pace of science. 1/2
Mixed emotions having been back from @Official_BSCB@_BSDB_#BSCBDB22 for 36h. Fantastic to be back in person, meet so many old friends and new faces; faces from screen in real life finally; some amazing and exciting science. And proud of how well the little one did. BUT…
The impact of Brexit on British science continues with notification from the ERC that awardees will need to move to the EU to take up their grants. The UK government is guaranteeing the grants in the UK but details remain sketchy.
And here it is — communication from @ERC_Research to UK-based #ERC starting grant grantees… UK awardees will have two months time to move their grant to the EU proper, otherwise they will lose their STG. Thank you #Brexit#BrexitReality. pic.twitter.com/AnJeCrCQrZ
— Thiemo Fetzer 🇪🇺🇺🇦 – same handle elsewhere (@fetzert) April 8, 2022
All the hard work put in to be awarded an #ERCStG, and we are now in the hands of a political decision. ☹️ https://t.co/YA1f4gezzX
Disaster for U.K. science. U.K. govt knew this was coming and let it happen. Where does the cash come from to replace this invaluable source? @ERC_Research is something we need to be part of. StG support the next generation…..Advanced funds half this old dawg’s group. https://t.co/skYlOv6Vkr
It’s been exciting to see the return of in-person meetings including the joint BSCB and BSDB meeting in the UK (we’ll post our meeting report soon) and #Dros22 in the US. Check out the list below for meetings with upcoming deadline and check our events page for our full listing.
If you are interested in science communication, would like to improve your writing skills and become part of the preLights community, don’t miss the open call for new preLighters.
Just for fun – our favourite April fools
We couldn't believe our eyes at first🤯🤯🤯! We just found the missing link between Cnidaria and Ctenophora. Introducing Pleurocoryna coelenterea, Pleurocorynidae fam. nov., Phylum: Coelenterata. pic.twitter.com/BSegc0B85e
After almost 5 years at the @czbiohub leading an awesome team of computationalists, microscopists and biologists, I have decided to move on and become a marine biologist. 1/n
Happy to announce that we are going to wind down our planarian research program in favor of studying brain regeneration in rodent models. We are all excited that we will finally be doing "real" neuroscience and working in a model relevant for human regeneration.
— Dr. Rachel Roberts-Galbraith (@awormwelcome) April 1, 2022
If you would like to write for the Node, check out our recent list of writing ideas. If you would like to contribute to our ‘Developing news’ blog, please get in touch at thenode@biologists.com
Join us in a Swedish enchanting scenario at the end of June (27-30) when it’s always light in Sweden!
If you are a Student (bachelor/Master’s/PhD), a Postdoc, or a young PI, and are using transcriptomics technologies, or you wish to approach them, this is the right event for you!
Plakoglobin is a mechanoresponsive regulator of naïve pluripotency Timo N. Kohler, Joachim De Jonghe, Anna L. Ellerman, Ayaka Yanagida, Michael Herger, Erin M. Slatery, Katrin Fischer, Carla Mulas, Alex Winkel, Connor Ross, Sophie Bergmann, Kristian Franze, Kevin Chalut, Jennifer Nichols, Thorsten E. Boroviak, Florian Hollfelder
Molecular Signatures and Cellular Diversity During Mouse Habenula Development Lieke L. van de Haar, Danai Riga, Juliska E. Boer, Youri Adolfs, Thomas E. Sieburgh, Roland E. van Dijk, Kyoko Watanabe, Nicky C.H. van Kronenburg, Mark H. Broekhoven, Danielle Posthuma, Frank J. Meye, Onur Basak, R. Jeroen Pasterkamp
TRIM28-dependent SUMOylation protects the adult ovary from activation of the testicular pathway Moïra Rossitto, Stephanie Déjardin, Chris M Rands, Stephanie Le Gras, Roberta Migale, Mahmoud-Reza Rafiee, Yasmine Neirijnck, Alain Pruvost, Anvi Laetitia Nguyen, Guillaume Bossis, Florence Cammas, Lionel Le Gallic, Dagmar Wilhelm, Robin Lovell-Badge, Brigitte Boizet-Bonhoure, Serge Nef, Francis Poulat
LINE-1 retrotransposon activation intrinsic to interneuron development Gabriela O. Bodea, Maria E. Ferreiro, Francisco J. Sanchez-Luque, Juan M. Botto, Jay Rasmussen, Muhammed A. Rahman, Laura R. Fenlon, Carolina Gubert, Patricia Gerdes, Liviu-Gabriel Bodea, Prabha Ajjikuttira, Peter Kozulin, Victor Billon, Santiago Morell, Marie-Jeanne H.C. Kempen, Chloe J. Love, Lucy M. Palmer, Adam D. Ewing, Dhanisha J. Jhaveri, Sandra R. Richardson, Anthony J. Hannan, Geoffrey J. Faulkner
Zfp503/Nlz2 is Required for RPE Differentiation and Optic Fissure Closure Elangovan Boobalan, Amy H. Thompson, Ramakrishna P. Alur, David McGaughey, Lijin Dong, Grace Shih, Emile R. Vieta-Ferrer, Ighovie F. Onojafe, Vijay K. Kalaskar, Gavin Arno, Andrew J. Lotery, Bin Guan, Chelsea Bender, Omar Memon, Lauren Brinster, Clement Soleilhavoup, Lia Panman, Tudor C. Badea, Andrea Minella, Antonio Jacobo Lopez, Sara Thomasy, Ala Moshiri, Genomics England Research Consortium, Delphine Blain, Robert B. Hufnagel, Tiziana Cogliati, Kapil Bharti, Brian P. Brooks
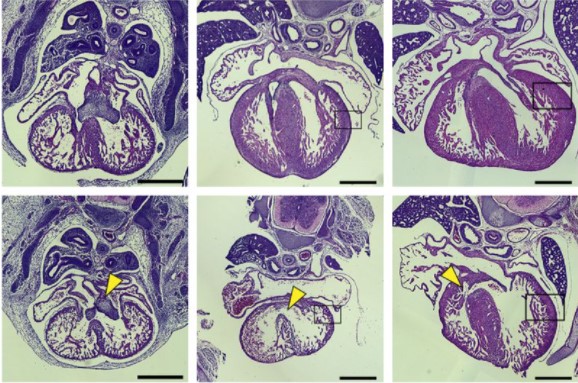
Embryonic mouse hearts stained with hematoxylin and eosin from Chang, et al.
A nuclear receptor facilitates differentiation of human PSCs into more mature hepatocytes Haiting Ma, Esmée de Zwaan, Yang Eric Guo, Paloma Cejas, Prathapan Thiru, Martijn van de Bunt, Jacob F. Jeppesen, Sudeepa Syamala, Alessandra Dall’Agnese, Brian J. Abraham, Dongdong Fu, Carrie Garrett-Engele, Tony Lee, Henry W Long, Linda G. Griffith, Richard A. Young, Rudolf Jaenisch
Stem cells partner with matrix remodeling cells during regeneration Blair W. Benham-Pyle, Frederick G. Mann Jr., Carolyn E. Brewster, Enya R. Dewars, Stephanie H. Nowotarski, Carlos Guerrero-Hernández, Seth Malloy, Kate E. Hall, Lucinda E. Maddera, Shiyuan Chen, Jason A. Morrison, Brian D. Slaughter, Anoja Perera, Alejandro Sánchez Alvarado
SARS-CoV-2 Can Infect Human Embryos Mauricio Montano, Andrea R. Victor, Darren K. Griffin, Tommy Duong, Nathalie Bolduc, Andrew Farmer, Vidur Garg, Anna-Katerina Hadjantonakis, Alison Coates, Frank L. Barnes, Christo G. Zouves, Warner C. Greene, Manuel Viotti
The conserved transcriptional program of metazoan male germ cells uncovers ancient origins of human infertility Rion Brattig Correia, Joana M. Almeida, Margot J. Wyrwoll, Irene Julca, Daniel Sobral, Chandra Shekhar Misra, Leonardo G. Guilgur, Hans-Christian Schuppe, Neide Silva, Pedro Prudêncio, Ana Nóvoa, Ana S. Leocádio, Joana Bom, Moisés Mallo, Sabine Kliesch, Marek Mutwil, Luis M. Rocha, Frank Tüttelmann, Jörg D. Becker, Paulo Navarro-Costa
Cell-intrinsic differences between human airway epithelial cells from children and adults Elizabeth F. Maughan, Robert E. Hynds, Adam Pennycuick, Ersilia Nigro, Kate H.C. Gowers, Celine Denais, Sandra Gómez-López, Kyren A. Lazarus, Jessica C. Orr, David R. Pearce, Sarah E. Clarke, Dani Do Hyang Lee, Maximillian N. J. Woodall, Tereza Masonou, Katie-Marie Case, Vitor H. Teixeira, Benjamin E. Hartley, Richard J. Hewitt, Chadwan Al Yaghchi, Gurpreet S. Sandhu, Martin A. Birchall, Christopher O’Callaghan, Claire M. Smith, Paolo De Coppi, Colin R. Butler, Sam M. Janes
Scalable Generation of Pseudo-Unipolar Sensory Neurons from Human Pluripotent Stem Cells Tao Deng, Carlos A. Tristan, Claire Weber, Pei-Hsuan Chu, Seungmi Ryu, Vukasin M. Jovanovic, Pinar Ormanoglu, Prisca Twumasi, Jaehoon Shim, Selwyn Jayakar, Han-Xiong Bear Zhang, Sooyeon Jo, Ty C. Voss, Anton Simeonov, Bruce P. Bean, Clifford J. Woolf, Ilyas Singeç
A cellular and molecular analysis of SoxB-driven neurogenesis in a cnidarian Eleni Chrysostomou, Hakima Flici, Sebastian G Gornik, Miguel Salinas-Saavedra, James M Gahan, Emma T McMahon, Kerry Thompson, Shirley Hanley, Michelle Kilcoyne, Christine E. Schnitzler, Paul Gonzalez, Andreas D Baxevanis, Uri Frank
Molecular characterization of a flatworm Girardia isolate from Guanajuato, Mexico Elizabeth M. Duncan, Stephanie H. Nowotarski, Carlos Guerrero-Hernández, Eric J. Ross, Julia A. D’Orazio, Clubes de Ciencia México Workshop for Developmental Biology, Sean McKinney, Mark C. McHargue, Longhua Guo, Melainia McClain, Alejandro Sánchez Alvarado
Single-cell atlas of human liver development reveals pathways directing hepatic cell fates Brandon T. Wesley, Alexander D. B. Ross, Daniele Muraro, Zhichao Miao, Sarah Saxton, Rute A. Tomaz, Carola M. Morell, Katherine Ridley, Ekaterini D. Zacharis, Sandra Petrus-Reurer, Judith Kraiczy, Krishnaa T. Mahbubani, Stephanie Brown, Jose Garcia-Bernardo, Clara Alsinet, Daniel Gaffney, Olivia C. Tysoe, Rachel A. Botting, Emily Stephenson, Dorin-Mirel Popescu, Sonya MacParland, Gary Bader, Ian D. McGilvray, Daniel Ortmann, Fotios Sampaziotis, Kourosh Saeb-Parsy, Muzlifah Haniffa, Kelly R. Stevens, Matthias Zilbauer, Sarah A. Teichmann, Ludovic Vallier
Multimodal spatiotemporal phenotyping of human organoid development Philipp Wahle, Giovanna Brancati, Christoph Harmel, Zhisong He, Gabriele Gut, Aline Santos, Qianhui Yu, Pascal Noser, Jonas Simon Fleck, Bruno Gjeta, Dinko Pavlinić, Simone Picelli, Maximilian Hess, Gregor Schmidt, Tom Lummen, Yanyan Hou, Patricia Galliker, Magdalena Renner, Lucas Pelkmans, Barbara Treutlein, J. Gray Camp
An integrated cell atlas of the human lung in health and disease L Sikkema, D Strobl, L Zappia, E Madissoon, NS Markov, L Zaragosi, M Ansari, M Arguel, L Apperloo, C Bécavin, M Berg, E Chichelnitskiy, M Chung, A Collin, ACA Gay, B Hooshiar Kashani, M Jain, T Kapellos, TM Kole, C Mayr, M von Papen, L Peter, C Ramírez-Suástegui, J Schniering, C Taylor, T Walzthoeni, C Xu, LT Bui, C de Donno, L Dony, M Guo, AJ Gutierrez, L Heumos, N Huang, I Ibarra, N Jackson, P Kadur Lakshminarasimha Murthy, M Lotfollahi, T Tabib, C Talavera-Lopez, K Travaglini, A Wilbrey-Clark, KB Worlock, M Yoshida, Lung Biological Network Consortium, T Desai, O Eickelberg, C Falk, N Kaminski, M Krasnow, R Lafyatis, M Nikolíc, J Powell, J Rajagopal, O Rozenblatt-Rosen, MA Seibold, D Sheppard, D Shepherd, SA Teichmann, A Tsankov, J Whitsett, Y Xu, NE Banovich, P Barbry, TE Duong, KB Meyer, JA Kropski, D Pe’er, HB Schiller, PR Tata, JL Schultze, AV Misharin, MC Nawijn, MD Luecken, F Theis
In the latest episode of the Genetics Unzipped podcast, we’re looking at the monkey in the mirror, investigating how flipped genetic switches and long-dead viruses make all the difference between our human faces and those of our closest primate relatives.
“The bits in our genome that encode functional elements like proteins are pretty much identical to a chimp’s. But straight away you can see that a chimp’s face is very different from our own, even though it’s made using the same biological ingredients.”
If you enjoy the show, please do rate and review on Apple podcasts and help to spread the word on social media. And you can always send feedback and suggestions for future episodes and guests to podcast@geneticsunzipped.com Follow us on Twitter – @geneticsunzip
December 3-7, 2022 | Walter E. Washington Convention Center | Washington, DC
Returning in person in 2022—this mid-sized unique joint meeting of the American Society for Cell Biology (ASCB) and the European Molecular Biology Organisation (EMBO) focuses on cell biology as the fundamental basis of biology, while also offering sessions on emerging interdisciplinary topics.
Besides offering attendees opportunities to present their research, the meeting provides an unparalleled forum for attendees to network and initiate collaborations across disciplines. Cell Bio 2022 offers eight different tracks—seven scientific and one covering educational, professional development, diversity, and inclusion.
Mansi Srivastava and I have edited a new book that’s packed with scientists’ stories about emerging model organisms. The book presents some interesting additions to the core set of model organisms, with contributions from people who have developed new model systems or advanced tools for emerging models. And it includes personal stories about how and why model systems were developed. The book isn’t comprehensive – we look forward to reading other scientists’ stories about these and other emerging models. But if you’re looking for 700+ pages of fascinating biology, we encourage you to take a look!
A PhD project proposal to be carried out in Patrick Lemaire’s lab at CRBM, Montpellier, France, in tight collaboration with Grégoire Malandain, Morpheme INRIA team, Sophia-Antipolis, France. Funding is conditional upon selection by the CBS2 doctoral school‘s entrance jury.
Why study ascidians?
The embryonic development of ascidians, a group of marine invertebrates, is remarkably conserved, at the single cell level, between individuals of a given species and between species, even if they diverged up to 400 MY ago. Ascidian genomes, however, evolve particularly rapidly. The remarkably simple and transparent ascidian embryos are thus ideal to study developmental systems drift and to identify constraints that could explain the exceptional evolutionary precision and stability of embryonic morphologies.
The proposed project
We propose a computational PhD project, which will use experimental data collected in the team or in public databases to characterise inter-individual and inter-species variability at the geometric, mechanical and transcriptional scales.
This project will involve the development of concepts and computer tools to study and measure, at each scale studied, the variability of different parameters (variability of cell lineages, cell lifetimes, orientation of cell division, pressures, surface and line tensions, gene expression, etc.). These studies will lead to a reflection on the concept of the average embryo and on its computational representation. The tools developed will open the way to the quantitative study of robustness to environmental and genetic perturbations, and to the identification of bridges between scales of analysis (search for co-varying parameters across scales).
What is available to start the project?
The project will benefit from the conceptual and methodological developments made over the last 10 years by the hosting teams and their collaborators (see references below). These breakthroughs (ASTEC, MorphoNet, Aniseed) place them in a unique position to analyse, experimentally and computationally, the variability of animal embryogenesis, whether natural or in response to environmental or experimental perturbations.
Using these advanced tools, we generated high resolution geometric and mechanical descriptions of 7 embryos of the ascidian Phallusia mammillata, over several hours of development and with a 2 minutes time resolution. You can view a short video highlighting this works.
These embryos and tools constitute a solid basis for the proposed PhD project. The embryo collection is currently expanding to include more WT Phallusia embryos as well as embryos cultured in response to environmental (temperature, salinity, pH) or genetic perturbations.
How to apply?
The ideal candidate will have successfully graduated from a Master’s programme in computer science, bioinformatics or physics recognized by France. S/He will have strong computational skills and some knowledge of developmental biology. A working knowledge of English (B2) is needed. There is no prerequisite in French.
To apply to the project, please contact P. Lemaire (patrick.lemaire[at]crbm.cnrs.fr) as soon as possible and by May 11, 2022 at the latest with a motivation letter, a CV and the names and contact details of 2 academic referees including the PhD supervisor.
References
Guignard L. *, Fiuza U.-M. *, Leggio B., Laussu J., Faure E., Michelin G., Biasuz K., Hufnagel L., Malandain G. #, Godin C. #, Lemaire P.# (2020) Contact-area dependent cell communications and the morphological invariance of ascidian embryogenesis. Science, 369 :6500 eaar5663
Dardaillon, J; Dauga, D; …; Dantec, C.#; Lemaire, P#. (2019) ANISEED 2019: 4D exploration of genetic data for an extended range of tunicates. Nucleic Acids Res. 48(D1): D668-D675
Leggio, B; Laussu J; Carlier, A; Godin, C; Lemaire, P and Faure, E (2019) MorphoNet: An interactive online morphological browser to explore complex multi-scale data. Nat Commun. 10(1):2812
The tunicate team at CRBM (Montpellier, France), headed by Patrick Lemaire, is offering a 2-year post-doctoral fellowship (or a 3-year PhD fellowship for an exceptional candidate) to study the robustness of animal embryonic development to genetic and environmental perturbations, using quantitative live imaging of ascidian embryos. A short video describes a recent piece of work of the team relevant to the project.
Why ascidians?
Ascidians are a group of marine invertebrates. Their embryonic cell lineages and early embryonic stage morphologies have remained essentially identical since the group’s emergence about 400 million years ago. This suggests that they are subject to very strong developmental or evolutionary constraints (Lemaire et al. 2011). Ascidian embryogenesis is also very robust to environmental perturbations of temperature and salinity. The extreme evolutionary and environmental robustness of embryonic geometries and cell lineages contrasts with a rapid genetic divergence between species and intra- and inter-specific variability in gene expression.
What is the project?
The proposed experimental project will provide a quantitative assessment of the developmental robustness of a critical morphogenetic process, ascidian gastrulation (1, 3, 5), to two key environmental parameters (water temperature and salinity) and to genetic perturbations of the morphogenetic driving force apparatus (myosin II, Rho kinase, …). The project will study the magnitude of environmental or genetic variations compatible with the production of a viable larva. It will seek to identify the least – and most – robust developmental processes and time points, i. e. those that collapse first, or resist best to the perturbations. Finally, it will characterize the structure of the natural and experimentally-induced variability in the geometry and mechanical properties of embryos. This may lead to the identification of developmental modules.
The project will involve advanced light-sheet imaging of live micro-injected embryos of the ascidian Phallusia mammillata, followed by the computational and statistical analysis of the acquired developmental movies (see 2, 4).
Who funds the project, and how to apply?
The project is funded by an ANR-NSF binational project grant and will be conducted in collaboration with 3 other teams: Prof. Atef Asnacios (MSC, Paris), Prof. Edwin Munro (U. Chicago, USA) and Prof. Madhav Mani (Northwestern University, Evanston, USA).
Expected candidates will have a PhD in cell and developmental biology, an excellent track record of publications and oral communications, strong skills in fluorescent live imaging and some experience in the computational analysis of large datasets. To apply, send Patrick Lemaire (patrick.lemaire[at]crbm.cnrs.fr) by May 11, 2022 at the latest a motivation letter, a CV and the names and contact details of 2 academic referees including the PhD supervisor. A working knowledge of English (B2) is needed, there is no prerequisite in French.
References
Fiuza U.-M. and Lemaire, P. (2021) Mechanical and genetic control of ascidian endoderm invagination during gastrulation, Semin Cell Dev Biol 120:108-118
Guignard L. *, Fiuza U.-M. *, Leggio B., Laussu J., Faure E., Michelin G., Biasuz K., Hufnagel L., Malandain G. #, Godin C. #, Lemaire P.# (2020) Contact-area dependent cell communications and the morphological invariance of ascidian embryogenesis. Science, 369 :6500 eaar5663
Fiuza U-M, Negishi T., Rouan A., Yasuo H.#, Lemaire P. # (2020) Nodal and Eph signalling relay drives the transition between apical constriction and apico-basal shortening during ascidian endoderm invagination. Development 147: dev186965
Leggio, B; Laussu J; Carlier, A; Godin, C; Lemaire, P and Faure, E (2019) MorphoNet: An interactive online morphological browser to explore complex multi-scale data. Nat Commun.10(1):2812
Sherrard, K., Robin, FB, Lemaire, P., and Munro, E. (2010) Sequential activation of apical and basolateral myosin drives endoderm invagination during ascidian gastrulation, Current Biology, 20(17):1499-510.
Read on for our news roundup of the past month, with an emphasis on what has caught our eyes on twitter. This month, we also include a list of meetings with upcoming deadlines, as well as a selection of preLights.
#leavingacademia
At the beginning of the month, it seemed like everyone was #leavingacademia. Below are a few of our favourite threads that highlight some of the challenges that academics face, but also the upsides of academic research and working with industry.
1/9. There has been a ton of discussion on academic Twitter about reasons to leave academia. This thread is for young scientists who want to stay in academia. Let me first comment on the things I agree with from the “time to leave” tweets:
(deep sigh) I think the current debate about academia vs. industry does not articulate how it really is better for all of us that there is robust competition and introspection about what job opportunities are present to us, and more importantly to our trainees.
It has been great to see the return of so many in-person conference. With ‘conference fever’ in mind, we have put together a list of #devbio and #stemcell meeting with upcoming abstract and registration deadlines. You can find a list of upcoming conferences on our events page. If we are missing your event, then please feel free to add it to our planner.
If you are interested in science communication, would like to improve your writing skills and become part of the preLights community, don’t miss the open call for new preLighters.
If you would like to write for the Node, check out our recent list of writing ideas. If you would like to contribute to our ‘Developing news’ blog, please get in touch at thenode@biologists.com
My journey in the field of mammalian developmental biology began when I joined the lab of Dr Ivan Bedzhov at the Max Planck Institute for Molecular Biomedicine (MPI-MB) in 2016 to pursue my doctoral research. After Ivan and I discussed the many important processes that take place during the peri-implantation phase of embryonic development, it was the unaddressed mechanisms by which the embryo forms the first contacts with the mother during implantation that caught my interest the most.
The process of implantation mediates the first direct interactions between the embryo and the mother. Specialized cells of the mouse embryo, known as trophoblast giant cells (TGCs), invade deep into the uterine tissues, enabling nutrient uptake and gas exchange with the maternal environment. In turn, the uterine stroma rapidly proliferates and completely engulfs the implanting embryo. Thus, studying the process of implantation is fundamentally challenging, as the embryo is concealed by the uterine tissues and its development depends on the maternal support.
According to clinical reports, almost half of the human pregnancies fail at the time of implantation (Boomsma et al., 2009; Koot et al., 2012). However, the cellular mechanisms of implantation and the factors causing termination of pregnancy are very poorly understood. The significance of these open questions and the excitement to understand the first interactions between an embryo and the mother motivated me to work on this project.
A glimpse at the trophoblast – maternal vasculature interactions in utero
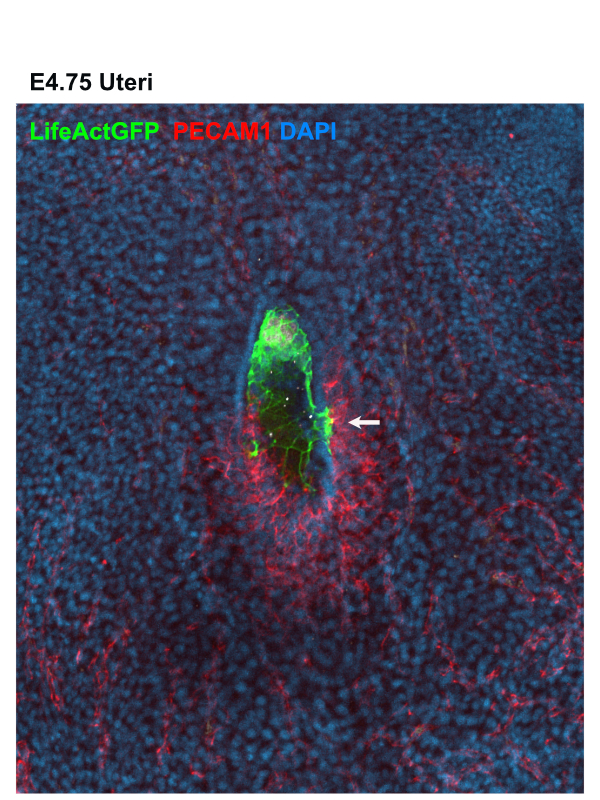
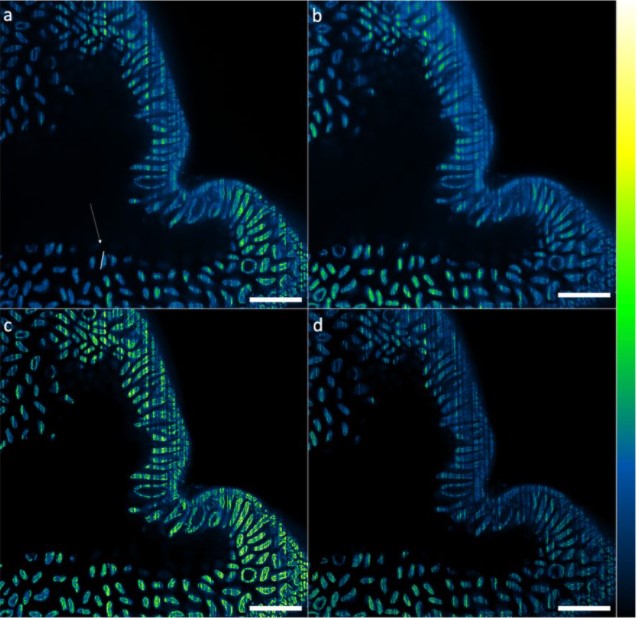
As an early miscarriage can happen because of poor trophoblast penetration and/or inadequate blood supply to the implantation site (Klauber et al., 1997; Reus et al., 2013; Torry et al., 2007), we were keen to understand whether an active crosstalk between the TGCs and the maternal blood vessels takes place during the implantation stages. We examined the spatial organization of the TGCs and determined that the invasive trophoblast is organised as strands of cells penetrating the uterine stroma. Moreover, we found that the TGCs of the implanting embryo intermingle with the surrounding blood vessels (Fig. 1). As static images were insufficient to study the dynamics of these interactions, we decided to establish a 3D biomimetic platform that resembles the biomechanical properties of the uterine stroma.
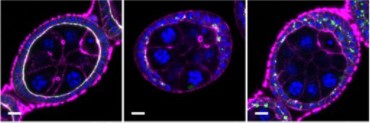
Figure 1: Whole mount image of uteri containing E4.75 LifeAct GFP Embryo stained for – LifeAct GFP Embryo (GFP) in green, blood vessels (PECAM1) in red and nuclei (DAPI) in blue. White arrow indicates invasive trophoblast intermingling with the surrounding blood vessel.
Establishment of the 3D biomimetic platform
First, we determined the stiffness of the maternal tissues using atomic force microscopy (AFM) and examined the cell adhesive properties of the endometrium. As we also had to take into account that the TGCs secrete matrix metalloproteinases (Zhu et al., 2012), we used biodegradable synthetic hydrogels as our artificial substrate. We tuned the stiffness and adhesive features of the hydrogels to mimic the biomechanical properties of the uterine stroma, (in collaboration with Dr Britta Trappmann’s and Dr Adrian Ranga’s labs). Culturing embryos in this 3D environment enabled us to directly observe ex utero implantation of the embryos for the first time, which was very exciting!
As our main focus was to understand the crosstalk between the invading TGCs and the maternal vasculature, we teamed up with Hongyan Long from Dr Britta Trappmann’s lab to incorporate the implanting embryos into a microfluidic device (Trappmann et al., 2017) which allowed us to model the interactions with the blood vessels. After several months of tireless efforts and optimization, we managed to establish the right conditions for co-culturing embryos and endothelial cells in the microfluidic chip. Using this platform, we found that the invasive trophoblast migrates towards the blood vessels to form direct cell-cell contacts, similar to the static images in vivo (Movie 1). This was truly fascinating and a very rewarding moment for us.
Movie 1: Timelapse movie of LifeAct GFP embryo and tdTomato expressing endothelial cells co-cultured in microfluidic device
Uncovering the molecular basis of the trophoblast – vasculature interactions
Next, we wanted to examine the signalling crosstalk that mediates these interactions. We are grateful to Dr Hyun-Woo Jeong from Professor Dr Ralf Adams lab, who helped us with the bioinformatic analysis. We found that the TGCs gain expression of cell surface receptors, ligands and adhesion molecules, similar to the ones expressed in the nearby blood vessels. By functionally examining the main players in this process, we found that platelet-derived growth factor receptor (PDGFR) signalling promotes the establishment of direct cell-cell contacts between the TGCs and the vasculature. As PDGFR signalling has been previously shown to mediate the recruitment of pericytes to the endothelial cells (Lindahl et al., 1997), our findings suggested that the TGCs exploit this signalling pathway to locate the endothelial cells of the maternal blood vessels. Consequently, the expression of compatible cell adhesion molecules in the TGCs and the vasculature, such as VE-cadherin, enables the formation of heterologous cell-cell contacts. This discovery also has a potential clinical relevance, as it has been shown that the PDGFR inhibitor used for the treatment of chronic myeloid leukaemia increases the risk of miscarriage in experimental animals and pregnant patients (Salem et al., 2019; Ault et al., 2006; Pye et al., 2008) and the reason for this side effect was so far obscure.
Bumps along the way
Like most of the projects in academic research, the road of our journey to understand the embryo maternal interaction was not always smooth. We did face unfortunate situations, but at the same time we also got a lot of positive comments from colleagues around the world about the novelty of the project and this encouraged us that we were on the right path. Most of the methods used in this study required intense optimization. We did get stuck sometimes but we kept trying and found new ways to overcome technical limitations, which also helped me to expand my skills and critical thinking. I am really thankful to all our collaborators for their time to discuss and troubleshoot challenging experimental approaches. We learned a lot through failed experiments and eventually when we look at the new discoveries that we’ve made, I think it was all worth the effort!
When we submitted our work to two of the top journals, we experienced major delays as reviewers that accepted the invitation to review our manuscript did not respond and kept us waiting for several months. At this point we had no choice but to move to another journal, as we were running out of time and funding. This happened twice and we lost 9 months just waiting for the initial review, despite the best efforts of the editors of these journals. Having to wait so long was very stressful. Unfortunately, other colleagues have also had similar experiences, so it is up to us, the research community as well as the journals to prevent such toxic behaviour. We finally found a good home for our manuscript in Developmental Cell, where the editors are doing their best to ensure fair peer-review process and present exciting new research to the community.
In the End…
Looking back, this journey has taught me that passion, compassion, clear objectives, perseverance, and interdisciplinary collaborations are key for making new discoveries. Looking forward, I hope that one day, our findings will help scientists understand how the human embryo establishes its first direct contacts with the mother and will lead to therapeutic approaches that substantially decrease the risk of an early miscarriage.
Ault, P., Kantarjian, H., O’Brien, S., Faderl, S., Beran, M., Rios, M.B., Koller, C., Giles, F., Keating, M., Talpaz, M., et al. (2006). Pregnancy among patients with chronic myeloid leukemia treated with imatinib. J Clin Oncol 24, 1204-1208.
Boomsma, C.M., Kavelaars, A., Eijkemans, M.J., Lentjes, E.G., Fauser, B.C., Heijnen, C.J., and Macklon, N.S. (2009). Endometrial secretion analysis identifies a cytokine profile predictive of pregnancy in IVF. Hum Reprod 24, 1427-1435.
Klauber, N., Rohan, R.M., Flynn, E., and D’Amato, R.J. (1997). Critical components of the female reproductive pathway are suppressed by the angiogenesis inhibitor AGM-1470. Nat Med 3, 443-446.
Lindahl, P., Johansson, B.R., Leveen, P., and Betsholtz, C. (1997). Pericyte loss and microaneurysm formation in PDGF-B-deficient mice. Science 277, 242-245.
Pye, S.M., Cortes, J., Ault, P., Hatfield, A., Kantarjian, H., Pilot, R., Rosti, G., and Apperley, J.F. (2008). The effects of imatinib on pregnancy outcome. Blood 111, 5505-5508.
Reus, A.D., El-Harbachi, H., Rousian, M., Willemsen, S.P., Steegers-Theunissen, R.P.M., Steegers, E.A.P., and Exalto, N. (2013). Early first-trimester trophoblast volume in pregnancies that result in live birth or miscarriage. Ultrasound Obst Gyn 42, 577-584.
Salem, W., Li, K., Krapp, C., Ingles, S.A., Bartolomei, M.S., Chung, K., Paulson, R.J., Nowak, R.A., and McGinnis, L.K. (2019). Imatinib treatments have long-term impact on placentation and embryo survival. Sci Rep 9, 2535.
Torry, D.S., Leavenworth, J., Chang, M., Maheshwari, V., Groesch, K., Ball, E.R., and Torry, R.J. (2007). Angiogenesis in implantation. J Assist Reprod Genet 24, 303-315.
 (No Ratings Yet)
(No Ratings Yet)


 (3 votes)
(3 votes)