The Sokol laboratory at the Icahn School of Medicine at Mount Sinai, New York, is looking for a motivated candidate to join our group as a postdoctoral associate. Our laboratory is in the Department of Cell, Developmental and Regenerative Biology, with a number of groups with highly collaborative, developmental and stem cell biology program. Besides high-quality research core facilities, career guidance and professional development training are provided for postdoctoral fellows.
Our group studies how Wnt signaling pathways branch out to control cell lineage and cell movements during vertebrate gastrulation and neurulation. We are also interested in the regulation of apicobasal and planar cell polarity during neural tissue and neural crest formation. A successful candidate will use live cell imaging and biochemical/proteomic approaches to identify new molecules regulating cell signaling and polarity in early vertebrate embryos and mammalian progenitor cells. See the description of our interests at http://labs.icahn.mssm.edu/sokollab/.
Preference will be given to highly motivated and interactive applicants with strong background in cell biology and biochemistry. Interested candidates may send their CV, a short description of relevant expertise, list of publications and the names of three references to Dr. Sergei Sokol (sergei.sokol@mssm.edu).
Closing date for applications 19th of January 2021
Starting date end of 2021, beginning of 2022
The 2020 call for the Spanish “Juan de la Cierva Postdoctoral Fellowships” is now open. This is a fantastic opportunity to join the Araújo group, who are focused on identifying the molecular mechanisms underlying single-cell branching during development and disease. We study this during morphogenesis of the Drosophila melanogaster tracheal and nervous systems (Ricolo et al. Current Biology 2016 and Ricolo and Araújo, eLife, 2020), and are also interested in knowing how single-cell branching affects the whole organismal behaviour. We are combining cell analysis and confocal tissue live imaging, with genetic approaches and CRISPR/Cas9 technologies.
We are based at the Institute of Biomedicine of the University of Barcelona (IBUB) and are looking for a motivated and enthusiastic candidate who will play a central role in the lab. You must have a Ph.D. in areas relevant to cell/developmental biology and/or in computational biology (and have defended your Ph.D. thesis after the 1st of January 2016). Applicants are expected to have excellent interpersonal and communication skills, be highly independent and committed to research in a fast-moving and exciting field.
If you’re interested, please write to sofiajaraujo [at] ub.edu
Nutrients such as phosphate are limiting in most soils. This has a great impact on crop yield in agriculture. Approaches to overcome these challenges often include using fertilizers, that in turn have a detrimental effect on the environment and our climate. Plants have developed adaptation mechanisms to efficiently forage the soil for phosphate such as modifying root system architecture and root hair density. In Wendrich et al., we unexpectedly identified a vascular transcription factor complex that controls root hair formation in response to limiting phosphate conditions.
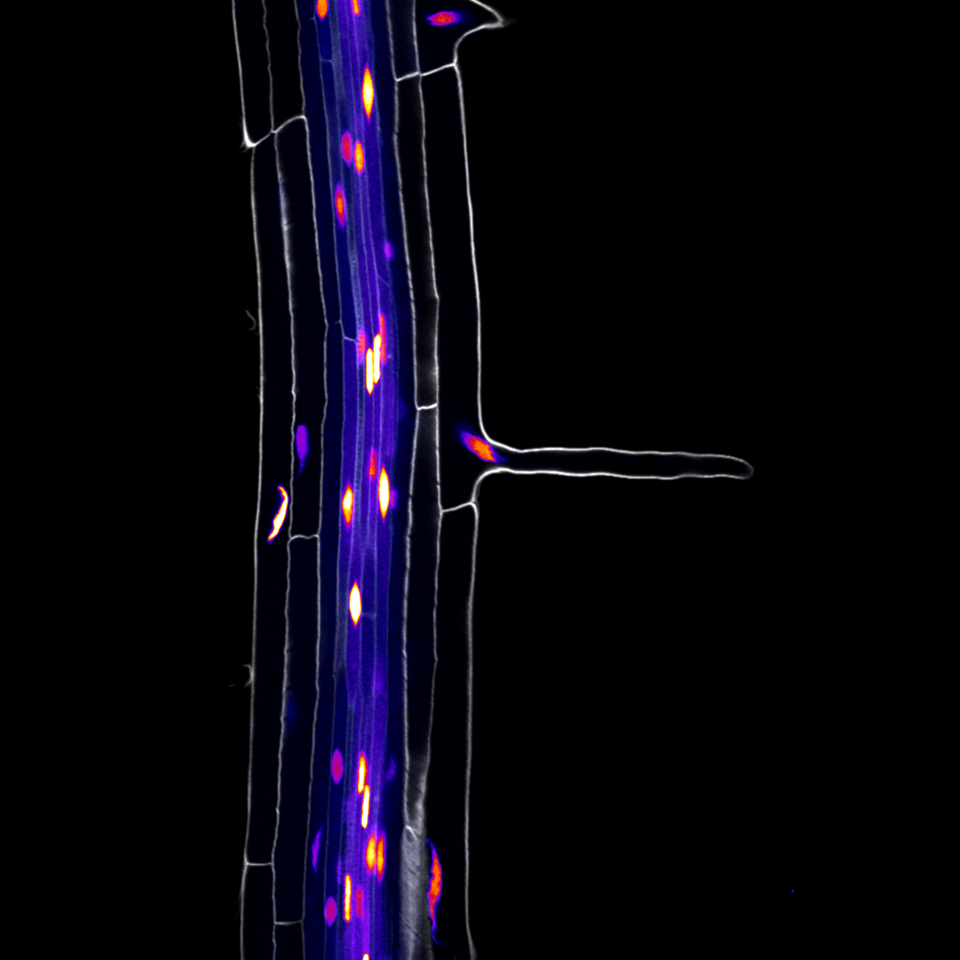
One of the main interests of our lab is identifying the molecular pathways controlled by the TARGET OF MONOPTEROS 5 and LONESOME HIGHWAY (TMO5/LHW) transcription factor heterodimer. In the root meristem, this complex is active in the inner most cells of the vasculature called the xylem. Here it controls the expression of LONELY GUY 4, which encodes an enzyme that is rate-limiting for the conversion of the phytohormone cytokinin into its bioactive form. Cytokinin is involved in many aspects of vascular development including patterning and proliferation. Using bulk transcriptomics, we previously revealed 273 target genes of the TMO5/LHW complex, but given the downstream intermediate cytokinin is mobile, we expected that many of these target genes would be expressed outside of the xylem cells. This is where this project started.
At that time, single-cell technologies were quickly becoming important in other fields and proved ideal to determine gene expression at unprecedented resolution. We thus decided we needed to get this technology up and running to answer our research question. Our institute had recently invested heavily to establish the technique, with commercial microfluidics systems and custom bioinformatic pipelines, but it remained unclear how reliable and efficient this would work on isolated plant cells. We teamed up with the group of Yvan Saeys in our neighboring VIB research center to generate a reference dataset for the Arabidopsis root meristem. We first needed to optimize the workflow leading up to the microfluidics single cell droplet emulsion. With their tough cell walls, plant cells are not easily dissociated from their tissue, requiring enzymatic digestion of the cell walls, called “protoplasting”. Cells in the outer layers are more accessible for these enzymes and tend to be overrepresented after in protoplast suspensions. We therefore first made sure that our single cell protoplast suspensions contained a good representation of each cell type, inner and outer layers, and was free of most debris and impurities, such as dead cells.
Unfortunately, this was the easy part. Now came the analysis, annotation and, importantly, full validation of the technology. This was where our collaboration effort truly started to pay off. The analysis pipelines were custom redesigned for our plant samples. This included building a visualization tool, for quick access to the data. Endless discussions, quick face-to-face check-ups and back-and-forth emailing ensured that we quickly learned from each other and from the dataset. Because of the rich resources available for Arabidopsis, we were able to annotate the main cell types of the root such as epidermis, ground tissues and vascular tissues. We however decided to take it a step further and zoom in to the level of sub-cell-types. Especially the different cell types of the vascular system were more difficult to annotate, as several cell types, like for example the procambium, only have limited known marker genes that are specifically expressed. Sub-cell-type predictions were confirmed by cell ploidy analysis and an extensive trajectory inference. By iteratively applying these trajectory methods, and having an open communication line between wet- and drylab, we were able to assess the robustness of the predicted trajectories and the resulting predicted subcluster annotations.
We next set out to evaluate the predictive power of this dataset, by construction of promoter reporter lines of genes with hitherto unknown expression patterns. All group members chipped in and a remarkable 100% of the lines with stable expression patterns validated the predicted cell type and developmental states. We were thus eager to apply this rich dataset on the 273 target genes of the TMO5/LHW complex previously identified. We identified an unexpected overrepresentation of genes expressed in root hairs (trichoblasts), suggesting that these vascular transcription factors may play a role in the development of the outermost trichoblast cells. Indeed, when overexpressing TMO5 and LHW in all cells of the root meristem, this dramatically increased the density of root hairs, resembling wild type roots grown in phosphate limiting conditions. Extensive follow-up genetic work allowed us to conclude that cytokinin, produced in inner vascular cells, acts as a mobile intermediate to control root hair response to low phosphate conditions in the outer most epidermal cell.
In our opinion, this is a good example of how collaborations within and between teams with different expertise can lead to new biological insights.
Postdoctoral Fellow (m/f/d) | RNA/Protein Biochemistry and Molecular Biology
MAX PLANCK INSTITUTE FOR HEART AND LUNG RESEARCH – W. G. KERCKHOFF INSTITUTE, BAD NAUHEIM
TYPE OF JOB
SCIENTIST
Job offer from December 08, 2020
The Max Planck Institute for Heart and Lung Research in Bad Nauheim (near Frankfurt, Germany), Department of Developmental Genetics (Prof. Dr. Didier Stainier) invites applications for a Postdoctoral Fellow (m/f/d) in RNA/Protein Biochemistry and Molecular Biology to study Genetic CompensationReference Number 2020_30.
A highly motivated postdoctoral candidate is invited to lead new projects to address fundamental questions in Genetic Compensation and Transcriptional Adaptation.
We have recently discovered a new form of genetic compensation that we have termed transcriptional adaptation. Briefly, transcriptional adaptation occurs when a premature termination codon leads to mutant mRNA degradation. The degradation fragments in turn modulate the expression of related genes, and consequently varying degrees of phenotypic rescue in some cases. We have observed this phenomenon in zebrafish, mouse, C. elegans, and more recently humans.
This is a novel and quickly developing field of research where many open questions remain to be addressed including 1) the nature and trafficking of the degradation fragments, and 2) the regulation of related genes by the degradation fragments, or their derivatives. For more detailed information please see our recent publications:
A Ph.D. in biology, biochemistry, genetics or a similar subject with a focus on molecular biology, cell biology, biochemistry and/or genetics. Knowledge of RNA and/or protein biochemistry and molecular biology is a plus.
About the employer:
The Max Planck Institute for Heart and Lung Research in Bad Nauheim is an interdisciplinary research institution with international flair. Our researchers have the opportunity to work on various model systems by making use of the latest cutting-edge technologies. Researchers are supported by state-of-the-art core facilities which offer services in next generation sequencing, proteomics, bioinformatics, cytometry, microscopy, and small animal imaging.
The Max Planck Society strives for gender and diversity equality; we welcome applications from all backgrounds. Furthermore, the Max Planck Society is committed to increasing the number of individuals with disabilities in its workforce and therefore encourages applications from such qualified individuals.
To apply, please submit the names and contact information for 2-3 references, a CV, and a short statement (2 pages max.) of your research experience and interests to didier.stainier@mpi-bn.mpg.de
Max Planck Institute for Heart and Lung Research
Department of Developmental Genetics
Ludwigstraße 43
61231 Bad Nauheim
Germany
Duke University’s Departments of Orthopaedic Surgery and Cell Biology invite applications for a tenure track faculty position in the field of musculoskeletal cell biology. The ideal candidate will have a PhD, MD, or MD/PhD and significant postdoctoral or faculty experience. They should have a record of outstanding scientific accomplishments and a well-developed research plan in a key research discipline such as: bone biology and osteoporosis, skeletal stem cell biology, skeletal regeneration, cartilage biology and arthritis, muscle/tendon biology and regeneration, or musculoskeletal and neuromuscular disorder research.
An appointment at any academic level is available depending on the expertise and track record of the candidate. The successful candidate will complement and enhance our substantial existing strengths in cell biology, orthopaedics, sarcoma, bioengineering, developmental biology, and musculoskeletal research utilizing a combination of in vivo and in vitro models. New faculty will receive a generous startup package and laboratory space. We expect the successful candidate to initiate and maintain an original, competitive, and independently funded research program.
Interested candidates should submit application materials including a cover letter, curriculum vitae, statement of accomplishments and future research goals (two-page limit), a diversity statement, electronic reprints of up to three recent articles, and arrange to have up to three letters of reference provided.
Applications will be reviewed on a rolling basis and should be submitted by January 31, 2021, however later applications will be considered until the position is filled. Initial interviews will be held virtually.
Matthew J. Hilton, PhD
Search Committee Chair
Associate Chair for Research, Basic/Translational Sciences
Department of Orthopaedic Surgery
Duke University School of Medicine
Policy Statement: Duke University prohibits discrimination and harassment, and provides equal employment opportunity without regard to an individual’s age, color, disability, gender, gender expression, gender identity, genetic information, national origin, race, religion, sex, sexual orientation, or veteran status. Duke is committed to recruiting, hiring, and promoting qualified women, minorities, individuals with disabilities, and veterans. Pursuant to Title IX of the Education Amendment of 1972, Duke prohibits discrimination on the basis of sex in any of its educational programs or activities. For more information regarding Duke’s Equal Opportunity/Affirmative Action policy and our commitment to diversity and equity go to: https://oie.duke.edu/
In the Genetics of Cell Behaviour in Development laboratory, at the Department of Genetics, Microbiology and Statistics at the University of Barcelona, we work with the terminal cells (TCs) of the tracheal system of Drosophila melanogaster, which are very interesting cells. During embryonic development, they are able to generate a tube within their cytoplasm, a so-called seamless, subcellular tube. As the larvae hatch and grow these tiny tubes are able to branch inside a single cell to provide oxygen to the various tissues in the animal.
Embryonic and larval tracheal TCs are a powerful model to understand the mechanisms that lead a single-cell to change its shape, and generate a subcellular seamless tube, in a process analogous to capillary blood-vessel sprouting. Cell elongation and subcellular tube formation are dependent on dynamic cytoskeletal reorganization. Microtubules and actin are major players in this active cytoskeletal process. We have been studying the initial events in the formation of this subcellular tube, focusing on what triggers these changes in the cytoskeleton.
When we investigated the role of centrosomes as Microtubule Organizing Centres (MTOCs) in TCs (Ricolo et al., 2016), we became interested in analysing microtubule bundles inside TCs. We tested the tracheal overexpression of several constructs consisting of MT-associated proteins fused with fluorophores. Among them, we used transgenic lines overexpressing the Drosophila spectraplakin Short-stop (Shot).
Shot is a giant multifunctional protein responsible for many aspects of cytoskeleton organization in different tissues. Shot can operate as a single cytoskeleton component, can coordinate cytoskeletal elements such as actin and microtubules, and can also interact with other cellular structures. The functional versatility of Shot is probably reflected by the abundant generation of different isoforms, and by the characteristics of its protein domains. The full-length isoform A (Shot-A) contains an NH2-terminal actin-binding domain (ABD), different central domains (rod-like domain, plakin spectrin-like repeats and an EF-hand), and a C-terminal domain involved in MT interaction (GRD and C-tail domains). Shot-A has been shown to act as an actin-microtubule crosslinker. Isoform C (Shot-C) lacks a proper actin-binding domain and has been described to interact mainly with MTs (Voelzmann et al., 2017).
Since we were interested in visualizing MTs in TCs in live embryos, we tested the tracheal overexpression of Shot-C using the transgenic line UAS-shotC-GFP. This was a clear winner! GFP staining of embryos overexpressing Shot-C-GFP, allowed us to beautifully label MTs; we could clearly see the MT-fibres inside the TCs and they were bright enough that we could image them live, as the embryo develops.
But… (there is always a but). When you want to see a biological structure through the use of an overexpression system in Drosophila embryos, as in other organisms or cell culture systems, you have to be very careful. Often the overexpression of a specific protein can perturb the physiological state of the cell; in other words, it can give rise to a mutant phenotype. So, we decided to analyze if the embryonic overexpression of Shot caused any changes during TC extension and lumen development.
Surprisingly, we found that Shot overexpression induced the generation of an extra-subcellular lumen inside the TC, a very similar phenotype to the one caused by extra centrosome numbers. So, by searching for ways of illuminating MTs, we had serendipitously found another way to induce luminal branching at embryonic stages. Of course, our next question was whether Shot was inducing higher numbers of centrosomes in TCs. Nicely, we were able to genetically prove that Shot acted in a different way to generate the extra-subcellular lumen branching event.
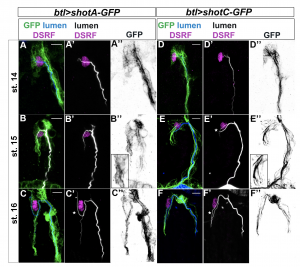
ShotOE induces luminal branching through its microtubule-binding domain. Lateral view of DB tip cells from st.14 to st.16, of btl >shotA-GFP embryos (A–C) and btl >shotCGFP (D–F). Embryos were stained with GFP (green in A-F and grey A’’-F’’) to visualise Shot-GFP, DSRF to mark the TC nuclei (in magenta) and CBP to stain the chitinous lumen (blue in A-F and white A’-F’). Both overexpressing conditions induced Extra subcellular lumina (white stars). Note the GFP was more diffuse in the cytoplasm of the TCs of embryos overexpressing shotA, and more organised in bundles in the TCs overexpressing shotC. Anterior side of embryo is on the left and dorsal side up. Scale bars 5 μm. From Ricolo and Araújo, 2020, eLife.
The results of our experiments told us that the more Shot we expressed in the TC, the more subcellular branching we could generate. So, how does this Shot overexpression generate branched subcellular lumina? One hypothesis was that an excess of MT-stabilization could induce the generation of acentrosomal branching points along pre-existent microtubule bundles along the luminal track. If this was true, the overexpression of other proteins involved in MT stabilization should be also able to induce extra subcellullar lumina. We tested this hypothesis and found that overexpression of the microtubule-associated protein Tau was able to generate the same luminal phenotype.
Analyzing the complete loss-of-function of shot we observed defects in luminal formation and cell elongation. We observed, using rescue experiments and analysis of endogenous Shot localization that this phenotype depended on Shot’s ability to crosslink MTs and actin. Surprisingly, we were able to rescue the shot null phenotype overexpressing Tau indicating a role for Tau in mediating the connection between actin and microtubules in TCs.
With our work, we showed the importance of actin-microtubule crosslinking in the formation of novel subcellular tubes by single-cells (Ricolo and Araújo, 2020). But also, this work has taught us that by doing research you can open new exploratory paths in an unpredictable way. Serendipity plays a role in science. You just need to keep an open mind and follow your curiosity!
The Evolutionary Developmental Biology Lab at the Francis Crick Institute is seeking a laboratory research scientist that will help establish the laboratory, manage its day-to-day operations, and lead the generation of large-scale genomics datasets.
The Evolutionary Developmental Biology Lab will open early 2021. We study how organs originate and how they diversify in form and function across species. We tackle this problem by combining evolutionary and developmental biology with large-scale comparative genomics. We are an interdisciplinary group and highly collaborative.
In our lab we will generate and analyse our own large-scale genomics datasets, including single-cell transcriptomics and epigenomics, whole-genome sequencing, and spatial transcriptomics. The laboratory research scientist will set-up the wet lab and lead the data production of the group. This will be done in close collaboration with several science technology platforms at the Institute (animal facilities, single-cell genomics, sequencing).
Evolutionary Developmental Biology Lab
One of the most fundamental problems in biology is understanding how novelties arise, whether they be new cells, new tissues, or whole new organs. Our group tackles this problem by identifying the genetic and developmental processes responsible for the origin, and subsequent evolution, of vertebrate organs.
Our group uses the placenta as a model to understand how organs originate and evolve. We study the placenta because placentas have evolved independently many times across vertebrates and because they exhibit an extraordinary diversity of forms and functions. We study the placenta within two distinct, but complementary, contexts. In mammals, we work on one of the most fascinating aspects of pregnancy, the mother’s tolerance to the intimate contact between its own cells and those of her foetus. We want to understand how these critical interactions between the mother and the fetal cells have evolved across mammals. In fish, we address the question of how organs are created. We study a family of fish where placentas have evolved independently multiple times. By studying multiple independent inventions of the placenta we try to identify general principles guiding the evolution of new organs at the level of genes, cells and developmental processes.
PhD position available in the lab of Dr Chema Martin (Queen Mary University of London) in collaboration with Oxford Nanopore Technologies, Dr Alex de Mendoza (QMUL) and Dr Paul Hurd (QMUL) to dissect the evolution of 3D genome structure with Pore-C Nanopore sequencing.
Background
The 3D structure of the genome – the set of DNA molecules with the guidelines to build an organism – is fundamental for proper expression of the genetic information contained in it [1]. However, how this 3D structure evolves and is influenced by other features of the genome is still unclear. This is because our understanding of this biological phenomenon is mostly limited to a handful of model organisms and by the fact that current methods to reconstruct the 3D structure of genomes depend on short-read sequencing. In late 2019, Oxford Nanopore Technologies (ONT) released Pore-C, a method to apply long-read sequencing to reconstruct 3D genome structure that overcomes current limitations in chromatin conformation approaches [2]. In parallel, the Martín-Durán lab is establishing a set of marine segmented worms (annelids) as research model systems for comparative evolutionary genomics [3]. What are the genomic features controlling 3D genome structure in annelids? How do these mechanisms generate variability in 3D genome structure among species? Can we develop Pore-C as a widespread approach to reconstruct 3D genome structure?
In this project, you will rigorously answer these questions combining state-of-the-art experimental and computation approaches in a unique academic-industry collaborative environment.
– You will have access to large genomic databases, and in-house live organisms to fuel your investigation.
– You will gain experience of molecular techniques (epigenomics, nanopore long-read sequencing), bioinformatics (pipelines to analyse Pore-C data), and statistics.
– You will be encouraged to develop your own ideas and hypotheses.
Benefits
This is a BBSRC LIDo iCASE PhD fully funded position, including home (UK) tuition fees and a tax-free stipend in the region of £17,285. The student will become part of Queen Mary’s Doctoral College, which provides training and development opportunities and financial support for research. The student will also have access to a Researcher Development Programme designed to help recognise and develop key skills and attributes needed to effectively manage research, and to prepare and plan for the next stages of their career. In addition, the student will enjoy a 6-months placement at Oxford Nanopore Technologies headquarters in Oxford (UK), where the students will integrate within the research and development team.
Skills/Qualifications
In a multidisciplinary project like this, candidates are unlikely to have a background in all disciplines involved. The most important qualification is motivation, enthusiasm and that the project appeals to you. However, previous computational experience would be important. We can envisage strong candidates coming through a variety of routes including:
– practical molecular biology
– developmental and cell biology
– biotechnology
– computational biology
Application
To apply, students should have a 1st class degree or have received a MSc in a relevant field (i.e. molecular biology, genetics and evolutionary genomics, developmental and cell biology, biotechnology, bioinformatics) or are about to finish their MSc.
1. Rowley, M.J., Corces, V.G. (2018) Organizational principles of 3D genome architecture. Nat Rev Genet 19, 789–800. https://doi.org/10.1038/s41576-018-0060-8
2. Netha Ulahannan, et al. (2019) Nanopore sequencing of DNA concatemers reveals higher-order features of chromatin structure. bioRxiv 833590; doi: https://doi.org/10.1101/833590
3. José M. Martín-Durán, et al. (2020) Conservative route to genome compaction in a miniature annelid. Nature Ecology and Evolution. https://doi.org/10.1038/s41559-020-01327-6
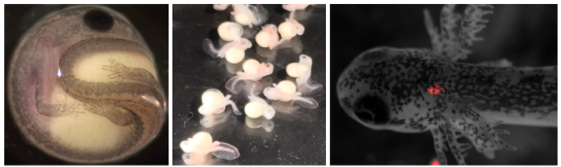
Drs. Hillary Maddin and Tetsuto Miyashita are seeking motivated students and fellows to join our Carleton-based team in Ottawa, Canada. Successful candidates will participate in a newly funded project aimed at understanding various aspects of the development and evolution of the vertebrate skeleton. In particular, projects will focus on investigating skull and limb development in model and non-model species of amphibians. Collaborators for these projects include Chris Joslin (Carleton), James Hanken (Harvard), and Ryan Kerney (Gettysburg). MSc, PhD, and postdoctoral positions available.
Carleton University has research partnerships with Canadian Museum of Nature, which houses natural history collections of over 14 million specimens, and the University of Ottawa, which offers an extensive network of biomedical and life science labs and the state-of-the-art next-generation sequencing facility. Ottawa is a vibrant, multicultural, and affordable city embraced in the green belt. Browse some overview and Google Image search summary of life in Canada’s capital.
 (No Ratings Yet)
(No Ratings Yet)
 (1 votes)
(1 votes)

 (7 votes)
(7 votes)