We are at the National Institute of Child Health and Human Development (NICHD) at NIH. Our lab is interested in understanding cell lineage differentiation, gene regulation and how non-coding DNA elements and the 3D architecture of chromosomes contribute to these processes during early mouse development.
Fully-funded postdoc positions up to five years including health benefits.
Opportunity to start your own research program or lead ongoing projects.
Who you are:
You share our enthusiasm for epigenetics, gene regulation, nuclear organization and mouse development.
You have PhD-experience in one or more of the following: mouse development, mouse genetics, epigenetics, massively-parallel sequencing techniques or computational biology.
Advantages of postdoctoral training at NIH
Fully-funded positions up to five years.
Large, diverse and extraordinary scientific network at the NIH/Bethesda campus. The NIH research community is unparalleled in its size, diversity and resources.
Possibility of living in a diverse, liberal and vibrant city: Washington DC
Or living in a calm residential area with great schools and good affordable housing, Bethesda and Rockville.
2 paragraph cover letter explaining your scientific trajectory and why you would like to join us.
CV and email contacts for 3 references.
The NIH is dedicated to building a diverse community in its training and employment programs.
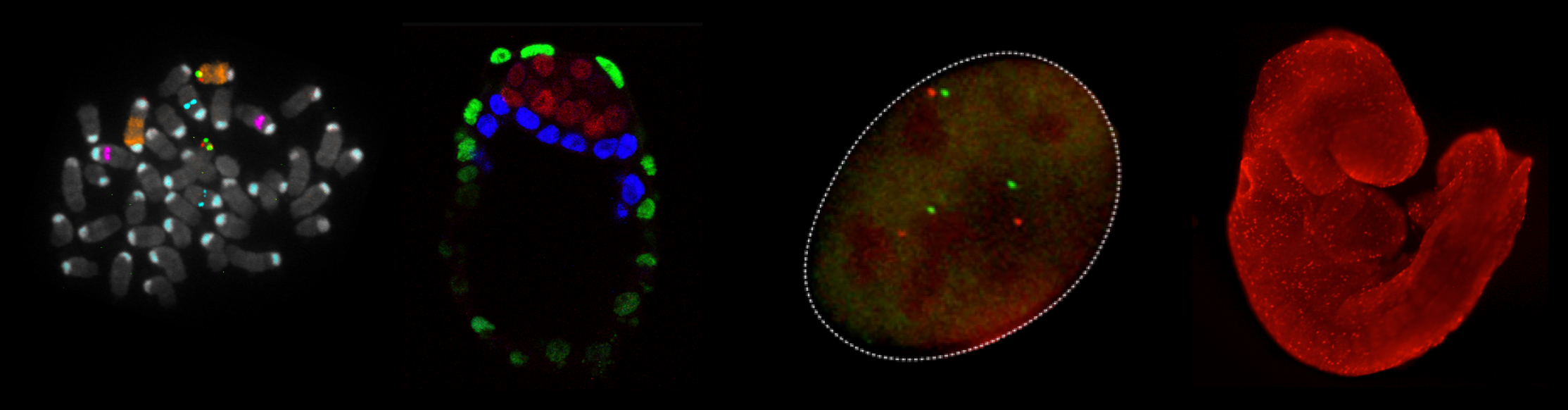
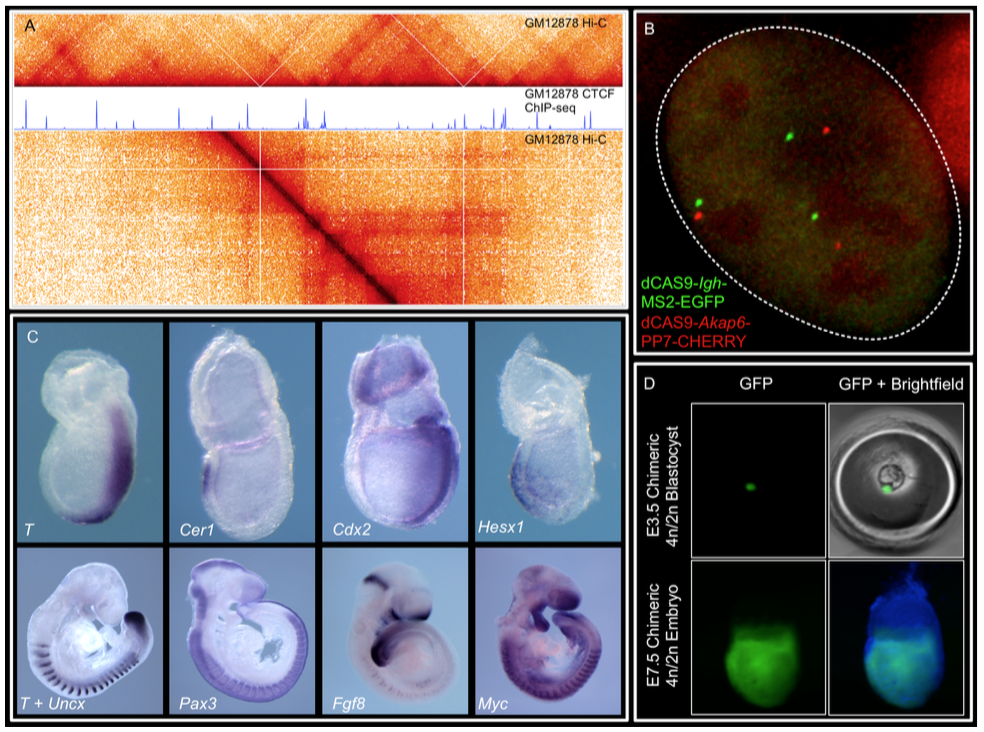
We combine imaging techniques in both fixed and living cells with sequencing- based genomic techniques that assess DNA-DNA interactions. (A) Hi-C and CTCF ChIP- seq of GM1278 cells (B) dCAS9 MCP-EGFP and PCP-CHERRY live imaging of the Igh and Akap6 loci. The mouse embryo is an unparalleled system in mammalian biology for understanding how tissue- specific gene expression is achieved. (C) Whole mount in-situ hybridization for patterning markers in mid and late gastrulating embryos. (D) Tetraploid aggregation with GFP ES cells allows generation of fully ES-cell derived embryos. (No Ratings Yet) Loading...
Postdoc in Developmental and Cancer Biology in the Arnes Group at BRIC, Faculty of Health and Medical Sciences, University of Copenhagen
The Biotechnology Research and Innovation Center (BRIC) at the University of Copenhagen is seeking to recruit a highly motivated postdoctoral researcher in the laboratory of Dr. Arnes. The position is initially for two years and can be filled early spring 2019.
We are looking for a Ph.D level molecular biologist with a strong background in developmental biology. In our group, we seek to answer fundamental questions in development and cancer using a combination of genetically modified mouse models and in vitro differentiation of pluripotent stem cells .
Our research
Terminally differentiated cells in the pancreas have the plasticity to adopt alternative cell fates through dedifferentiation and reprogramming. This cellular plasticity, although considered a physiological process of tissue homeostasis, renders the tissue susceptible to diseases, such as cancer and diabetes. Cell-type specific regulatory programs maintain the identity and function of mature cell types, and although we have a clear picture of the steady-state transcriptional and epigenetic landscape in development and disease, the molecular regulators of cell fate transitions are not well understood. Our goal is to define the mechanism controlling cellular plasticity in pancreas development and cancer. To achieve this general goal, we focus on the following areas of research:
Define the dynamic molecular events, with emphasis in non-coding regulatory elements, leading to cell fate transition in development and cancer.
Identify and characterize functional long non-coding RNAs regulators of cellular plasticity in vivo and in vitro.
Examine the therapeutic potential of cellular plasticity in regenerative medicine and cancer.
These studies are intended to discover novel molecular markers of tumor progression and targetable molecular mediators of cellular identity to prevent the initiation and progression of pancreatic cancer.
Relevant publications and reviews:
Arnes L*#, Liu Z#, Wang J#, Maurer HC, Bommakanti N, Garofalo DC, Balderes DA, Sussel L, Olive KP*, Rabadan R*. Comprehensive characterisation of compartment specific long non-coding RNAs associated with pancreatic ductal adenocarcinoma. Gut. 2018 Feb 10. *Corresponding author
Arnes L, Akerman I, Balderes D, Ferrer J, Sussel L. Blinc1 encodes a long non-coding RNA that regulates islet beta cell formation and function. Genes Dev. 2016 Mar 1;30(5):502-7
Arnes L and Sussel L. Epigenetic modifications and long non-coding RNAs influence pancreas development and function. Trends Genet. 2015 Jun;31(6):290-9.
Your profile
The successful candidate must be able to work independently: plan experiments, develop methods and conduct data analysis as well as communicate the results. (S)he will be encouraged to develop her/his ideas within the laboratory goals. (S)he must be able to work independently, as well as collaborate within the group and with other research groups. (S)he is expected to interact with experimental, computational and clinical laboratories in translational studies. (S)he must actively participate in journal clubs, seminars, lab meetings, mentoring of students, etc. in the laboratory, and research activities organized at BRIC and DanStem.
Your qualifications
We are looking for a highly motivated and enthusiastic scientist with the following competences and experience:
Curious and driven researcher with independent and critical thinking.
Strong background in tissue culture and molecular biology, genome editing and next-generation sequencing (ChIP-, RNA-, ATAC-sequencing) is mandatory.
Experience with high throughput screening will be positively evaluated.
Previous experience in live imaging of tumor progression, in-vivo mouse models of cancer, and bioinformatic analysis of large gene expression datasets is highly desirable.
Basic knowledge in systems biology and bioinformatics is an advantage and opportunities for further education will be given.
Publications in the area of gene regulation, transcriptional networks, non-coding RNA is desirable but not mandatory.
Excellent written and oral communication skills in English.
About BRIC
BRIC is a dynamic and international research center. We offer good working and career conditions in an open and collaborative atmosphere. Our state-of-the-art research and core facilities include modern laboratories shared between 25 research groups at BRIC and Finsen Laboratory.
The research groups at BRIC and Finsen are closely collaborating. We have our own PhD programme, MoMeD, a young researchers club ASAP, scientific clubs, a Group Leader forum with regular meetings and Liaison Committee with focus on work environment. On a weekly basis, we have our BRIC-CEHA seminars with invited speakers. To ease the daily administrative tasks for our researchers, the well-functioning administration assists researchers with HR, order and delivery, communication, funding and much more.
Internationally, BRIC actively participates in the European alliance, EU-life, consisting of 13 excellent life science research institutions. This will further strengthen and promote BRIC research and research education programme.
The place of employment is at BRIC, University of Copenhagen, Ole Maaløes Vej 5, 2200 Copenhagen N.
The employment as postdoc is a full time and fixed-term position for 2 years. Starting date is 1 March 2019 or as soon as possible thereafter.
Salary, pension and terms of employment will be in accordance with the agreement between the Danish Ministry of Finance and AC (Danish Confederation of Professional Associations). Currently, the monthly salary starts at 33,700 DKK/ approx. 4,500 Euro (October 2018-level). Depending on qualifications, a supplement may be negotiated. The employer will pay an additional 17.1 % to your pension fund.
Non-Danish and Danish applicants may be eligible for tax reductions, if they hold a PhD degree and have not lived in Denmark the last 10 years.
The position is covered by the “Memorandum on Job Structure for Academic Staff at the Universities” of 18 September 2015.
International applicants may find this link useful: www.ism.ku.dk to the university’s International Staff Mobility unit.
Application procedure
Your online application must be submitted in English by clicking ‘Apply now’ below and must include the following documents/attachments – all in PDF format:
1. Motivated letter of application (max. one page).
2. CV incl. education, work/research experience, language skills and other skills relevant for the position.
3. A certified/signed copy of a) PhD certificate and b) Master of Science certificate. If the PhD is not completed, a written statement from the supervisor will do.
4. List of publications.
Application deadline: February 1, 2019, 23.59pm CET
We reserve the right not to consider material received after the deadline, and not to consider applications that do not live up to the abovementioned requirements.
The further process
After the expiry of the deadline for applications, the authorized recruitment manager selects applicants for assessment on the advice of the hiring committee. All applicants are then immediately notified whether their application has been passed for assessment by an unbiased assessor. Once the assessment work has been completed each applicant has the opportunity to comment on the part of the assessment that relates to the applicant him/herself.
The applicant will be assessed according to the Ministerial Order no. 242 of 13 March 2012 on the Appointment of Academic Staff at Universities.
University of Copenhagen wish to reflect the diversity of society and welcome applications from all qualified candidates regardless of age, disability, gender, nationality, race, religion or sexual orientation. Appointment will be based on merit alone.
The Department of Biology at Swarthmore College invites applications for a two-year visiting assistant professor position starting in August of 2019. Teaching responsibilities consist of one course with weekly laboratory sections each semester. The applicant’s course offerings are anticipated to include a team-taught introductory cell and molecular biology course, an intermediate-level genetics course, and another intermediate course aligned with the applicant’s interests. Additionally, there may be an opportunity to teach an advanced seminar-style course (with laboratory projects) in an area that is complementary to our existing curriculum. Funds are available for travel to professional meetings and to support undergraduate research students during the academic year and the summer.
Located in the immediate suburbs of Philadelphia and just 20 miles from Wilmington DE, Swarthmore College is a highly selective liberal arts college whose mission combines academic rigor with social responsibility. Swarthmore has a strong institutional commitment to diversity, and actively seeks and welcomes applications from candidates with exceptional qualifications, particularly those with demonstrable commitments to a more inclusive society and world. Swarthmore is an Equal Opportunity Employer. Applicants from traditionally underrepresented groups are strongly encouraged to apply. For more information on Faculty Diversity and Excellence at Swarthmore, see http://www.swarthmore.edu/faculty-diversity-excellence/information-candidates-new-faculty
Applicants should have a Ph.D., teaching experience, and a strong commitment to undergraduate education. The strongest candidates will be expected to demonstrate a commitment to teaching that speaks to and motivates undergraduates from diverse backgrounds.
All application materials (cover letter, curriculum vitae, statements of teaching and research interests, and three letters of recommendation) should be submitted online at apply.interfolio.com/59011. Review of applications will begin on February 25th, 2019. For more information, please visit our website at www.swarthmore.edu/biology. Questions regarding this position should be addressed to the search chair, Brad Davidson, at genetics_search@swarthmore.edu
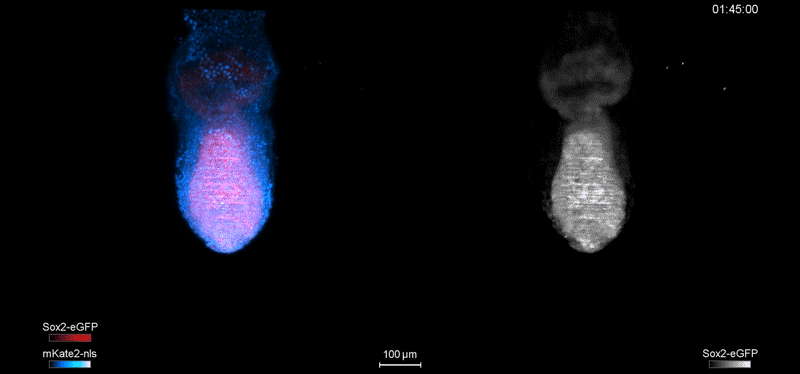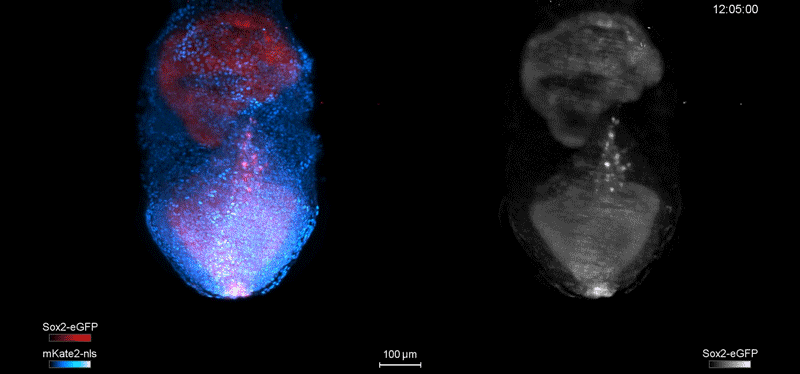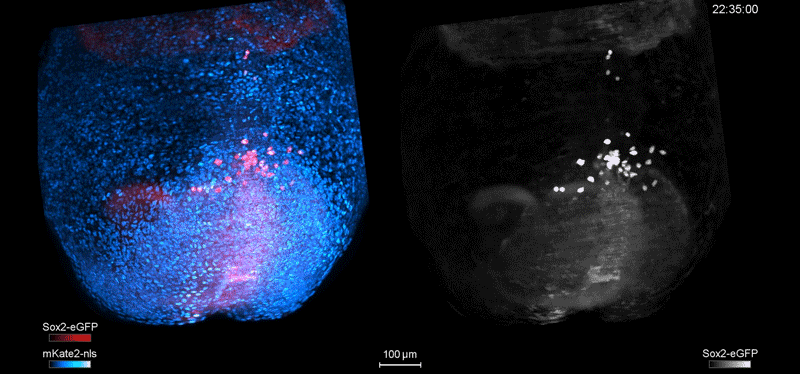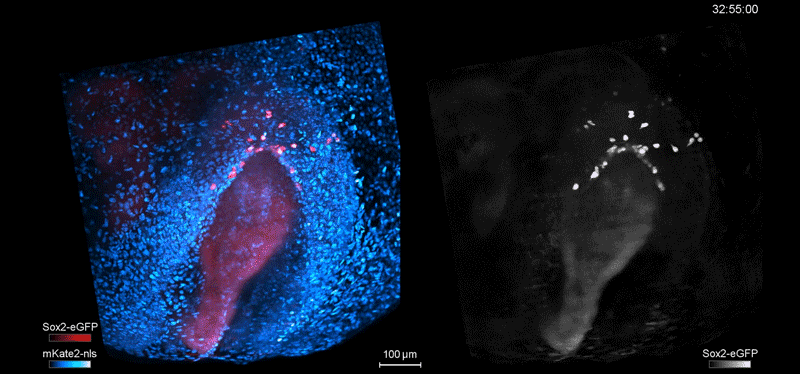
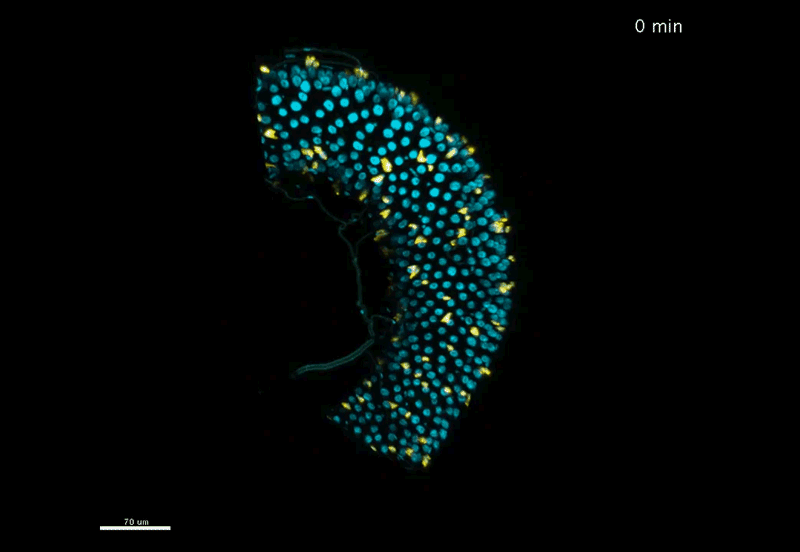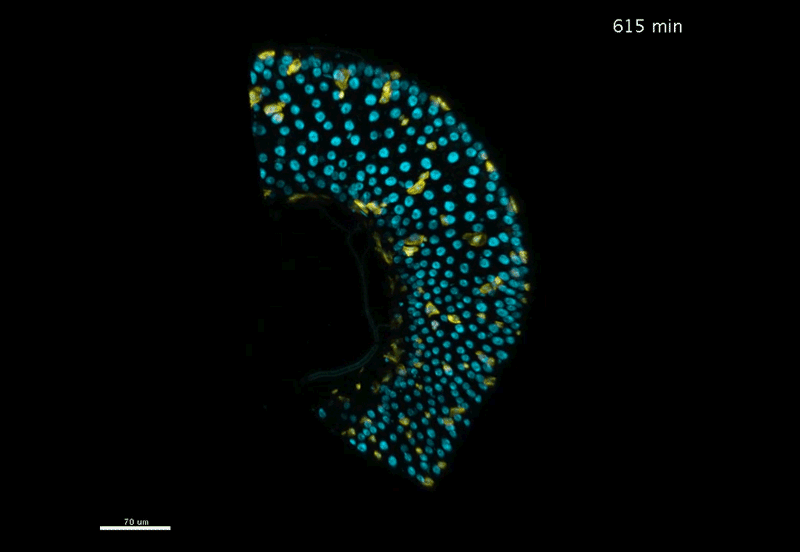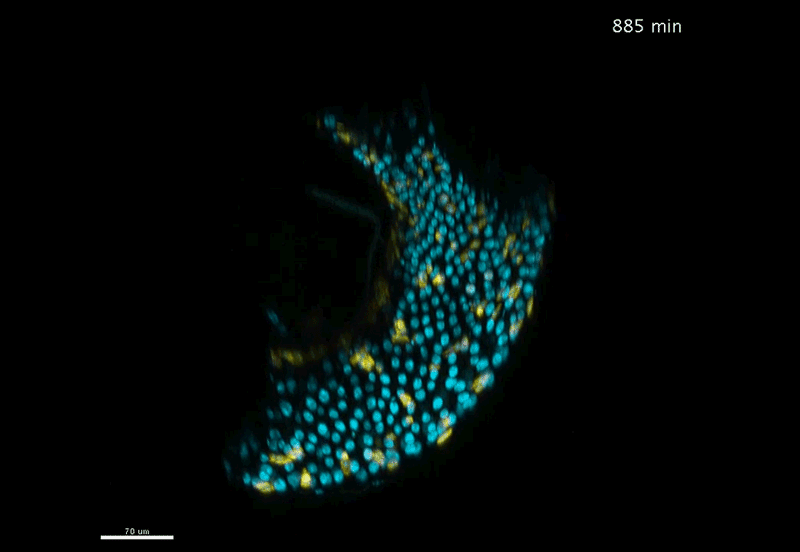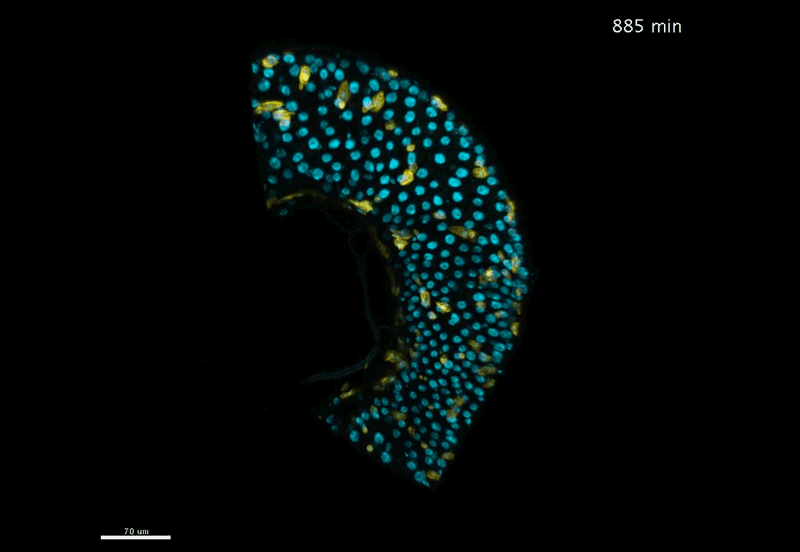
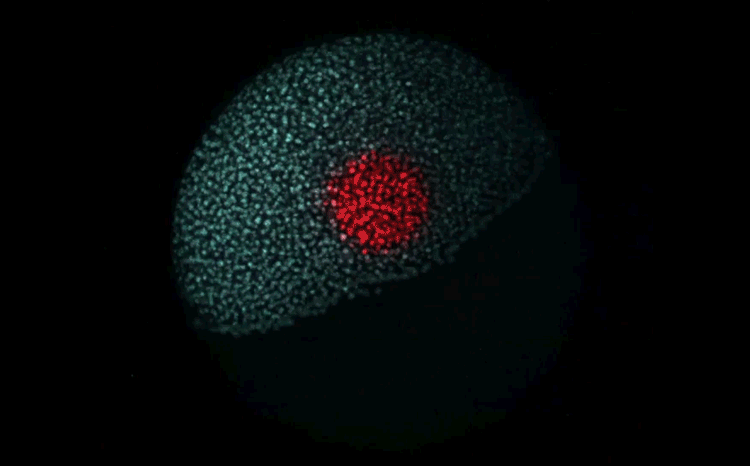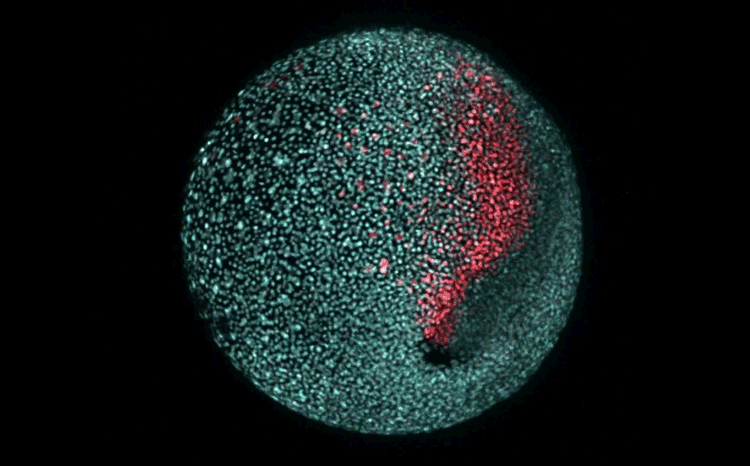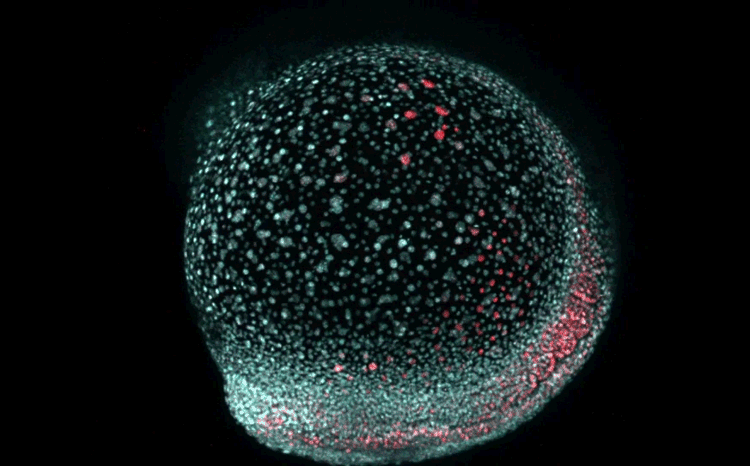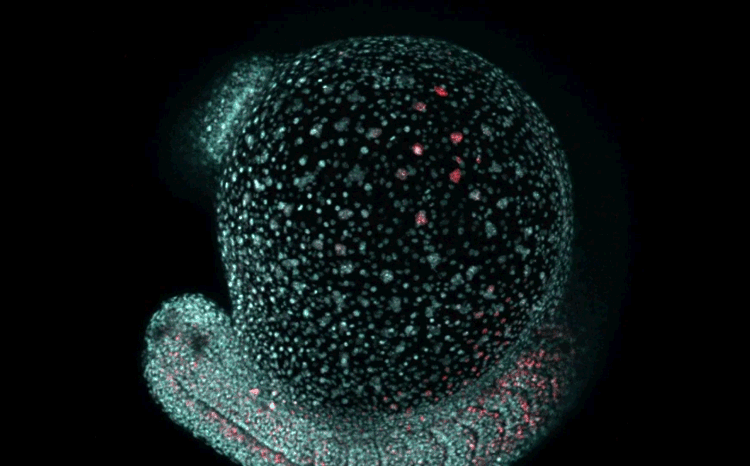
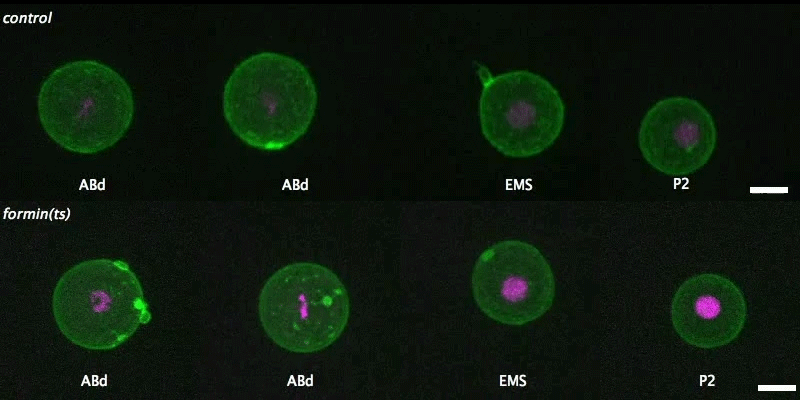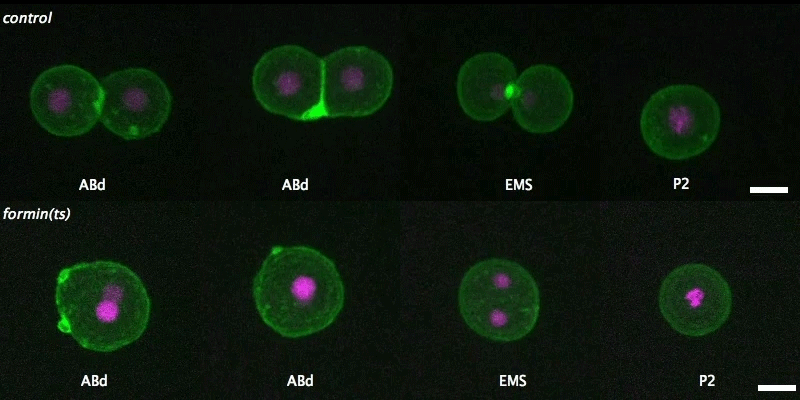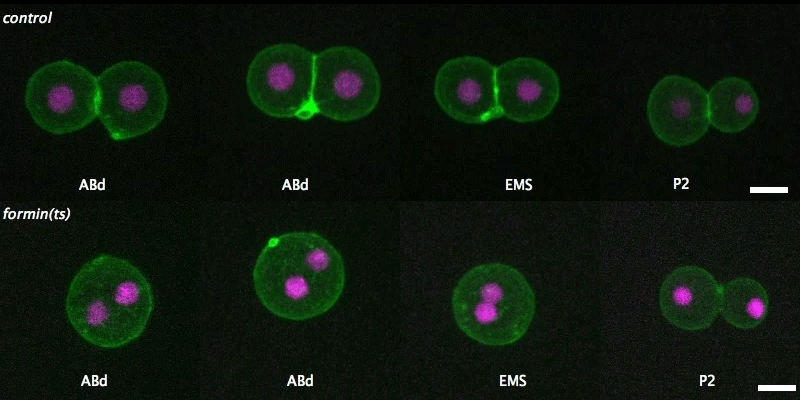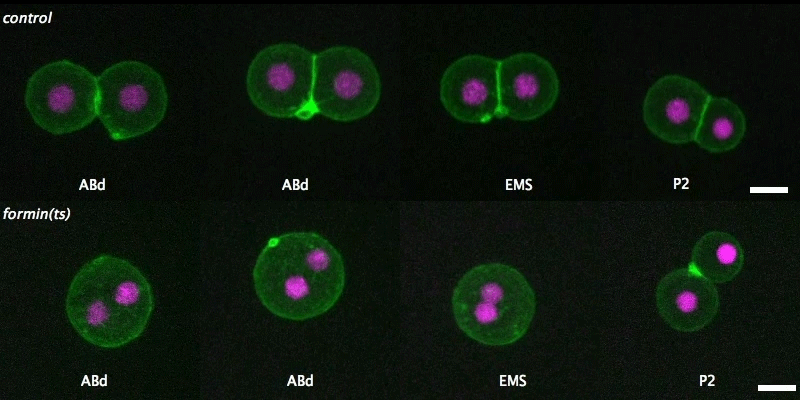
Over on Twitter we’ve been having fun with our third instalment of the 12 GIFs of Christmas. For those not on Twitter, here are the GIFs – they represent some of the most cutting edge and inventive developmental biology of 2018, and also showcase the beauty of timelapse microscopy.
Transcription overlaid onto the rapid cell divisions of the early Drosophila embryo.
Jeremy Dufourt, Antonio Trullo, Jennifer Hunter, Carola Fernandez, Jorge Lazaro, Matthieu Dejean, Lucas Morales, Saida Nait-Amer, Katharine N. Schulz, Melissa M. Harrison, Cyril Favard, Ovidiu Radulescu & Mounia Lagha
Andrea Attardi, Timothy Fulton, Maria Florescu, Gopi Shah, Leila Muresan, Martin O. Lenz, Courtney Lancaster, Jan Huisken, Alexander van Oudenaarden, Benjamin Steventon
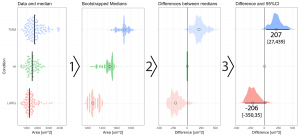
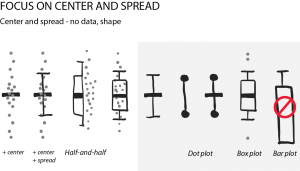
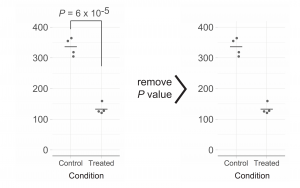
2018 was a fun year on the Node, with a continued diversity of posts, more jobs than ever and our highest number of readers since our launch (regularly breaking the 30k page views per month barrier). Good vibes, and a good time to celebrate our most-read from the year, which includes three posts on statistics and data visualisation by Joachim Goedhart, a cover competition and a group ‘op-ed’ about the advantages of preprints.
We gathered reactions from developmental biologists, reproductive biologists and ethicists to one of the year’s biggest and most controversial science stories
And if you haven’t, why not sign up to our email alerts – your weekly dose of developmental biology news, research highlights, interviews, meeting reports and jobs!
Shoshkes Carmel lab has an opening for a passionate postdoctoral fellow in the field of Telocytes as niche cells. We are seeking for a candidate with a strong mouse genetics background. We apply “-omics”, genetics and live-imaging approaches to uncover key aspects in the cell biology of Telocytes, large stromal cells recently emerged to constitute the intestinal stem cell niche, and their role in intestinal homeostasis.
For more details please see: Shoshkes-Carmel M et al., Subepithelial telocytes are an important source of Wnts that support intestinal crypts. Nature 2018 May 2.
Please apply via email including a cover letter with a short statement of research interests and Curriculum Vitae to:
Dr. Michal Shoshkes-Carmel
Dept. of Developmental Biology and Cancer Research
The Harvey Laboratory is looking to employ motivated and talented postdoctoral fellows to study the role of the Hippo pathway in organ size control and cancer. Our research is situated at both the Peter MacCallum Cancer Centre and Monash University in Melbourne, Australia.
We employ a range of techniques including Drosophila genetics, advanced microscopy, transcriptomics, bioinformatics, molecular biology and cancer cell biology. You will work in a supportive team, be a good communicator and also have the ability to work independently. You will possess expertise in a range of molecular and cell biology, biochemical and genetic techniques. Experience in Drosophila genetics is advantageous but not essential.
Peter MacCallum Cancer Centre is Australia’s largest specialist cancer centre with 600 research staff and students. We are located in Australia’s largest and most vibrant biomedical precinct with more than 10,000 researchers across multiple Universities, Research Institutes and Hospitals. Melbourne is a multi-cultural city with great food, weather, culture and sport and is often voted the world’s most liveable city.
For more information visit this link: https://www.petermac.org/research/labs/kieran-harvey
Poon et al., (2018). A Hippo-like signaling pathway controls tracheal morphogenesis in Drosophila melanogaster.Developmental Cell.47: 564-575.
Manning et al., (2018). Dynamic fluctuations in subcellular localization of the Hippo pathway effector Yorkie in vivo.Current Biology.28: 1651-1660.
Degoutin et al., (2013).Riquiqui and Minibrain, regulators of the Hippo pathway downstream of Dachsous.Nature Cell Biology.15: 1176-1185.
Harvey et al., (2013). The Hippo pathway and human cancer. Nature Reviews Cancer.13: 246-257.
Poon, et al.,(2011). The sterile 20-like kinase Tao-1 controls tissue growth by regulating the Salvador-Warts-Hippo pathway. Developmental Cell.21: 896-906.
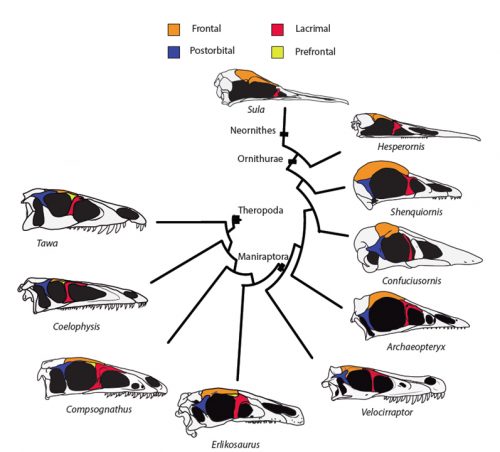
Birds are a dominant group of land Vertebrates (probably the largest in numbers with +10000 species described), highly successful and diverse. Birds originated from members of the Theropoda: the meat-eating dinosaurs that included famous forms like T. rex or Velociraptor, well-known from the movies. The fact that birds are a kind of dinosaur has been a matter of debate not except from controversy, but largely accepted today. The two most important lines of evidence that allowed us to understand the true identity of birds have been the (incredibly detailed) fossil record of the non-avian to avian dinosaur transition, and birds’ embryological development. Embryology reveals the dinosaur within birds and in cases development can parallel in short time what we see to have happened in millions of years of fossil record. Alexander Vargas’s lab at the University of Chile focuses on this evolutionary transition, employing both fossils and embryos to understand the workings of evolution.
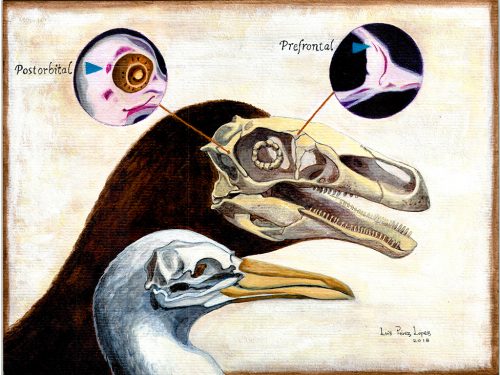
Researchers of comparative anatomy have documented how embryos show the dinosaur-bird link since the times of Darwin, mostly focusing on the postcranial skeleton. With this idea in mind, we set to study the development and evolution of the dinosaur skull. The head of birds is very unique, and two key differences when comparing birds to other reptiles are their skull and the huge brains and eyes birds possess. The bones composing the skull of birds are thin, light and mostly fused together in the adult. In addition, two bones (the postorbital and the prefrontal, located behind and in front of the eye respectively), were lost at different moments during the evolution of the dinosaurs leading to modern birds. Both in the fossil record and during embryonic time, how and when these bones disappeared, and if there were remnants of their presence as suggested by previous reports, were some of the questions we wanted to resolve in our recent paper in Nature Ecology and Evolution.
Fig.1: Traditional view of the evolution of the theropod skull and the loss of prefrontal and postorbital bones in the line leading to modern birds
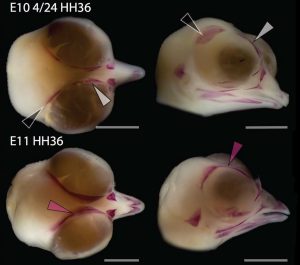
By looking at the fossil record of Theropods, we found two distinct “moments” when these bones were lost, at the origin of two recognizable clades: the postorbital was lost at the Euornithes node, close to modern birds; while the prefrontal was lost at the Pennaraptora node, right at the base of the clade containing dinosaurs like Oviraptor, Velociraptor, Archaeopteryx and modern birds. This bone has an interesting story, as in most early members of this clade, the prefrontal was not present, while the lacrimal bone acquired a T-shape (in contrast to the inverted L-shape of other dinosaurs that did possess a prefrontal). This T-shape, in which the T’s posterior tip occupies the position otherwise used by the prefrontal, suggested these bones were the result of fusion between the prefrontal and the lacrimal. The surprising part was that some dinosaurs seemed to be reverting into having a separate prefrontal bone. Some specimens of different species of these dinosaurs were showing a separate prefrontal bone, while at the same time losing that “tip” of the lacrimal that could have been the fused prefrontal. Even members of the same species would show variability in the presence or absence of the adult bone, as Deinonychus or Archaeopteryx specimens could have a prefrontal separated. To understand the basis of these changes in the cranium, we went to embryos.
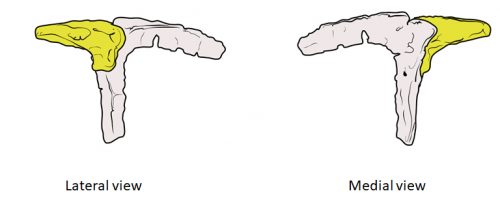
Fig. 2: In the specimen of Deinonychus MOR747, the lacrimal has an inverted-L shape and is accompanied by a separated prefrontal bone (yellow), just like in earlier theropod dinosaurs but unlike closely related species like Velociraptor.
Chicken embryos have been a preferred animal model for those willing to study embryonic development for more than 2000 years, and have provided the vast majority of the information we have today about skull development for birds. However, they are just one species among thousands, representing one of hundreds of distinct lineages. In an effort to include more of the disparity and diversity of birds, we collected embryonic series of six families of birds, including (of course) chicken, but also ducks, and species not normally used for developmental studies like lapwings, coots, budgerigar and tinamous (a member of the palaeognathae related to ostriches and emus), and also included embryonic stages of Alligator mississippiensis to broaden our sample to members of the two living archosaur groups, birds and crocodylians. By looking at the avian embryos and their developing ossifications alone, we confirmed the presence of more ossification centers than adult bones in the embryos of birds, but only by comparing them with alligator embryos and the fossil record were we able to interpret them in light of their evolutionary history.
Fig. 3: Authors in the hunt of red-gartered coot nests, with permission of the Agriculture Service of Chile.
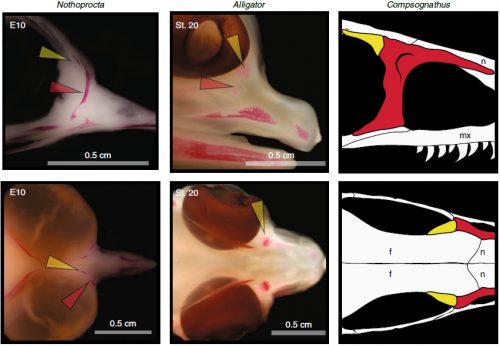
In embryos of all the birds observed, two ossification centers develop and make up the adult lacrimal bone. These two ossifications were observed and identified as the lacrimal and prefrontal bones of chicken by Erdmann in 1940, but we also found two separate ossifications giving rise to the lacrimal of alligator embryos, which do form a separate prefrontal from another ossification center, meaning Erdmann’s identification of a prefrontal bone in the chick was mistaken. However, we did find an embryonic bone in one species, the Chilean tinamou, where in addition to the two ossification centers of the lacrimal, a third, well-developed ossification develops in a position similar to that of the embryonic Alligator and adult dinosaur prefrontal. This bone, however fused to the nasal bone, and becomes indistinguishable from its other pieces. In a way, the bone is doing what we think it did in dinosaurs like Velociraptor: developing as a separate center and later fusing to a neighboring bone. This separate embryonic origin can explain why it re-appeared in different specimens as a separate adult bone, as it likely failed to fuse. However, in the tinamou, it is doing something it did not do in Velociraptor as instead of the lacrimal, it is fusing to a different neighbor.
Fig. 4: The prefrontal ossification of archosaur embryos (tinamou and alligator) and the adult bone in dinosaurs. Taken from Figure 1 of the original paper.
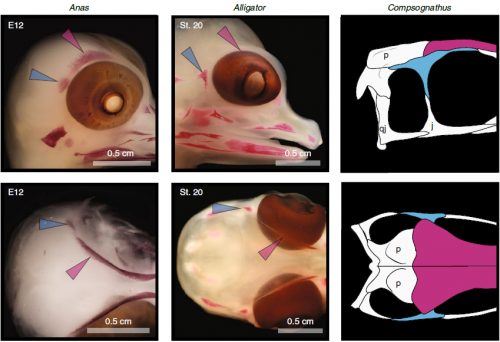
We also found an “extra” ossification not corresponding to any adult avian bone and just behind the eye that looks like the embryonic postorbital ossification of Alligator and the adult postorbital bone of dinosaurs, but instead of remaining separate as in these animals, fuses to the back of the frontal bone.
Fig. 5: The postorbital ossification of archosaur embryos (duck and alligator) and the adult bone in dinosaurs. Taken from Figure 2 of the original paper.
In a way, we were expecting to find this ossification, since the frontal of birds has been described as being formed from two separate portions, derived of two distinct embryonic germ layers; the mesoderm and the neural crest. Bones in the skull of vertebrates come in two flavors, as the most-rostral ones derive from neural crest cells and the ones in the back of the skull come from the mesoderm. We have known of this for years, thanks to careful chick-quail chimera experiments, and the double origin of the frontal has in fact created a lot of debate on the frontal identity itself. In other vertebrates, bones usually derive from one of these embryonic sources, and the frontal in particular is made up cells of neural crest origin. While other researchers also proposed the mesodermal portion could correspond to a separate bone that ended up fusing to the frontal, the identification of this portion as the parietal bone did not agree with many morphological criteria, anatomical correspondences nor with the evidence presented by the fossils. Our suggestion that the back portion of the avian frontal comes from the ossification that gives rise to the postorbital in other reptiles is consistent with the compared embryology of birds and crocodylians, as well as with the fossil record. This proposed homology implies the postorbital of Alligator should derive from mesoderm cells, something not yet corroborated as fate mapping of crocodilian embryos has not yet been done.
Fig. 6: The postorbital and frontal ossifications of chicken make up the adult frontal bone. Taken from Supplementary figure 4 of the original paper
The history of transformations and fusions of the prefrontal and postorbital provides us with some interesting lessons on how evolution takes place, but also leaves us with a number of intriguing questions of how skull development is regulated. Upon fusing with a neighboring bone, both the prefrontal and postorbital apparently lost their own identities, becoming a non-independent part of the larger bone, not showing any morphological similarity to the separated bone of other species. Moreover, the cells that make up the independent ossification centers end up forming a structure in which there’s no trace or clue of their origin, even when coming from two very distinct embryonic sources as are the neural crest and mesoderm. Contrary to most of the bones in the body, which form a mold or cast of cartilage that is later replaced by bony tissue, many bones in the skull (particularly those of the face, skull roof, palate and jaws) develop directly into bone from mesenchymal condensations. We know a (staggering) lot more about how these cells invade the head and face or find their location than what we know about how the mesenchymal condensations are established and how they turn into individual bones. Between the migration of neural crest cells into the head, for example, and the onset of bone formation, there’s a gap in our understanding that has still to be bridged. Questions like what signals regulate the condensation mesenchyme, the beginning of bone formation or the spatial distribution of the ossification centers that form the bones are still poorly answered. Our knowledge on this kind of bone development derives from (and has been limited by) histological sections, which do not allow us to have a whole-picture of what’s going on in the whole head, and the study a few molecular markers that mostly label post-mesenchymatic condensation and pre-osteogenic stages of maturation. It is, in truth, by these limitations that our study relied only on bone staining to observe and compare the ossification patterns of different species. More understanding of the whole picture was not going to come from digging into deeper molecular mechanisms in the chicken, but from studying and comparing the development of more species, in a maybe simpler way, like Alizarin staining.
One interesting thing to consider is that one possible driver behind the huge modification of the skull in the evolution of birds might be the evolution of a big brain. Birds have enlarged brains compared to other reptiles and non-avian dinosaurs, and the enlargement of the brain would necessitate a re-structuring of the skull covering it. Other cases of evolution of big brains also result in evolution of the skull as a whole, and even loss of bones, maybe in a similar way than what we see in birds. In mammals, bones of mixed origin proved to be composites resulting from fusion of elements, including those supposedly lost long ago in the earliest mammalian lineage. Although these bones are located in the back of the skull, mammals also lost bones like the prefrontal or postorbital. The brain, being a huge structure lying directly underneath the developing skull-roof, can possibly influence mesenchymal condensations, bone deposition, ossification rate and overall timing and spatial development of the skull. How these structures interact and how they evolve in concert is yet to be studied and understood.
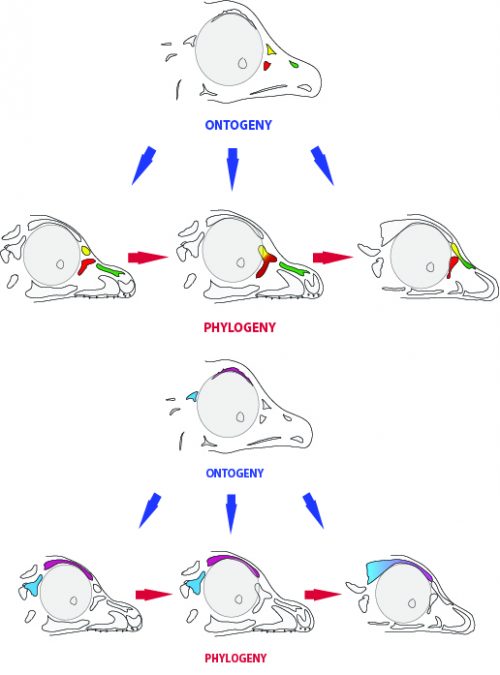
One last lesson we can derive from our results is the evolutionary meaning of the embryonic persistence of a structure. The ossification centers we found add to a longer list of stories in which the embryo retains at least a rudiment of a long-lost structure, which enables the evolution of new morphologies in later lineages. By retaining the prefrontal ossification, for example, some dinosaurs were (and still are) able to “experiment” a variety of morphological outcomes that include fusion with the lacrimal or nasal bones. It also worth noting that, at least in the case of the skull of birds, the adult bones end up fused together. So in a sense, the order or pattern in which ossification centers are beginning to fuse to each other could be less of adaptive significance than it is just a phenomenon of developmental drifting, in which a pattern is established and conserved. Itself, the high degree of bone fusion in the avian adult skull could be an extension of a trajectory started by those dinosaurs in which first two bones, then two others began fusing.
Fig. 7: Ontogeny and phylogeny. Possible developmental outcomes can be possible adult phenotypes of different lineages.
How does evolutionary change take place as shown by the fossil record and during embryonic time can be a question not just of “how dinosaurs looked like” but also an important source of understanding of developmental mechanisms at play every day.
Artwork by Luis Pérez López [CC BY 4.0]
Erdmann, K. (1940). Zur Entwicklungsgeschichte der Knochen im Schädel des Huhnes bis zum Zeitpunkt des Ausschlüpfens aus dem Ei. Zeitschrift für Morphologie und Ökologie der Tiere, 36(3), 315-400.
Le Lièvre, C. S. (1978). Participation of neural crest-derived cells in the genesis of the skull in birds. Development, 47(1), 17-37.
Noden, D. M. (1983). The role of the neural crest in patterning of avian cranial skeletal, connective, and muscle tissues. Developmental biology, 96(1), 144-165.
Evans, D. J., & Noden, D. M. (2006). Spatial relations between avian craniofacial neural crest and paraxial mesoderm cells. Developmental dynamics: an official publication of the American Association of Anatomists, 235(5), 1310-1325.
Piekarski, N., Gross, J. B., & Hanken, J. (2014). Evolutionary innovation and conservation in the embryonic derivation of the vertebrate skull. Nature communications, 5, 5661.
Maddin, H. C., Piekarski, N., Sefton, E. M., & Hanken, J. (2016). Homology of the cranial vault in birds: new insights based on embryonic fate-mapping and character analysis. Royal Society open science, 3(8), 160356.
Abzhanov, A., Rodda, S. J., McMahon, A. P., & Tabin, C. J. (2007). Regulation of skeletogenic differentiation in cranial dermal bone. Development, 134(17), 3133-3144.
Stockholm University, Sweden, invites applications for one postdoctoral position in the laboratory of Professor Mattias Mannervik at the Department of Molecular Biosciences, The Wenner-Gren Institute (http://www.su.se/mbw). The position is scheduled to start as soon as possible.
Transcriptional coregulators are proteins that facilitate communication between transcription factors and the basal transcription apparatus, in part by affecting chromatin through post-translational modification of histones. As such, they contribute to generation of cell-type specific gene regulatory networks and epigenetic control of animal development (see Mannervik et al. Science, 284, 606-609, Boija et al. Mol Cell, 68, 491-503). This laboratory is using genomic, genetic, and transgenic approaches in Drosophila melanogaster to elucidate the molecular mechanisms of transcriptional and chromatin regulator function during development. In this project two approaches are used to investigate the in vivo role of histone modifications in gene expression. A histone replacement system is used to examine the effects of amino acid substitutions in the histones on organismal development, and a modified CRISPR/Cas9 system is employed to target chromatin modifying enzymes to endogenous loci.
The position is available immediately and requires a recent Ph.D. as well as extensive experience in molecular biology techniques. The successful applicant should have a high-quality publication record, and motivation to study underlying mechanisms of gene regulation in development. The position will be funded with a fellowship, and includes health insurance.
Stockholm University is one of the largest and most prominent universities in Sweden, located in the nation’s capital city, beautifully surrounded by the first national city park in the world. For further information, see http://www.su.se/english/ and http://www.academicstockholm.se/
Application: Applications marked with reference number SU 465-0156-18 should be submitted electronically as a single PDF file to mattias.mannervik@su.se and to birgitta.olsson@su.se
The application deadline is February 1, 2019.
Applications should comprise the following:
1) a personal statement describing your interest in this project (1-2 paragraphs), research experience (1–2 paragraphs) and career goals (1-2 paragraphs)
2) curriculum vitae
3) bibliography
4) names, e-mail adresses, and phone numbers of three references
Established by the British Society for Developmental Biology in 2014, The Gurdon/The Company of Biologists Summer Studentship scheme provides financial support to allow highly motivated undergraduate students an opportunity to engage in practical research during their summer vacation. Each year, ten successful applicants spend eight weeks in the research laboratories of their choices, and the feedback we receive is outstanding. You can read accounts from previous years here.
Our final report from the 2018 group of student awardees comes from Rachel Wong (student at Queen’s University Belfast) who undertook her research with Karen Liu(King’s College, London).
Analysis of Rapgerf5 and canonical Wnt signalling in embryonic mouse development
During the summer of 2018, I worked with Dr John Griffin in Dr Karen Liu’s lab at King’s College London. My focus was on the gene RAPGEF5, which was previously identified as a candidate gene for heterotaxy, a congenital disease affecting heart development and the spatial arrangement of organs. It is estimated that 1 in 10,000 people are diagnosed with heterotaxy, and is the cause of 3% of all congenital heart cases [1]. However, the genetics of heterotaxy are still unclear. Thus research is necessary to understand the disease mechanism in more detail.
Not much is known about RAPGEF5 protein, but we know it is involved in the canonical Wnt pathway, in the transportation of beta-catenin into the nucleus [2]. When Wnt is active, a cascade of chemical reactions prevent the degradation of beta-catenin in the cytoplasm, allowing it to bind to a transporter protein to enter the nucleus. Our current model suggests that in response to Rap-GDP conversion to Rap-GTP by RAPGEF5, beta-catenin can dissociate from the transporter protein. This frees beta-catenin, allowing it to interact with DNA-binding proteins to alter gene expression. Therefore, my project aimed to answer three key questions:
Where is canonical Wnt signalling active during embryonic development?
Where is RAPGEF5 expressed during embryonic development?
Does loss of RAPGEF5 lead to any developmental abnormalities such as heterotaxy?
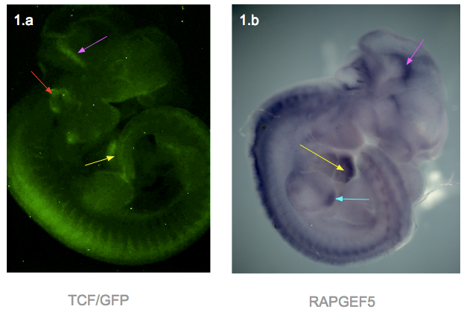
To answer my first question, I used genetically modified mouse embryos carrying a TCF/Lef-dependent reporter to visualize the areas with active Wnt signalling. When Wnt is active, the transcription factors TCF/Lef are active and bind to specific binding sites on DNA. This activates a promoter that causes the GFP reporter gene to be expressed and produce proteins that fluoresce under a specific wavelength.
I dissected embryos from their sacs at weeks E11.5, E12.5, E13.5 and E14.5. They were then photographed using a fluorescence microscope. In general, fluorescence can be seen in the ears, edges of limbs, spine, branchial arches, whiskers and brain. The heart was only visible in the E11.5, as the skin over the heart was too thick at later stages (Figure 1.a). At week E14.5, fluorescence was only faintly visible at the ear, due to the thicker skin.
In response to question 2, I fixed and dehydrated the E11.5 embryos for whole mount mRNA in situ hybridization. A Rapgef5-specific probe was used to stain the embryo, and the result was shown in figure 1.b. RAPGEF5 mRNA was expressed in the heart, brain, spine and the tip of the hind limb.
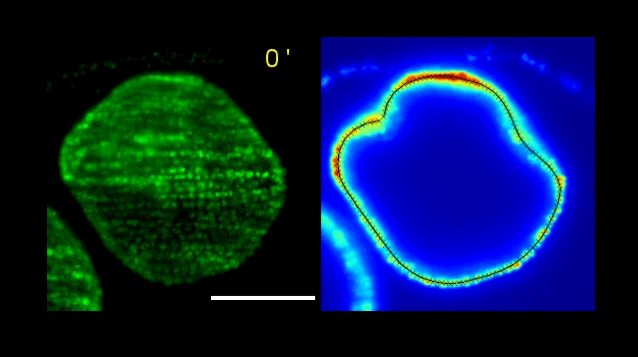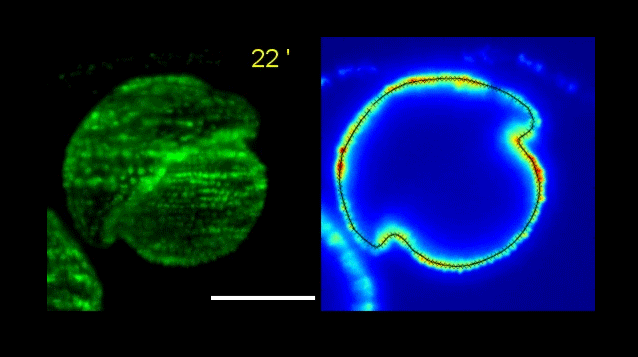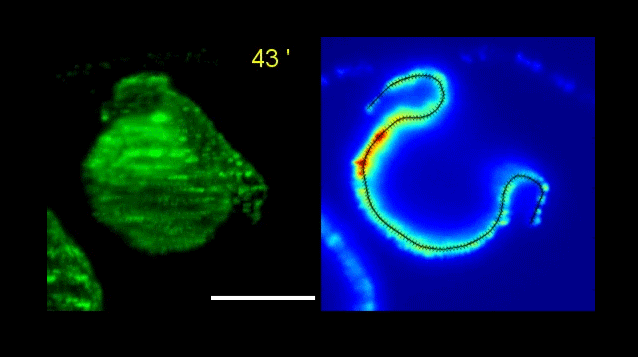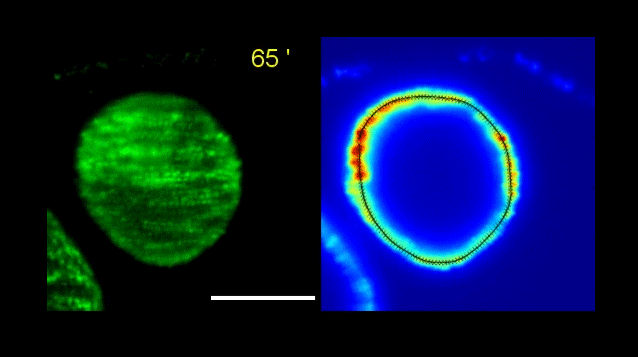
Figure 1 – Both photographs are of a TCF/Lef E11.5 mouse embryo. The left shows GFP signal reporting active Wnt., and the right shows the whole mount in situ hybridization with Rapgef5 probe Structures highlighted are the heart (yellow arrows), mid brain (pink), ear (red), tip of hind limb (blue).
Figure 1 presents two photographs of an E11.5 embryo, one showing the distribution of RAPGEF5 specifically, and the other for active Wnt signalling. When compared, the distribution appears similar, however there are discrepancies such as the ear, and tip of the hind limb.
Finally, for my third question, we bred RAPGEF5 mutant mice and inspected them at stages E9.5, E10.5, E14.5 and 6 weeks after birth. Tail clippings were taken from the embryos and ear clippings from the pups for DNA extraction and PCR to confirm their genotype, as they could either be wild-type, heterozygous or homozygous. Unfortunately the genotyping was still in the stages of trial-and-error, as the bands in the gel electrophoresis did not match the reference DNA ladder, and further tweaking with the PCR temperature and primers is necessary.
However, there were some phenotypic changes found. Out of the total of 9 E9.5 amniotic sacs, 4 were had healthy embryos, 4 were empty and 1 was malformed and underdeveloped. It is possible that this embryo was in the process of being reabsorbed to match the 4 other empty sacs. For the E10.5, there were 8 embryos in total, and 2 had underdeveloped heads and lacked proper surface morphology. All 8 of the E14.5 embryos were phenotypically normal.
Figure 2
Interestingly, as seen in figure 2, when compared with the wild-type, the 6 week old pup had bald patches in a ‘Christmas tree’ pattern and a possible front limb deformity. Both were similar in size and behaviour.
I thoroughly enjoyed my summer studentship at King’s, and learnt many new techniques such as wax sectioning and mounting, in situ hybridisation and using fluorescence microscopy. I would like to thank Mr John Griffin, Dr Karen Liu and the Liu lab for taking time out of their schedule for their help and guidance.
 (No Ratings Yet)
(No Ratings Yet)






 (4 votes)
(4 votes)