In this quarterly webinar series, hosted by Pablo J. Sáez and Valeria Venturini, we aim to showcase the latest research in the field, and we’d love to hear from researchers (especially early-career researchers) who would like to present their work. You can get in touch with us at focalplane@biologists.com or fill in our short application form.
Development, host of the Node, invites you to submit your latest research to our upcoming Special Issue – Plant and Algae Development. This issue will be coordinated by Academic Editor Dominique Bergmann (Stanford University, USA) and Guest Editor Susana Coelho (Max Planck Institute for Biology, Tübingen, Germany).
Developmental biology aims to understand how a single cell, the zygote, becomes a complex multicellular organism with specialised cell types organised into functional units. Derived from unicellular ancestors, algae, plants and animals use shared developmental principles, such as biophysical interactions, signalling, patterning and cell fate determination, to innovate and overcome obstacles for multicellular development, such as coordinating growth. The highly plastic development of photosynthetic organisms have long offered specific opportunities to answer questions regarding the origins of multicellularity, stem cell maintenance and regeneration, as well as environmental and microbial interactions and integration. In recent years, advances in genomics, imaging and synthetic biology are revealing how gene regulatory networks, mechanical biology and signalling interact to shape form and function in plants and algae. Furthermore, mathematical, computational and modelling approaches combined with traditional experimental biology have revealed, explained and predicted quantifiable properties behind phenotypes and across scales. We also see an exciting range of research organisms in use today, including species of algae, moss, ferns, flowering plants and others. In this special issue, we aim to showcase the contribution of plants and multicellular algae to our understanding of development, regeneration and evolution, highlighting quality research across the entire breadth of developmental biology.
The deadline for submitting research papers is 2 November 2026.
Note: it appears that some people are having issues with google forms, apologies if this has affected you. If you cannot use the form, please email your entry to us (see below for what is needed) and we will add it for you!
Looking forward to seeing all your images!
Many thanks,
Timothy
On behalf of YEN committee
Competition details:
Submit an original image you have generated related to developmental or stem biology. This may be a microscopy image, an image analysis output, or an artwork inspired by your scientific work.
Provide a title and a short description of the image, explaining the scientific context and techniques used (e.g. microscopy method or analysis strategy).
Provide your full name and affiliation. This information will be used for our internal records and future acknowledgement, but will not be displayed during voting.
You can view examples from the YEN 2025 Image Competition here.
Terms and Conditions
By sending us an image for the YEN Image Competition, you agree to the YEN Committee displaying the image publicly (including on our website and social media) and using it on YEN materials to promote future events. We will always credit you unless you request otherwise.
All submissions will undergo an initial review by the YEN Committee based on quality and visual impact. Applicants will be notified of the outcome of this selection through the email address provided in this form.
I was fortunate to receive an EMBRC-supported grant to carry out a small project on calcium imaging in sea urchin larvae at the Institut de la Mer de Villefranche (IMEV).
What makes IMEV special is not only its location on the French Riviera, but also the scientific history embedded in the place itself. Since the late 19th century, researchers have come to Villefranche-sur-Mer to study marine embryos, fertilization, plankton, and development; taking advantage of the bay’s extraordinary biodiversity and direct access to living marine organisms. In many ways, modern marine developmental biology was shaped in places like this.
And you still feel that spirit today.
Although IMEV is relatively small, it is well equipped and highly efficient for experimental work (Fig. 1).
Fig. 1. Institut de la Mer de Villefranche (IMEV), Villefranche-sur-Mer, France.
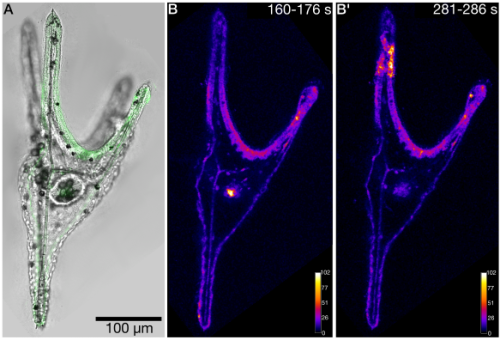
Access to marine organisms, imaging platforms, and technical support made it possible to rapidly test several GCaMP variants and explore live imaging approaches in sea urchin larvae (Fig. 2).
Fig. 2. Calcium imaging in the sea urchin larva Paracentrotus lividus expressing GCaMP6s. A. Brightfield image overlaid with GCaMP6s fluorescence signal (green). B – B’. MAX projections from selected time windows of an XYT recording, showing representative spontaneous calcium activity patterns. Fluorescence intensity is displayed using the Fire (LUT).
A key part of what made this project so productive was being hosted by Luis Bezares Calderon.
I had never met Luis before this visit. I knew his work on neuronal and behavioral mechanisms in Platynereis, so during the proposal stage I sent him a simple email asking whether he would consider hosting me at IMEV. He generously agreed.
From the beginning, he gave me full access to his lab resources and created a highly collaborative environment. But what shaped the experience most was not only the experimental support, it was the way he constantly challenged the thinking behind the experiments.
His questions were persistent and fundamental: Why this approach? How will it answer your question? Are you just going to look at traces forever, or understand their function?
Coming from a molecular biology background, I was used to thinking mainly in terms of genes, molecules, and cellular mechanisms. Working with Luis pushed me to think more directly about nervous system function and behavior. For the first time in my life as a researcher, I witnessed behavioral assays in sea urchin larvae with my own eyes while simultaneously trying to connect activity dynamics to biological function.
This connection between molecules, cells, neural activity, and behavior is something Luis actively builds into his science. I found that intellectually challenging, stimulating, and genuinely exciting.
This experience reminded me that marine stations are not just places to run experiments. They are environments where questions can evolve quickly because of access, collaboration, and the intensity of scientific interaction.
I am very grateful to everyone at IMEV, and especially to Luis, for making this possible.
“Why do some hearts regenerate, while others do not?”
This question has followed me for more than a decade. Not as a single project, but as a thread that kept resurfacing—each time forcing us to rethink what we thought we understood.
The idea of comparing regenerative zebrafish with non-regenerative medaka started as one of several proposals I discussed with Didier Stainier when I joined his lab in 2014. At the time, it felt simple: let biology provide the contrast, instead of trying to extract answers from a single system.
Zebrafish regenerate their hearts. Medaka do not. That difference was too striking to ignore (1).
I was in my second postdoc then, aware that I needed something I could carry forward. Didier encouraged me to pursue it, even though it was clearly high-risk. Looking back, that decision shaped everything that followed.
Didier visited IBMS at Academia Sinica in Taiwan in 2019 for an Institutional Lecture—a full-circle moment early in the journey.
When the data pointed to something unexpected
Together with Rubén Marín-Juez —who later became both a key collaborator and a close friend—we established a cryoinjury model in medaka and generated our first comparative RNA-seq datasets (2).
What we saw was not what we expected.
Instead of cardiomyocyte-centered differences, which at the time were widely viewed as the primary drivers of cardiac regeneration, the strongest signals pointed toward immune responses and angiogenesis (2). I remember hesitating. It felt like we were drifting away from what many would consider the “core” of cardiac regeneration.
But the data were clear, and we decided to follow it.
We used to joke that we were like the Maze Runners—moving forward without knowing what was coming next, or how things would end. Rubén focused on angiogenesis and uncovered how revascularization is an early and essential step in regeneration (3, 4). In parallel, I moved toward immunity—despite being warned, quite accurately, that “immunity is too complicated and messy to work with.”
In hindsight, that hesitation reflects something broader. Work in non-mammalian models is often judged by how directly it translates to human biology, rather than by the clarity of the biological principles it can reveal. Yet it is precisely these systems—and the people willing to pursue them—that allow us to uncover mechanisms that are otherwise difficult to see.
Rubén and I returned to Bad Nauheim, Germany, in 2025 for the 30th anniversary of the Stainier lab—revisiting the place where this journey first began.
A result we thought we understood—but didn’t
What stood out early was that macrophage infiltration appeared delayed and reduced in medaka (2). So, we asked a simple question: what happens if we delay macrophage recruitment in zebrafish?
Using clodrosome, we transiently depleted macrophages prior to injury and observed impaired regeneration.
At the time, we interpreted this primarily as a timing effect—an early delay that irreversibly disrupts regeneration.
This seemed to fit the data. Macrophages eventually came back, and their numbers recovered within about a week. Yet the heart still failed to regenerate.
Through temporal single-cell profiling—driven in large part by the careful and persistent work of Ke-Hsuan Wei, one of the first PhD students in my lab—we realized something we had completely missed before: clodrosome was not simply delaying macrophages—it was preferentially depleting the resident macrophage population (5).
These cardiac resident-like macrophage subsets turned out to be essential for heart regeneration—coordinating revascularization, cardiomyocyte survival, debris clearance, and extracellular matrix remodeling.
That realization reframed everything.
Even when we allowed extended recovery, giving circulating, monocyte-derived macrophages ample time to repopulate the heart, regeneration did not recover.
At that moment, the entire story finally made sense.
It was not only about timing. It was also about identity.
What initially appeared to be a delay in macrophage function was, in fact, the loss of a specific and irreplaceable cell population.
Turning a non-regenerative system on
We then asked the opposite question: instead of suppressing the immune response, could activating it change the outcome?
Poly I:C—identified through comparative transcriptomics—enhanced macrophage recruitment and, unexpectedly, enabled de novo regeneration in medaka (2).
That was one of those moments when you don’t immediately trust the data. We repeated the experiments, trying to convince ourselves it wasn’t an artifact.
But it held.
Regenerative capacity began to look less like a fixed property and more like something that could be modulated.
From immune identity to regenerative signal
Our recent paper in PNAS represents the latest step in this progression (6).
Led by Kaushik Chowdhury, this phase of the work brought together comparative analysis, single-cell profiling, and functional experiments to identify a regeneration-associated macrophage population induced by poly I:C.
These macrophages localize to the injury border zone and express Granulin.
What started as a candidate marker became a functional insight. Through a series of carefully executed experiments, the team showed that recombinant Granulin alone is sufficient to promote cardiomyocyte proliferation and reduce scarring—linking immune activation to a concrete regenerative outcome.
Closing the loop, granulin expression is also activated in zebrafish following cardiac injury and is essential for heart regeneration.
Basic science, unexpected translation
I always consider myself a basic scientist and a developmental biologist. None of this work started with a translational goal.
It was driven by curiosity—by a question that seemed fundamental, but not immediately “useful” or “applicable”.
And yet, it led us to a concept that is inherently translational: that regeneration might be induced through immune modulation.
As Didier once put it:
“One never really knows when a basic science finding will transform translational research… CRISPR/Cas9 is just one recent example.”(7)
That perspective has stayed with me throughout this journey. Especially at moments when the work felt uncertain, or when its relevance was not immediately obvious.
Still the same question
If there is one idea that has gradually emerged, it is that regenerative capacity is not fixed.
It is governed, at least in part, by the identity and function of immune cells—and therefore potentially modifiable.
That doesn’t make the problem simple. But it reframes it.
And in many ways, we are still following the same question.
Just with a clearer understanding of what actually matters—and with a team that made it possible to see it.
A glimpse of the team behind the work—reminding us that every figure is built on many shared moments that never make it into the paper. Kaushik (back row, fourth from the left) and Ke-Hsuan (fifth from the right).
References
1. Ito K, Morioka M, Kimura S, Tasaki M, Inohaya K, Kudo A. Differential reparative phenotypes between zebrafish and medaka after cardiac injury. Developmental Dynamics. 2014;243(9):1106-15.
2. Lai S-L, Marín-Juez R, Moura PL, Kuenne C, Lai JKH, Tsedeke AT, et al. Reciprocal analyses in zebrafish and medaka reveal that harnessing the immune response promotes cardiac regeneration. eLife. 2017;6:e25605.
3. Marin-Juez R, Marass M, Gauvrit S, Rossi A, Lai S-L, Materna SC, et al. Fast revascularization of the injured area is essential to support zebrafish heart regeneration. Proceedings of the National Academy of Sciences. 2016;113(40):11237-42.
4. Marín-Juez R, El-Sammak H, Helker CSM, Kamezaki A, Mullapuli ST, Bibli S-I, et al. Coronary Revascularization During Heart Regeneration Is Regulated by Epicardial and Endocardial Cues and Forms a Scaffold for Cardiomyocyte Repopulation. Developmental Cell. 2019;51(4):503-15.e4.
5. Wei K-H, Lin IT, Chowdhury K, Lim KL, Liu K-T, Ko T-M, et al. Comparative single-cell profiling reveals distinct cardiac resident macrophages essential for zebrafish heart regeneration. eLife. 2023;12:e84679.
6. Chowdhury K, Huang C-L, Lin IT, Hung Y-J, Lim KL, Liu H-W, et al. Immune modulation promotes heart regeneration through macrophage and Granulin functions in medaka. Proceedings of the National Academy of Sciences. 2026;123(16):e2524705123.
7. Grewal S. An interview with Didier Stainier. Development. 2015;142(17):2861-3.
From the helpimascientist.com archive, an in-depth discussion covering the many purposes of laboratory group meetings and why you should care about all of them.
The Marín-Juez laboratory, at the CHU Sainte-Justine Research Center, is recruiting PhD students and postdoctoral fellows (up to 5 years fully funded position). Our laboratory is interested in the cellular and molecular mechanisms regulating cardiac regeneration and development.
The successful applicant will join the Marín-Juez laboratory at the CHU Sainte-Justine Research Center, where they will have access to state-of-the-art facilities and technology platforms including Advanced imaging platform (light-sheet, spinning-disc confocal, multiphoton, STED super-resolution, etc.), genomics (DropSeq, 10x, Illumina Novaseq, Visium), IPSC Cell Reprograming and bioinformatics platforms. The CHU Sainte-Justine Research Center provides a thriving scientific environment where the successful applicant will have the opportunity to work with multidisciplinary scientific teams and to collaborate with talented clinicians and researchers.
Research project description
We have previously uncovered mechanisms governing coronary network replenishment, including the formation of a vascular scaffold that supports cardiomyocyte regeneration and mediates coronary-epicardial interactions and immune responses (Marín-Juez et al., PNAS 2016; Marín-Juez et al., Dev Cell 2019; El-Sammak et al., Circ Res 2022; Wang et al., Development 2024; Gupta et al. Dev Bio 2025; Rouf et al. Development 2026). Our recent work identifies the epicardium as a master regulator of cardiac fibrosis resolution and tissue replenishment (Kayman-Kürekçi et al. NCVR2026).
Building on these findings, we now aim to elucidate how the cardiac endothelium, epicardium, and immune system components cooperate to regulate tissue replenishment, as well as the specific mechanisms underlying their roles in cardiomyocyte regeneration and development.
PhD student position
Applicants should have training in molecular biology, cell biology, or related fields. Candidates should be enthusiastic about regenerative and developmental biology. Previous research experience with zebrafish and/or heart regeneration is highly valued but not essential. Candidates with experience in confocal/light-sheet imaging and/or genome engineering are strongly encouraged to apply.
Postdoc position
We are looking for candidates with a Ph.D. in the biological sciences and laboratory experience in tissue repair/regeneration, cellular, molecular biology, or genetics. Previous experience working with zebrafish, imaging and histology are highly valued but not essential. Candidates with experience in confocal/light-sheet imaging and/or genome engineering are strongly encouraged to apply. Preference will be given to applicants with excellent collaborative and communication skills.
How to apply
Candidates must send the required documentsto Rubén Marín Juez at ruben.marin.juez.hsj@ssss.gouv.qc.ca
The Ruiz lab is offering fully funded postdoctoral positions up to five years in the Laboratory of Genome Integrity located at the National Institutes of Health (NIH, Bethesda, MD). NIH is the largest biomedical research agency in the world, fosters world-renowned researchers and provides access to state-of-the art innovative technologies and scientific resources.
Our laboratory uses human and mouse embryonic stem cells (ESCs) as well as mouse embryos to understand the molecular mechanisms underlying cell fate decisions. The applicant should have or be about to have a PhD in Developmental Biology, Genetics, Molecular Biology or similar, and must have demonstrated expertise in mouse early embryology (mouse pre-implantation embryo isolation and in vitro mouse embryo culture). Expertise in embryo microinjection and manipulation will be considered an advantage. In addition, molecular biology/mammalian cell culture (preferably in embryonic stem cells) and knowledge of next generation sequencing technologies will also be relevant for the position.
The applicant will be involved in a variety of exciting projects studying cell plasticity/totipotency to explore the underlying mechanisms of new regulators of Zygotic Genome Activation. We seek highly motivated, creative individuals, eager to learn and develop new technologies and complex cell systems based on live cell/embryo imaging, single-cell technologies and CRISPR-based editing, interested in understanding how a single cell can develop into a complex multicellular organism in vitro and in vivo.
Please send a brief cover letter and the names of three references via e-mail to:
So, recently I attended a developmental biology conference – my first one of 2026, with six more to go! While socialising and networking with a group of truly amazing stem cell researchers, many of them asked, after I introduced myself as a Reviews Editor at Development, “What exactly does a Reviews Editor do?” After answering this question at least five times – across scientists at different career and life stages – I realised it might be time to share with you all what we ‘cool kids’ actually do.
Normally, at The Company of Biologists (Development’s publisher), we follow a hybrid working model, which means that we work from home for half of the week (which is usually 2-3 days a week for me) and the rest in the office. If you haven’t seen a photo of our office building yet, it is a rather beautiful building, combining the charm of a cottage-style exterior (complete with hipped roofs and classic sash windows) with a bright, modern and open-style office inside.
At Development, Ingrid Tsang and I are the Reviews Editors and we mainly handle the journal’s front-section content (so, that includes Reviews, Spotlights, Perspectives, Hypotheses, Primers, interviews – yeah, we have an extensive list!) and we work closely with Alex Eve, Executive Editor of Development.
Image shows a Reviews Editor, handling several deadline-oriented projects. Generated with CoPilot.
I normally start my workday between 9:30-10 am, and my first task is always to reply to emails while sipping my morning coffee (a strong flat white if I am home and a long cappuccino when in the office).
After the first half an hour to attending to emails regarding submissions, chasing authors for their submissions, or finishing off a pending task from the previous day (which often involves taking a final read through a decision letter), I move on to the main tasks of the day.
If I am working on a chunky edit – meaning a developmental edit of a Review article – I would usually block off an entire day for it (at least 7–8 hours). This typically happens once a Review-type article (which we commission in-house and invite authors to submit) has gone through peer review. At that stage, we, the in-house Reviews Editors, read through the full manuscript in detail, commenting on scientific accuracy, language and structure, conciseness and accessibility, journal style, article length and references – all while helping authors address the Reviewers’ comments more effectively. I also go through the figures and legends (we take our display items very seriously, as a single figure often speaks a thousand words), commenting on visual appeal, labelling and other finer details. Developmentally editing an article is usually the most rigorous part of the job, at least in my opinion, as it ensures that the final piece is not only of high quality but also forward-looking and engaging for our wide readership. At Development, we pride ourselves on being extremely hands-on when guiding authors and helping them address both our feedback and the Reviewers’ comments, in order to publish the best possible version of their review.
Another crucial part of our job is commissioning. We have our in-house commissioning meetings every two weeks. So, if you catch me the afternoon before, I am usually frantically reading articles on a certain topic of interest, trying to prepare somewhat cohesive pitches to discuss with the rest of the team. We mainly invite authors to write peer-reviewed, review-type content for us. To identify emerging topics in the field, we attend important conferences, chat with researchers across a wide range of developmental biology disciplines, keep an eye on their websites, analyse research trends across primary research articles and participate in extensive 1-1.5-hour long commissioning meetings.
Once we have agreed on a topic and a suitable author, we invite them to write for us. If they accept our invitation, the author will then often involve their students and collaborators as co-authors, and at that point we discuss the potential scope and type of the article. Of course, we also have a thorough in-house pipeline that monitors the status of all articles from invitation through to acceptance.
When we’re in the office, Alex, Ingrid, Andrea (Community Manager of the Node) and I often chat about various aspects of the job throughout the day – both formally and informally – because our work requires teamwork and collaboration. So, Ingrid and I will often discuss scheduling to make sure our publication pipeline stays tight (i.e. that every Issue publishes a few front-section content). If we have just returned from a conference, we chat with Alex and Andrea about emerging research trends and potential blogs/ posts for the community site. We also bounce around ideas for commissioning topics and share feedback on each other’s pitches, amidst a healthy dose of random life chats.
For me, the day usually ends with a quick catch-up on plans for the next day. This is also when I respond to any remaining email replies from the morning. I usually like to do a final run-through of my to-do list, ticking things off and marking any pending tasks (if there are any). And with that, I sign off for the day!
It’s been just over 8 months since I finished my PhD and posted my introduction as Development’s newest Reviews Editor on the Node. In this time, a constant question I’ve been asked by the ECRs around me from both my past life in academia and my current life at conferences has been “What do you do now? What does your day look like?”.
Of course, it’s not just me. Saanjbati – my partner in crime on the Reviews Editors team – has also been fielding these questions since she started this job as well. So, given the appetite from ECRs in hearing about our jobs, we’re lifting the lid on the elusive title of ‘Reviews Editor’ to show you what really goes on behind the scenes to deliver Development’s review articles and other front section content. To kick things off, Saanjbati has an article on “What does a Reviews Editor do?” and I am providing a run-down here of one sunny (!) day in April 2026, randomly chosen by my prettiest d20 die for your perusal.
It’s probably worth noting here that, as with most jobs, each day in the journal’s office is very different to the next. So do let me know if you would like to see another day in my life. But for now, hope you enjoy reading about this one!
Microsoft 365 Co-Pilot’s graphical interpretation of myday.
Whilst I am writing about my experiences as a Reviews Editor at Development, all views here are my own and do not represent the journal.
——
Cambridge, England – 2026 One of April’s many Mondays.
10:00 – Get to work, catch up with the office, grab a coffee. Our core office hours are 10am till 2pm so, as a night owl, I take full advantage of this and start at 10.
10:05 – Check emails that have come in through the weekend, as well as the various reports and notification we automatically receive from our online submissions system. An author who I am really excited about has agreed to write a review for us – whoop! I respond immediately and get stuck into clearing my inbox, which seems to be perennially full no matter how much I try to empty it.
10:30 – Soreen* break! Have a quick chat with the lovely preLights Community Manager, Reinier, about possible exciting preprints to highlight between bites of sticky sugary goodness. I promise to also upload the preprints list I collated at BSDB a few weeks ago. But for now, back to emails.
11:30 – Just received a message from Saanjbati about moving an article around between scheduled issues, so we have a quick chat about this. Then back to working through my inbox.
12:30 – Done with emails! Whew. A few commissioning emails have been sent out, feedback on synopsis given, reviewers chased, submission deadlines updated. Deep dives into synthetic biology and photoreceptors surfaced from. Just a little bit more admin to go…
12:45 – … and we’re done! Time for lunch outside in some suprisingly good weather.
13:15 – After a little bit of sun (and a touch of wind), it’s back to the desk for me. This afternoon, I have a slightly overdue meeting report to send round. It’s been delayed because I lost all the notes I’d been taking on it when my computer decided to restart itself whilst I was at the conference dinner and disco. Obviously a massive shame, but it also pushes me to ruminate harder over my notes in an attempt to rescue them, which might lead me to find other ideas…? Or at least that’s what I tell myself.
13:30 – I’ve realised I need to book my hotel for a trip later this year, which I’ve already forgotten to do four times. I get this out of the way first before I forget a fifth. Makes me excited for EuroEvoDevo in Glasgow!
14:30 – Somehow got sidetracked into looking up biorxiv references made during BSDB, which prompted another deep dive into recent preprints published under ‘Developmental Biology’. Spend a few minutes in awe of how quickly research is moving in certain directions and the seemingly masses of interest in biophysical/quantitative biology. Send some ramblings to Reinier and feel mildly envious of his job.
14:40 – Back to work on my core responsibilities! I’m only halfway through thinking about the meeting report and doing follow-ups on it. But it’s already time to look through the list of articles that were accepted in the past week and think about which should be highlighted.
14:58 – Done, just in time for the research highlight (RH) meeting! This is our weekly meeting where we discuss all the back section (i.e. primary research) articles and decide which to highlight. There are so many interesting papers this week, it’ll be quite hard to choose and I’m excited for the discussion.
15:35 – RHs have been picked, and I’m back at my desk to do a final read-through of the paper I’ve been assigned, just to make sure it’s as interesting as it seemed from my initial read-through.
15:40 – An email from our production team has just come through about a review article I’ve handled. Spend 10 minutes on this. Then another message comes through regarding some travel admin. Another 10 minutes gone.
16:00 – Back to reading this week’s RH paper. I personally really like it and am even more excited to write the highlight, although some of it is feeling quite anatomically complicated and will be difficult to describe in just 200 words without an illustration… Ohh well, that’s a problem for the future.
16:15 – OK, the various miscellaneous items that came in are done. Time to really lock-in on the meeting report, which needs to be sent today. It’s a nice creative exercise to reflect on all the science discussed at the meeting, but quite stressful to go through the whole programme and all my notes and synthesise something coherent for the rest of the team when under a time pressure.
16:50 – My brain is fried, my fingers feel like they’re about to fall off and my spirits are in dire need of chocolate. But the meeting report has finally been sent off! Time to wrap up a few things. As always, some important emails trickle through right as I’m about to leave. I resolve to address them on my way home.
17:00 – I leave early on Mondays so rush out for my bus. Bye!
*(other brands of malt loaves exist etc. etc.)
——
Did you know that anyone can publish on the Node? If you’ve been inspired to write a piece for the developmental biology community, feel free to register an account and then make your own blog post here: https://thenode.biologists.com/wp-admin/post-new.php

 (No Ratings Yet)
(No Ratings Yet)



 (2 votes)
(2 votes)



