The Mammalian Genetics and Development Workshop is an annual meeting focusing on the development and genetics of mammals. The Meeting is based on the submitted abstracts, and include diverse topics ranging from mammalian development (not exclusively human or mouse) and identification of disease genes and developmental mechanisms, to human genetics and epigenetics. Other model systems (including Drosophila, zebrafish and chick) are also welcomed where these relate to general developmental questions and/or disease models.
The meeting will be similar format to the 28th Meeting in 2017, which had a fantastic set of short talks covering various aspects of developmental biology, mouse models of human disorders and genetics of human disease (abstracts are published at https://doi.org/10.1017/S0016672317000076)
The workshop is traditionally a venue for post-docs and PhD students to talk rather than laboratory heads and is an excellent training ground and a friendly, informal forum. In keeping with this objective, we offer TWO PRIZES of £150 to individual post-graduate/post-doctoral presenters. In addition, thanks to the generosity of Mammalian Genome (https://link.springer.com/journal/335) publishers, we will offer an additional PRIZE of £150.
Registration: A £10 registration fee is payable by all attendees on arrival at the meeting. This fee covers the abstract booklet, tea and coffee refreshments and the wine reception. Speakers and chairpersons will be provided with lunch, free of charge, on the day of their presentation.
Abstract Submission: All Workshop presentations will be in lecture format (15 or 20 minutes). Please send your abstract by e-mail (Word or rtf file) to ich.mgdwshop@ucl.ac.uk by 5pm on 8th November. In addition please indicate the length of talk you prefer.
Abstract format is as follows: Title (bold), AUTHORS (ALL CAPS), Address (italics) and text (200 word limit). Abstracts will be published (with the authors’ permission) in Genetics Research (https://www.cambridge.org/core/journals/genetics-research).
We have a vacancy for a postdoctoral research associate, funded by a BBSRC grant, to work on the roles of spastic paraplegia gene products in formation of axonal ER, and identification of new genes and cellular processes involved in this. For some background on the project, see: https://elifesciences.org/articles/23882.
Applicants should have a PhD in a biological science, interests in neuronal cell biology, and experience in as many as possible of the following: model organism genetics, genetic screens, neuronal cell biology, light or electron microscopy, live imaging, molecular biology, and bioinformatics.
The Academy of Medical Sciences is convening a scientific meeting on the 19 and 20 March 2019, in Oxford. Abstract submission deadline: 5 November 2018.
This meeting will explore key areas of neurodevelopmental research in a unique forum that emphasises discussion and collaboration between disciplines, career stages and sectors.
One of the key aims will be to highlight the latest advances in neurodevelopmental research and identify key research questions that could bring a real impact to the field. The meeting’s talks and discussions will span a broad range of neurodevelopmental and disorder research, from in utero to adolescence, and from multiple perspectives, including molecular, cellular, circuit/systems and behavioural levels.
We welcome researchers from all backgrounds to attend this meeting and strongly encourage early career researchers, from PhD level onwards, to apply. Registration is free.
Calculation and reporting of p-values is common in scientific publications and presentations (Cristea and Ioannidis, 2018). Usually, the p-value is calculated to decide whether two conditions, e.g. control and treatment, are different. Although a p-value can flag differences, it cannot quantify the difference itself (footnote 1). Therefore, p-values fail to answer a very relevant question: “How large is the difference between the conditions?” (Gardner and Altman, 1986; Cumming, 2014). The aforementioned question can be answered by calculating the “effect size”, which quantifies differences. Calculation and interpretation of effect sizes is straightforward and therefore a good alternative for calculating p-values (Ho, 2018). In the figure below, I transformed an ordinary plot with p-values into graphs that depict (i) the data and (ii) the difference between median values as the effect size. The calculation and application of effect sizes is the topic of this blog.
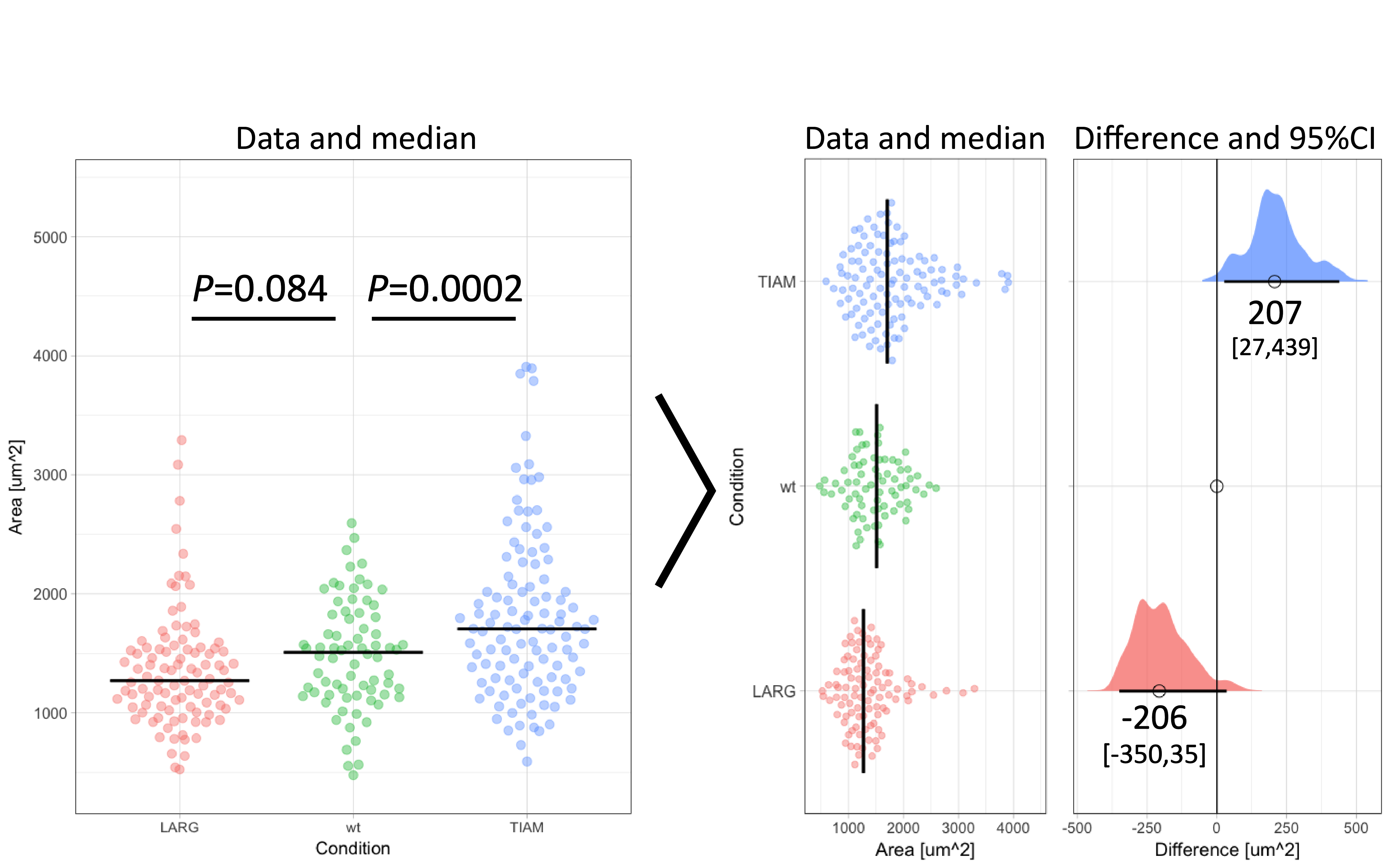
Figure 1: Transformation of an ordinary graph with p-values into a visualization of the data and the difference between median values as the effect size. Values indicate the difference between the median and the median of the ‘wt’ Condition. The horizontal lines and the values between brackets indicate the 95% confidence interval.
Why do we need an alternative for p-values?
The null-hypothesis significance test (NHST) compares the central tendency of two samples, e.g. the mean for parametric or median for nonparametric tests. Next to the central tendency (or location), it takes into account the precision of that value. Since the precision increases with increasing sample size, the outcome of a NHST depends on sample size (Nakagawa and Cuthill, 2007) as also explained here. In other words, the p-value reflects both the difference in central tendency and the number of observations. This is just one of the issues with p-values, other issues that should be considered when using p-values are discussed elsewhere (Goodman, 2008). Due to the issues with p-values, it has been recommended to replace (Cumming, 2014; Halsey et al., 2015) or supplement (Wasserstein and Lazar, 2016) p-values with alternative statistics. Effect sizes, aka estimation statistics, are a good alternative (Claridge-Chang and Assam, 2016).
Effect size
In statistics, the effect size reflects the difference between conditions. In contrast with p-values the effect size is independent of sample size. Effect sizes are expressed as relative (footnote 2) or absolute values. Here, I will only discuss the absolute effect size. The absolute effect size is the difference between central tendency of samples. Commonly used measures of central tendency are the mean and median for normal and skewed distributions respectively. Hence, for a normal distribution the effect size is the difference between the two means.
Quantification of differences
To illustrate the calculation and use of effect sizes, I will use data from an experiment in which the area of individual cells was measured under three conditions (Figure 1). There is an untreated (wt) condition and two treated samples. The question is how the treatment affects the cell area. When the distribution of the observed areas is non-normal (as is the case here), the median better reflects the central tendency (i.e. typical value) of the sample. Therefore, it is more adequate for this experiment to calculate the difference between the medians (Wilcox, 2010). The median values for TIAM, wt and LARG are 1705 µm², 1509 µm² and 1271 µm² respectively. From these numbers, it follows that the effect of TIAM is an increase of area by 1705 µm² – 1509 µm² = 196 µm². The effect of LARG is a decrease in cell area of 1271 µm² – 1509 µm² = -238 µm². Note that the difference is expressed in the same units (e.g. µm²) as the original measurement and is therefore easy to understand.
Confidence intervals
As mentioned before, the difference will not depend on sample size. On the other hand, the precision will depend on sample size. More observation per experimental condition will generate median values with a lower error. As a consequence, the difference will be more precisely defined with a larger sample size (Drummond and Tom, 2011). To indicate the precision of the difference, the 95% confidence interval (95CI) is a suitable measure (Cumming, 2014). For differences between means, the 95CI can be calculated directly (see here or here for calculation with R) but for the difference between medians it is not straightforward.
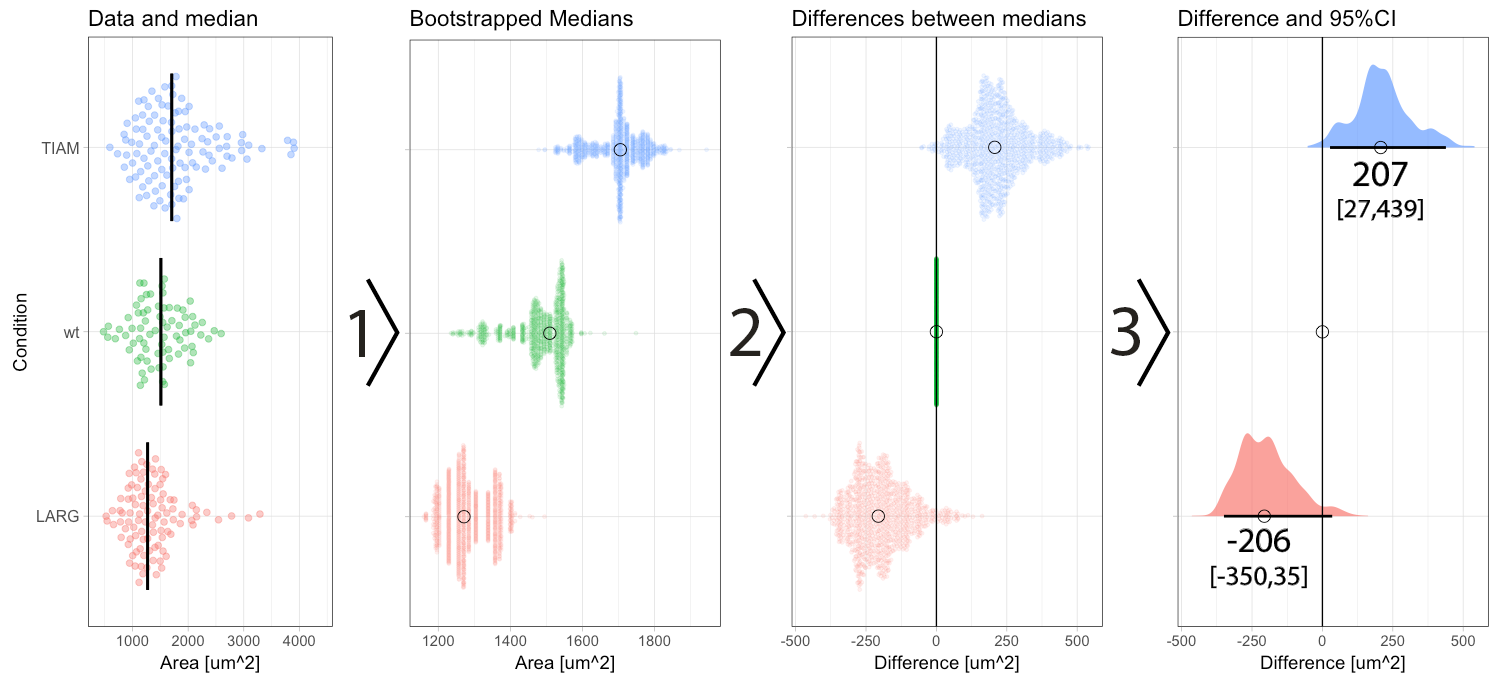
Figure 2: Step-by-step explanation of the calculation and visualization of the effect size and 95% confidence intervals. Step 1) bootstrapping of the data results in a collection of 1000 new median values. Step 2) the difference between the conditions (LARG and TIAM) and the control (wt) is calculated by subtraction, yielding 1000 differences (or effect sizes). Step 3) the middle value (median) of the distribution defines the effect size and the borders encompassing 95% of the differences define the 95% confidence interval (95CI, values between brackets). Medians are indicated with a circle and the 95CI with a horizontal line.
Median difference with confidence intervals
In a previous blog, I explained how bootstrapping can be used derive the 95CI of the median. Briefly, bootstrapping is resampling of existing data in silico, typically 1000 times. This process simulates repeating the actual experiment (footnote 3). Bootstrapping can be done with the data of the experiment, for each of the conditions, yielding 1000 median values which are displayed in figure 2 (step 1). The 1000 medians reflect a distribution of values that would have been obtained, if the experiment were to be repeated 1000x. These bootstrapped medians can be used to calculate the effect size, i.e. difference between the median of the control group and the other conditions. The result of the calculation is 1000 difference values per condition (figure 2, step 2). The resulting distribution of the differences can be used to determine the 95CI. As explained previously, the 95CI is taken from the middle 95% of the data, i.e. the values at the 2.5th and 97.5th percentile (figure 2, step 3). Thus, by bootstrapping we obtain the difference between medians and the 95CI (footnote 4).
Result and interpretation
The resulting graph shows two effect sizes with confidence intervals. All values are in the original units (µm²) and therefore these numbers make sense. The effect of TIAM is an area change of 207 µm² with a 95CI that ranges from 27 µm² to 439 µm² (footnote 5). The confidence interval can be listed between brackets: [27,439]. Since the 95CI does not overlap with zero (as also clear from the graph in figure 1 and 2), this is interpreted as a significant change (at an alpha level of 0.05). On the other hand, the effect of LARG is -206 [-350,35] µm². Since the 95CI does overlap with zero for this condition, this implies that there is no conclusive evidence for an effect. Note that the interpretation of the confidence intervals is consistent with the p-values listed in figure 1. For more information on the interpretation of effect size and 95CI see table 5.2 by Yatano (2016).
Conclusion
The effect size is the difference between conditions. It is of great value for the quantitative comparison of data and it is sample size independent. Another advantage is that the effect size is easy to understand (in contrast to p-values, which are often misunderstood or misinterpreted). The benefit of the bootstrap procedure to derive the confidence intervals is that it delivers a distribution of differences. The distribution conveys a message of variability and uncertainty, reducing binary thinking. So, make a difference by adding the effect size to your graph. The effect size facilitates interpretation and can supplement or replace p-values.
Acknowledgments: Thanks to Franka van der Linden and Eike Mahlandt for their comments.
Footnote 1: The actual definition of a p-value is: The probability of the data (or more extreme values) given that (all) the assumptions of the null hypothesis are true.
Footnote 2: Examples of relative effect size are Cohen’s d and Cliff’s delta. The Cliff’s delta is distribution independent (i.e. robust to outliers) and can be calculated using an excel macro, see: Goedhart (2016)
Footnote 3: Bootstrapping assumes that the sample accurately reflects the original population that was sampled. In other words, it assumes that the sampling was random and that the samples are independent. These assumptions also apply to other inferential statistics, i.e. significance tests.
Footnote 4: The website www.estimationstats.com (developed by Adam Claridge-Chang and Joses Ho) uses bootstrapping to calculate the effect size and 95CI for the difference of means. It is also a good source for information on effect sizes.
Footnote 5: Bootstrapping depends on random resampling, and therefore the numbers will be slightly different each time that resampling is performed.
If you’re into developmental biology, chances are you’ve spent some time in your life thinking about how cells change the shapes of tissues. What would cells need to do in order to prevent change of tissue shape, though? In the text below, I summarize my thoughts on why the question of not changing shape during growth might be of interest and discuss the findings from our PLoS Biology publication on the topic (Matejčić et al., 2018). I performed the experiments for this work in the lab of Caren Norden from the Max Planck Institute of Molecular Cell Biology and Genetics in Dresden and the theoretical work was done by Guillaume Salbreux from The Francis Crick Institute in London.
As tissues develop, they need to grow into their correct size and shape in order to function optimally. Biological studies in the past, as well as those today, investigated changes of tissue shapes during the process of morphogenesis, and how cells that build these tissues contribute to shape changes (for example, invagination or elongation). However, many tissues, such as the human ear or foot, establish their shape very early in development, and then need to continue to grow without changing their shape.
At first, such maintenance of the shape of tissues might seem like a very simple task; the cells building the tissue should only maintain an already established shape. But, as the size of a tissue changes, so might the conditions around it, making this task more complicated. A larger tissue may, among other changes, experience new physical constraints as its cells become more packed, stretched and/or compressed. As an example, imagine you have a cultured, 1000 µm x 1000 µm x 10 µm sheet of a simple, monolayered epithelial tissue. You want to scale this culture up 10 times while keeping its proportions constant (i.e., maintain its surface-to-thickness ratio). How would you go about this scaling task? You might wait for your epithelial sheet to proliferate and grow more, but it might only spread, making its apical surface bigger but keeping its thickness the same. You might then constrain its growth laterally, but cells might create a multilayered epithelium. In this case, cells might be extruded, stop dividing or the tissue might buckle. Therefore, keeping a tissue shape unchanged while it is increasing its size is not always as simple as adding more cells through proliferation. For similar reasons, it is also not trivial to predict which cellular parameters will affect tissue shape as its size changes. Despite the importance of such scaling of tissue shape during growth, this complex question had not previously been addressed.
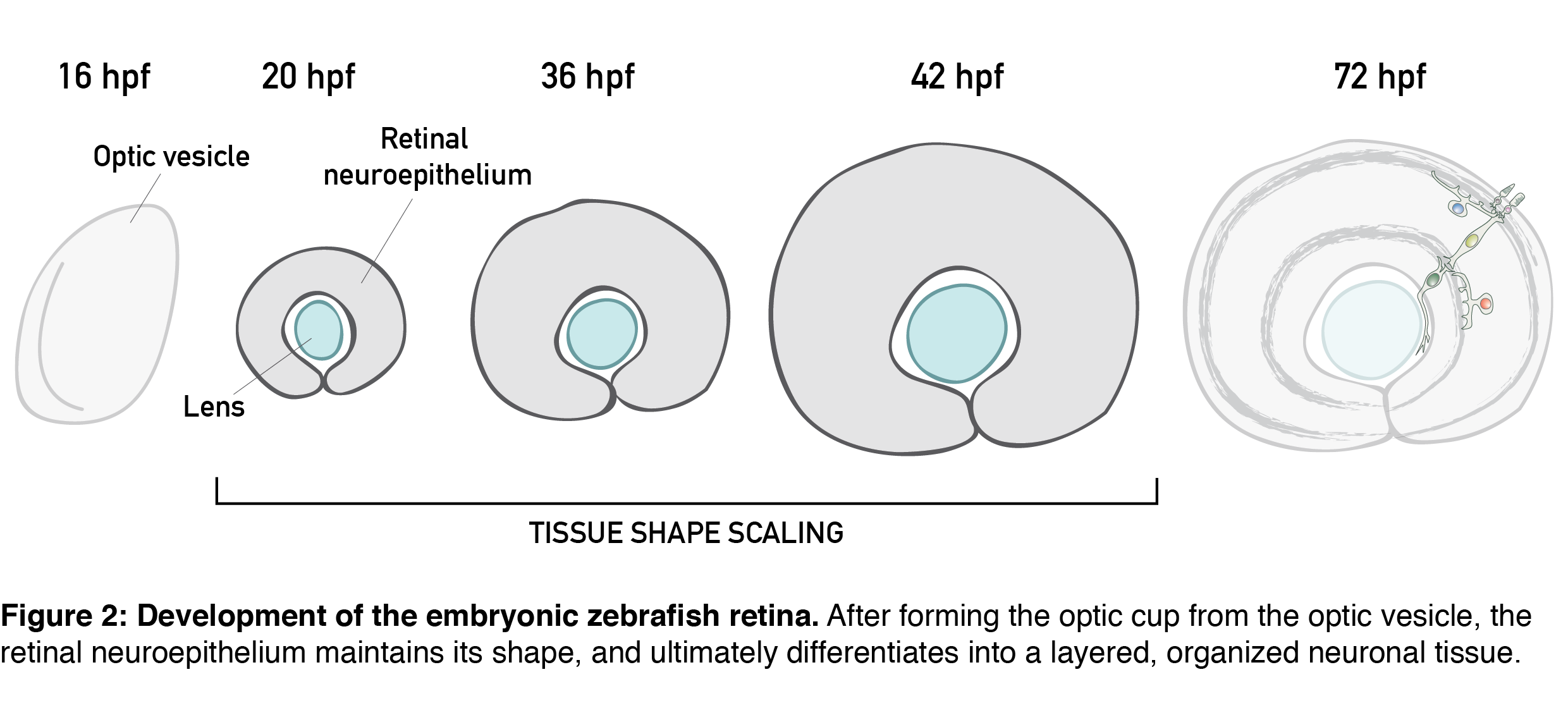
The topic of tissue shape scaling during growth is best investigated in a tissue with a geometrically simple shape, that allows quantitative analysis, and where shape perturbations are easily detectable, as well as functionally significant. Such a tissue is the developing vertebrate retina, that establishes its shape early in development (during the first trimester in humans) and then maintains shape as it grows. It has a shape of a smooth hemispherical cup (Figure 1, left panel) and lies in the back of the vertebrate eye to transmit light impulses to the brain. As within the light path of a microscope, each component within the eye, including the retina, must be correctly shaped and aligned for light to propagate effectively. It is, therefore, not surprising that even small glitches in human retinal structure, in the form of wrinkles or thickenings, lead to distorted vision (macular pucker) and that its smooth shape is essential for optimal visual function. During embryonic development, the retina is a pseudostratified epithelium, a conserved and widely utilized tissue arrangement (Figure 1, right panel). As in all other pseudostratified epithelia, in the developing retina cells are long, spindle-shaped and all divide at the apical tissue surface.
All these features are present in the retinal neuroepithelium of the zebrafish – an experimental model with the advantages of easy manipulation, tractable tissue-wide live and fixed imaging as well as fast development. In the zebrafish, the retinal neuroepithelium undergoes a phase of shape scaling in just 24 hours (stages ~20-48 hours-post-fertilization (hpf), Figure 2), which allowed me to study this entire developmental process within a single imaging session. Together, these factors make the developing zebrafish retina an excellent system to study tissue shape maintenance during growth.
Our project on retinal growth began out of a curiosity; I dug into how cell parameters such as number, size, shape and differentiation contribute to the increase in retinal size. Through these 3D, tissue-wide quantitative analyses (Video 1), we learned that, initially, all retinal cells divide, and after the developmental stage of 36 hpf, they have a 35% chance of exiting the cell cycle. Cell loss is negligible for growth in this system, and cells during development get smaller and smaller and increasingly packed. Overall, we were able to show that retinal tissue grows exponentially, by proliferative increase in cell number.
Video 1
However, I mentioned earlier that to scale tissue shape during growth is not necessarily as simple as adding more cells through proliferation. For starters, we should know where in the tissue new cells are added; we know oriented divisions and non-uniform proliferation can affect tissue shape by causing non-isotropic growth. That’s why we first needed to confirm that cell divisions do not orient along a preferred axis. We also found that their spatial distribution is homogeneous in all developmental stages, meaning that new cells are added uniformly throughout the tissue. Together with the previous finding, we here showed that proliferation itself does not disturb isotropic retinal growth. But, as I will hopefully manage to convince you, there is still more to retinal scaling than “just” adding more cells. In the second part of this text, I describe our quantitative analysis of retinal shape, and the development of the shape of cells that build it.
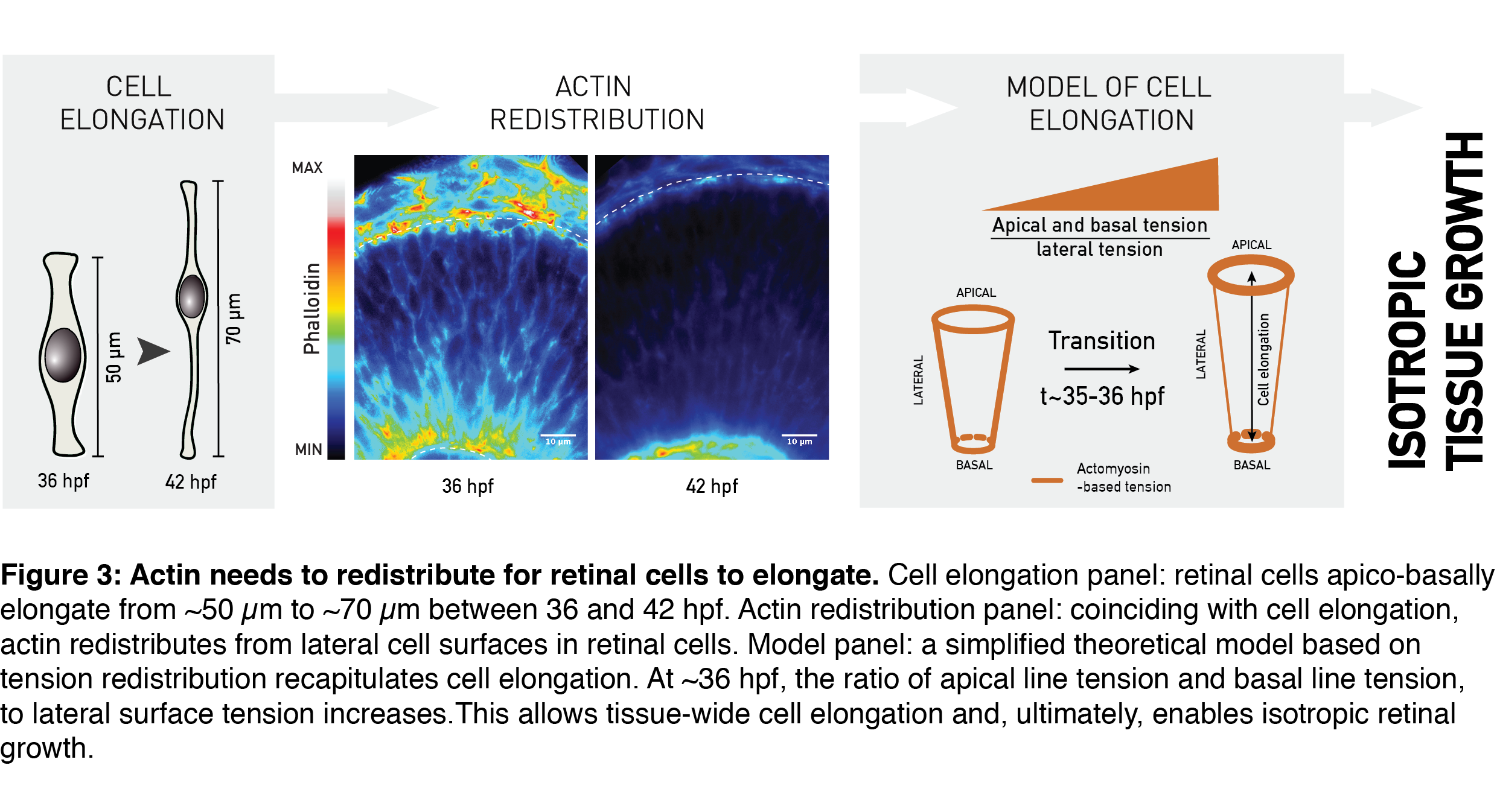
One important metric to assess proportions of a geometrical object (e.g. your computer screen) is its aspect ratio. The aspect ratio – or height-to-width ratio – can serve as a measure of shape and shape scaling of the tissue, as in my epithelial-sheet-example from the beginning. Using the aspect ratio as a metric of shape, we could quantitatively show that the shape of the whole retinal tissue does not change during growth. In other words, retinal thickness and surface area increased in concert, allowing developmental shape scaling. The thick retinal neuroepithelial tissue is still an epithelial monolayer, with each cell attached at both the basal and the apical surface of the tissue. Due to this, this apico-basal thickening of the tissue we observed translates to an elongation of retinal cells; the already slender cells thin-out and elongate even further (Figure 3, Cell elongation panel). Was it possible that this change in the shape of cells was necessary to keep tissue shape unchanged by increasing its thickness as it grew?
Cell shape in many different developmental contexts is controlled by actin (rev. in Lecuit and Lenne, 2007; Paluch and Heisenberg, 2009; Salbreux et al., 2012). To understand what governs developmental cell elongation I measured, I looked at how actin is distributed in the retina. Indeed, the subcellular distribution of actin changed as the tissue grew. Precisely coinciding with cell elongation, actin was depleted from the lateral cell regions and remained prominent only at the apical- and basal-most cell processes (Figure 3, Actin redistribution panel). A simplified theoretical model by Guillaume Salbreux could explain such actin-based cell elongation by using the actin signal redistribution as a proxy for redistribution of the cellular cortical tension (Figure 3, Model panel). In combination with the measured rate of tissue volume increase, this model also recapitulated the overall constant aspect ratio of the retinal tissue during growth.
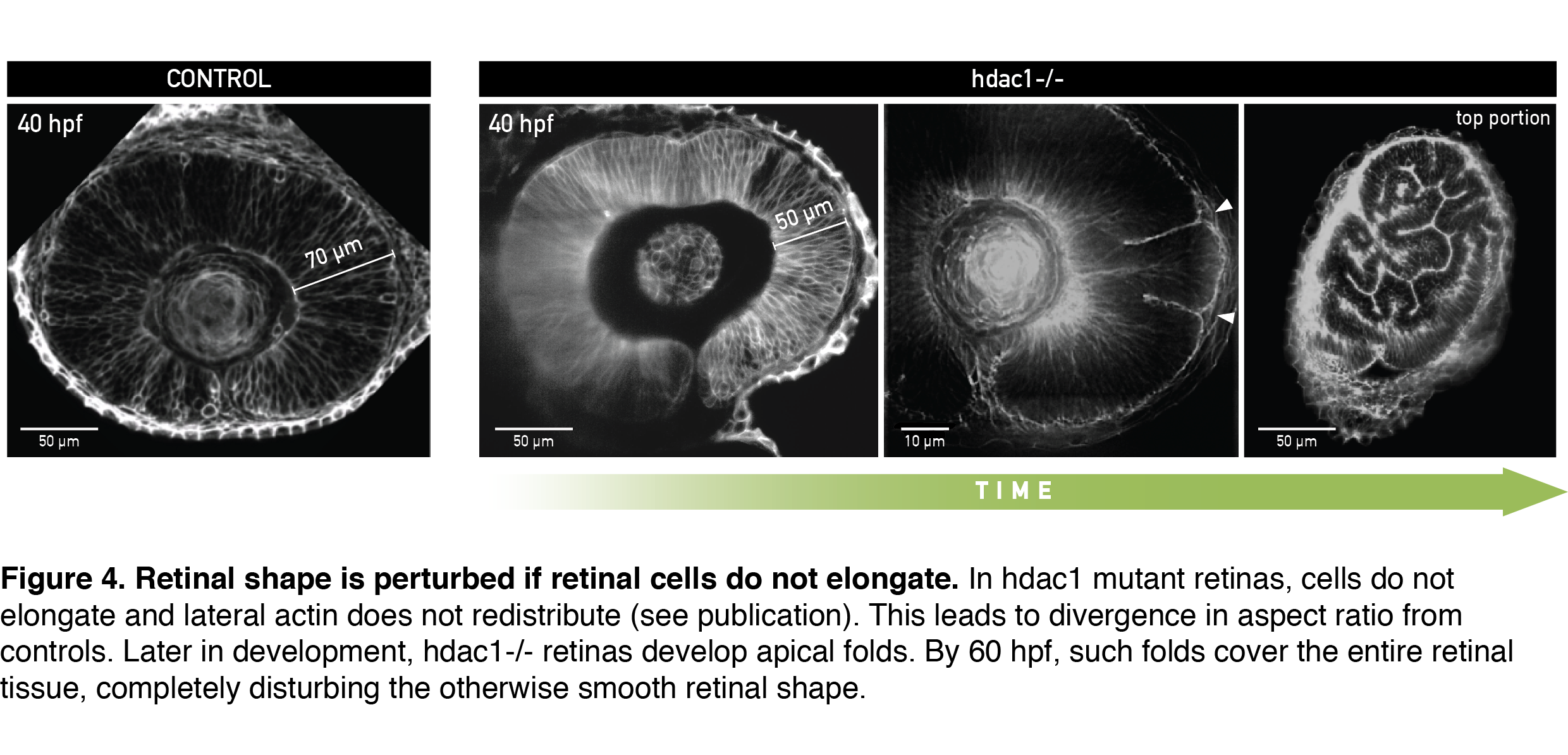
To test if cells indeed needed to elongate to allow scaling of tissue shape, I analyzed retinal development in the zebrafish mutant for histone deacetylase 1 (hdac1-/-), where eye shape was shown to be severely perturbed in the late stages of development (Stadler et al., 2005; Yamaguchi et al., 2005). From live light sheet imaging datasets, I found that cells in the hdac1-/- retina indeed failed to elongate (Figure 4, 40 hpf). In a very exciting finding, completely consistent with our working model, the lateral cell region in these mutants remained rich in actin signal even in late developmental stages. By simply keeping the cell tension ratio in our model unchanged we could theoretically recapitulate the divergence in shape between control and mutant tissue that we measured experimentally. This convinced us that redistribution of the lateral actin pool is necessary for cells to elongate and keep overall retinal shape intact. Upon growing further, the short-celled hdac1-/- retinal epithelia started disturbing their otherwise smooth tissue surface by folding apically (Figure 4, Video 2).
Video 2
A day later, retinal folds appeared throughout the hdac1-/- retinal tissue and created a structure resembling more a miniature folded brain than a retina (Figure 4, rightmost panel). This severe shape perturbation demonstrated again how important it is for the retinal cells to elongate to maintain tissue’s aspect ratio during growth.
In an interesting side observation, folded retinas of hdac1 mutants could still differentiate into a neuronal tissue, despite the tissue having a heavily disturbed shape that would subsequently interfere with light propagation. This fact decouples retinal differentiation from tissue shape and highlights the importance of precise timing of developmental events; proliferative growth, actin redistribution and differentiation all need to be temporally coordinated to give rise to a fully functioning retina.
We also found that the synchronous, tissue-wide redistribution of actin and cell elongation are non-cell-autonomous events that depend on the extracellular matrix (ECM). We think that the ECM composition (or structure) changes at ~36 hpf in an Hdac1-dependent manner, and that this change affects actin tissue-wide, coordinating its redistribution within all retinal cells. In the zebrafish retina, Hdac1 functions upstream of Wnt/ß-catenin and Notch signaling. In turn, Wnt/ß-catenin can affect expression of ECM-genes (rev. in Astudillo and Larraín, 2014). So, by inhibiting Wnt/ß-catenin at the right time during development, Hdac1 might indirectly cause a change in composition of the basement membrane, and the ECM – actin axis might then act on actin redistribution inside retinal cells. Considering the evolutionary conserved tissue architecture and the players governing shape maintenance in the retina, similar mechanisms might be at play in other growing pseudostratified epithelia, such as the human retina or brain.
…
Overall, we combined experiments and theory to investigate the maintenance of retinal tissue shape during growth. We identified actin redistribution from the lateral cell membranes as a simple developmental cue necessary to elongate cells and thus keep the overall aspect ratio of the retinal tissue unchanged. Our story highlights that uniform growth and constant shape during development are not default states but require maintenance at the cell level (Shraiman, 2005), and I hope it will motivate further studies of coordination growth and shape during development.
Altogether, we generated a rich, 3D tissue-wide dataset of retinal growth and shape, quantifying many cell-level parameters during development of the retinal neuroepithelium. It would be great to have such detailed studies of many other organs. I think such knowledge should then be used as a ground truth of wildtype development to get closer to rebuilding synthetic and simulated tissues, but also to “debug” development in systems such as organoids, to ultimately really understand in vivo development.
References
Astudillo, P. and Larraín, J. (2014). Wnt Signaling and Cell-Matrix Adhesion. Curr. Mol. Med.14, 209–220.
Fish, J. L., Dehay, C., Kennedy, H. and Huttner, W. B. (2008). Making bigger brains-the evolution of neural-progenitor-cell division. J Cell Sci121, 2783–2793.
Lecuit, T. and Lenne, P.-F. (2007). Cell surface mechanics and the control of cell shape, tissue patterns and morphogenesis. Nature Reviews Molecular Cell Biology8, 633–644.
Matejčić, M., Salbreux, G. and Norden, C. (2018). A non-cell-autonomous actin redistribution enables isotropic retinal growth. PLOS Biol16, e2006018.
Paluch, E. and Heisenberg, C.-P. (2009). Biology and Physics of Cell Shape Changes in Development. Current Biology19, R790–R799.
Salbreux, G., Charras, G. and Paluch, E. (2012). Actin cortex mechanics and cellular morphogenesis. – PubMed – NCBI. Trends in Cell Biology22, 536–545.
Shraiman, B. I. (2005). Mechanical feedback as a possible regulator of tissue growth. Proc. Natl. Acad. Sci. U.S.A.102, 3318–3323.
Stadler, J. A., Shkumatava, A., Norton, W. H. J., Rau, M. J., Geisler, R., Fischer, S. and Neumann, C. J. (2005). Histone deacetylase 1 is required for cell cycle exit and differentiation in the zebrafish retina. Dev. Dyn.233, 883–889.
Yamaguchi, M., Tonou-Fujimori, N., Komori, A., Maeda, R., Nojima, Y., Li, H., Okamoto, H. and Masai, I. (2005). Histone deacetylase 1 regulates retinal neurogenesis in zebrafish by suppressing Wnt and Notch signaling pathways. Development132, 3027–3043.
The winners of Nikon’s Small World in Motion 2018 Competition have just been announced, and overall first place has gone to a stunning developmental biology SPIM movie.
The Francis Crick Institute is recruitingEarly Career Researchers who wish to set up their first independent research programme at the Crick in any area related to biomedicine. We welcome applications from those who wish to work on a flexible and/or part-time basis.
Successful candidates will be offered a competitive salary with a 6-year contract, renewable once for a total of 12 years. The package includes:
Salaries and consumables for up to five researchers, including graduate students
Opportunity to expand through external grant funding
Ready access to Crick Core Facilities
Full lab setup in state-of-the-art laboratory space
Package applies to the duration of the contract
The Crick will provide mentoring and support to ensure its early career Group Leaders make the most of their time at the institute and develop a world-class research programme. Towards the end of the 12-year period we will support them to find leadership positions elsewhere, with potential for a transition start-up package for those remaining in the UK.
Applications from candidates with a PhD and postdoctoral experience should be submitted online at:
Location: University of Southampton, UK
Salary: £30,395 to £36,261 per annum Full Time – Fixed Term for 3 years
Closing Date: Monday 05 November 2018
Interview Date: See advert
Reference: 1061818BJ
A Research Fellow position is available in the laboratory of Mammary Stem Cell Biology & Breast Cancer headed by Dr. Salah Elias at the School of Biological Sciences (SoBS) – University of Southampton (UoS), to study the mechanisms of asymmetric cell division during mammary gland development and homeostasis. The position is available for 3 years tenable from February 2019, funded by the Medical Research Council (MRC). On appointment your post title will be Research Fellow.
The Project
Our lab focusses on studying the mechanisms that regulate mammary stem cell fate and dynamics in normal development and breast cancer. This exciting project is a collaboration between our group and Dr. Philip Greulich group based at the Department of Mathematical Sciences at UoS. It will employ combined cutting-edge in vivo single-cell lineage tracing, quantitative three-dimensional (3D) high-resolution imaging and single-cell RNA-sequencing as well as mathematical/computational modelling to (1) identify novel mechanisms that control mitotic spindle orientation in mammary stem cells; and (2) determine how these mechanisms influence cell fate outcomes in the differentiating mammary epithelium. The outcomes of this collaborative project are expected to provide important novel insight into the identity, dynamics and potential of mammary stem cells.
The Successful Candidate
We are looking for a creative, ambitious and skilled Postdoctoral Researcher Scientist willing to challenge an innovative project by adopting a pro-active attitude and an analytical approach, with a strong interest in interdisciplinary collaboration.
You will be responsible for the development of the project, which includes experimental design, data collection and interpretation. You will work in collaboration with Dr. Greulich group who will use mathematical modelling to compare the generated experimental data with predictions from stochastic models for cell fate dynamics, and test the project’s hypotheses via Bayesian inference. You are also expected to contribute to new ideas for research projects, develop ideas for writing grant proposals, prepare scientific reports, write up results for publication in international peer-reviewed journals, assist other members of our group or people working on collaborative projects to become familiar with new methodologies, act as a source of information and advice on scientific protocols.
You will hold a PhD* or equivalent professional qualifications and experience in epithelial stem cell and/or cancer biology (or related field). A strong evidence of proficiency in cell biology and quantitative advanced microscopy in vivo is necessary. You will have experience in animal models and have a personal licence to work with rodents or be prepared to obtain such a licence via attendance of in-house courses. Experience in molecular biology techniques including Next Generation Sequencing is desirable. Basic understanding of computational/mathematical modelling would be advantageous. You will be friendly and have excellent interpersonal skills with a desire to communicate with other researchers whilst maintaining the highest level of professionalism at all times.
The Environment
At SoBS, we use cutting-edge technologies for innovative research to define the basis of human health and disease, and develop interventions that benefit peoples’ lives. There is a strong interdisciplinary research focus bringing together researchers from Biological and Medical Sciences, Computer Sciences, Physics and Mathematical Sciences with experimental work housed in a £45 million building that encompasses cutting-edge research infrastructures. Our Imaging Microscopy Centre (IMC) provides access to several advanced optical microscopic modalities equipped to perform the 3D imaging of this proposal. Excellent Specific-Pathogen-Free (SPF) Animal and Histology Facilities are also available. SoBS offers a supportive and dynamic research environment, with comprehensive training and career development opportunities. You will have full access to all resources and undertake appropriate training in the use of the equipment of our state-of-the-art core facilities to accomplish your studies. SoBS has an outstanding Stem Cell and Developmental research theme, in which our group is perfectly embedded, with excellent potential for collaborations that extend to the closely located Cancer Sciences where breast cancer research is very active. You will thrive within a unique international, stimulating and challenging research environment, with an outstanding international research seminar series in addition to Stem Cells and Quantitative Biology-centred meetings co-organized by SoBS and Mathematical Sciences. The proximity of SoBS and Mathematical Sciences and the specialized expertise that each contributes provides a unique environment needed to achieve the maximum impact of this project.
For informal enquiries, please contact Dr Salah Elias S.K.Elias@soton.ac.uk and/or Dr. Philip Greulich P.S.Greulich@soton.ac.uk
It is anticipated that interviews will take place at the end of November 2018.
Equal Opportunities and Benefits
SoBS holds an Athena SWAN Silver Award, demonstrating commitment to equal opportunities and gender balance in the workplace.
The University of Southampton has a generous maternity policy and onsite childcare facilities; employees are able to participate in the childcare vouchers scheme. Other benefits include state-of-the-art on-campus sports, arts and culture
facilities, a full programme of events and a range of staff discounts.
Application Procedure
Closing Date: 5th November 2018. However, we encourage early applications as we will be reviewing applications on an on-going basis. Therefore the advert may close before the deadline if suitable candidates are identified. First review date: 29th October 2018
How to Apply: You should submit your completed application form online at www.jobs.soton.ac.uk. Please include (1) a cover letter outlining your scientific interests, describing how you meet the requirements of the position, and an outline of future goals; (2) a curriculum vitae, (3) contact information for at least two references.
References are requested along with your application, so please allow time for these to be received prior to the close date, to assist the department with shortlisting.
If you need any assistance, please call Samantha Stubbs (Recruitment Team) on +44 (0) 23 8059 4046. Please quote vacancy reference number 1061818BJ on all correspondence.
*Applications will be considered from candidates who are working towards or nearing completion of a relevant PhD qualification. The title of Research Fellow will be applied upon successful completion of the PhD. Prior to the qualification being awarded the title of Senior Research Assistant will be given.
Antibodies.com is proud to support researchers with travel grants up to £500.
The Award:
Each quarter, Antibodies.com offers a travel grant up to £500 to help cover the cost of attending a conference.
These travel grants are open to PhD candidates, lab managers, and post-docs from academic research institutions across Europe. The grant is intended to help cover the costs of registration, accommodation, and travel to a conference of choice.
For a chance to win, simply complete the application form at Antibodies.com; including a summary of your research or abstract for the conference.
Winners:
A team of scientists will read all research summaries / abstracts that are submitted and select the winner based on which research they find most interesting. The winner will be notified of the award by email.
Soon after the rediscovery of Mendel’s laws of inheritance in plants, French scientist Lucien Cuénot published a paper in 1902, reporting his studies of the inheritance of pigmentation in the house mouse.
Cuénot’s results showed that Mendel’s laws of inheritance also applied to animals. This is a fundamental paper in the field of genetics.
The original paper was published in French & many people may not read French. Therefore, Google, myself, and Phil Soriano (Mount Sinai, New York) translated the original manuscript into English.
Shuo-Ting Yen & Chang-Ru Tsai (MD Anderson Cancer Center) did the Chinese translation. Seol Hee Im (Haverford) did the Korean translation. Vanessa Barone (Scripps Institution of Oceanography) did the Italian translation.
You can find these translations at the University of Texas Genetics & Epigenetics Graduate Program website under Historical Translations (https://bit.ly/2P0gJ4v). We hope this provides the opportunity for many people to read this classic paper in genetics.
 (No Ratings Yet)
(No Ratings Yet)
 (20 votes)
(20 votes)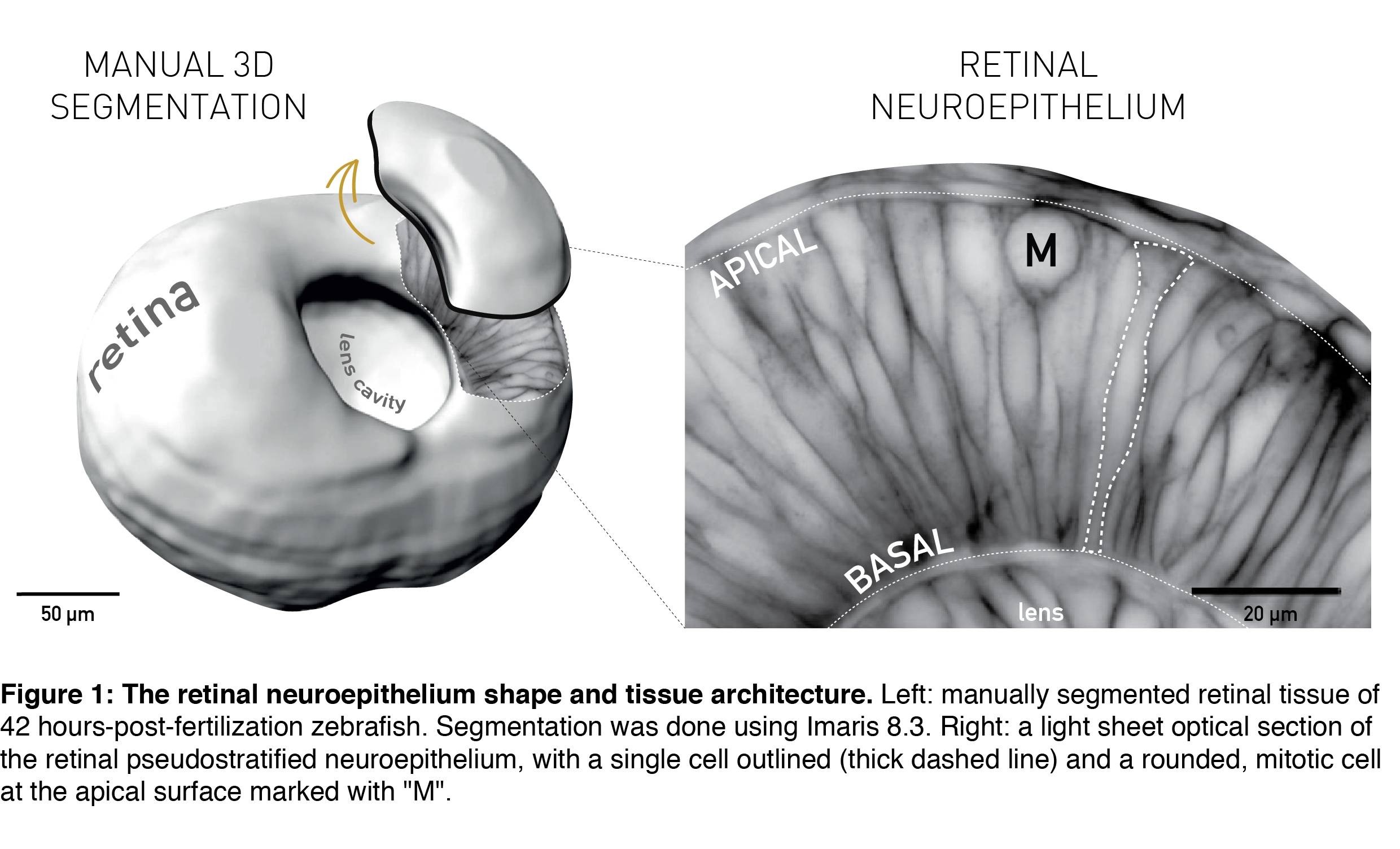
 (3 votes)
(3 votes)
